



Introduction
Breast cancer is one of the most common malignant tumors in women in the world, and it is also the malignant tumor with the highest fatality rate among women [1, 2]. According to data released by the International Cancer Society in 2018, breast cancer accounts for 11.6% of all cancers and 6.6% of all cancers. At present, the clinical treatment of breast cancer is mainly surgical treatment, but there are many molecular subtypes of breast cancer, some of which are prone to recurrence and metastasis, resulting in poor prognosis after surgical treatment [3–5]. Therefore, the search for new diagnosis and treatment has become the key to the diagnosis and treatment of breast cancer.
Circular RNA (circRNA) is a kind of endogenous RNA molecule with the closed ring structure, which exists widely and variously in eukaryotic cells and can regulate gene expression [6, 7]. In the 1990s, the existence of circRNAs was found in human transcripts, but not enough attention was paid to it [8]. With the development of high-throughput sequencing technology, a large number of circRNAs have been discovered and confirmed to have the function of post-transcriptional regulation. CircRNAs have attracted more and more attention [9]. Studies have confirmed that circRNAs mainly affects the occurrence, development, invasion and metastasis of tumor by evading growth inhibition and apoptosis, activating invasion and metastasis and angiogenesis, and maintaining proliferation signal [10]. The expression of CircRNAs is tissue-specific and related to tumor staging [11]. In addition, it has been found that circRNAs could stably exist in peripheral blood, exocrine and other body fluids, so it has the potential to become a marker for tumor diagnosis, prognosis and efficacy monitoring [12]. Yang Rui used RNA sequencing technology (RNA sequencing, RNA-seq) to analyze 4 pairs of breast tissues and paracancerous tissues, and found that the expression of annular RNA hsa_circ__0058514 in tumor tissues increased. Yang Rui et al. found that down-regulation of hsa_circ_0058514 expression could significantly reduce the proliferation, migration and invasion of breast cancer cells, and promote apoptosis. The above experiments suggest that hsa_circ_0058514 is expected to be a new target for the treatment of breast cancer [13]. Tang screened 1705 abnormally expressed circRNAs, in breast cancer by sequencing and found that the expression of hsa_circ_0001982 was increased in breast cancer by further study. Moreover, Tang uses bioinformatics methods to predict that miR-143 is the target of hsa_circ_0001982. Cytological experiments have verified that knockout of hsa_circ_0001982 in breast cancer cells inhibits breast cancer proliferation and induces apoptosis mainly through targeting miR-143 [14]. Circ_Foxo3 was first discovered as a down-regulated circRNA in breast cancer and other tumor tissues. Experimental data show that circ_ Foxo3 can promote the level of Foxo3 protein by binding to several microRNA shared by Foxo3 linear mRNA. In addition, ectopic Foxo3 pseudogenes and Foxo3 play a role in inhibiting the proliferation, survival and progression of cancer cells. These results suggest that circ_Foxo3 can up-regulate the level of Foxo3 protein through two different mechanisms. It is suggested that circ_Foxo3 could regulate the cellular function of tissue cells and inhibit the progression of cancer cells [15–17].
Here, We demonstrated that the role of the underlying mechanism of circular RNA RBM33 (circ_RBM33) in breast cancer, which would be a novel therapy target of breast cancer.
Results
CircRBM33 knockdown prevents breast cancer cells development
In previous research, circRBM33 was upregulated in different tumor tissues and cells [18, 19]. Here, we found that circRBM33 was also upregulated in different breast cancer cell lines (Figure 1A). Next, we constructed siRNA for inhibiting the expression level of circRBM33 in MCF7, MB-MDA-231, BT20, and HS578T breast cancer cell lines and immortalized breast epithelial MCF-10A cell line. The knockdown efficiency was confirmed in breast cancer cells (Figure 1B and Supplementary Figure 1A). Ki-67 staining was performed to detect the cell proliferation in breast cancer cells. We observed that si-circ RBM33 inhibited cell growth in MCF-7 and MDA-MB-231 cells (Figure 1C). Flow cytometry assay showed that si-circRBM33 blocked cell cycle from G0/G1 phase into S phase in breast cancer cells (Figure 1D). Meanwhile, si-circRBM33 prevented clone formation in MCF-7 and MDA-MB-231 cells (Figure 1E). Transwell invasion assay results revealed that silencing of circRBM33 would inhibit cell invasion in breast cancer cells (Figure 1F). TUNEL assay also performed that silencing of circRBM33 induced apoptosis in MCF-7 and MDA-MB-231 cells (Figure 1G). Meanwhile, we observed that the depletion of circRBM33 had no effect on proliferation, invasion, and apoptosis of MCF-10A cells (Supplementary Figure 1B–1E). Taken together, silencing of circRBM33 prevented tumor progression by inhibiting tumor progression in breast cancer cells.
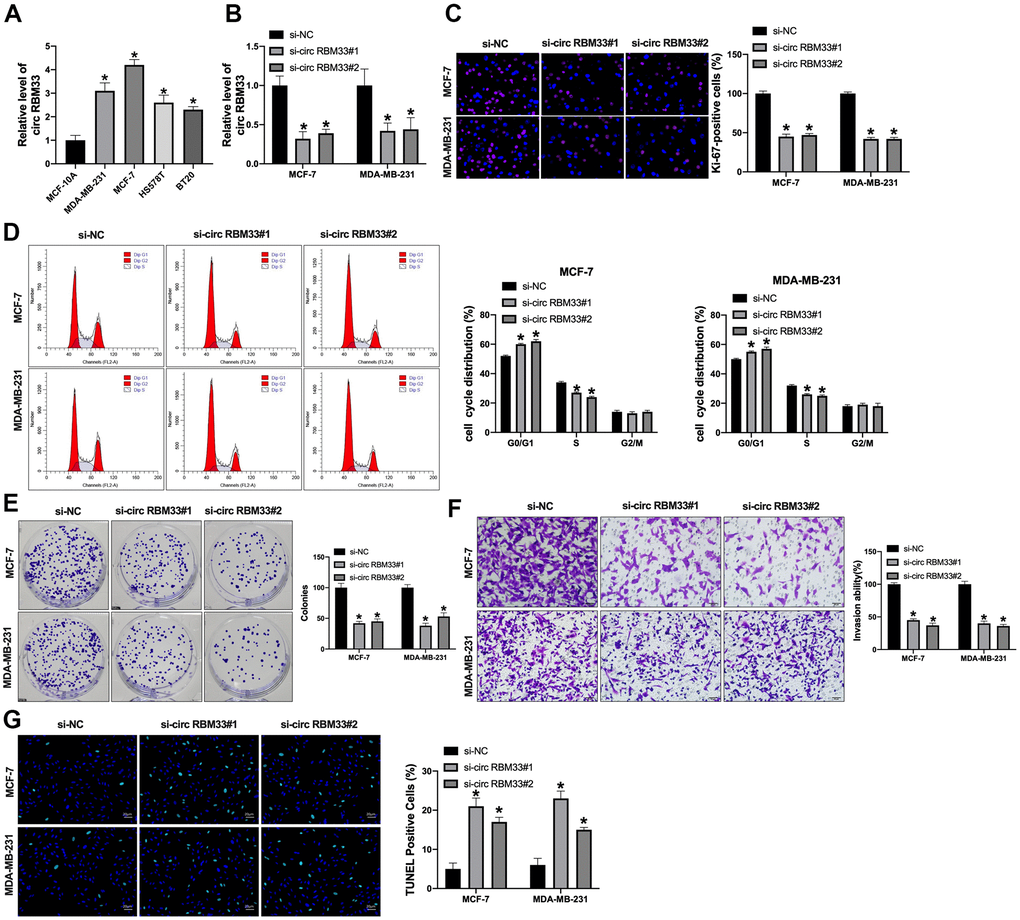
Figure 1. The expression of circRBM33 is upregulated in breast cancer cells. (A) The expression of circRBM33 was analyzed by qPCR in different breast cancer cells. (B–G) The MCF-7 and MDA-MB-231 cells were treated with circRBM33 siRNAs. (B) qRT-PCR was performed to assess the circRBM33 level. (C) Ki-67 staining was used to detect the cell proliferation (D) Cell cycle determined by flow cytometry analysis. (E) The cell proliferation was detected by clone formation assay. (F) The cell invasion was measured by Transwell assay. (G) The cell apoptosis was determined by TUNEL assay. Data are presented as mean ± SEM. Statistic significant differences were indicated: * P < 0.05, ** P < 0.01.
CircRBM33 regulates tumor progression via Wnt pathway
Wnt/β-catenin signal pathway involved in many tumor progression. We treated the Wnt agonist 1 into cells after si-circRBM33 transfection. Then we found that silencing of circRBM33 inhibited cell cycle, while Wnt agonist 1 prevented the function of si-circRBM33 (Figure 2A). Further, clone formation assay revealed that Wnt signal activation abolished the function of si-circRBM33 (Figure 2B). Transwell invasion assay also declared that the inhibition effect of si-circRBM33 was blocked by si-circRBM33 (Figure 2C). Similarly, si-circRBM33 induced apoptosis level in breast cancer cells, which was inhibited by Wnt agonist 1 (Figure 2D). Consistently, the depletion of circRBM33 enhanced Bax and cleaved caspase-3 expression and reduced Bcl-2 expression, while Wnt agonist 1 reversed this effect in the cells (Figure 2E). Taken together, circRBM33 may regulate breast cancer progression via Wnt signal pathway.
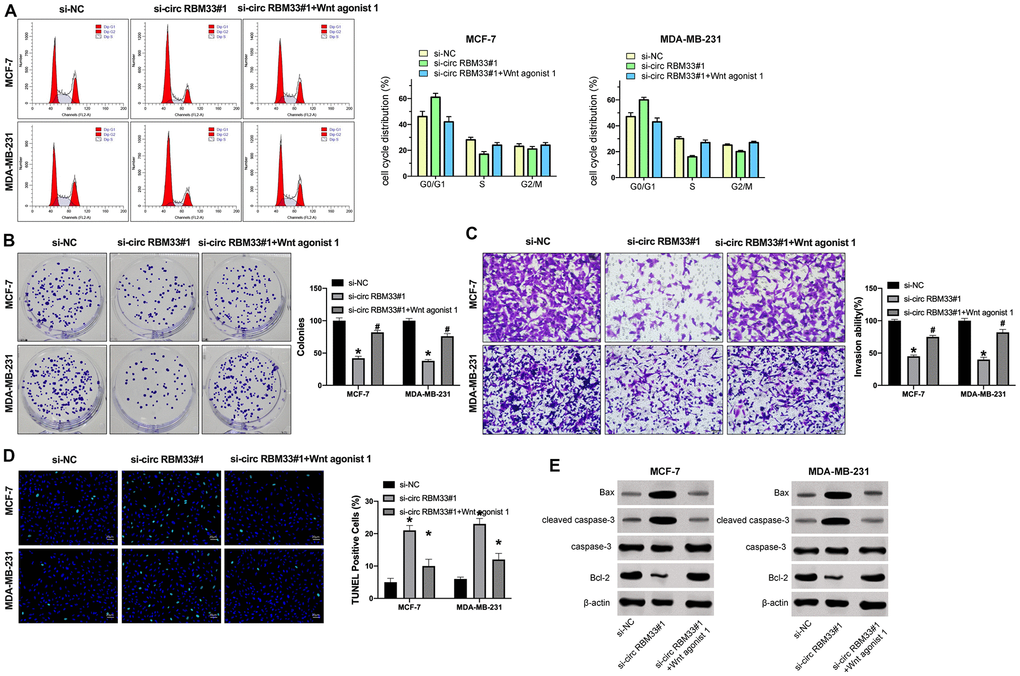
Figure 2. Wnt signal activation prevents si-circRBM33 effect on breast cancer cells. (A–D) MCF-7 and MDA-MB-231 cells were treated with circRBM33 siRNA or co-treated with circRBM33 siRNA and Wnt agonist 1 (10 μM). (A) The cell cycle was determined by flow cytometry analysis. (B) The cell proliferation was measured by clone formation assay. (C) The cell invasion was detected by Transwell assay. (D) The cell apoptosis was assessed by TUNEL assay. (E) The expression of Bax, cleaved caspase-3, caspase-3, and Bcl-2 was tested by Western blot analysis. Data are presented as mean ± SEM. Statistic significant differences were indicated: * P < 0.05 vs. si-NC, #P < 0.05 vs. si-circRBM33.
CircRBM33 could interact with miR-17-5p
Then, we confirmed that circRBM33 was located in MCF-7 and MDA-MB-231 cell cytoplasm (Figure 3A, 3B). Bioinformatics website predicted that miR-17-5p, miR-33a-5p, miR-20a-5p could interact with circRBM33 (Figure 3C). Then we detected the expression level of miR-17-5p, miR-33a-5p, miR-20a-5p in breast cancer cells after circRBM33 transfection. Then we found that miR-17-5p was downregulated in MCF-7 and MDA-MB-231 cells (Figure 3D). The predicted binding sites were shown in Figure 3E (upper). Luciferase assay demonstrated that miR-17-5p mimic co-transfected with circRBM33-wild type (circRBM33-WT) performed decreased luciferase activity, while not circRBM33-mutant (circRBM33-MUT), the results determined that miR-17-5p could interact with circRBM33 (Figure 3E). Furthermore, RIP assay and pull-down assay also verified that circRBM33 could interact with miR-17-5p (Figure 3F, 3G). We also found that miR-17-5p was downregulated in different breast cancer cells (Figure 3H).
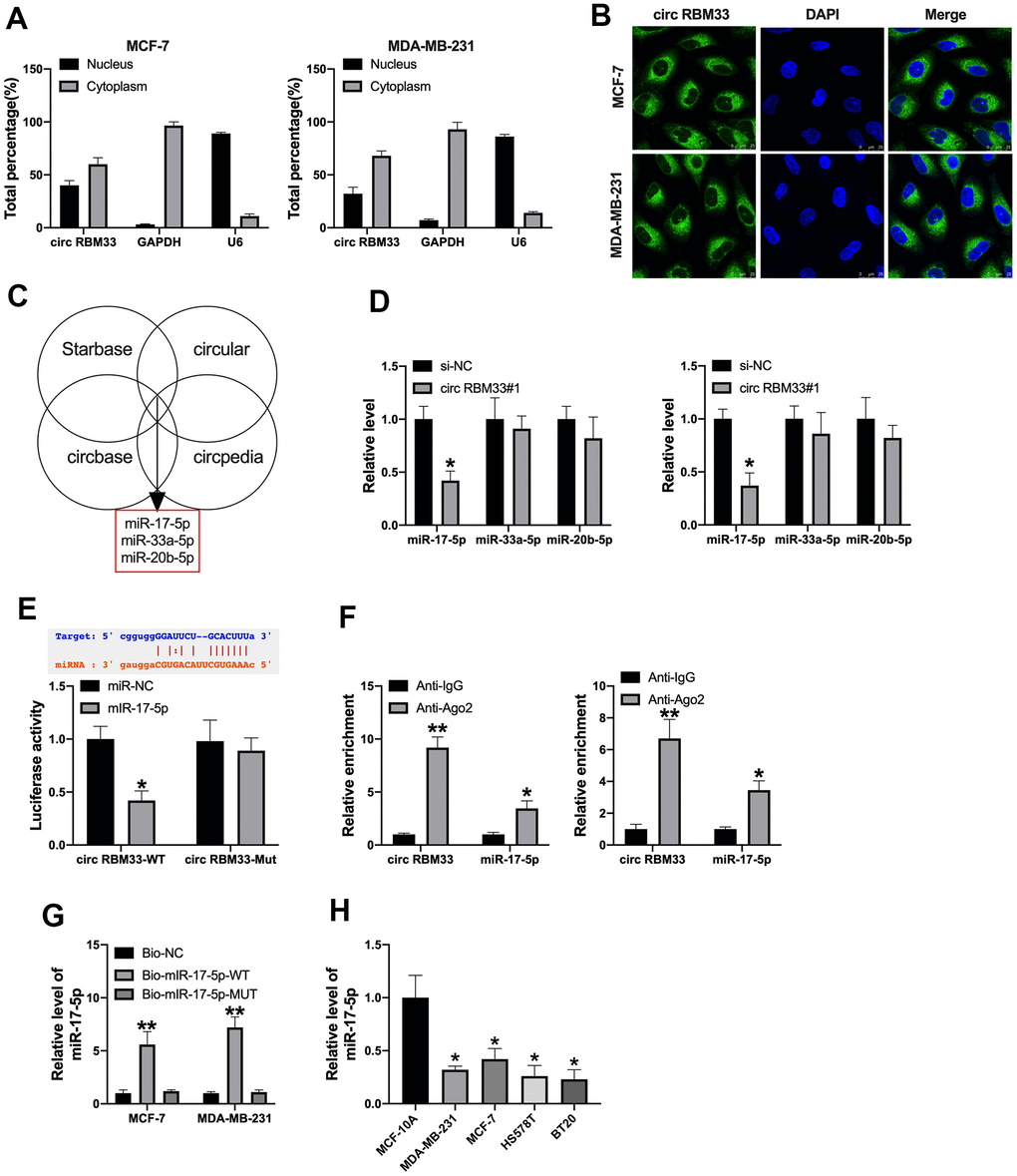
Figure 3. CircRBM33 acts as a sponge for miR-17a-5p. (A, B) Subcellular fractionation assay was performed to identify the location of circRBM33; (C) The predicted target of circRBM33; (D) The underlying target level of circRBM33, miR-17-5p, miR-33a-5p, miR-20b-5p; (E) The binding sites between circRBM33and miR-17a-5p; Luciferase activity of miR-17a-5p mimic with PITPNA-AS1-WT or PITPNA-AS1-MUT; (F) AGO2 RIP assay; (G) Pull-down assay. (H) MiR-17a-5p expression in breast cancer cell. Data are presented as mean ± SEM. Statistic significant differences were indicated: ns, no significance, * P < 0.05, ** P < 0.01.
CBX7 is an underlying target of miR-17a-5p
Bioinformatics website predicted that CBX7 was the target of miR-17-5p (Figure 4A). Database also performed that there was a negative relationship between CBX7 and miR-17a-5p (Figure 4B). The predicted binding sites were shown in Figure 3E (upper). Luciferase assay demonstrated that miR-17-5p mimic co-transfected with CBX7-wild type (CBX7-WT) performed decreased luciferase activity, while not CBX7-mutant (CBX7-MUT), the results determined that miR-17-5p could interact with CBX7 (Figure 4C). We also found that CBX7 was upregulated in different breast cancer cells (Figure 4D). AGO2 RIP and pull-down assay also revealed the similar results (Figure 4E, 4F). Overexpression of miR-17a-5p and silencing of circRBM33 prevented CBX7 expression in MCF-7 and MDA-MB-231 cells (Figure 4G, 4H). Taken together, CBX7 could be downstream of circRBM33.
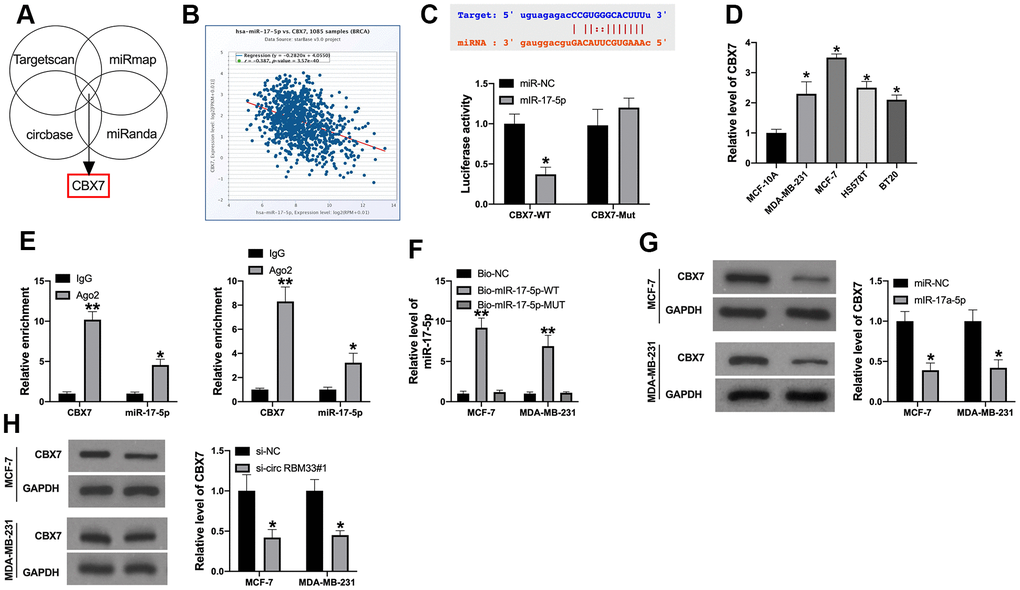
Figure 4. MiR-17a-5p regulates CBX7 expression in breast cancer cells. (A) Possible mRNA regulated by miR-17a-5p were selected and are presented in a Venn diagram; (B) The negative relationship between miR-17a-5p and CBX7. (C) The binding sites between CBX7 and miR-17a-5p; Luciferase activity of miR-17a-5p mimic with CBX7-WT or CBX7-MUT; (D) CBX7 expression in breast cancer cell. (E) AGO2 RIP assay; (F) Pull-down assay. (G, H) CBX7 expression regulation by miR-17a-5p mimic or circRBM33 inhibitor in breast cancer cells; Data are presented as mean ± SEM. Statistic significant differences were indicated: * P < 0.05, ** P < 0.01.
Functionally, we found that miR-17a-5p repressed proliferation, invasion, and induced apoptosis of MCF-7 and MDA-MB-231 cells, while the overexpression of CBX7 could reverse this effect (Supplementary Figure 2), suggesting that miR-17a-5p suppresses breast cancer progression by targeting CBX7.
CBX7 prevents the function of si-circRBM33 on tumor progression
Further, we co-transfected si-circRBM33 and CBX7 into breast cancer cells (Figure 5A). Silencing of circRBM33 prevented cell proliferation which was abolished by CBX7 overexpression (Figure 5B). Meanwhile, cell cycle and clone formation assay shown that si-circRBM33 inhibited cell proliferation, and CBX7 prevented the function of circRBM33 on breast cancer cells (Figure 5C, 5D). circRBM33 inhibition also prevented invasion ability, while CBX7 blocked circRBM33 function in breast cancer cells (Figure 5D).
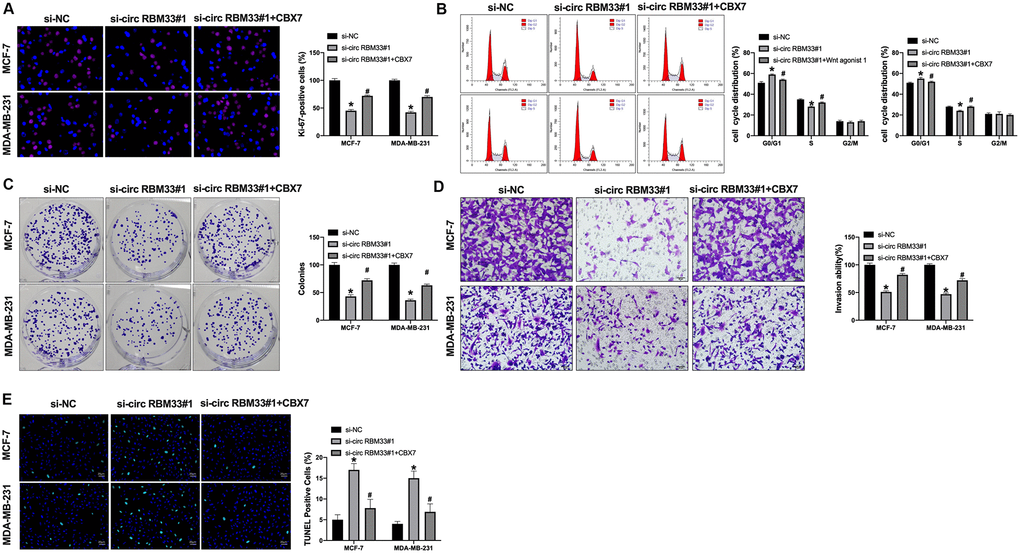
Figure 5. CIRCRBM33 regulates tumor growth via targeting CBX7. (A) Ki-67 staining was used to detect the cell proliferation; (B) Cell cycle determined by flow cytometry analysis. (C) The cell proliferation was detected by clone formation assay. (D) The cell invasion was measured by Transwell assay. (E) The cell apoptosis was determined by TUNEL assay. Data are presented as mean ± SEM. Statistic significant differences were indicated: ** P < 0.01 vs. si-NC; #P <0.05 vs.si-circRBM33.
TUNEL assay results also performed that CBX7 prevented si-circRBM33 on apoptosis level in breast cancer cells (Figure 5E). Taken together, CBX7 would be a target of circRBM33 and involve in regulating breast cancer progression.
CircRBM33/cbx7 regulates tumor growth in vivo
We constructed stabled circRBM33 low expression breast cancer cells and normal breast cancer cells, and injected into nude mice to observe the tumor growth. Silencing of circRBM33 prevented tumor volume and weight, which was inhibited by CBX7 overexpression (Figure 6A, 6B). The decreased expression of circRBM33 and CBX7, increased expression of miR-17a-5p were observed in tumor tissues, which was prevented by CBX7 (Figure 6C).
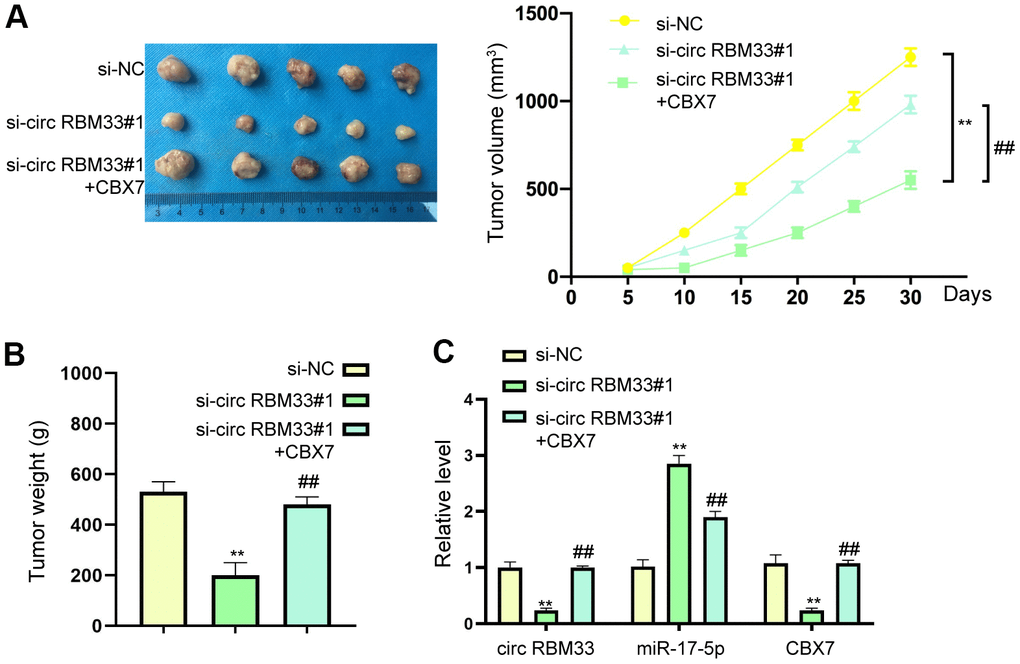
Figure 6. CIRCRBM33 regulates the tumor growth in vivo. (A–C) The SCID nude mice were randomly divided into two groups, then MCF7 cells (1×106/100μL/mouse) transfected with si-control (si-NC) si-circRBM33, or co-transfected with si-circRBM33 and CBX7 overexpressing plasmid were hypodermically injected in the nude mice. The width and length of tumor, and the body weight of mice were measured at indicated time. Tumor volume was calculated by the formula: width (mm)2 × length (mm)/2. The mice were anesthetized to death when tumor size reached 1000 mm3, and the tumors were collected. (A) The tumor volume was shown. (B) The tumor weight was shown. (C) The expression of PITPNA-AS1, miR-17a-5p, CBX7 was determined. N=5. Data are presented as mean ± SEM. Statistic significant differences were indicated: ** P < 0.01, ## P < 0.01.
Discussion
The incidence of breast cancer is increasing, which brings heavy burden to individuals and society [20, 21]. Early diagnosis of breast cancer is the key to improve the curative effect and prognosis of breast cancer. However, there are no obvious symptoms in the early stage of breast cancer, and the clinical diagnosis of breast cancer is lack of specific methods, which brings great difficulties to the early diagnosis of breast cancer. Because circRNAs are insensitive to RNA enzyme R and more stable than linear RNAs, they have unique advantages as markers [22]. Current studies have investigated the feasibility of circ RNAs as diagnostic markers for breast cancer. Nair et al. analyzed 885 samples from breast cancer patients provided by the Cancer Genome Genome (TCGA) database and found that more circRNAs were identified in the paracancerous tissues of estrogen receptor positive patients than in tumor tissues. These circRNAs can be used to distinguish different breast cancer subtypes, and the number of circRNAs was negatively correlated with cell proliferation. Yin et al. found that the accuracy of plasma hsa_circ_0001785 in breast cancer was higher than that of CEA and CA15-3. In addition, the expression level of plasma hsa_circ_0001785 was correlated with histological grade, TNM stage and distant metastasis, and the level of plasma hsa_circ_0001785 in postoperative patients was lower than that in preoperative patients [23, 24]. The expression level of circRNAs in breast cancer tissues is promising for prognostic determination and subtype classification of breast cancer in the future.
Here, we determined that circRBM33 was upregulated in breast cancer cells. Silencing of circRBM33 inhibited tumor progression and induced apoptosis in breast cancer cells. Activation Wnt signal pathway would prevent the inhibition effect of si-circRBM33 on tumor progression. However, the mechanism of circRBM33 on the inhibition of Wnt pathway should be explored by more investigations in the future. Further, we identified that miR-17a-5p was target of circRBM33. The potential relationship of other circRNAs with circRBM33 or miR-17a-5p needs to be investigation in future studies Chromobox (CBX) family is a class of PCG family proteins. CBX family includes 5 subtypes, namely CBX2, CBX4, CBX6, CBX7, and CBX8, which have different expression patterns and biological functions in different tissues [25]. Immunohistochemical analysis of sporadic colorectal cancer lesions in tissue microarray showed that CBX7 expression was reduced or absent in a large number of colorectal cancer tissues compared with non-pathological colorectal mucosa, and the reduced or absent CBX7 expression was associated with poor prognosis [26]. CBX7 can inhibit p16 gene and protein kinase B (protein kinase B, PKB/Akt)/nuclear factor kappa B predominate (nuclear factor kappa B, the nf-kappa B)/miR - 21 ways to enhance stem cell characteristics of gastric cancer cells and block CBX7 expression of gastric cancer cells, p16 gene expression, weakened stem cell properties, in addition also found Akt/nf-kappa B/miR- 21 signal axis is CBX7 startup gastric cancer stem cell characteristics of key signaling pathways downstream effect [27]. Abnormal expression of CBX7 is closely related to the occurrence, development, metastasis and aggressiveness of malignant tumors, and has important clinical significance in guiding the prognosis of malignant tumors. Here, we found that CBX7 was upregulated in breast cancer cells, which would be as an underlying target of circRBM33.
Existing studies have shown that circRNAs are involved in the whole process of the occurrence, development, invasion, metastasis and drug resistance of breast cancer. CircRNAs are expected to be novel biomarkers for diagnosis of breast cancer and new targets for treatment of breast cancer. However, it should be noted that although a large number of studies have reported the effect of circRNAs on the whole process of breast cancer, some studies have the following shortcomings. Therefore, the study of circ RNAs is still in the preliminary stage, and its effect on breast cancer needs to be further verified.
Conclusions
In summary, our results demonstrated that circRBM33 facilitates the progression of breast cancer through binding with miR-17a-5p and modulating CBX7levels. CircRBM33 may act as a tumor biomarker and may be used as a potential therapeutic target in the future.
Materials and Methods
Cell culture
Human breast cancer cell lines MCF7, MB-MDA-231, BT20, HS578T and immortalized breast epithelial cell MCF-10A were purchased from Shanghai Cell Bank of Chinese Academy of Sciences. MB-MDA-231 and MCF-10A were cultured in DMEM/F12 medium containing 10% fetal bovine serum. MCF7, BT20, MCF-10A were cultured in RPMI-1640 medium containing 10% fetal bovine serum and cultured in 5% CO2 incubator at 37° C. Wnt agonist 1 was purchased from Selleck Chemicals (Shanghai, China) and used at a final concentration of 10 μM at 37° C.
CCK-8
The cells were inoculated into 96-well plate. CCK-8 method was used to detect the cells for 3 days, and the cell growth curve was drawn. During the detection, 10 μL of 5mg/ml ‘s CCK-8 reagent was added to each well, incubated in the incubator for 4 hours, and 150 μL dimethyl sulfoxide was added to each well. The absorbance (A) value of each well at the wavelength 450 nm was detected by SpectraMax plus 384 full-wavelength enzyme labeling instrument.
Transwell invasion assay
Transwell was prepared with Matrigel matrix glue; cells were resuscitated with serum culture medium and inoculated in Transwell. Mitomycin-c was used to stop proliferation. RPMI-1640 medium containing 10%FBS was added to the lower chamber; after 24 hours of culture, the upper chamber was taken out and fixed with paraformaldehyde for 10 min 0.1% crystal violet staining solution for rinse with 10 min PBS solution; after drying, the number of transmembrane cells was counted and counted by inverted microscope.
Western blot
The cells were lysed and the total protein was extracted, then sodium dodecyl sulfate (SDS)-polyacrylamide gel (PAGE) electrophoresis was used, polyvinylidene fluoride (PVDF) membrane was transferred to the membrane, and 5% skim milk was sealed for 1 hour. The first antibody was added and incubated overnight at 4° C, and the second antibody was incubated for 2 h at room temperature. ECL developer was developed in gel imaging system.
Clone formation experiment
The cells were suspended and inoculated in a petri dish containing 10 mL 37° C preheated culture medium, placed at 37° C containing 5% CO2, and cultured in an incubator for 14 days. It was washed twice with PBS, fixed with 4% paraformaldehyde for 20 min, and treated with 0.1% crystal violet for 30 min at room temperature. The number of cell clones was observed under microscope.
In vivo tumor xenograft
ALL animal experiments in this work were conducted under the approval of Ethics Committee of the Xinjiang Medical University. SCID nude mice (5-weeks old) were ordered from Vital River Laboratory (China) and fed in a SPF environment. The mice were randomly divided into two groups, then MCF7 cells (1×106/100μL/mouse) transfected with si-control (si-NC) si-circRBM33, or co-transfected with si-circRBM33 and CBX7 overexpressing plasmid were hypodermically injected in the nude mice. The width and length of tumor, and the body weight of mice were measured at indicated time. Tumor volume was calculated by the formula: width (mm)2 × length (mm)/2. The mice were anesthetized to death when tumor size reached 1000 mm3, and the tumors were collected.
Statistical methods
Prism 8.0 statistical software was used to analyze the data, the measurement data were expressed by mean ± SEM, the data of multiple groups were compared by one-way ANOVA, and the data of the two groups were compared by t-test.
Data availability statement
The data that support the findings of this study are available from the corresponding author, upon reasonable request.
Supplementary Materials
Author Contributions
Minxia Li and Shaobo Liu designed the study; Xihui Zhang and Zhusheng Sun performed experiments; Shaobo Liu collected and analysed data; Zhusheng Sun wrote the manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- 1. Duffy SW, Vulkan D, Cuckle H, Parmar D, Sheikh S, Smith RA, Evans A, Blyuss O, Johns L, Ellis IO, Myles J, Sasieni PD, Moss SM. Effect of mammographic screening from age 40 years on breast cancer mortality (UK Age trial): final results of a randomised, controlled trial. Lancet Oncol. 2020; 21:1165–72. https://doi.org/10.1016/S1470-2045(20)30398-3 [PubMed]
- 2. Tamirisa N, Lin H, Shen Y, Shaitelman SF, Sri Karuturi M, Giordano SH, Babiera G, Bedrosian I. Association of Chemotherapy With Survival in Elderly Patients With Multiple Comorbidities and Estrogen Receptor-Positive, Node-Positive Breast Cancer. JAMA Oncol. 2020; 6:1548–54. https://doi.org/10.1001/jamaoncol.2020.2388 [PubMed]
- 3. Britt KL, Cuzick J, Phillips KA. Key steps for effective breast cancer prevention. Nat Rev Cancer. 2020; 20:417–36. https://doi.org/10.1038/s41568-020-0266-x [PubMed]
- 4. Steenbruggen TG, Steggink LC, Seynaeve CM, van der Hoeven JJ, Hooning MJ, Jager A, Konings IR, Kroep JR, Smit WM, Tjan-Heijnen VC, van der Wall E, Bins AD, Linn SC, et al. High-Dose Chemotherapy With Hematopoietic Stem Cell Transplant in Patients With High-Risk Breast Cancer and 4 or More Involved Axillary Lymph Nodes: 20-Year Follow-up of a Phase 3 Randomized Clinical Trial. JAMA Oncol. 2020; 6:528–34. https://doi.org/10.1001/jamaoncol.2019.6276 [PubMed]
- 5. Pashayan N, Antoniou AC, Ivanus U, Esserman LJ, Easton DF, French D, Sroczynski G, Hall P, Cuzick J, Evans DG, Simard J, Garcia-Closas M, Schmutzler R, et al. Personalized early detection and prevention of breast cancer: ENVISION consensus statement. Nat Rev Clin Oncol. 2020; 17:687–705. https://doi.org/10.1038/s41571-020-0388-9 [PubMed]
- 6. Vo JN, Cieslik M, Zhang Y, Shukla S, Xiao L, Zhang Y, Wu YM, Dhanasekaran SM, Engelke CG, Cao X, Robinson DR, Nesvizhskii AI, Chinnaiyan AM. The Landscape of Circular RNA in Cancer. Cell. 2019; 176:869–81.e13. https://doi.org/10.1016/j.cell.2018.12.021 [PubMed]
- 7. Flemming A. The enigma of circular RNA. Nat Rev Immunol. 2019; 19:351. https://doi.org/10.1038/s41577-019-0173-0 [PubMed]
- 8. Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J, Chen D, Gu J, He X, Huang S. Circular RNA is enriched and stable in exosomes: a promising biomarker for cancer diagnosis. Cell Res. 2015; 25:981–4. https://doi.org/10.1038/cr.2015.82 [PubMed]
- 9. Chen LL. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat Rev Mol Cell Biol. 2020; 21:475–90. https://doi.org/10.1038/s41580-020-0243-y [PubMed]
- 10. Guarnerio J, Zhang Y, Cheloni G, Panella R, Mae Katon J, Simpson M, Matsumoto A, Papa A, Loretelli C, Petri A, Kauppinen S, Garbutt C, Nielsen GP, et al. Intragenic antagonistic roles of protein and circRNA in tumorigenesis. Cell Res. 2019; 29:628–40. https://doi.org/10.1038/s41422-019-0192-1 [PubMed]
- 11. Kristensen LS, Andersen MS, Stagsted LV, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019; 20:675–91. https://doi.org/10.1038/s41576-019-0158-7 [PubMed]
- 12. Szabo L, Salzman J. Detecting circular RNAs: bioinformatic and experimental challenges. Nat Rev Genet. 2016; 17:679–92. https://doi.org/10.1038/nrg.2016.114 [PubMed]
- 13. Wu J, Liu S, Fan Z, Zhang L, Tian Y, Yang R. A novel and selective inhibitor of PKC ζ potently inhibits human breast cancer metastasis in vitro and in mice. Tumour Biol. 2016; 37:8391–401. https://doi.org/10.1007/s13277-015-4744-9 [PubMed]
- 14. Tang YY, Zhao P, Zou TN, Duan JJ, Zhi R, Yang SY, Yang DC, Wang XL. Circular RNA hsa_circ_0001982 Promotes Breast Cancer Cell Carcinogenesis Through Decreasing miR-143. DNA Cell Biol. 2017; 36:901–8. https://doi.org/10.1089/dna.2017.3862 [PubMed]
- 15. Du WW, Fang L, Yang W, Wu N, Awan FM, Yang Z, Yang BB. Induction of tumor apoptosis through a circular RNA enhancing Foxo3 activity. Cell Death Differ. 2017; 24:357–70. https://doi.org/10.1038/cdd.2016.133 [PubMed]
- 16. Lu WY. Roles of the circular RNA circ-Foxo3 in breast cancer progression. Cell Cycle. 2017; 16:589–90. https://doi.org/10.1080/15384101.2017.1278935 [PubMed]
- 17. Wang C, Tao W, Ni S, Chen Q. Circular RNA circ-Foxo3 induced cell apoptosis in urothelial carcinoma via interaction with miR-191-5p. Onco Targets Ther. 2019; 12:8085–94. https://doi.org/10.2147/OTT.S215823 [PubMed]
- 18. Ding Y, Yuan X, Gu W. Circular RNA RBM33 contributes to cervical cancer progression via modulation of the miR-758-3p/PUM2 axis. J Mol Histol. 2021; 52:173–85. https://doi.org/10.1007/s10735-020-09933-1 [PubMed]
- 19. Wang N, Lu K, Qu H, Wang H, Chen Y, Shan T, Ge X, Wei Y, Zhou P, Xia J. CircRBM33 regulates IL-6 to promote gastric cancer progression through targeting miR-149. Biomed Pharmacother. 2020; 125:109876. https://doi.org/10.1016/j.biopha.2020.109876 [PubMed]
- 20. DeSantis CE, Ma J, Gaudet MM, Newman LA, Miller KD, Goding Sauer A, Jemal A, Siegel RL. Breast cancer statistics, 2019. CA Cancer J Clin. 2019; 69:438–51. https://doi.org/10.3322/caac.21583 [PubMed]
- 21. Huo D, Hu H, Rhie SK, Gamazon ER, Cherniack AD, Liu J, Yoshimatsu TF, Pitt JJ, Hoadley KA, Troester M, Ru Y, Lichtenberg T, Sturtz LA, et al. Comparison of Breast Cancer Molecular Features and Survival by African and European Ancestry in The Cancer Genome Atlas. JAMA Oncol. 2017; 3:1654–62. https://doi.org/10.1001/jamaoncol.2017.0595 [PubMed]
- 22. Circular RNA. Nat Biotechnol. 2021; 39:23. https://doi.org/10.1038/s41587-020-00787-2 [PubMed]
- 23. Yin WB, Yan MG, Fang X, Guo JJ, Xiong W, Zhang RP. Circulating circular RNA hsa_circ_0001785 acts as a diagnostic biomarker for breast cancer detection. Clin Chim Acta. 2018; 487:363–8. https://doi.org/10.1016/j.cca.2017.10.011 [PubMed]
- 24. Li Z, Zheng J, Lin W, Weng J, Hong W, Zou J, Zhang T, Ye C, Chen Y. Circular RNA hsa_circ_0001785 inhibits the proliferation, migration and invasion of breast cancer cells in vitro and in vivo by sponging miR-942 to upregulate SOCS3. Cell Cycle. 2020; 19:2811–25. https://doi.org/10.1080/15384101.2020.1824717 [PubMed]
- 25. Gao Z, Zhang J, Bonasio R, Strino F, Sawai A, Parisi F, Kluger Y, Reinberg D. PCGF homologs, CBX proteins, and RYBP define functionally distinct PRC1 family complexes. Mol Cell. 2012; 45:344–56. https://doi.org/10.1016/j.molcel.2012.01.002 [PubMed]
- 26. Pallante P, Terracciano L, Carafa V, Schneider S, Zlobec I, Lugli A, Bianco M, Ferraro A, Sacchetti S, Troncone G, Fusco A, Tornillo L. The loss of the CBX7 gene expression represents an adverse prognostic marker for survival of colon carcinoma patients. Eur J Cancer. 2010; 46:2304–13. https://doi.org/10.1016/j.ejca.2010.05.011 [PubMed]
- 27. Ni SJ, Zhao LQ, Wang XF, Wu ZH, Hua RX, Wan CH, Zhang JY, Zhang XW, Huang MZ, Gan L, Sun HL, Dimri GP, Guo WJ. CBX7 regulates stem cell-like properties of gastric cancer cells via p16 and AKT-NF-κB-miR-21 pathways. J Hematol Oncol. 2018; 11:17. https://doi.org/10.1186/s13045-018-0562-z [PubMed]