



Introduction
Osteosarcoma (OS), a primary malignant bone tumor originating from mesenchymal tissue, ranks as the third most prevalent malignancy in children and adolescents, following leukemia and lymphoma. Characterized by a bimodal age distribution peaking in the second decade and post-65 years, OS is notorious for its high metastatic and recurrent nature [1]. Recent advancements in chemotherapy and surgical interventions have improved the five-year survival rate for primary OS patients to 65-70%. However, persistent challenges like tumor metastasis and drug resistance impede patient recovery [2]. Among these challenges, multidrug resistance (MDR), contributing to 40-50% of the adverse prognoses in OS cases, necessitates urgent attention to enhance treatment outcomes [3].
Currently, strategies to overcome MDR in osteosarcoma mainly include two approaches: targeting resistance molecules and improving drug delivery methods [4]. Targeting resistance molecules primarily involves inhibiting cell resistance by targeting membrane transporters or related signaling pathways, while improving drug delivery methods mainly involves encapsulating drugs in nanoparticles, which can be designed to co-deliver chemotherapeutic drugs and membrane transporter-targeting agents. Combining these two approaches, it is clear that discovering new drugs that target membrane transporters is a highly valuable topic in overcoming MDR in osteosarcoma.
The ATP-binding cassette (ABC) transporter superfamily, encompassing a broad spectrum of membrane transporters, is a crucial mediator of MDR, hindering drug accumulation effectivity in cancer cells [5]. Among these transporters, ABCB1 (also known as P-glycoprotein, P-gp), ABCC1, and ABCG2 are particularly notable for their enhanced expression across various MDR cancer types, transcending organ specificity [6]. In OS, ABCB1 is a recognized indicator of reduced chemo-/immune-sensitivity, heightened lung metastasis risk, and poorer clinical outcomes [7–9]. Survey on ABCB1 in OS emerges its specific involvement in resistance against doxorubicin and the essential role in cross-resistance to paclitaxel, docetaxel, vincristine, taxotere, etoposide, and vinorelbine [10, 11]. ABCB1 has been considered a significant target for overcoming MDR. Thereby, numerous strategies have been proposed to reverse drug resistance by targeting ABCB1. However, due to the limited drug absorption and extensive side effects, the discovery of novel targeting drugs remains crucial [12, 13].
Conventional ABCB1 inhibitors have been associated with increased doxorubicin accumulation in the heart, leading to cardiotoxicity [14]. However, phytochemicals and their derivatives are emerging as novel, promising ABCB1 inhibitors, offering the advantage of reduced side effects on normal tissues compared to conventional inhibitors [15]. They have been reported to enhance intracellular drug retention without exacerbating toxicity [16]. Among these phytochemicals, ginsenosides, the primary active ingredients of Panax ginseng, possess a wide range of pharmacological effects [17–19]. The ABCB1 inhibitory properties have been demonstrated in several ginsenosides, such as ginsenosides Rg1 and CK, which control temozolomide resistance in glioblastoma cells by regulating cholesterol metabolism [20]. Ginsenoside Rh2 inhibits P-gp levels by modulating the NF-kB pathway, reversing drug resistance, and showing promising medicine values when combined with thermogel [21, 22]. However, numerous ginsenosides have yet to be investigated for their potential roles in modulating tumor drug resistance. Among them, ginsenoside Rh4, a rare and novel type of ginsenoside, has exhibited anti-aging, anti-inflammatory, and anti-tumor activities [23]. However, its role in alleviating tumor drug resistance in conjunction with chemotherapy remains under-explored.
This study explores the potential MDR-modulatory effect [18, 24] of ginsenoside Rh4 in reversing drug resistance, surpassing other ginsenosides like Rh2, known for their MDR-modulatory effects [18, 24]. Ginsenoside Rh4 diminishes ABCB1 expression by inhibiting the PI3K/AKT pathways, enhancing the intracellular retention of chemotherapeutic drugs, effectively reversing ABCB1-associated tumor chemotherapy resistance both in vitro and in vivo. Additionally, Rh4 shows promise in preventing OS migration and invasion in vitro, indicating its therapeutic potential in treating resistant and refractory tumors, particularly OS.
Results
Comparative reversal effect of eight ginsenosides on doxorubicin insensitivity of MG63/DXR cells
To investigate the effects of ginsenosides on doxorubicin-resistant OS cells, we conducted a CCK-8 assay for eight ginsenosides (Figure 1) based on MG63/DXR cells, a cell line derived from OS and resistant to doxorubicin. The IC50 values of the eight distinct ginsenosides to doxorubicin in MG63/DXR cells ranged from 0.2041 to 1.370 μM (Figure 2A). A comparative assessment of the impact of the eight ginsenosides on the doxorubicin sensitivity of MG63/DXR cells (Figure 2B) revealed that among the selected ginsenosides, ginsenoside Rh4 exhibited the most potent doxorubicin-resistance reversal effect (Reversal Factor, RF = 0.1872), even surpassing Rh2, which has been previously reported to possess such reversal activity (RF = 0.3881) [25]. This indicates that ginsenoside Rh4 represents a novel ginseng ingredient with an even more significant potential for reversing drug resistance than the widely studied ginsenoside Rh2. Ginsenoside Rh4, a bioactive component derived from Panax Notoginseng, is known for its remarkable pharmacological properties, particularly its anti-tumor effects [26]. However, its role in combination with chemotherapy and in reversing drug resistance has yet to be extensively researched. Consequently, ginsenoside Rh4 was selected for further investigation on its underlying mechanism of reversing chemotherapy resistance and its application in tumor treatment.
Figure 1. The types and structures of ginsenosides.
Ginsenoside Rh4 enhances doxorubicin-induced cell apoptosis in vitro
To further elucidate the impact of ginsenoside Rh4 on the MG63/DXR cell line, it was crucial first to determine its optimal concentration on this cell line. From non-inhibitory to completely inhibitory, a range of doxorubicin concentrations (0-12.8 μM) in combination with ginsenoside Rh4 (0-80 μM) was administered to the MG63/DXR cells for 48 hours, and the cell proliferation was assessed by the CCK-8 assay (Figure 2C). The findings demonstrated that the increased Rh4 concentration enhanced the sensitivity of MG63/DXR cells to doxorubicin, with the IC50 value decreasing from 2.158 μM in the control group to approximately 0.17 μM at 40 μM of Rh4. We found that 40 μM Rh4 did not significantly inhibit cell proliferation but effectively reversed drug resistance, while increasing the concentration to 80 μM resulted in the suppression of cell proliferation (Supplementary Figure 1). The apoptosis rates of the MG63/DXR cells were determined by flow cytometry with an Annexin V-FITC apoptosis assay. The treatments included doxorubicin (0.6 μM) alone, Rh4 (40 μM) alone, and a combination of both for 24 hours. The cells treated with Rh4 alone did not show significant apoptosis or necrosis compared to the control group. However, the combined treatment significantly increased the rate of apoptosis (18.64%) compared to that treated with doxorubicin alone (5%). These results suggest that ginsenoside Rh4 can significantly enhance the efficacy of doxorubicin on the MG63/DXR cell line, highlighting its potential as a synergistic agent for treating cancer with MDR.
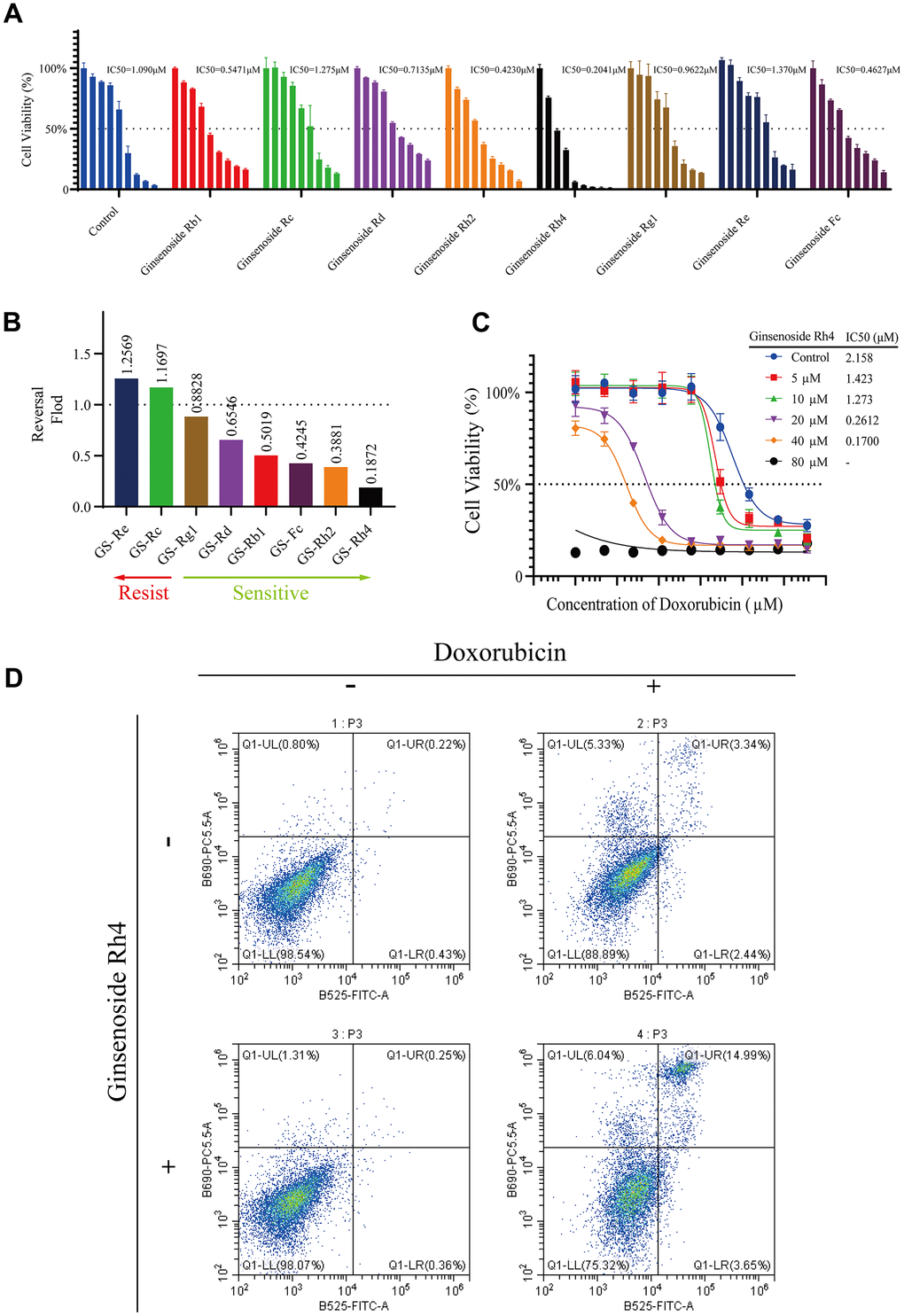
Figure 2. Modulation of doxorubicin sensitivity in MG63/DXR cells by ginsenosides. (A) The CCK-8 assay demonstrates a comparative analysis of eight ginsenosides (40 μM) in reversing doxorubicin resistance in MG63/DXR cells over 48 hours, showing the dose-response and corresponding IC50 values for each ginsenoside (40 μM). (B) The bar graph illustrates the fold change in doxorubicin sensitivity (IC50 with ginsenoside treatment/IC50 of control) in MG63/DXR cells post-treatment with the eight ginsenosides, with Rh4 exhibiting the lowest fold change, indicating the strongest reversal of resistance among the tested compounds. (C) Dose-response curves for MG63/DXR cells treated with incremental concentrations of ginsenoside Rh4 (0-80 μM) in combination with doxorubicin over 48 hours, demonstrating enhanced chemosensitivity. (D) Flow cytometry analysis of apoptosis in MG63/DXR cells treated with ginsenoside Rh4 (40 μM) and doxorubicin (0.6 μM) for 24 hours using Annexin V-FITC/PI staining. The vertical axis represents PI staining fluorescence intensity (y-axis), and the horizontal axis represents Annexin V-FITC fluorescence intensity (x-axis). Each flow cytometry plot is divided into four quadrants: LL (low signal on both axes), LR (high signal on the x-axis and low on the y-axis), UL (low signal on the x-axis and high on the y-axis), and UR (high signal on both axes). These quadrants indicate different cell states: LL for viable cells, LR for early apoptotic cells, UL for necrotic cells, and UR for late apoptotic cells. The results show a significant increase in the proportion of late apoptotic cells in the Rh4 and doxorubicin co-treatment group.
Ginsenoside Rh4 suppresses migration and invasion of MG63/DXR cells in vitro
Both MDR and metastasis are recognized as the adverse prognostic factors for OS [27]. To further investigate the impact of ginsenoside Rh4 on OS metastasis and invasion, we conducted wound healing and Transwell assays. As shown in Figure 3A, 3B, based on the wound healing assay, ginsenoside Rh4 significantly inhibited the migration of MG63/DXR cells at concentrations ranging from 20 to 40 μM. As shown in Figure 3C, 3D, based on the Transwell experiments, ginsenoside Rh4 significantly suppressed both the migration and invasion of MG63/DXR cells. These findings suggest that beyond reversing drug resistance, ginsenoside Rh4 can potentially treat OS by inhibiting metastasis and cell invasion.
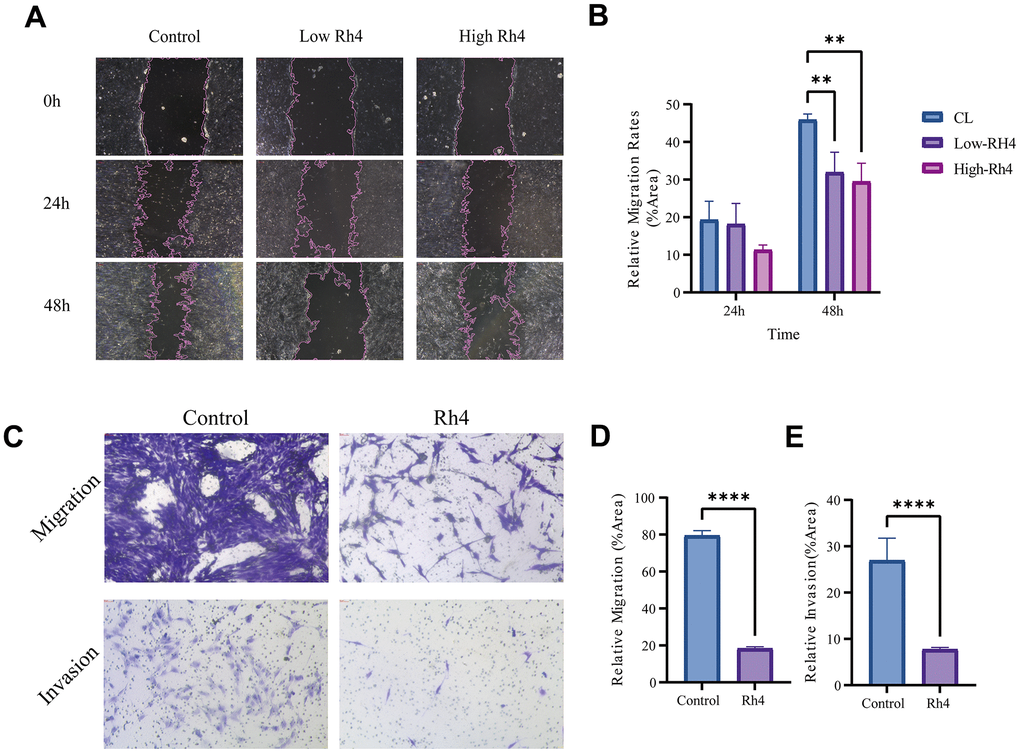
Figure 3. Impact of ginsenoside Rh4 on MG63/DXR cell migration and invasion. (A) Wound healing assay outcomes for MG63/DXR cells at 0-, 24-, and 48-hours post-treatment with control, low (20 μM), and high (40 μM) concentrations of Ginsenoside Rh4, depicting migration inhibition. (B) Quantitative analysis of the wound healing assay from panel A, indicating Ginsenoside Rh4’s effectiveness in reducing MG63/DXR cell migration. (C) Results from Transwell assays measuring MG63/DXR cell migration and invasion post-treatment with control and 40 μM ginsenoside Rh4 at 48h, revealing reduced migratory and invasive behaviors. (D) Quantitative assessment of migration from panel C, highlighting the inhibitory effect of ginsenoside Rh4 on cell migration. (E) Quantitative assessment of invasion from panel C, demonstrating ginsenoside Rh4’s ability to diminish cell invasion.
Ginsenoside Rh4 enhances the efficacy of doxorubicin in inhibiting tumor growth in vivo
The capacity of ginsenoside Rh4 to counteract doxorubicin resistance in MG63/DXR cells was evaluated by establishing a subcutaneous xenograft tumor model in the NCG-mHc1-V3 mice. The experimental methodology is elaborated in Figure 4A, with the condition of subcutaneous MG63/DXR tumors in the mice depicted in Figure 4B, 4C. The mean tumor volumes during the concurrent administration of doxorubicin and ginsenoside Rh4 are documented in Figure 4E, demonstrating that Rh4 significantly bolstered the suppressive impact of doxorubicin on tumor proliferation in vivo. Histological analyses via H&E staining (Figure 4F) unveiled the pronounced vacuolation and necrosis within the tumor cells of the xenografted tumors in the NCG-mHc1-V3 mice following treatment with both doxorubicin and Rh4, in contrast to that received monotherapy of doxorubicin. Moreover, doxorubicin administration is frequently accompanied by adverse effects, including cardiotoxicity and weight reduction [28]. As depicted in Figure 4C, a discernible trend towards weight gain was observed in the NCG-mHc1-V3 mice under the dual treatment regime compared to those received only doxorubicin (p < 0.05), intimating a potential mitigative effect of Rh4 on doxorubicin-induced toxicity. Additionally, histopathological evaluation of cardiac (Figure 4G) and hepatic (Figure 4H) tissues revealed no significant pathological alterations or exacerbation of cardiac damage in the mice treated with the combined regimen, affirming the therapeutic safety of this approach.
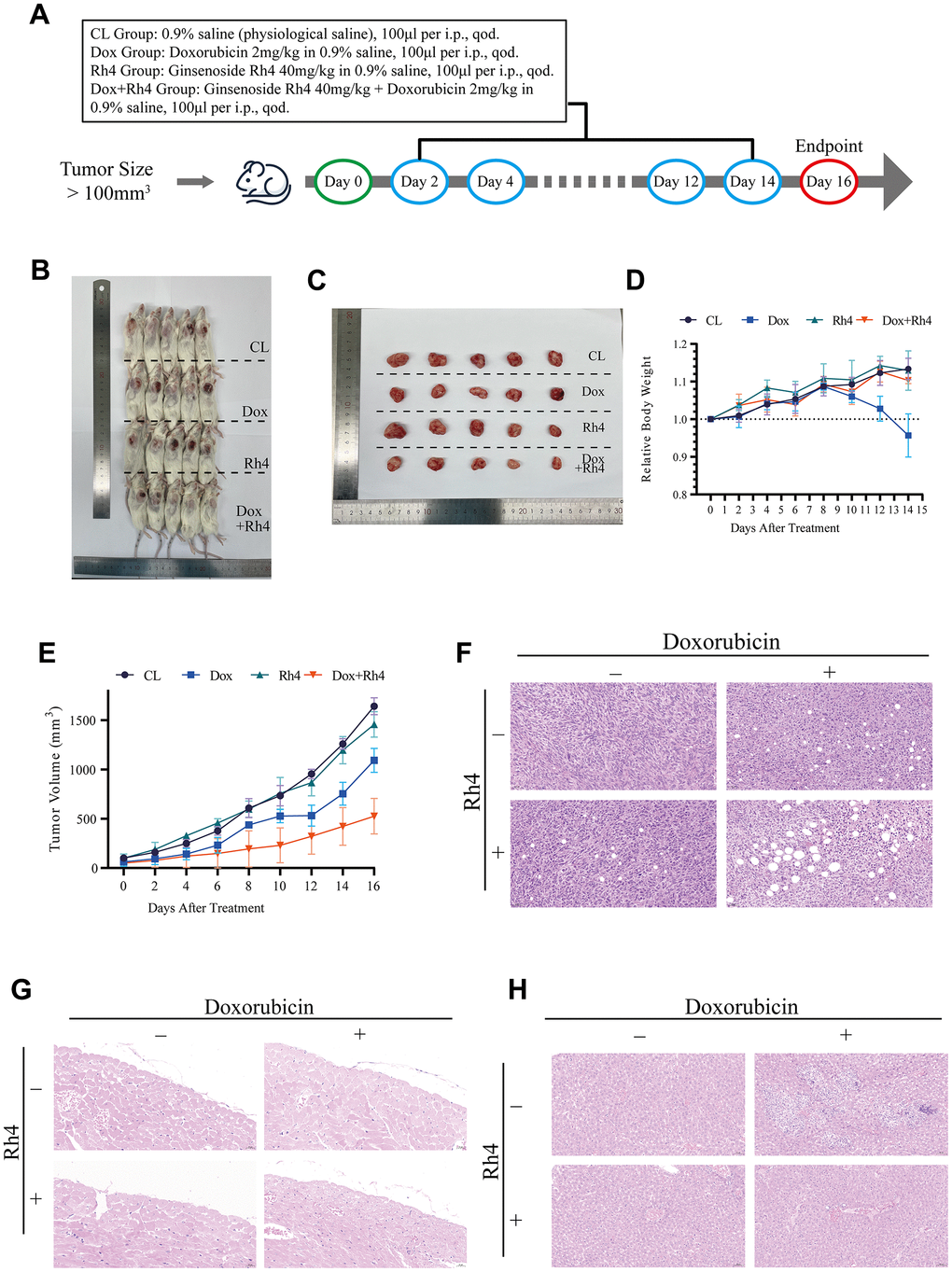
Figure 4. Therapeutic efficacy of doxorubicin combined with ginsenoside Rh4 in vivo. (A) Schematic representation of the treatment schedule for NCG mice, detailing drug administration intervals and evaluation time points. (B) Images of excised tumors at study conclusion, illustrating variance in tumor size among treatment groups. (C) Comparison of resected tumor sizes against a ruler, underscoring the impact of treatment modalities on tumor growth. (D) Line graph depicting the relative body weight changes across treatment groups, noting the absence of significant weight loss in the combination therapy group compared to the doxorubicin group. (E) Tumor volume progression curves for each treatment group, indicating a decelerated growth rate in the combination therapy cohort. (F) Histological analysis of tumor tissue sections stained with H&E, showing pronounced apoptotic and necrotic alterations in the combination therapy group. (G) H&E-stained heart tissue sections, affirming the absence of detrimental effects attributable to ginsenoside Rh4 or the combination therapy. (H) H&E-stained liver tissue sections, revealing no exacerbation of side effects due to ginsenoside Rh4 or the combined treatment.
Ginsenoside Rh4 enhances doxorubicin accumulation by inhibiting ABCB1 expression both in vitro and in vivo
The reduced intracellular drug accumulation is one of the most common causes of MDR, especially for doxorubicin resistance [29]. To clarify the effect of ginsenoside Rh4 on the intracellular accumulation of doxorubicin in the MG63/DXR cell line, we employed flow cytometry to measure the autofluorescence of doxorubicin in cells and thereby assess its intracellular accumulation [30]. As demonstrated in Figure 5A, the combined application of ginsenoside Rh4 (40 μM) and doxorubicin (0.6 μM) significantly increased the intensity of doxorubicin’s autofluorescence in cells, indicating a higher intracellular accumulation of the drug. Overexpression of ABCB1, identified as one of the most typical and common causes leading to reduced intracellular accumulation, functions by employing ATP to expel drug molecules from the cytoplasm for diminishing drug accumulation [5, 31]. The effect of ginsenoside Rh4 on ABCB1 expression was quantified via Western blot analysis in vitro (Figure 5B) and IHC staining in vivo (Figure 5C). Compared to the control group, the protein expression level of ABCB1 was significantly downregulated in the MG63/DXR cells after 48 hours treatment of Rh4 (40 μM) with or without doxorubicin (0.6 μM). These results demonstrated that ginsenoside Rh4 significantly decreased the expression levels of ABCB1 both in vitro and in vivo, leading to an increased doxorubicin accumulation.
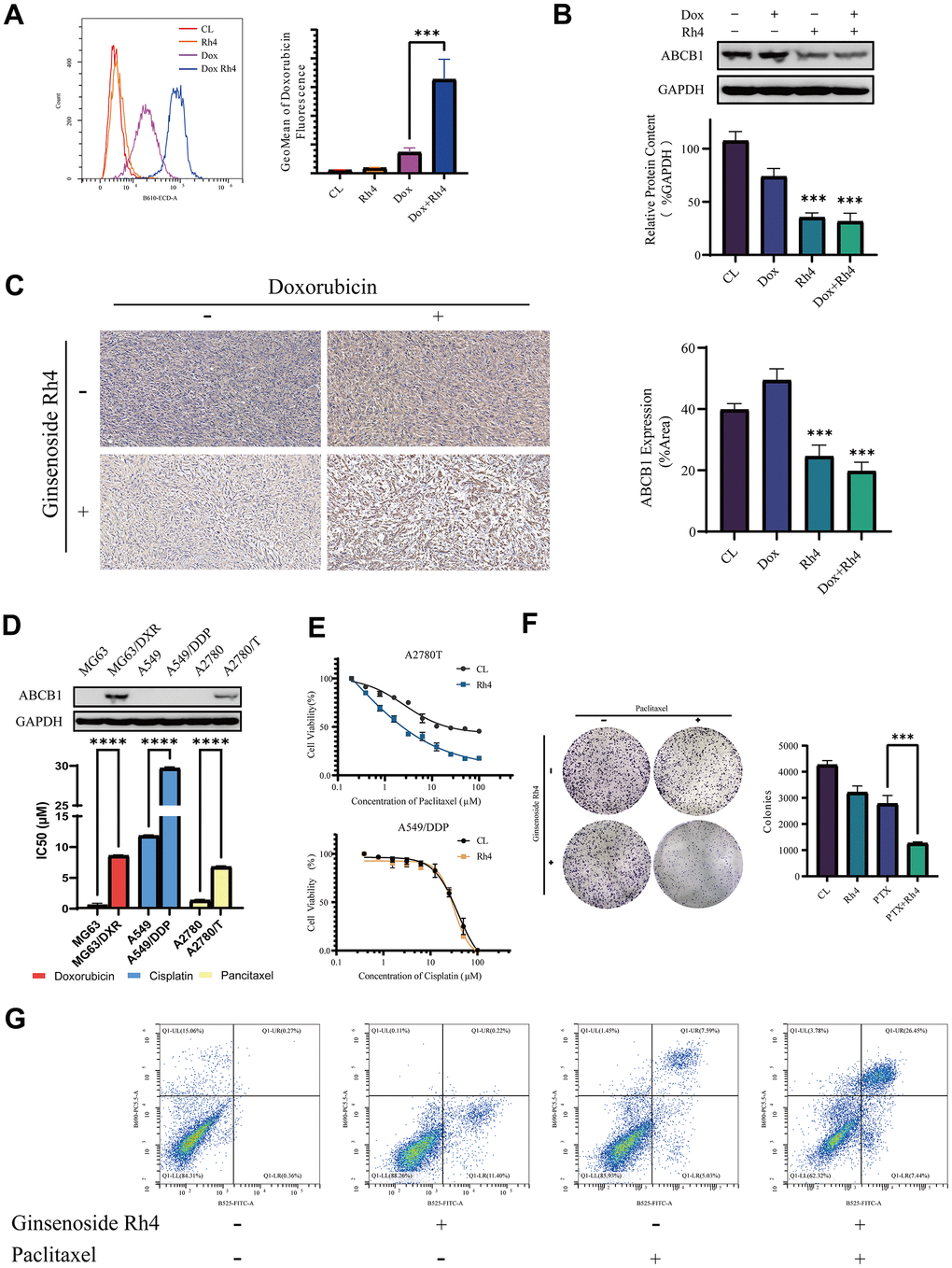
Figure 5. Molecular mechanisms underlying the reversal of drug resistance in MG63/DXR by ginsenoside Rh4. (A) An intracellular accumulation of doxorubicin in MG63/DXR cells was measured by flow cytometry, with autofluorescence intensity in the B610-ECD-A channel. The x-axis represents the fluorescence intensity, while the y-axis denotes cell count. The further right the peak of fluorescence intensity for each group, the greater the intracellular retention of doxorubicin in that group. The accompanying bar graph uses the GeoMean of fluorescence intensity to represent the amount of doxorubicin uptake for each group. (B) The expression levels of ABCB1 protein analyzed through Western blotting, employing GAPDH as an internal control. A bar graph illustrates the relative protein expression levels among the study groups. (C) Immunohistochemistry (IHC) employed to assess ABCB1 expression in vivo, with accompanying bar graphs depicting protein expression levels within each experimental group. (D) Comparative ABCB1 expression in drug-resistant versus sensitive cell lines analyzed by Western blot, including a bar graph detailing IC50 values for various cell lines and drugs, elucidating the role of ABCB1 in drug resistance. (E) Dose-response curves highlighting the sensitivities of A2780T cells to paclitaxel and A549/DDP cells to cisplatin, with and without Rh4 treatment, detailing shifts in IC50 values that signify modulation of drug sensitivity by Rh4. (F) Colony formation assays in A2780T cells exploring the synergistic effects of paclitaxel (2 μM) and Rh4 (40 μM) on cell proliferation and survival. (G) Apoptosis in A2780T cells treated with and without ginsenoside Rh4 (40 μM) and paclitaxel (2 μM) for 24 hours was analyzed using flow cytometry with Annexin V-FITC/PI staining to distinguish apoptotic cells from viable cells, highlighting the pro-apoptotic effect of Rh4 in the context of chemotherapy. In the flow cytometry plots, the y-axis represents PI staining fluorescence intensity, and the x-axis represents Annexin V-FITC fluorescence intensity. Each plot is divided into four quadrants: LL (low signal on both axes), LR (high signal on the x-axis and low on the y-axis), UL (low signal on the x-axis and high on the y-axis), and UR (high signal on both axes). These quadrants correspond to different cell states: LL for viable cells, LR for early apoptotic cells, UL for necrotic cells, and UR for late apoptotic cells. The results show a significant increase in the proportion of late apoptotic cells in the Rh4 and paclitaxel co-treatment group.
Ginsenoside Rh4 suppresses the PI3Kδ/AKT signaling pathway
We investigated the transcriptomic changes through RNA sequencing to further elucidate the mechanism underlying the reversal of MDR by ginsenoside Rh4 in the MG63/DXR cell line. We identified a total of 1,830 DEGs between the Dox+Rh4 and the Dox-only groups. Volcano plots revealed significant DEGs between the treatment and control groups, with 624 genes upregulated and 1,206 genes downregulated (Figure 6A). To pinpoint the potential pathway alterations among these DEGs, we performed KEGG pathway enrichment analysis through KOBAS-i, a machine learning-based-gene set enrichment method (Figure 6B) [36]. The results highlighted the PI3K/Akt, TNF, and mTOR signaling pathways with the highest pathway enrichment scores of 3.9779, 3.8761, and 3.8587, respectively. The DEGs related to the PI3K/AKT signaling pathway (p < 0.05) are visualized in a heatmap (Figure 6C), revealing the inhibition of the vital protein PI3KCD (PI3Kδ) at the mRNA level.
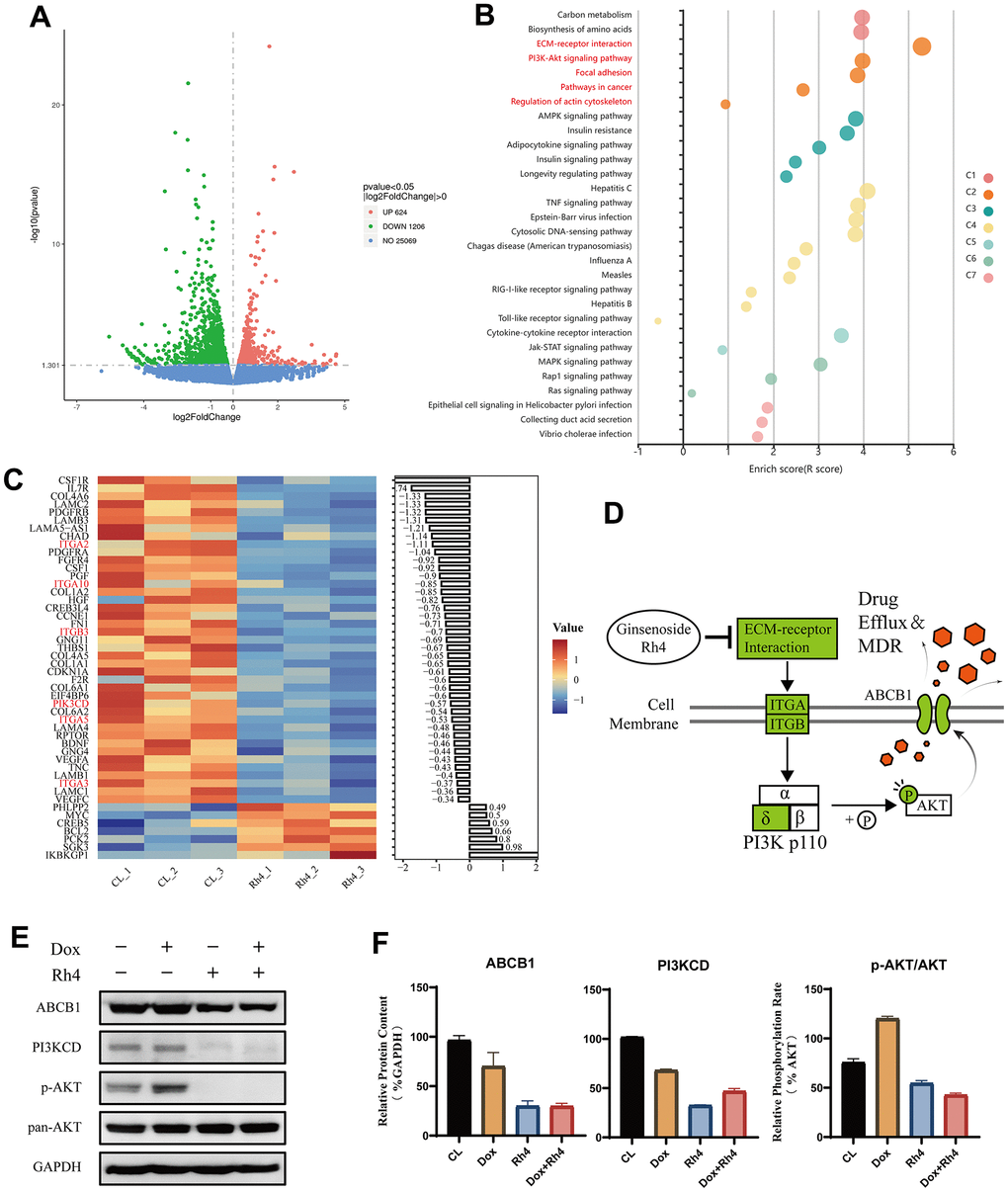
Figure 6. Differential gene expression and pathway analysis influenced by ginsenoside Rh4. (A) Volcano plot visualization of differentially expressed genes (DEGs) between the control (CL) and Rh4-treated groups, highlighting significant gene expression changes. (B) Enrichment results of KEGG pathways for DEGs between CL and Rh4 groups, depicted in various colors to represent different clusters as identified by the KOBAS-i model. (C) Heatmap representation of the regulation of PI3K/AKT pathway-related genes among DEGs between CL and Rh4 groups, illustrating the detailed gene expression modulation within this pathway. (D) Schematic representation of Ginsenoside Rh4’s modulation of ABCB1 expression via the PI3K/AKT pathway, where green denotes suppression of related gene expression. (E) Western blot analysis of ABCB1 and PI3K/AKT pathway-related protein levels, using GAPDH as an internal control. A bar graph shows the relative protein expression levels across the study groups, indicating the impact of Rh4 on these key regulatory proteins.
PI3Ks (phosphoinositide 3-kinases) are dimers composed of a regulatory subunit, p85 (Class II), and a catalytic subunit, p110 (Class I), including four isoforms: p110α, -β, -γ, and -δ [37]. Previous reports suggest that CRISPR/Cas9 knocking out the PI3Kα and β subunits to inhibit the PI3K/AKT signaling pathway can overcome P-gp and BCRP (Breast cancer resistance protein, ABCG2)-mediated MDR in cancer [38]. We proposed that ginsenoside Rh4 could inhibit the PI3Kδ subunit, thus obstructing the PI3K/AKT signaling pathway and reducing the levels of ABCB1 (Figure 6D). To confirm our hypothesis, the protein levels of ABCB1, PI3KCD, p-AKT, and AKT were evaluated. A marked decrease was found in the levels of ABCB1 and PI3KCD following a 24 hours treatment of ginsenoside Rh4 (40 μM), regardless of the co-treatment with doxorubicin (0.6 μM). The p-AKT/AKT ratio, considered as an indicator of the activation of the PI3K/AKT pathway, was reduced by Rh4 treatment along as well as in combination with doxorubicin, while it was elevated by doxorubicin treatment alone. These findings reinforce the results from the transcriptomic analysis, verifying that ginsenoside Rh4 could effectively suppress the PI3Kδ/AKT signaling pathway.
Ginsenoside Rh4 inhibits ABCB1 and reverses drug resistance via the PI3Kδ/AKT signaling pathway
To elucidate the mechanism through which ginsenoside Rh4 reverses drug resistance, we conducted experiments validating the correlation between the PI3Kδ/AKT pathway and doxorubicin resistance in the MG63/DXR cells. We designed RNA interference against PI3KCD and a strategy of transient transfection for overexpression. The MG63/DXR cells were categorized into the control, siPI3KCD (RNA interference), OE-PI3KCD (overexpression), and OE-PI3KCD-Rh4 (overexpression with Rh4 co-treatment) groups to evaluate the doxorubicin-induced cell proliferation over 48 hours by a CCK-8 assay (Figure 7A). The results demonstrate a direct correlation between the expression levels of PI3KCD and the IC50 of doxorubicin in the MG63/DXR cell lines. Overexpression of PI3KCD resulted in increased resistance to doxorubicin (RF = 1.0627), whereas inhibition of PI3KCD increased the sensitivity (RF = 0.2452). Moreover, the Rh4-induced reversal of drug resistance was significantly diminished by PI3KCD overexpression, with the RF increasing from 0.2481 (Rh4 alone) to 0.6151, indicating a positive correlation between the PI3KCD levels and doxorubicin resistance in the MG63/DXR cells. Further validation at the protein level revealed that inhibition of PI3KCD expression (Figure 7B) could suppress the PI3Kδ/AKT pathway and decrease the ABCB1 expression compared to the controls. Conversely, overexpressing PI3KCD activated the PI3Kδ/AKT pathway and increased ABCB1 expression. In the presence of Rh4 co-treatment, the inhibitory effect of Rh4 on the PI3Kδ/AKT pathway and ABCB1 expression was counteracted by PI3KCD overexpression (Figure 7C). These results confirm our hypothesis that ginsenoside Rh4 could inhibit the PI3Kδ subunit, thereby obstructing the PI3K/AKT signaling pathway and downregulating ABCB1.
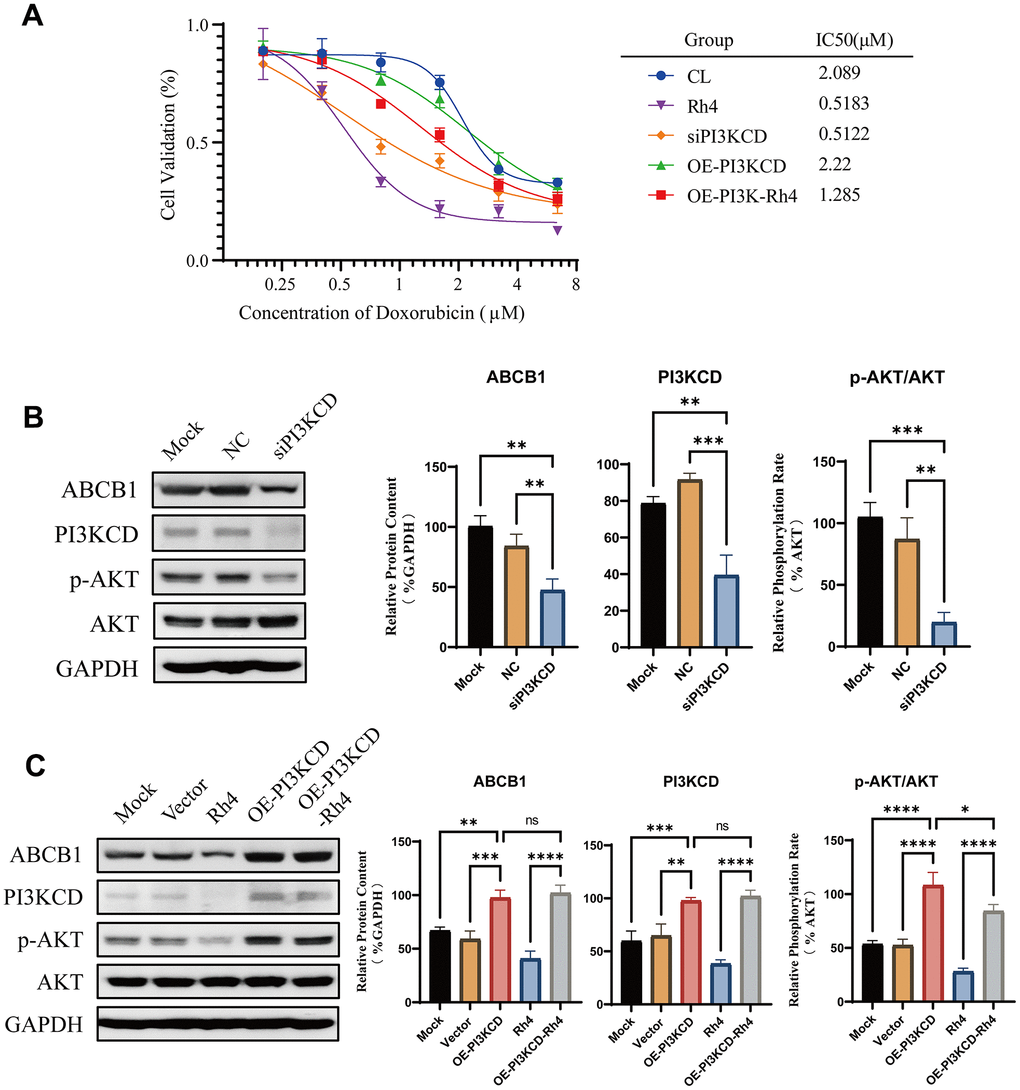
Figure 7. Rescue experiment validation of ginsenoside Rh4 reversing drug resistance via the PI3K/AKT axis. (A) Drug sensitivity curves for MG63/DXR cells to doxorubicin, illustrating the effects of PI3KCD inhibition or overexpression on drug inhibition efficiency. (B) Western blot analysis of ABCB1 and PI3K/AKT pathway-related protein levels upon PI3KCD inhibition, employing GAPDH as an internal standard to ensure consistent quantification across samples. (C) Western blot analysis detailing the levels of ABCB1 and PI3K/AKT pathway-related proteins following PI3KCD overexpression, with GAPDH used as a baseline for normalization.
Discussion
MDR in OS treatment remains a significant clinical challenge [39–41]. With the enrichment of the chemical compound library, numerous natural products have emerged as potential anticancer and anti-MDR agents, such as curcumin, quercetin, ginsenoside, tanshinone, etc. [42–44]. These phytochemicals possess distinct advantages in combating MDR by providing unique benefits such as organ protection and multifaceted efficacy. Ginsenosides are compounds extracted from Panax Ginseng. Ginsenosides Rh2 and Rg1 have been reported for their effectiveness in reversing drug resistance [20, 22], while many other ginsenoside types remain to be examined.
The present study identifies ginsenoside Rh4 as an effective medicinal ingredient in reversing ABCB1-related drug resistance, particularly doxorubicin resistance in OS. This study extends research on reversing tumor drug resistance by ginsenosides and represents the first report on using ginsenoside Rh4 for OS treatment. We first conducted a CCK-8 assay to screen potential ginsenosides, including those previously reported to reverse drug resistance, such as Rh2 and Rg1, as well as the unexplored variants like Rb1 and Rh4. This screening identified Rh4 with superior capabilities in reversing drug resistance. Ginsenoside Rh4, as a rare and novel active ingredient of ginsenosides, has shown therapeutic potential, including for cancer treatment, cardiovascular health, and neuroprotection [45–47]. In the realm of cancer therapy, Rh4 is recognized to induce cell cycle arrest, foster apoptosis in cancer cells, and curb metastasis through the modulation of critical signaling pathways, notably JAK2/STAT3 and Wnt/β-catenin pathways [46]. The neuroprotective attributes of Rh4 are particularly evident in models of Alzheimer’s disease and depression, where it contributes to the reduction of amyloid-beta plaque formation and the alleviation of depression-like behaviors [45]. However, the associations of Rh4 with drug resistance and OS treatment remain unexplored. Given the lack of research on Rh4’s efficacy in OS treatment, we evaluated the cell viability following exposure to Rh4 at different concentrations. We found that 40 μM Rh4 notably enhanced the effect of doxorubicin on MG63/DXR cells, establishing this concentration as optimal both in vitro and in vivo. In contrast, higher concentrations, specifically 80 μM, of Rh4, induced cytotoxicity in MG63/DXR cells. Our study also indicated that 40 mg/kg of Rh4 could improve body weight without inducing significant adverse effects on cardiac or hepatic tissues, compared to chemotherapy alone. Furthermore, our in vitro assessments demonstrated Rh4’s capability to inhibit the migration and invasion of OS cells, highlighting its multifaceted therapeutic advantages in cancer treatment.
Cancer cells can employ diverse strategies to evade or neutralize the cytotoxic effects induced by anticancer treatments. One of the primary mechanisms is through reducing drug accumulation, especially by the overexpression of ABCB1 [48]. To elucidate the mechanisms underlying the reversal of drug resistance by ginsenoside Rh4, we initially examined the intracellular doxorubicin retention and found that it was increased by ginsenoside Rh4. Subsequently, we introduced Rh4 to the A549/DDP cell line, which is resistant to cisplatin without ABCB1 overexpression, and the A2780T cell line, which is resistant to paclitaxel with high ABCB1 expression, to examine if other mechanisms exist. We found that ginsenoside Rh4 could reverse the drug resistance in the A2780T cell line but not in A549/DDP, highlighting its targeted selectivity towards ABCB1-related resistance.
We conducted RNA sequencing to investigate the molecular mechanisms behind such effects of ginsenoside Rh4. The results from RNASeq and KEGG pathway enrichment analysis indicated that a majority of the genes significantly enriched the PI3K/AKT signaling pathway. This finding was further validated at the protein level, demonstrating that ginsenoside Rh4 could inhibit the PI3Kδ/AKT pathway. Suppression of the PI3K/AKT signaling pathway is known to play a crucial role in cell survival, apoptosis, and drug resistance [49–53]. Lei et al. have shown that knocking out the α and β subunits of the PI3K 110p can inhibit the expression level of ABCB1, while the effect of PI3Kδ inhibition remains unverified [38]. To confirm such an effect, we employed RNA interference and transient transfection for PI3KCD gene overexpression. The results elucidated that ginsenoside Rh4 could reverse drug resistance by inhibiting PI3Kδ levels.
In the treatment of osteosarcoma and various solid tumors, MDR (multidrug resistance) and metastasis are significant factors leading to poor patient prognosis. In this study, ginsenoside Rh4 demonstrated positive effects in combating both MDR and tumor metastasis, making it a promising candidate for the clinical management of osteosarcoma and other tumors at risk for MDR and metastasis. Additionally, this study reported the PI3K/AKT targeting properties of ginsenoside Rh4, which shares common targets with some PI3K/AKT targeted drugs, such as Copanlisib and Alpelisib [54, 55]. These drugs have been approved for the treatment of advanced breast cancer and relapsed follicular lymphoma (FL) in second- and third-line therapies, respectively. Therefore, we suggest that future research should explore the potential application of ginsenoside Rh4 in breast cancer and lymphoma.
Despite obtaining some exciting results, this study has certain limitations that need to be explored in future research.
Firstly, ginsenoside Rh4 inhibited the migration and invasion of MG63/DXR cells in our study, but its underlying molecular mechanisms remain unclear. Our RNA sequencing results showed significant inhibition of extracellular matrix (ECM) receptors by ginsenoside Rh4. In future research, we suggest an in-depth investigation into the inhibitory effects of ginsenoside Rh4 on ECM-related receptors to understand its comprehensive impact on cell migration and invasion.
Secondly, this study did not elucidate the effects of ginsenoside Rh4 on the upstream and downstream molecules of the PI3K/AKT pathway. Based on the results of this study, we recommend focusing on ITGA2, an upstream regulator of PI3K, as a potential target of ginsenoside Rh4 in subsequent research. We also suggest further exploration of the regulatory mechanisms of downstream molecules of the PI3K/AKT pathway, such as mTOR and NF-κB, which are competitive downstream targets of AKT. These studies will help us to better understand the diverse pharmacological effects of ginsenoside Rh4.
Thirdly, this study reported that the reversal effect of ginsenoside Rh4 on drug resistance was effective in some resistant cell lines (MG63/DXR, A2780T) but not in others (A549/DDP). However, this study did not further validate the reasons for this difference. Based on our results, we believe this discrepancy is due to the different primary drug resistance mechanisms in the two resistant cell lines (A549/DDP does not have high ABCB1 expression). We suggest that further research should focus on how the reversal effect of ginsenoside Rh4 varies in resistant cell lines with different resistance mechanisms (e.g., MRP1, BCRP) to draw comprehensive conclusions about the role of ginsenoside Rh4 in overcoming tumor chemotherapy resistance.
In summary, this study employed a comprehensive approach, including both in vitro and in vivo models, to report for the first time the application of ginsenoside Rh4 in the treatment of osteosarcoma (OS). This study not only confirmed the effective reversal activity of ginsenoside Rh4 against doxorubicin resistance but also highlighted its safety. At the molecular biology level, ginsenoside Rh4 has the potential to act as a natural PI3Kδ inhibitor to mitigate ABCB1-related MDR.
Materials and Methods
Chemicals and reagents
Ginsenosides Rb1 (DR0006), Rc (DR0013), Rd (DR0015), Re (DR0014), Rg1 (DR0009), Rh2 (DS0036), Rh4 (DR0020), and Fc (DS0195), with purity exceeding 95%, were purchased from Lemeitian Medicine Co., Ltd. (Chengdu, China), and their chemical structures are illustrated in Figure 1. Cisplatin (HY-17394), doxorubicin (HY-15142A), paclitaxel (HY-B0015), N,N-dimethylformamide (DMF, HY-Y0345), and dimethyl sulfoxide (DMSO, HY-Y0320) were supplied by MedChemExpress (Shanghai, China). Cell Counting Kit-8 (CCK-8, CX001) reagent, phosphate-buffered saline (PBS, CB012), Dulbecco’s Modified Eagle Medium (DMEM, CB001), and Roswell Park Memorial Institute 1640 (RPMI-1640, CB004) media were procured from Epizyme Biomedical Technology (Shanghai, China). Fetal bovine serum (FBS, CSFBS1500) was obtained from CellSera Biotechnology Co., Ltd. (Rutherford, Australia). The Annexin V-FITC apoptosis detection kit (C1062M) was purchased from Beyotime Biotechnology Co., Ltd. (Shanghai, China). Antibodies against ABCB1 (22336-1-AP) and GAPDH (60004-1-Ig) were acquired from Proteintech Group, Inc. (MA, USA). Additionally, antibodies targeting phosphorylated AKT (#9271), AKT (#4691), and PI3Kδ (ab109006) were sourced from CST (MA, USA) and Abcam Group, Inc. (Cambridge, UK), respectively.
Cell lines and animals
The OS cell lines MG63 and MG63/DXR were graciously provided by Professor Zhang Chunlin at the Department of Orthopedic Surgery, Institute of Bone Tumor, Shanghai Tenth People’s Hospital, affiliated with Tongji University School of Medicine (Shanghai, China). The ovarian cancer cell lines A2780 and A2780T were kindly donated by Professor Han Yonglong at the Department of Pharmacy, Shanghai Jiao Tong University Affiliated Sixth People’s Hospital (Shanghai, China). The lung cancer cell lines A549 and A549/DDP were purchased from Shanghai Whelab Bioscience Ltd. The cell lines were cultured in either DMEM or RPMI-1640 medium in 10 cm diameter dishes, supplemented with 10% FBS, in a humidified atmosphere of 5% CO2 at 37° C. Subculturing was performed every two days at a 1:3 ratio. Female NCG-mHc1-V3 mice, aged four weeks and weighing approximately 20 ± 2 g, were acquired from GemPharmatech Co., Ltd. (Nanjing, China). They were maintained in a specific pathogen-free environment to ensure optimal health and experimental conditions. Twenty-four female NCG-mHc1-V3 mice were utilized for the establishment of a subcutaneous xenograft tumor model. Following acclimatization for three days, the mice received subcutaneous injections of 1*10^6 cells of MG63/DXR per mouse under the skin of the right forelimb axilla. Daily observations were made until the subcutaneously implanted tumor volume grew to over 100 mm3. After the implanted tumor model reached the load standard, a regimen of intraperitoneal injection every other day was adopted for drug administration. The drug was prepared using DMSO to make a stock solution, which was then diluted with saline in a 1:9 ratio just before injection. Body weight and tumor size changes were monitored every two days before drug administration, continuing for two weeks. At the end of the experiment, the mice were euthanized under deep anesthesia to collect samples of the implanted tumors, heart, and liver.
Cell proliferation assay
The cytotoxic effects of ginsenosides (Rb1, Rc, Rd, Re, Rg1, Rh2, Rh4, and Fc), cisplatin, doxorubicin, and paclitaxel were evaluated by the CCK-8 assay. After detachment with 0.25% trypsin, the cells were plated in 96-well plates at a density of 5 × 10^4 cells/well. Upon reaching over 80% confluence with spindle-shaped adherence (typically after 24 hours of culture), the cells were treated with the test compounds for 48 hours. The concentrations of the test compounds were determined based on the IC50 values obtained from preliminary experiments, typically using a range of IC50 x 0.0625, x 0.125, x 0.25, x 0.5, x 1, x 2, x 4, x 8, and x 16. The ginsenosides and cisplatin were dissolved in DMSO and DMF, respectively, with solvent concentrations below 0.1%. The experiments were performed in triplicate. Cell proliferation levels were determined using the CCK-8 assay kit. Optical density (OD) was measured at 480 nm using a Varioskan Flash microplate reader (Thermo Fisher Scientific, MA, USA). The inhibition rate for cell proliferation was calculated as (1 - average OD of the treated samples / average OD of the control samples) × 100%. Half-maximal inhibitory concentration (IC50) values were determined using the GraphPad Prism software.
Hematoxylin and eosin (H&E) staining and histological analysis
Tissue samples were fixed in 4% paraformaldehyde for 24 hours at 4° C, dehydrated in an ethanol series, cleared in xylene, and embedded in paraffin. The sections (5 μm) were cut and placed on slides for staining. For H&E staining, the sections were deparaffinized, rehydrated, and stained with hematoxylin for nuclei visualization, followed by eosin for cytoplasmic visualization. The slides were then dehydrated, cleared, and mounted. For immunohistochemistry (IHC), the sections underwent antigen retrieval in citrate buffer (pH 6.0), blocked for non-specific binding, and incubated with primary antibodies specific to target proteins overnight at 4° C. After washing, the sections were incubated with horseradish peroxidase-conjugated secondary antibodies, developed using 3,3’-Diaminobenzidine (DAB), counterstained with hematoxylin, and mounted. The stained slides were examined under a light microscope. Images were captured for qualitative assessment of tissue morphology and specific protein expression. The staining areas were compared between different tissue samples based on the statistical color models (n ≥ 5) [56].
Colony formation assay
The cell proliferation capacity was evaluated through a colony formation assay. Cells were seeded in six-well plates at a density of 5,000 cells per well and treated with Rh4 (40 μM), paclitaxel (5 μM), and their combination for 48h. The culture was maintained for a week, with media refreshments every two days. Following incubation, the cells were fixed with 95% methanol for 15 minutes at room temperature and stained with 0.1% crystal violet for five minutes. After washing with PBS, the plates were photographed. The stained areas were compared across different samples (n ≥ 5), quantitatively measuring the colony formation ability.
Wound healing assay
For the wound healing assay, MG63/DXR cells in logarithmic growth phase were seeded into six-well plates at 1 × 106 cells per well and incubated for 24 hours. A sterile pipette tip was used to create a scratch in a tri-horizontal pattern across the cell layer, after which the cells were washed with PBS to capture images under a microscope. The cells were then treated with Rh4 in serum-free medium at 0, 20, and 40 μM concentrations during the assay. At least five images per area were taken at the same location at 0, 24, and 48 hours. The efficiency of cell migration was evaluated by measuring the wound healing rate, which was calculated using the Image J software.
Transwell migration and invasion assays
Migration and invasion assays were conducted using 24-well plates (Corning Incorporated, NY, USA). MG63/DXR cells were seeded at 1 × 104 cells per well in the upper chamber of a Transwell apparatus with serum-free medium for the migration assay. The lower chamber was filled with 600 μL of 20% FBS medium. The cells were incubated with or without Rh4 (40 μM) for 24 hours. The invasion assay was differentiated by pre-coating the upper chamber with a 1:5 dilution of Matrigel (Corning, NY, USA) in a serum-free medium. Following incubation, the filters fixed in methanol were stained with 0.1% crystal violet. Images were captured, and the cells were counted in five randomly selected fields.
Doxorubicin accumulation assay
The intracellular accumulation of doxorubicin was quantified by flow cytometry, with its autofluorescence detected at 590 nm in the B610-ECD-A channel. Cells were cultured in six-well plates for eight hours before doxorubicin treatment for an additional eight hours, with experimental groups receiving a 24-hour pre-treatment with Rh4 (40μM). Following the treatment, the cells were trypsinized, collected, and washed with PBS twice before resuspension in 0.5 ml PBS for analysis. A Cytoflex XL (Beckman Coulter, CA, USA) was utilized to analyze 10,000 events per sample, quantifying drug uptake through mean fluorescence intensity in the B610-ECD-A channel.
Cell apoptosis assay
Apoptosis was evaluated using an Annexin V-FITC/ propidium iodide (PI) Apoptosis Detection Kit (Beyotime Biotechnology Co., Ltd., Shanghai, China). Cells were treated under various conditions in six-well plates for 48 hours, harvested, and PBS-resuspended. Following centrifugation, 195 μl of Annexin V-FITC binding buffer was added to the cell pellet. The cells were stained with Annexin V-FITC and PI for 15 minutes and then analyzed using a Cytoflex XL (Beckman Coulter, CA, USA), with approximately 10,000 cells evaluated per sample. Data were processed using the CytExpert 2.4 software for detailed apoptosis assessment across different groups.
RNA sequencing analysis, identification of differentially expressed genes (DEGs), and KEGG enrichment analysis
Cell samples from the control (CON), doxorubicin treatment (Dox), and combination treatment (Dox+Rh4) groups (n = 3 for each) were treated with doxorubicin and Rh4 at concentrations of 0.5 μM and 40 μM, respectively, for 24 hours. Samples were submitted to Novogene Bioinformatics Technology Co., Ltd. (Tianjin, China) for RNA sequencing. RNA was extracted using the TRIzol reagent, followed by quality assessment and library preparation for sequencing on the Illumina NovaSeq 6000 platform. Sequencing data were processed to capture and convert fluorescence signals into sequence information via the CASAVA software. Read counts per gene were determined using FeatureCounts, and the FPKM (fragments per kilobase of transcript per million fragments mapped) values were calculated to quantify gene expression levels. DEGs were identified using the DESeq2 software, with a significance threshold set at p < 0.05. Functional enrichment of the DEGs was conducted through KEGG pathway analysis using the KOBAS-i software to understand the biological pathways affected by the treatments [36].
Western blotting analysis
The cells were cultured and treated as described above. Cells were treated with Rh4 (40 μM) and Dox (0.5 μM) as needed for 48 hours. After 48 hours of stimulation, the cell precipitates were collected and lysed on ice using 200 μl of RIPA lysis buffer for five minutes. Subsequently, total protein was extracted by centrifugation for 20 minutes at 15,000 rpm and 4° C, and the protein concentrations were determined using an enhanced BCA protein assay kit (Epizyme Biomedical Technology, Shanghai, China). For each sample, 1 μl of SDS-PAGE protein loading buffer (5X) was added to every 4 μl of the protein sample. The 7.5% SDS-PAGE kit (Epizyme Biomedical Technology, Shanghai, China) was prepared according to the manufacturer’s instructions. A total of 10 μg of protein per lane was loaded, and the dilution of primary and secondary antibodies was determined based on preliminary experiments. In this study, the antibody dilution ratios were as follows: ABCB1 1:10,000, PIK3CD 1:2,500, p-AKT and AKT 1:5,000, and GAPDH 1:20,000. Finally, protein bands were visualized using the BeyoECL Star Kit (Merck, NJ, USA) and a chemiluminescence module on the Amersham ImageQuant 800 Gel imaging system (GE Healthcare, IL, USA). Band intensities were semi-quantified using the Image J software, with relative protein expression normalized to GAPDH (internal control protein).
Overexpression and siRNA of PI3Kδ and transfection
The siRNA and overexpression plasmid for human PI3KCD, alongside their respective controls, were obtained from GenePharma (Shanghai, China), with sequences detailed in Supplementary File 1 and Supplementary Table 1. Transfection was performed using the jetPRIME reagent (Polyplus, NY, USA). The siRNA or overexpression plasmid was diluted in jetPRIME buffer, mixed with the jetPRIME reagent, and incubated for 10 minutes. This mixture was then applied to the cells. After eight hours of incubation with the siRNA or plasmid transfection mixture, the medium was replaced with fresh culture medium. The cells were then further treated with ginsenoside Rh4 and/or doxorubicin for 48 hours, and gene expression was assessed by Western blot.
Statistical analysis
Statistical analysis was performed with the SPSS 24.0 and GraphPad Prism software. One-way analysis of variance and Dunnett’s test was conducted to analyze the difference between groups, with p < 0.05 considered statistically significant. “*” was used to denote significance levels, as * for p < 0.05, ** for p < 0.01, and *** for p < 0.001.
Author Contributions
TX completed the cell proliferation assay, doxorubicin accumulation assay, cell apoptosis assay, RNA sequencing analysis, Western blotting analysis, PI3Kδ siRNA inhibition and overexpression, and animal experiments, and contributed to the conceptualization, methodology, formal analysis, visualization, and preparation of the original draft. HC, YZ, XS, and YX completed the colony formation assay, wound healing assay, Transwell migration and invasion assays, assisted with the animal experiments, and contributed to the investigation, data curation, validation, and resources. ZS provided guidance for the animal experiments and contributed to the supervision of the work and funding acquisition. DM and YW contributed to the guidance and supervision of the work, funding acquisition, and review of the manuscript. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Ethical Statement
All animal-related experiments were conducted in strict accordance with the ethical standards and guidelines for animal care as established by the Ethical Committee of Shanghai Jiao Tong University Affiliated Sixth People’s Hospital. The animal welfare and ethical review for this study were provided by Shanghai Yishang Biotechnology Co., Ltd. The approval number is YS-JL-001.
Funding
This study was supported by National Key Research Project of Science and Technology Ministry (grant number: 2023YFC2411405), National Natural Science Foundation of China (grant number: 81972521) and Science and Technology Innovation Action Plan of Shanghai Municipal Science and Technology Commission (grant number: 21ZR1449000).
References
- 1. Belayneh R, Fourman MS, Bhogal S, Weiss KR. Update on Osteosarcoma. Curr Oncol Rep. 2021; 23:71. https://doi.org/10.1007/s11912-021-01053-7 [PubMed]
- 2. Aljubran AH, Griffin A, Pintilie M, Blackstein M. Osteosarcoma in adolescents and adults: survival analysis with and without lung metastases. Ann Oncol. 2009; 20:1136–41. https://doi.org/10.1093/annonc/mdn731 [PubMed]
- 3. Lilienthal I, Herold N. Targeting Molecular Mechanisms Underlying Treatment Efficacy and Resistance in Osteosarcoma: A Review of Current and Future Strategies. Int J Mol Sci. 2020; 21:6885. https://doi.org/10.3390/ijms21186885 [PubMed]
- 4. Gill J, Gorlick R. Advancing therapy for osteosarcoma. Nat Rev Clin Oncol. 2021; 18:609–24. https://doi.org/10.1038/s41571-021-00519-8 [PubMed]
- 5. Dean M, Moitra K, Allikmets R. The human ATP-binding cassette (ABC) transporter superfamily. Hum Mutat. 2022; 43:1162–82. https://doi.org/10.1002/humu.24418 [PubMed]
- 6. Huang J, Ecker GF. A Structure-Based View on ABC-Transporter Linked to Multidrug Resistance. Molecules. 2023; 28:495. https://doi.org/10.3390/molecules28020495 [PubMed]
- 7. Serra M, Pasello M, Manara MC, Scotlandi K, Ferrari S, Bertoni F, Mercuri M, Alvegard TA, Picci P, Bacci G, Smeland S. May P-glycoprotein status be used to stratify high-grade osteosarcoma patients? Results from the Italian/Scandinavian Sarcoma Group 1 treatment protocol. Int J Oncol. 2006; 29:1459–68. https://doi.org/10.3892/ijo.29.6.1459 [PubMed]
- 8. Belisario DC, Akman M, Godel M, Campani V, Patrizio MP, Scotti L, Hattinger CM, De Rosa G, Donadelli M, Serra M, Kopecka J, Riganti C. ABCA1/ABCB1 Ratio Determines Chemo- and Immune-Sensitivity in Human Osteosarcoma. Cells. 2020; 9:647. https://doi.org/10.3390/cells9030647 [PubMed]
- 9. Serra M, Picci P, Ferrari S, Bacci G. Prognostic value of P-glycoprotein in high-grade osteosarcoma. J Clin Oncol. 2007; 25:4858–60. https://doi.org/10.1200/JCO.2007.13.0534 [PubMed]
- 10. Yang X, Yang P, Shen J, Osaka E, Choy E, Cote G, Harmon D, Zhang Z, Mankin H, Hornicek FJ, Duan Z. Prevention of multidrug resistance (MDR) in osteosarcoma by NSC23925. Br J Cancer. 2014; 110:2896–904. https://doi.org/10.1038/bjc.2014.254 [PubMed]
- 11. Liu T, Li Z, Zhang Q, De Amorim Bernstein K, Lozano-Calderon S, Choy E, Hornicek FJ, Duan Z. Targeting ABCB1 (MDR1) in multi-drug resistant osteosarcoma cells using the CRISPR-Cas9 system to reverse drug resistance. Oncotarget. 2016; 7:83502–13. https://doi.org/10.18632/oncotarget.13148 [PubMed]
- 12. Dong J, Qin Z, Zhang WD, Cheng G, Yehuda AG, Ashby CR
Jr , Chen ZS, Cheng XD, Qin JJ. Medicinal chemistry strategies to discover P-glycoprotein inhibitors: An update. Drug Resist Updat. 2020; 49:100681. https://doi.org/10.1016/j.drup.2020.100681 [PubMed] - 13. He ZX, Zhao TQ, Gong YP, Zhang X, Ma LY, Liu HM. Pyrimidine: A promising scaffold for optimization to develop the inhibitors of ABC transporters. Eur J Med Chem. 2020; 200:112458. https://doi.org/10.1016/j.ejmech.2020.112458 [PubMed]
- 14. Qu J, Ke F, Yang X, Wang Y, Xu H, Li Q, Bi K. Induction of P-glycoprotein expression by dandelion in tumor and heart tissues: Impact on the anti-tumor activity and cardiotoxicity of doxorubicin. Phytomedicine. 2022; 104:154275. https://doi.org/10.1016/j.phymed.2022.154275 [PubMed]
- 15. Lee E, Enomoto R, Koshiba C, Hirano H. Inhibition of P-glycoprotein by wogonin is involved with the potentiation of etoposide-induced apoptosis in cancer cells. Ann N Y Acad Sci. 2009; 1171:132–6. https://doi.org/10.1111/j.1749-6632.2009.04722.x [PubMed]
- 16. Kim TH, Shin S, Yoo SD, Shin BS. Effects of Phytochemical P-Glycoprotein Modulators on the Pharmacokinetics and Tissue Distribution of Doxorubicin in Mice. Molecules. 2018; 23:349. https://doi.org/10.3390/molecules23020349 [PubMed]
- 17. Zhao W, Ma J, Zhang Q, Zhang H, Ma W, Li S, Piao Y, Zhao S, Dai S, Tang D. Ginsenoside Rg3 overcomes tamoxifen resistance through inhibiting glycolysis in breast cancer cells. Cell Biol Int. 2024; 48:496–509. https://doi.org/10.1002/cbin.12123 [PubMed]
- 18. Zhou B, Xiao X, Xu L, Zhu L, Tan L, Tang H, Zhang Y, Xie Q, Yao S. A dynamic study on reversal of multidrug resistance by ginsenoside Rh2, in adriamycin-resistant human breast cancer MCF-7 cells. Talanta. 2012; 88:345–51. https://doi.org/10.1016/j.talanta.2011.10.051 [PubMed]
- 19. Liu GW, Liu YH, Jiang GS, Ren WD. The reversal effect of Ginsenoside Rh2 on drug resistance in human colorectal carcinoma cells and its mechanism. Hum Cell. 2018; 31:189–98. https://doi.org/10.1007/s13577-017-0189-3 [PubMed]
- 20. Qiu R, Zhang J, Ge C, Zhong Y, Liu S, Li Q, Zou J, Fan H, Li Y. Ginsenosides Rg1 and CK Control Temozolomide Resistance in Glioblastoma Cells by Modulating Cholesterol Efflux and Lipid Raft Distribution. Evid Based Complement Alternat Med. 2022; 2022:1897508. https://doi.org/10.1155/2022/1897508 [PubMed]
- 21. Zhang J, Lu M, Zhou F, Sun H, Hao G, Wu X, Wang G. Key role of nuclear factor-κB in the cellular pharmacokinetics of adriamycin in MCF-7/Adr cells: the potential mechanism for synergy with 20(S)-ginsenoside Rh2. Drug Metab Dispos. 2012; 40:1900–8. https://doi.org/10.1124/dmd.112.045187 [PubMed]
- 22. Long J, Hu W, Ren T, Wang X, Lu C, Pan X, Wu C, Peng T. Combating multidrug resistance of breast cancer with ginsenoside Rh2-irrigated nano-in-thermogel. Int J Pharm. 2024; 650:123718. https://doi.org/10.1016/j.ijpharm.2023.123718 [PubMed]
- 23. Fan W, Fan L, Wang Z, Mei Y, Liu L, Li L, Yang L, Wang Z. Rare ginsenosides: A unique perspective of ginseng research. J Adv Res. 2024. [Epub ahead of print]. https://doi.org/10.1016/j.jare.2024.01.003 [PubMed]
- 24. Popov A, Klimovich A, Styshova O, Tsybulsky A, Hushpulian D, Osipyants A, Khristichenko A, Kazakov S, Ahuja M, Kaidery N, Thomas B, Tishkov V, Brown A, et al. Probable Mechanisms of Doxorubicin Antitumor Activity Enhancement by Ginsenoside Rh2. Molecules. 2022; 27:628. https://doi.org/10.3390/molecules27030628 [PubMed]
- 25. Hou JG, Jeon BM, Yun YJ, Cui CH, Kim SC. Ginsenoside Rh2 Ameliorates Doxorubicin-Induced Senescence Bystander Effect in Breast Carcinoma Cell MDA-MB-231 and Normal Epithelial Cell MCF-10A. Int J Mol Sci. 2019; 20:1244. https://doi.org/10.3390/ijms20051244 [PubMed]
- 26. Wu Q, Deng J, Fan D, Duan Z, Zhu C, Fu R, Wang S. Ginsenoside Rh4 induces apoptosis and autophagic cell death through activation of the ROS/JNK/p53 pathway in colorectal cancer cells. Biochem Pharmacol. 2018; 148:64–74. https://doi.org/10.1016/j.bcp.2017.12.004 [PubMed]
- 27. Chen J, Duan Z, Liu Y, Fu R, Zhu C. Ginsenoside Rh4 Suppresses Metastasis of Esophageal Cancer and Expression of c-Myc via Targeting the Wnt/β-Catenin Signaling Pathway. Nutrients. 2022; 14:3042. https://doi.org/10.3390/nu14153042 [PubMed]
- 28. Kciuk M, Gielecińska A, Mujwar S, Kołat D, Kałuzińska-Kołat Ż, Celik I, Kontek R. Doxorubicin-An Agent with Multiple Mechanisms of Anticancer Activity. Cells. 2023; 12:659. https://doi.org/10.3390/cells12040659 [PubMed]
- 29. Housman G, Byler S, Heerboth S, Lapinska K, Longacre M, Snyder N, Sarkar S. Drug resistance in cancer: an overview. Cancers (Basel). 2014; 6:1769–92. https://doi.org/10.3390/cancers6031769 [PubMed]
- 30. Li Y, Tan M, Sun S, Stea E, Pang B. Targeted CRISPR activation and knockout screenings identify novel doxorubicin transporters. Cell Oncol (Dordr). 2023; 46:1807–20. https://doi.org/10.1007/s13402-023-00847-0 [PubMed]
- 31. Skinner KT, Palkar AM, Hong AL. Genetics of ABCB1 in Cancer. Cancers (Basel). 2023; 15:4236. https://doi.org/10.3390/cancers15174236 [PubMed]
- 32. Al-Malky HS, Al Harthi SE, Osman AM. Major obstacles to doxorubicin therapy: Cardiotoxicity and drug resistance. J Oncol Pharm Pract. 2020; 26:434–44. https://doi.org/10.1177/1078155219877931 [PubMed]
- 33. Hu J, Jiang Y, Tan S, Zhu K, Cai T, Zhan T, He S, Chen F, Zhang C. Selenium-doped calcium phosphate biomineral reverses multidrug resistance to enhance bone tumor chemotherapy. Nanomedicine. 2021; 32:102322. https://doi.org/10.1016/j.nano.2020.102322 [PubMed]
- 34. Kong QW, Yang J, Li D, Ding YW, Hu YJ, Xue XC, Shi MZ, Jiang B, Zhou YY, Zhang M, Hu JD, Guo C, Chen JJ, Han YL. Tongguanteng injection reverses paclitaxel resistance via upregulation of TAB1 expression in ovarian cancer in vitro and in vivo. J Ethnopharmacol. 2023; 300:115728. https://doi.org/10.1016/j.jep.2022.115728 [PubMed]
- 35. Sajid A, Rahman H, Ambudkar SV. Advances in the structure, mechanism and targeting of chemoresistance-linked ABC transporters. Nat Rev Cancer. 2023; 23:762–79. https://doi.org/10.1038/s41568-023-00612-3 [PubMed]
- 36. Bu D, Luo H, Huo P, Wang Z, Zhang S, He Z, Wu Y, Zhao L, Liu J, Guo J, Fang S, Cao W, Yi L, et al. KOBAS-i: intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021; 49:W317–25. https://doi.org/10.1093/nar/gkab447 [PubMed]
- 37. Kang S, Denley A, Vanhaesebroeck B, Vogt PK. Oncogenic transformation induced by the p110beta, -gamma, and -delta isoforms of class I phosphoinositide 3-kinase. Proc Natl Acad Sci USA. 2006; 103:1289–94. https://doi.org/10.1073/pnas.0510772103 [PubMed]
- 38. Zhang L, Li Y, Wang Q, Chen Z, Li X, Wu Z, Hu C, Liao D, Zhang W, Chen ZS. The PI3K subunits, P110α and P110β are potential targets for overcoming P-gp and BCRP-mediated MDR in cancer. Mol Cancer. 2020; 19:10. https://doi.org/10.1186/s12943-019-1112-1 [PubMed]
- 39. PDQ Pediatric Treatment Editorial Board. Childhood Soft Tissue Sarcoma Treatment (PDQ®): Health Professional Version. 2024. In: PDQ Cancer Information Summaries [Internet]. Bethesda (MD): National Cancer Institute (US). 2002. [PubMed]
- 40. Thebault E, Piperno-Neumann S, Tran D, Pacquement H, Marec-Berard P, Lervat C, Castex MP, Cleirec M, Bompas E, Vannier JP, Plantaz D, Saumet L, Verite C, et al. Successive Osteosarcoma Relapses after the First Line O2006/Sarcome-09 Trial: What Can We Learn for Further Phase-II Trials? Cancers (Basel). 2021; 13:1683. https://doi.org/10.3390/cancers13071683 [PubMed]
- 41. Bielack SS, Smeland S, Whelan JS, Marina N, Jovic G, Hook JM, Krailo MD, Gebhardt M, Pápai Z, Meyer J, Nadel H, Randall RL, Deffenbaugh C, et al, and EURAMOS-1 investigators. Methotrexate, Doxorubicin, and Cisplatin (MAP) Plus Maintenance Pegylated Interferon Alfa-2b Versus MAP Alone in Patients With Resectable High-Grade Osteosarcoma and Good Histologic Response to Preoperative MAP: First Results of the EURAMOS-1 Good Response Randomized Controlled Trial. J Clin Oncol. 2015; 33:2279–87. https://doi.org/10.1200/JCO.2014.60.0734 [PubMed]
- 42. Ganta S, Amiji M. Coadministration of Paclitaxel and curcumin in nanoemulsion formulations to overcome multidrug resistance in tumor cells. Mol Pharm. 2009; 6:928–39. https://doi.org/10.1021/mp800240j [PubMed]
- 43. Ghafouri-Fard S, Shabestari FA, Vaezi S, Abak A, Shoorei H, Karimi A, Taheri M, Basiri A. Emerging impact of quercetin in the treatment of prostate cancer. Biomed Pharmacother. 2021; 138:111548. https://doi.org/10.1016/j.biopha.2021.111548 [PubMed]
- 44. Li QF, Shi SL, Liu QR, Tang J, Song J, Liang Y. Anticancer effects of ginsenoside Rg1, cinnamic acid, and tanshinone IIA in osteosarcoma MG-63 cells: nuclear matrix downregulation and cytoplasmic trafficking of nucleophosmin. Int J Biochem Cell Biol. 2008; 40:1918–29. https://doi.org/10.1016/j.biocel.2008.01.031 [PubMed]
- 45. Ren Z, Yang H, Zhu C, Deng J, Fan D. Ginsenoside Rh4 Alleviates Amyloid β Plaque and Tau Hyperphosphorylation by Regulating Neuroinflammation and the Glycogen Synthase Kinase 3β Signaling Pathway. J Agric Food Chem. 2023; 71:13783–94. https://doi.org/10.1021/acs.jafc.3c02550 [PubMed]
- 46. Zhang Y, Ma P, Duan Z, Liu Y, Mi Y, Fan D. Ginsenoside Rh4 Suppressed Metastasis of Lung Adenocarcinoma via Inhibiting JAK2/STAT3 Signaling. Int J Mol Sci. 2022; 23:2018. https://doi.org/10.3390/ijms23042018 [PubMed]
- 47. Wang Y, An X, Wang F, Jiang Y. Ginsenoside RH4 inhibits Ang II-induced myocardial remodeling by interfering with NFIL3. Biomed Pharmacother. 2024; 172:116253. https://doi.org/10.1016/j.biopha.2024.116253 [PubMed]
- 48. Marchandet L, Lallier M, Charrier C, Baud’huin M, Ory B, Lamoureux F. Mechanisms of Resistance to Conventional Therapies for Osteosarcoma. Cancers (Basel). 2021; 13:683. https://doi.org/10.3390/cancers13040683 [PubMed]
- 49. Zou Z, Zhang J, Zhang H, Liu H, Li Z, Cheng D, Chen J, Liu L, Ni M, Zhang Y, Yao J, Zhou J, Fu J, Liang Y. 3-Methyladenine can depress drug efflux transporters via blocking the PI3K-AKT-mTOR pathway thus sensitizing MDR cancer to chemotherapy. J Drug Target. 2014; 22:839–48. https://doi.org/10.3109/1061186X.2014.936870 [PubMed]
- 50. Wu DP, Zhou Y, Hou LX, Zhu XX, Yi W, Yang SM, Lin TY, Huang JL, Zhang B, Yin XX. Erratum: Cx43 deficiency confers EMT-mediated tamoxifen resistance to breast cancer via c-Src/PI3K/Akt pathway: Erratum. Int J Biol Sci. 2022; 18:5309–11. https://doi.org/10.7150/ijbs.75891 [PubMed]
- 51. Qiu C, Su W, Shen N, Qi X, Wu X, Wang K, Li L, Guo Z, Tao H, Wang G, Chen B, Xiang H. MNAT1 promotes proliferation and the chemo-resistance of osteosarcoma cell to cisplatin through regulating PI3K/Akt/mTOR pathway. BMC Cancer. 2020; 20:1187. https://doi.org/10.1186/s12885-020-07687-3 [PubMed]
- 52. Lu X, Yang F, Chen D, Zhao Q, Chen D, Ping H, Xing N. Quercetin reverses docetaxel resistance in prostate cancer via androgen receptor and PI3K/Akt signaling pathways. Int J Biol Sci. 2020; 16:1121–34. https://doi.org/10.7150/ijbs.41686 [PubMed]
- 53. Kalinina EV, Hasan AAS, Tatarskiy VV, Volodina YL, Petrova AS, Novichkova MD, Zhdanov DD, Nurmuradov NK, Chernov NN, Shtil AA. Suppression of PI3K/Akt/mTOR Signaling Pathway and Antioxidant System and Reversal of Cancer Cells Resistance to Cisplatin under the Effect of Curcumin. Bull Exp Biol Med. 2022; 173:371–5. https://doi.org/10.1007/s10517-022-05551-x [PubMed]
- 54. Munoz J, Follows GA, Nastoupil LJ. Copanlisib for the Treatment of Malignant Lymphoma: Clinical Experience and Future Perspectives. Target Oncol. 2021; 16:295–308. https://doi.org/10.1007/s11523-021-00802-9 [PubMed]
- 55. André F, Ciruelos E, Rubovszky G, Campone M, Loibl S, Rugo HS, Iwata H, Conte P, Mayer IA, Kaufman B, Yamashita T, Lu YS, Inoue K, et al, and SOLAR-1 Study Group. Alpelisib for PIK3CA-Mutated, Hormone Receptor-Positive Advanced Breast Cancer. N Engl J Med. 2019; 380:1929–40. https://doi.org/10.1056/NEJMoa1813904 [PubMed]
- 56. Shu J, Dolman GE, Duan J, Qiu G, Ilyas M. Statistical colour models: an automated digital image analysis method for quantification of histological biomarkers. Biomed Eng Online. 2016; 15:46. https://doi.org/10.1186/s12938-016-0161-6 [PubMed]