



Introduction
Breast cancer (BC) is the most common cancer in women worldwide and the leading cause of cancer death in women. The ER-positive (ER+) breast cancer is the most common subtype of breast cancer, accounting for 68% of all breast cancer types. Because of its heterogeneous nature, it is particularly difficult to diagnose and evaluate clinically, and therefore the incidence and mortality rates are on the rise and the disease burden is increasing [1]. BC patients may experience bone metastasis (BM), which is currently considered incurable [2]. About 70% or more of advanced breast cancer will develop distant metastases, and about 83% of them have bone as the first metastatic site. Once bone metastases occur, osteolytic destruction is formed, which is prone to fracture and nerve compression in late stage, and obvious pain and paralysis symptoms, which seriously affects patients’ survival quality and survival rate [3]. According to the World Health Organization’s International Agency for Research on Cancer (IARC) 2022 cancer statistics report, breast cancer accounts for the first place among all cancers in women with 31% of new cases and the second place among all cancer types in women with 15% of mortality. However, because the molecular mechanisms of BM have not been fully elucidated, the efficacy of existing treatments is limited and they have failed to significantly improve the overall survival rate of patients [1, 4, 5].
BM is a complex multi-step process, including the disruption of the dynamic balance of bone remodeling, vascularization in the tumor, regulation of immune cells such as bone marrow mesenchymal stem cells (BMSCs), adipocytes and macrophages, etc. The molecular mechanisms are very complex, and these cell types and their secreted factors together constitute the tumor microenvironment (TME). These cell types and their secreted factors together constitute the TME, and are closely related to the occurrence of breast cancer bone metastasis [6, 7]. The complex composition of the TME includes fibroblasts, immune cells, adipocytes, vascular endothelial cells and extracellular matrix, etc. The complex molecular components and cellular changes in the TME are essential for promoting cancer metastasis [8]. Previous studies have shown that relevant fibroblasts can promote the metastasis of breast cancer cells. These fibroblasts play a key role in the bone colonization of breast cancer cells by influencing the intrinsic tumor characteristics and TME [9]. In addition, MSC localized in breast cancer form “tumor xenografts” with tumor cells, leading to tumor cell growth and bone metastasis [10]. In addition, patients with breast cancer may induce oxidative reactions in the body through a variety of pathways, placing the body in a state of oxidative stress and adversely affecting the prognosis of the patient [11]. The ability of breast cancer to metastasize is closely related to the redox status of cells [12]. However, the ecology of specific cells in BM is still unknown to a large extent. Therefore, further research on the cell level in BM is of great scientific significance and is of great significance for developing new strategies for BM treatment.
In this study, we used single cell technology to construct a global single cell landscape atlas of BM, comprehensively discussed the ecosystem of BM microenvironment, and revealed the imbalance of BM bone remodeling balance and the important role of some cells in BM development.
Materials and Methods
Data collection and processing
BM-related single cell RNA sequencing (scRNA-seq) data were obtained from Gene Expression Omnibus (GEO, https://www.ncbi.nlm.nih.gov/geo/), dataset GSE190772 based on the GPL24676 platform, including BM of two bilateral bone metastases collected in a patient initially diagnosed with ER+ primary breast cancer. In addition, dataset GSE131007 based on the GPL20301 and GPL24676 platforms, including three primary tumor tissues from mouse xenografts of human BC patient origin and one BM tissue. Among them, cells from murine cells or human-murine doublets were excluded. Total 3 BM tissues and 3 primary samples, and the primary samples were the control samples in this study.
Construction of single cell atlas
Single-cell data were merged using the IntegrateData function [13] of the Seurat package [14] in R language, and cell clustering analysis was performed according to default parameters, filtering for cells with top and bottom 1% gene count and >10% mitochondrial content. The clustering results were downscaled and visualized [15] based on a uniform manifold approximation and projection (UMAP) for dimension reduction technique and projected onto a two-dimensional image defined as a single-cell atlas. In addition, cell types were annotated according to cell markers known from previous studies [16].
Differential gene expression analysis
The differentially expressed genes (DEGs) in each cluster between single cells of primary tumor tissue and BM tissues were identified using the “FindAllMarkers” function, and differences with a adjusted P-values < 0.05 and |log fold change (logFC)| > 0.5 were considered significant.
Functional enrichment and gene enrichment analysis
To further explore the biological processes and pathways involved with genes that showed dysregulated expression in different cell clusters, Gene Ontology (GO) terminology and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed based on the expression of marker genes. The R package clusterProfiler [17] for enrichment analysis regarding biological processes (BPs) of GO and KEGG signaling pathways, and P<0.05 were considered significant.
Pseudo-time analysis
The differentiation developmental trajectory of dysregulated cells in primary tumor tissues and BM tissues was reconstructed using the Monocle 3 package [19] in R language and visualized by UMAP. Subsequently, the cells were sorted according to their progression through the developmental program.
Gene regulatory network (GRN) analysis
In addition, using the Python module tool pySCENIC [20], this study comprehensively reconstructed the transcription factor-centered gene regulatory network to further explore the regulatory mechanisms of dysregulated cells.
The workflow started with describing the input single-cell expression level profile matrix, and then using a regression method for each target (GRNBoost2) to infer co-expression modules. The results allowed us to determine which indirect targets were trimmed based on the discovery of cis-regulatory patterns (cisTarget). Subsequently, AUcell was used to quantify the activity of those regulators by enriching and scoring the regulator target genes to obtain a regulon activity score (RAS). The single-cell data were further downscaled using the RAS matrix and a regulon specificity score (RSS) was calculated based on the Jensen-Shannon divergence (JS scatter) and used to identify regulators specific for certain cell populations. The most specific and significant regulons were mapped to single cell cluster profiles and validated using massively parallel sample sequencing (SEEK database). Finally, a connection specificity index (CSI) matrix was calculated, and the regulators were hierarchically clustered according to CSI to define regulator modules that could be used to identify relationships between regulator modules and regulators. Those relationships were then visualized using the R package ComplexHeatmap.
Cellular communication
Signal transduction emphasizes the manner and outcome of signal reception and the signal conversion after reception, with ligand-receptor binding being one of the main forms of signal transduction between neighboring cells. In this study, high confidence ligand-receptor interactions between subpopulations of cells were identified by the R language package iTALK. It preferentially identifies genes that are highly or differentially expressed in cell clusters that will be matched by a ligand-receptor database to discover important intercellular communication events.
Data analysis and statistics
Comparisons between the two groups were made using Student’s t test and correlation coefficients were calculated using Spearman analysis. P<0.05 was considered significant. Regarding the code used in this study we have uploaded it as Supplementary File 1.
Data availability statement
Data used in this study were obtained from Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo).
Results
Global single-cell landscape of bone metastases from breast cancer
We try to draw the global single cell atlas of BM through scRNA-seq technology, and further explore the potential ecological panorama of BM, in order to find the potential therapeutic target of BM. The flow of this study was shown in Figure 1A. After standardized data processing and quality control, a total of 16,409 high-quality single-cell transcriptional profiles were captured and clustered to generate 43 cell clusters, and differential gene expression analysis revealed a wide range of gene expression dysregulation in different kinds of cell types in BM compared to controls. The cell clusters were further identified into eight cell types (Figure 1B), including BC cells, osteoblasts (OC), mesenchymal stem cells (MSCs), B cells, CD8+ T cells, CD4+ T cells, fibroblasts and endothelial cells (En). The markers positively expressed by the cells are consistent with recent published gene signatures such as scRNA-seq and laboratory studies (Figure 1C). In addition, BC cells were reduced in the BM microenvironment compared to controls, whereas MSCs, Fibroblasts and other cells were significantly more abundant in the BM (Figure 1D). In addition, we assessed the differences in single-cell oxidative stress levels between control and BM samples and found that GO_CELL_DEATH_IN_RESPONSE_TO_OXIDATIVE_STRESS and GO_RESPONSE_TO_OXIDATIVE_STRESS had higher scores in bone metastasis samples (Figure 1E). In summary, we initially constructed a global landscape of the dynamic single-cell ecology of the BM microenvironment by single-cell histology, and we found significant concomitant dysregulation of gene expression between different cell types and explored the altered cellular ecology of BM patients, in which BC, MSCs and fibroblasts may play an important role in BM. In addition to this, oxidative stress may play a key role in BM.
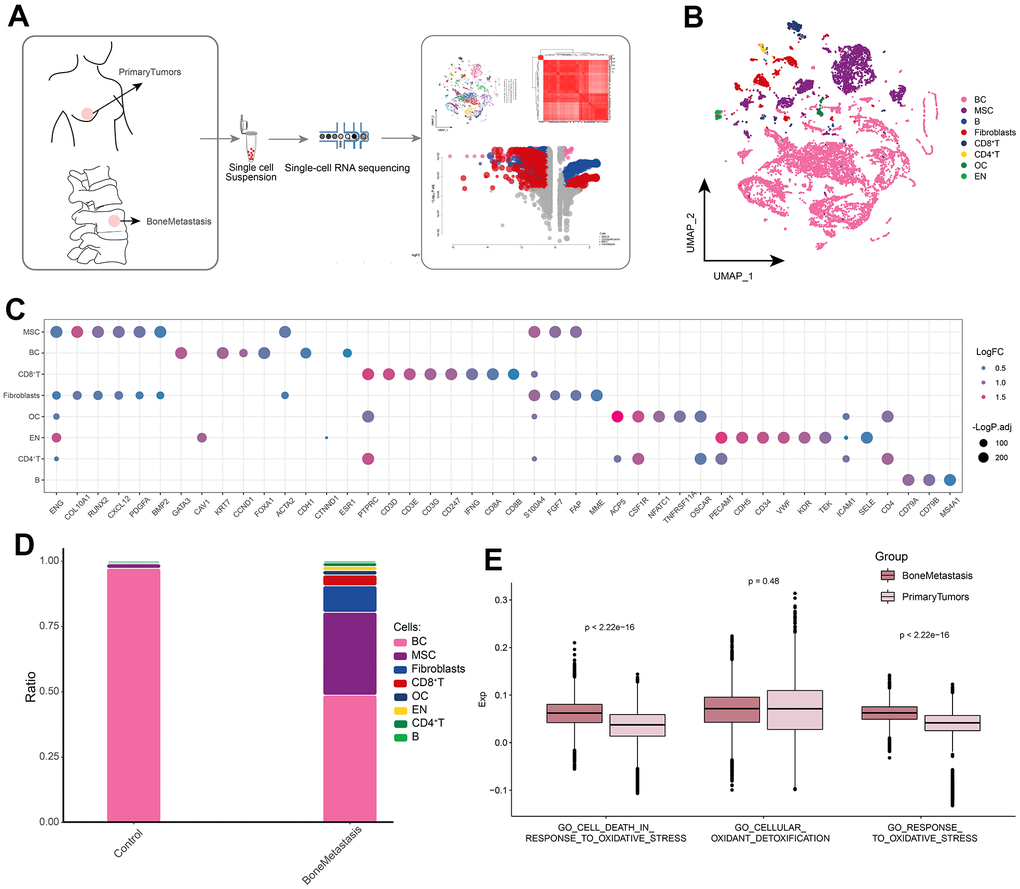
Figure 1. Global single-cell landscape of patients with bone metastases from breast cancer. (A) Flow chart underlying this study. Primary tumors were used as controls in the follow-up study. (B) Single-cell atlas mapping cell types. (C) Cell marker genes for annotation. (D) Differences in cell abundance between control and breast cancer bone metastasis patients. (E) Comparison of differences in single-cell oxidative stress levels between control and bone metastasis samples.
Landscape of BC cell subpopulations in breast cancer bone metastases
When cancer cells metastasize to the bone, these cells enter some part of the bone through the blood or lymphatic system and become metastatic cancer cells. When these cancer cells deposit in bone, they release substances that form osteoclasts and osteoblasts. This may account for the decrease in BC cells in Figure 1D. Based on the cellular ecological atlas at single-cell resolution, we explored the subpopulations of BC cells in depth and identified a total of 10 subpopulations of BC cells (Figure 2A) and found that these subpopulations were heterogeneous among different subgroups (Figure 2B). These BC cell subpopulations all significantly expressed their marker genes, with BC_MUC1, BC_SCGB2A2, BC_FN1, BC_BGN, and BC_PEG10 subpopulations being significantly more abundant in BM (Figure 2C, 2D). By enrichment analysis, we found that these subpopulations were significantly enriched in oxidative stress response and BPs associated with oxidative stress and cell death (Figure 2E). In addition, Oxidative phosphorylation, HIF-1 signaling pathway, TGF-beta signaling pathway, Wnt signaling pathway, Apoptosis-associated oxidative stress and cell death-related pathways, MAPK signaling pathway, p53 signaling pathway, PI3K-Akt signaling pathway Breast cancer, and JAK-STAT signaling pathway of cancer-related signaling pathways were also significantly enriched (Figure 2F). Meanwhile, in control and BM samples, BC cells were significantly different from GO_CELL_DEATH_IN_RESPONSE_TO_OXIDATIVE_STRESS, GO_CELLULAR_OXIDANT_DETOXIFICATION and GO_RESPONSE_TO_OXIDATIVE_ STRESS oxidative stress pathway scores were significantly different (Figure 2G and Supplementary Figure 1). Oxidative stress-related pathway scores were significantly higher in BM samples than in controls. These results further suggest that BC cells in BM are susceptible to oxidative damage and exhibit high levels of oxidative stress, which plays a key role in apoptosis.
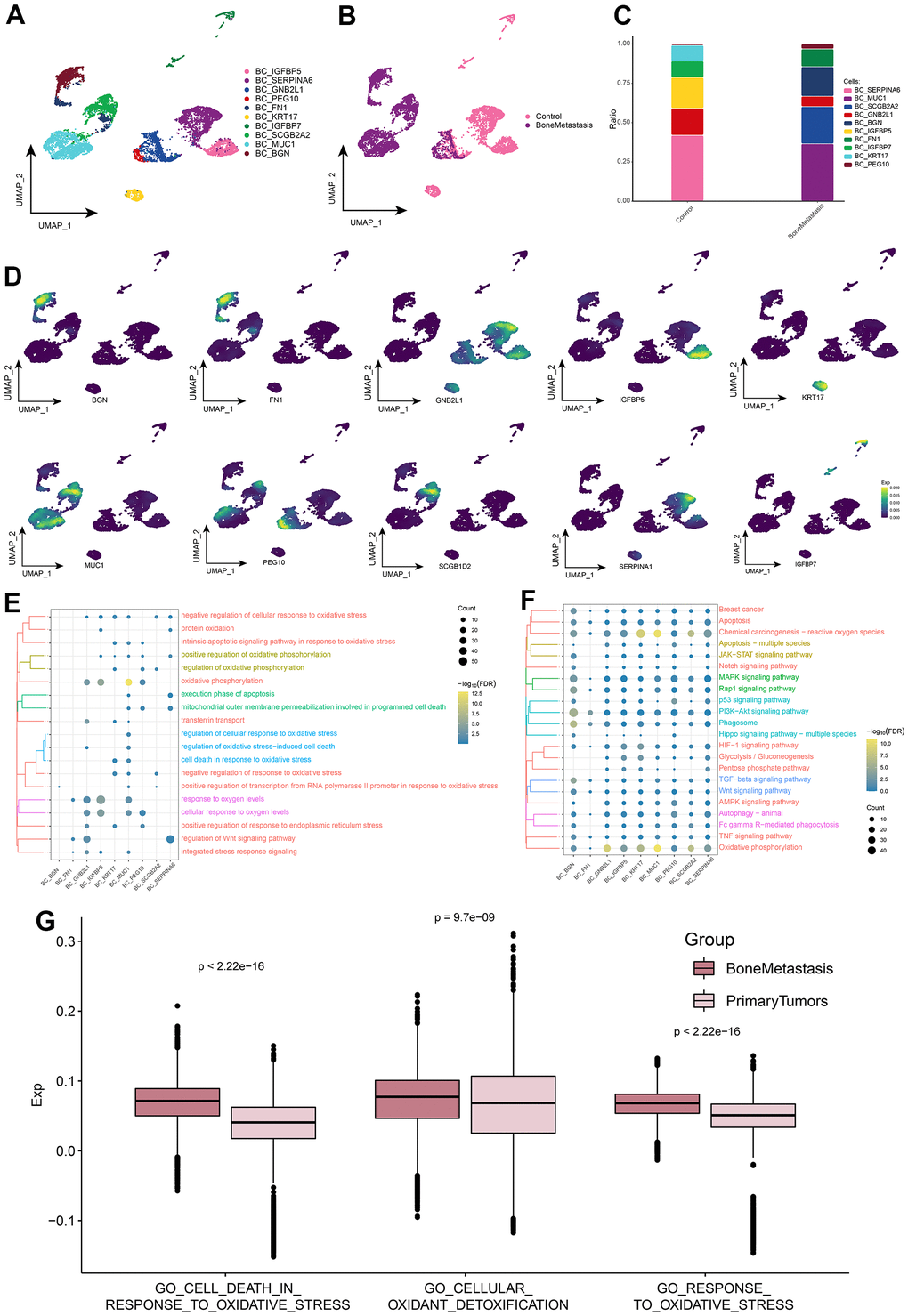
Figure 2. Breast cancer cell subpopulations in patients with bone metastases from breast cancer. (A) Single-cell atlas showing breast cancer cell subpopulations. (B) Single-cell atlas showing breast cancer cell subpopulations in control and breast cancer bone metastasis patients. (C) Differential abundance of breast cancer cell subpopulations in control and breast cancer bone metastasis patients. (D) Marker genes specifically and highly expressed in subpopulations of breast cancer cells. (E, F) Biological processes (E) and signaling pathways (F) enriched in breast cancer cell subpopulations. (G) Comparison of the differences in oxidative stress levels in breast cancer cells between control and bone metastasis samples.
Clonal evolution of BC cells in bone metastases from breast cancer
We then depicted the differentiated developmental trajectories of tumor cell subpopulations. BC_MUC1, BC_SCGB2A2, and BC_BGN subpopulations were at the end of the developmental trajectory of tumor cells, while BC_SERPIINA6 subpopulation was at the beginning of the developmental trajectory and gradually differentiated into different other subpopulations as BM progressed (Figure 3A, 3B). Markers were clustered into four modules, where BC_MUC1 and BC_SERPIINA6 were regulated by the same transcription factor HOXB2 and BC_BGN was regulated by the TCF4 transcription factor (Figure 3C), and the expression of these transcription factors was mapped in a single cell atlas (Figure 3D). In conclusion, we further clarified the differentiation and development trajectory of tumor cell subsets in BM and the transcriptional regulation targets of different subsets.
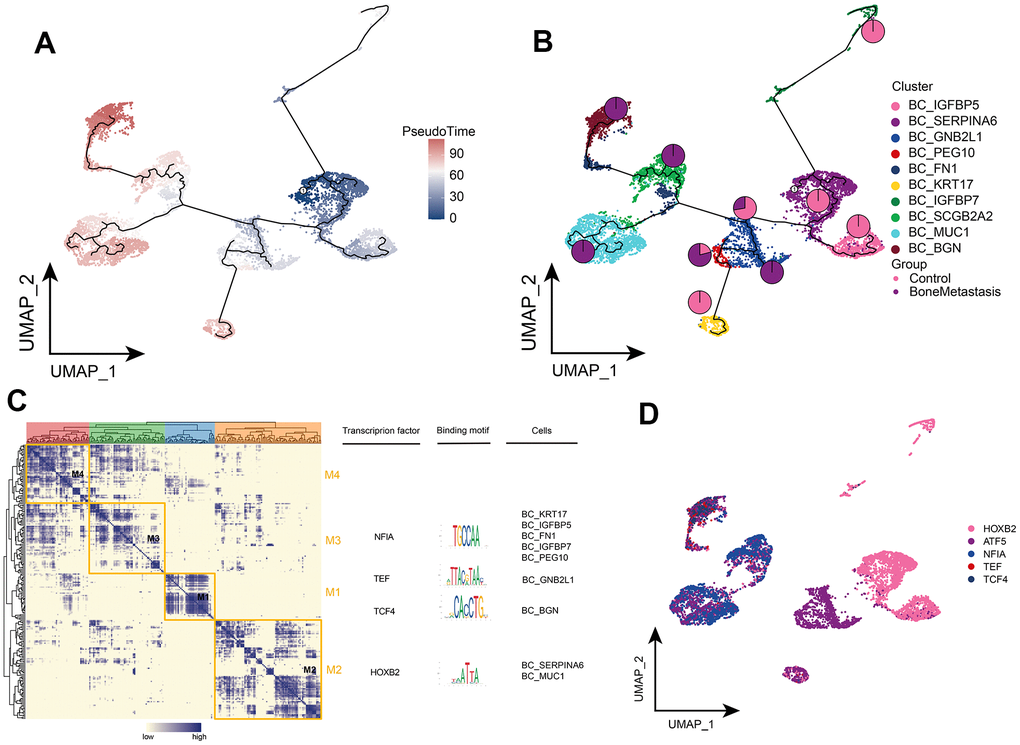
Figure 3. Clonal evolution of breast cancer cell subpopulations in patients with bone metastases from breast cancer. (A, B) Pseudo-time values (A) and developmental trajectories (B) of breast cancer cell subpopulations, with pie charts representing the proportion of control and breast cancer bone metastasis patients in breast cancer cell subpopulations. (C) Co-expression modules of transcription factors in breast cancer cell subpopulations of patients with breast cancer bone metastases. Left: Identification of regulator modules based on the regulator’s linkage specificity index matrix. Middle: representative transcription factors and their binding patterns in the modules. Right panel: cell subpopulations in which transcription factors are located. (D) Single-cell atlas showing transcription factors regulating breast cancer cell subpopulations.
Landscape of fibroblast subpopulations in bone metastases from breast cancer
The aforementioned of others’ analysis revealed the presence of a large number of fibroblasts in bone metastasis sites, and fibroblasts are becoming important cellular players in bone metastasis [21]. Eight fibroblast subpopulations were further identified by subpopulation analysis (Figure 4A), and these fibroblast subpopulations were essentially highly enriched in BM (Figure 4B), with differentially expressed genes for each subpopulation displayed in Supplementary Table 1. These fibroblast subpopulations all expressed different specific markers, and studies of their variable levels revealed significant abundance of Fibroblasts_CLDN10 and Fibroblasts_S100P in controls, and Fibroblasts_IBSP, Fibroblasts_TAGLN, Fibroblasts_ASPN, and Fibroblasts_OLFML2B, Fibroblasts_KRT19 and Fibroblasts_MGST1 were significantly abundant in BM (Figure 4C, 4D). Further, the BPs and KEGG signaling pathways involved in these fibroblast subpopulations were explored, and fibroblast subpopulations were found to be significantly involved in the biological processes of ossification and bone remodeling (Figure 4E), in addition to ECM-receptor interaction, Cytokine-cytokine receptor interaction, Apoptosis, Focal adhesion, TNF signaling pathway, and TGF-beta signaling pathway were significantly enriched (Figure 4F). Taken together, our results suggest that fibroblasts in BM may regulate the ecological niche formation of bone metastases and play an important role for tumor cell inoculation into the bone marrow and growth.
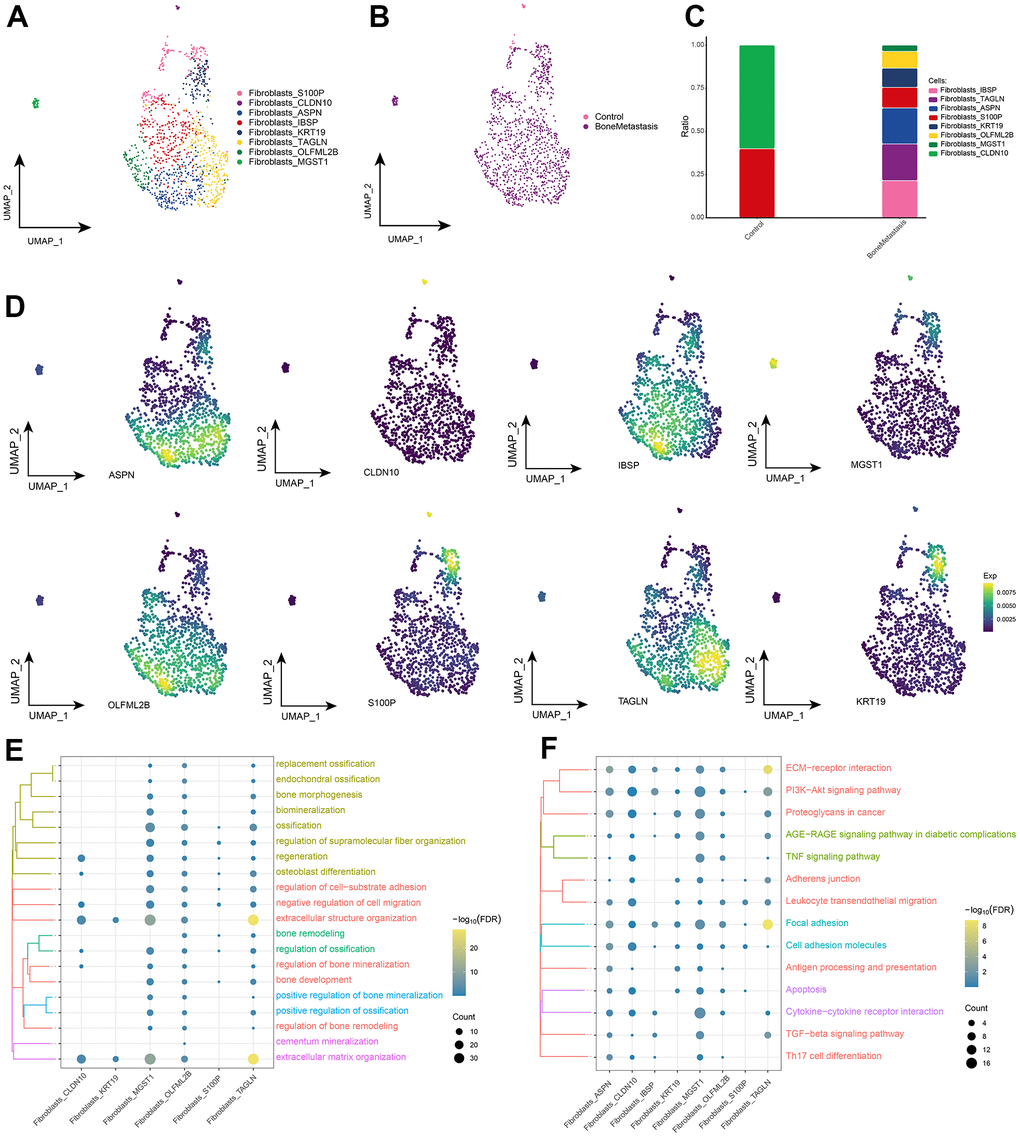
Figure 4. Fibroblast subpopulations in patients with bone metastases from breast cancer. (A) Single-cell atlas showing fibroblast subpopulations. (B) Single-cell atlas showing fibroblast subpopulations in control and breast cancer bone metastasis patients. (C) Differences in abundance of fibroblast subpopulations in control and breast cancer bone metastasis patients. (D) Marker genes specifically and highly expressed in subpopulations of fibroblast subpopulations. (E, F). Biological processes (E) and signaling pathways (F) that enrich fibroblast subpopulations.
Clonal evolution of fibroblasts in bone metastases from breast cancer
The pseudo-time differentiation trajectory showed that the Fibroblasts_S100P and Fibroblasts_CLDN10 subpopulations were in an early position of differentiation development, while Fibroblasts_IBSP, Fibroblasts_TAGLN, Fibroblasts_ASPN, Fibroblasts_ OLFML2B, Fibroblasts_KRT19, and Fibroblasts_MGST1 were at the end stage and all were highly enriched in BM (Figure 5A, 5B). Subsequent GRN analysis showed that fibroblast subpopulation genes were organized into three modules (Figure 5C) and that different fibroblast subpopulations were regulated by different TFs to guide cell fate selection, respectively (Figure 5D). These results reflect the developmental trajectory of fibroblast subpopulations in BM and the transcriptional regulatory targets of different subpopulations.
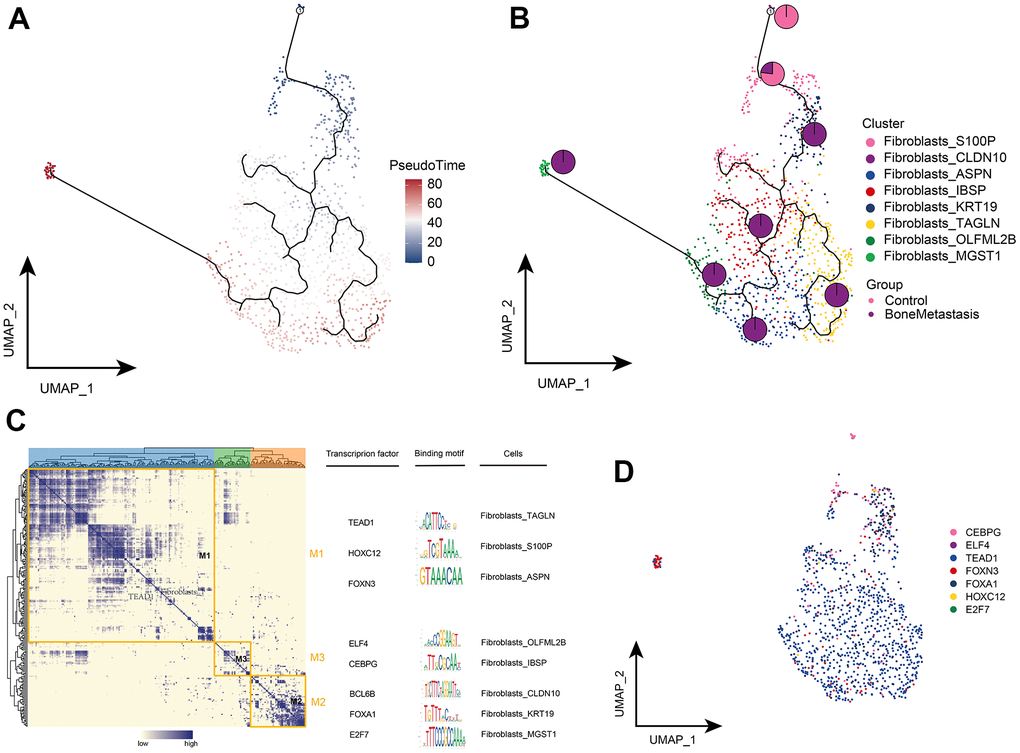
Figure 5. Clonal evolution of fibroblast subpopulations in patients with bone metastases from breast cancer. (A, B) Pseudo-time values (A) and developmental trajectories (B) of fibroblast subpopulations, with pie charts representing the proportion of fibroblast subpopulations in control and breast cancer bone metastasis patients. (C) Co-expression modules of transcription factors in fibroblast subpopulations of patients with breast cancer bone metastases. Left: Identification of regulator modules based on the regulator’s linkage specificity index matrix. Middle: representative transcription factors and their binding patterns in the modules. Right panel: cell subpopulations in which transcription factors are located. (D) Single-cell atlas showing transcription factors regulating fibroblast subpopulations.
The MSC subpopulations landscape of breast cancer bone metastases
MSCs can differentiate into a variety of cell types, including osteogenic osteoblasts, chondrocytes, and adipocytes [22]. In addition, MSCs are a key component of tumor cell homing and adhesion to bone metastasis ecological niches [2]. Exploring the MSC subpopulation landscape by single cell resolution identified ten MSC subpopulations (Figure 6A), all of which were essentially significantly abundant in BM (Figure 6B), with differentially expressed genes for each subpopulation displayed in Supplementary Table 2. Further exploring the expression of specific markers for these subpopulations and differences in the abundance of subpopulations, the MSC_MARCKSL1 subpopulation was significantly more abundant in the control group, while all other subpopulations were significantly present in the BM group (Figure 6C, 6D). Enrichment analysis revealed that MSC subpopulations were involved in the regulation of BMP signaling pathway, tumor necrosis factor production, tumor necrosis factor superfamily cytokine production, and some skeletal growth and development and oxidative stress-related BPs (Figure 6E), Focal adhesion, Cell adhesion molecules, ECM-receptor interaction, which are KEGG signaling pathways associated with tumor metastasis, were enriched, in addition, PI3K-Akt signaling pathway, TGF-beta signaling pathway, Hippo signaling pathway and NF-kappa B signaling pathway, which are cancer-related KEGG signaling pathways, were also significantly enriched (Figure 6F). GO_RESPONSE_TO_OXIDATIVE_ STRESS scores were higher in the BM samples (Figure 6G and Supplementary Figure 2). In conclusion, by identifying MSC subpopulations and uncovering the functional roles of their subpopulations, we found that MSCs and oxidative stress may play an important role in BM.
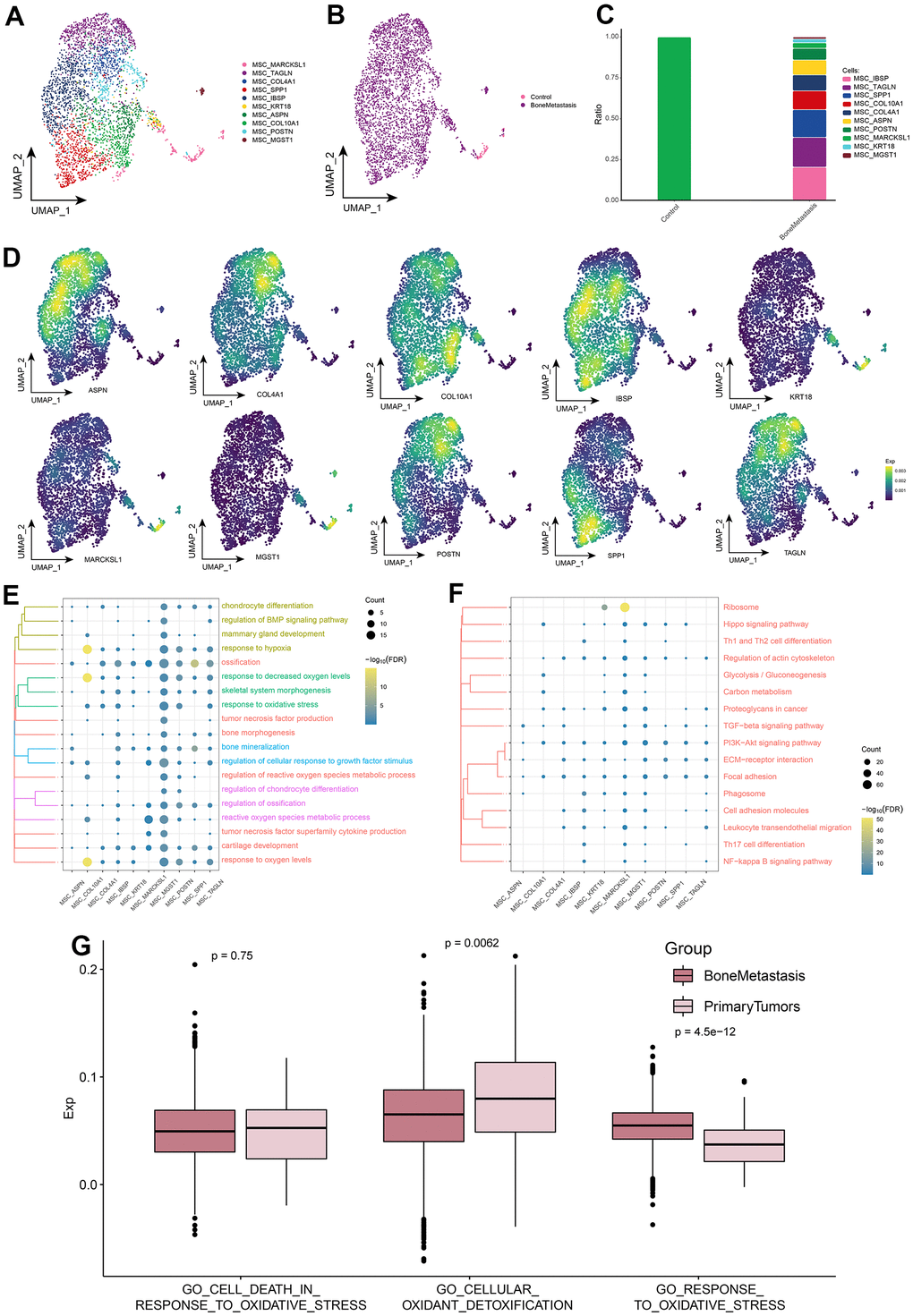
Figure 6. MSC subpopulations in patients with bone metastases from breast cancer. (A) Single-cell atlas showing MSC subpopulations. (B) Single-cell atlas showing MSC subpopulations in control and breast cancer bone metastasis patients. (C) Differential abundance of MSC subpopulations in control and breast cancer bone metastasis patients. (D) Marker genes specifically and highly expressed in subpopulations of MSC subpopulations. (E, F) Biological processes (E) and signaling pathways (F) enriched in MSC subpopulations. (G) Comparison of the differences in oxidative stress levels in MSC between control and bone metastasis samples.
Clonal evolution of MSCs with bone metastases from breast cancer
Further exploring the differentiation trajectory of the MSC subpopulations, the MSC_MARCKSL1 subpopulation was located at an early position in the development of differentiation and had a high ability to differentiate further toward subpopulations specifically present in the BM (Figure 7A, 7B). Confirming the accuracy of the results in Figure 6C, the MSC_MARCKSL1 subpopulation was significantly more abundant in the primary lesions. By GRN, we explored the TFs regulating MSC subpopulations that are regulated by different TFs, including HOXB3, PPARG, SPI1, XBP1, ELF5, and ZBTB7B, respectively (Figure 7C, 7D). Taken together, we can know the differentiated developmental trajectory of MSC subpopulations and explored their transcriptional regulation.
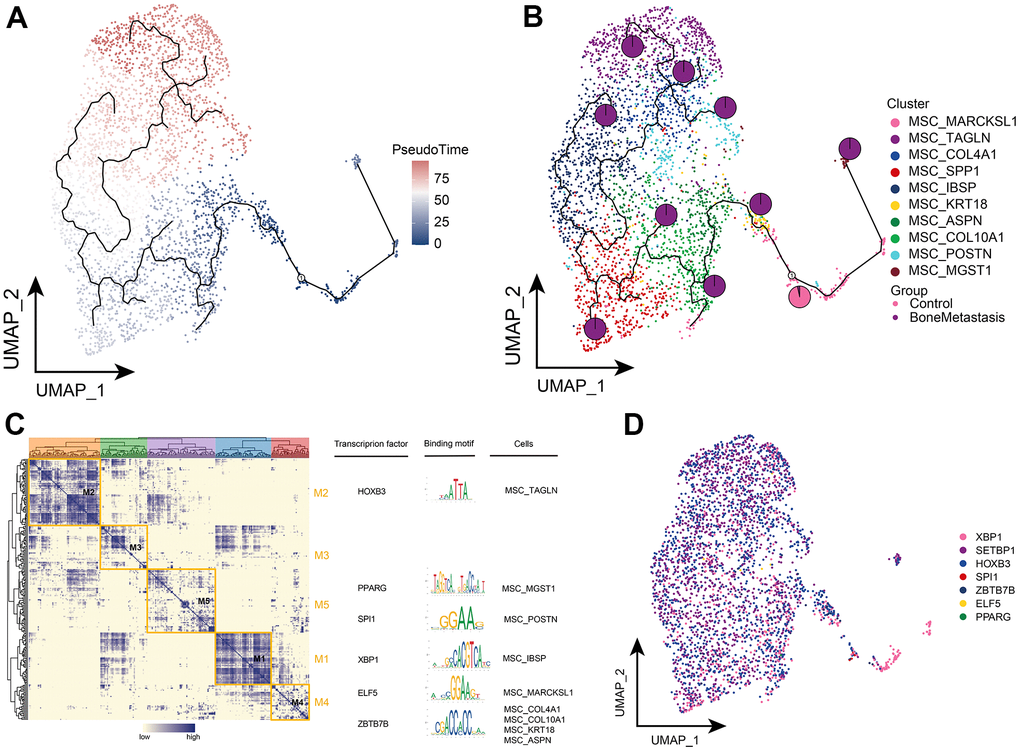
Figure 7. Clonal evolution of MSC subpopulations in patients with bone metastases from breast cancer. (A, B) Pseudo-time values (A) and developmental trajectories (B) of MSC subpopulations, pie charts representing the proportion of control and breast cancer bone metastasis patients in MSC subpopulations. (C) Co-expression modules of transcription factors in MSC subpopulations of patients with breast cancer bone metastases. Left: Identification of regulator modules based on the regulator’s linkage specificity index matrix. Middle: representative transcription factors and their binding patterns in the modules. Right panel: cellular subpopulations in which transcription factors are located. (D) Single-cell atlas showing transcription factors regulating MSC subpopulations.
Intercellular communication in breast cancer bone metastases
Since we have successfully outlined cellular-level alterations in BM and characterized the functional and transcriptional profiles of different cellular subpopulations, we used a public ligand-receptor database to infer intercellular communication during BM. By comparing cell identity-specific genes with ligand receptors, we classified hypothetical ligand-receptor pairs for different cell populations in control and BM samples. For controls, we found a stronger interaction between fibroblasts and BC cells (Figure 8A). Notably, in BM, BC cells showed the most interactions with other cell types (Figure 8B and Supplementary Table 3), and we observed strong ligand-receptor pairs in BC cell subpopulations.
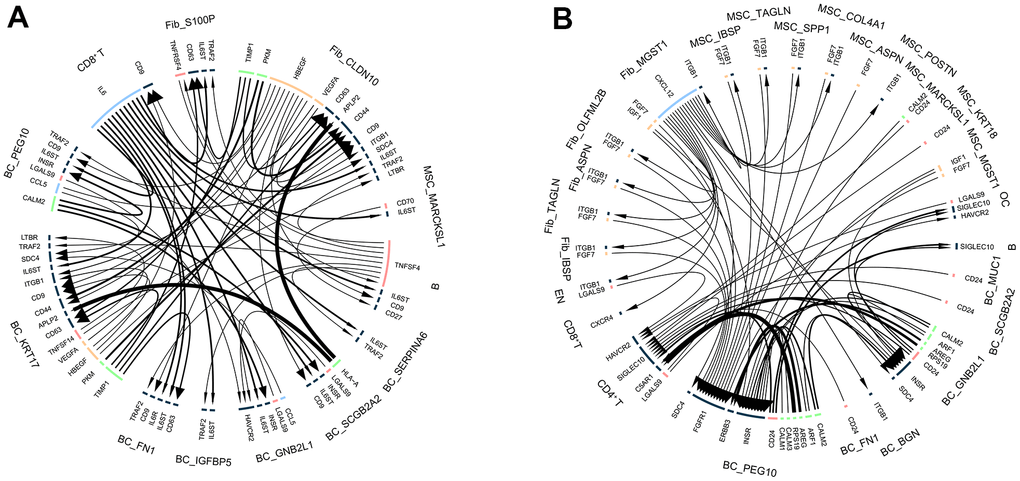
Figure 8. Intercellular communication in breast cancer bone metastases. (A) Intercellular communication of cell subpopulations in control samples. (B) Intercellular communication of cell subpopulations in bone metastasis samples.
Discussion
Recurrence of ER+ breast cancer leads to high mortality rates every year, therefore the specificity of BM must be identified by dissecting the mechanism of action between single cells. The development of bone metastases in breast cancer patients is characterized by complications and poor prognosis, as well as leading to a reduced quality of life for patients. In BM, different cells have different roles in cancer progression and metastasis, and some cells with significant differences are emerging as important cellular players in BM [21, 22]. Here, we constructed a global single-cell landscape of BM based on scRNA-seq data from primary tumor tissues of BM patients and BM tissues, and further explored the pathways, differentiation developmental trajectories and transcriptional regulatory targets involved in cell subpopulations that may have important roles in BM.
Bone is the preferred site of BC cells metastasis, and after metastasis to bone, tumor cells regulate the interactions between different cell types through molecular mechanisms that alter bone homeostasis and thus tumor cell survival, dormancy and/or proliferation [23]. In the present study, tumor cells were observed to be reduced in BM and highly expressed MUC1, SCGB2A2, FN1, BGN, and PEG10. Mucin 1 (MUC1), also known as cancer antigen 15-3 (CA 15-3), has the potential to promote BC cell motility and metastasis. It has been demonstrated that MUC1 adheres to E-selectin and intercellular adhesion molecule-1 (ICAM-1) on the endothelial surface and that ICAM-1 activates the Src oncogene, thereby enhancing BC cell motility and metastatic potential [24]. A study by Iman Mamdouh Talaat et al. tentatively demonstrated that bone marrow horse injection hemoglobin-1 (SCGB2A2) can be used as a tool to study breast cancer early BM. Fibronectin 1 (FN1) is an extracellular matrix protein that may play an important role in inhibiting BC-associated bone loss. In contrast, BGN and PEG10 have not been studied in BM. In addition, enrichment analysis revealed that breast cancer cells in BM are susceptible to oxidative damage and exhibit high levels of oxidative stress, which plays a key role in apoptosis. The mechanistic role of the oxidative microenvironment on BC cells in bone remains largely controversial [25, 26]. Previous studies have shown that in breast cancer, oxidative stress has different effects on primary tumors and distal metastatic organs at different pathological stages [27]. However, recent studies have found that accumulation of oxidative stress may lead to tumor cell death [28]. This is consistent with our study.
To date, only preliminary studies have been performed on the phenotype and transcript levels of fibroblasts in patients with bone metastases. BM often proceeds through multiple steps, including multiple metastases from the primary site of cancer cell growth, invasion, migration through the body circulation and extravasation, seeding to distant organs and subsequent steps of proliferation therein [29, 30]. Subpopulation analysis revealed significant abundance of Fibroblasts_CLDN10 and Fibroblasts_S100P in controls, Fibroblasts_IBSP, Fibroblasts_TAGLN, Fibroblasts_ASPN, Fibroblasts_OLFML2B, Fibroblasts Fibroblasts_KRT19 and Fibroblasts_MGST1 were significantly abundant in BM. analysis by Jinling Liao et al. revealed that CLDN10 expression levels were reduced in breast cancer tissues compared to normal breast tissues [31]. In addition, a pro-metastatic and developmental role of S100P in BC has been identified. IBSP attracts osteoclasts and creates an osteoclast-rich environment in bone, assisting in the delivery of exosomal miR-19a to osteoclasts to induce osteoclastogenesis [32]. And TAGLN, ASPN, KRT19 and MGST1 have important roles in the prognosis, invasion, metastasis and drug resistance of breast cancer [33–36]. In the present study, fibroblast subpopulations were found to be significantly involved in Cytokine-cytokine receptor interaction, TNF signaling pathway and TGF-beta signaling pathway, and Focal adhesion, and fibroblasts can contact in an intercellular dependence to regulate migration and invasion capacity. Force transmission is mediated by heterogeneous adhesion involving N-calmodulin on fibroblast membranes and E-calmodulin on cancer cell membranes. Fibroblast-derived cytokines and chemokines can contribute to the immunosuppressive tumor microenvironment by recruiting and producing immunosuppressive cells [21]. These results suggest a role of fibroblast subpopulations in the promotion of BM.
During the BM process, BC cells migrate together with MSCs from the primary foci to the bone marrow, a process that is dependent on bone bridge proteins [37]. Interestingly, the MSC_MARCKSL1 subpopulation was found in this study to be located at an early position in differentiation development, with a high differentiation capacity to further differentiate to subpopulations specifically present in BM. In addition, the prognostic value of MARCKSL1 in breast cancer has been gradually investigated [38, 39]. However, its regulatory role in BM is still unknown, and this study proposes to imagine MARCKSL1 as an important regulatory molecule in the BM process, and its study may provide help for the treatment of BM. In addition, MSCs have a strong osteogenic potential [22], and in the present study it was also found that MSC subpopulations were significantly enriched in some BPs related to skeletal growth and development and oxidative stress. Compared to highly differentiated cell types, MSCs have a weaker antioxidant capacity and is more sensitive to oxidative responses [40]. These studies highlight the complexity of MSCs and further studies are needed to understand whether MSCs can be used clinically in the treatment of bone metastases.
Previous studies focused on the exploration of a single gene or a unique type of cell in BM. However, no comprehensive single cell profiling analysis of BM has been conducted, largely ignoring the impact of intercellular interactions on BM. In conclusion, our results provide a preliminary subpopulation landscape of the BM tumor microenvironment and reveal the differentiated developmental trajectories and transcriptional regulatory targets of these subpopulations, revealing the transcriptional heterogeneity hidden in the population-averaged measurements and providing ideas for identifying new targets for personalized therapeutic approaches. However, the results need to be validated using experiments and larger samples.
Supplementary Materials
Author Contributions
Minmin Zhang responsible for manuscript writing; Xiao Chai responsible for manuscript writing and proofreading; Li Wang responsible for data collection; Ke Mo responsible for data analysis; Wenyang Chen responsible for data collation; Xiangtao Xie responsible for the construction of ideas. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This study was supported by the Guangxi Natural Science Foundation (2023GXNSFBA026169 and 2023GXNSFAA026407), the Project of Liuzhou Scientific Research and Technology Development Plan (2022SB002 and 2022SB014) and the Scientific Research Project of Guangxi Health Commission (Z20210033 and GXZYZ20210011).
References
- 1. Waks AG, Winer EP. Breast Cancer Treatment: A Review. JAMA. 2019; 321:288–300. https://doi.org/10.1001/jama.2018.19323 [PubMed]
- 2. Tulotta C, Ottewell P. The role of IL-1B in breast cancer bone metastasis. Endocr Relat Cancer. 2018; 25:R421–34. https://doi.org/10.1530/ERC-17-0309 [PubMed]
- 3. Momenimovahed Z, Salehiniya H. Epidemiological characteristics of and risk factors for breast cancer in the world. Breast Cancer (Dove Med Press). 2019; 11:151–64. https://doi.org/10.2147/BCTT.S176070 [PubMed]
- 4. Britt KL, Cuzick J, Phillips KA. Key steps for effective breast cancer prevention. Nat Rev Cancer. 2020; 20:417–36. https://doi.org/10.1038/s41568-020-0266-x [PubMed]
- 5. Hu C, Hart SN, Gnanaolivu R, Huang H, Lee KY, Na J, Gao C, Lilyquist J, Yadav S, Boddicker NJ, Samara R, Klebba J, Ambrosone CB, et al. A Population-Based Study of Genes Previously Implicated in Breast Cancer. N Engl J Med. 2021; 384:440–51. https://doi.org/10.1056/NEJMoa2005936 [PubMed]
- 6. Tahara RK, Brewer TM, Theriault RL, Ueno NT. Bone Metastasis of Breast Cancer. Adv Exp Med Biol. 2019; 1152:105–29. https://doi.org/10.1007/978-3-030-20301-6_7 [PubMed]
- 7. Anderson NM, Simon MC. The tumor microenvironment. Curr Biol. 2020; 30:R921–5. https://doi.org/10.1016/j.cub.2020.06.081 [PubMed]
- 8. Hinshaw DC, Shevde LA. The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res. 2019; 79:4557–66. https://doi.org/10.1158/0008-5472.CAN-18-3962 [PubMed]
- 9. Li H, Lin X, Yang D, Chen Z, Wang X, Re F, Wei J, Chen J. Cancer-associated fibroblasts support bone tropic metastasis by acting as coordinators between the tumor microenvironment and bone matrix in breast cancer. Neoplasma. 2021; 68:10–22. https://doi.org/10.4149/neo_2020_200905N951 [PubMed]
- 10. Karnoub AE, Dash AB, Vo AP, Sullivan A, Brooks MW, Bell GW, Richardson AL, Polyak K, Tubo R, Weinberg RA. Mesenchymal stem cells within tumour stroma promote breast cancer metastasis. Nature. 2007; 449:557–63. https://doi.org/10.1038/nature06188 [PubMed]
- 11. Jezierska-Drutel A, Rosenzweig SA, Neumann CA. Role of oxidative stress and the microenvironment in breast cancer development and progression. Adv Cancer Res. 2013; 119:107–25. https://doi.org/10.1016/B978-0-12-407190-2.00003-4 [PubMed]
- 12. Padmanaban V, Krol I, Suhail Y, Szczerba BM, Aceto N, Bader JS, Ewald AJ. E-cadherin is required for metastasis in multiple models of breast cancer. Nature. 2019; 573:439–44. https://doi.org/10.1038/s41586-019-1526-3 [PubMed]
- 13. Butler A, Hoffman P, Smibert P, Papalexi E, Satija R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat Biotechnol. 2018; 36:411–20. https://doi.org/10.1038/nbt.4096 [PubMed]
- 14. Stuart T, Butler A, Hoffman P, Hafemeister C, Papalexi E, Mauck WM 3rd, Hao Y, Stoeckius M, Smibert P, Satija R. Comprehensive Integration of Single-Cell Data. Cell. 2019; 177:1888–902.e21. https://doi.org/10.1016/j.cell.2019.05.031 [PubMed]
- 15. Becht E, McInnes L, Healy J, Dutertre CA, Kwok IWH, Ng LG, Ginhoux F, Newell EW. Dimensionality reduction for visualizing single-cell data using UMAP. Nat Biotechnol. 2018. [Epub ahead of print]. https://doi.org/10.1038/nbt.4314 [PubMed]
- 16. Li Z, McGinn O, Wu Y, Bahreini A, Priedigkeit NM, Ding K, Onkar S, Lampenfeld C, Sartorius CA, Miller L, Rosenzweig M, Cohen O, Wagle N, et al. ESR1 mutant breast cancers show elevated basal cytokeratins and immune activation. Nat Commun. 2022; 13:2011. https://doi.org/10.1038/s41467-022-29498-9 [PubMed]
- 17. Yu G, Wang LG, Han Y, He QY. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS. 2012; 16:284–7. https://doi.org/10.1089/omi.2011.0118 [PubMed]
- 18. Tirosh I, Izar B, Prakadan SM, Wadsworth MH 2nd, Treacy D, Trombetta JJ, Rotem A, Rodman C, Lian C, Murphy G, Fallahi-Sichani M, Dutton-Regester K, Lin JR, et al. Dissecting the multicellular ecosystem of metastatic melanoma by single-cell RNA-seq. Science. 2016; 352:189–96. https://doi.org/10.1126/science.aad0501 [PubMed]
- 19. Trapnell C, Cacchiarelli D, Grimsby J, Pokharel P, Li S, Morse M, Lennon NJ, Livak KJ, Mikkelsen TS, Rinn JL. The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat Biotechnol. 2014; 32:381–6. https://doi.org/10.1038/nbt.2859 [PubMed]
- 20. Van de Sande B, Flerin C, Davie K, De Waegeneer M, Hulselmans G, Aibar S, Seurinck R, Saelens W, Cannoodt R, Rouchon Q, Verbeiren T, De Maeyer D, Reumers J, et al. A scalable SCENIC workflow for single-cell gene regulatory network analysis. Nat Protoc. 2020; 15:2247–76. https://doi.org/10.1038/s41596-020-0336-2 [PubMed]
- 21. Mukaida N, Zhang D, Sasaki SI. Emergence of Cancer-Associated Fibroblasts as an Indispensable Cellular Player in Bone Metastasis Process. Cancers (Basel). 2020; 12:2896. https://doi.org/10.3390/cancers12102896 [PubMed]
- 22. Sun X, Li K, Zha R, Liu S, Fan Y, Wu D, Hase M, Aryal UK, Lin CC, Li BY, Yokota H. Preventing tumor progression to the bone by induced tumor-suppressing MSCs. Theranostics. 2021; 11:5143–59. https://doi.org/10.7150/thno.58779 [PubMed]
- 23. Mishra A, Shiozawa Y, Pienta KJ, Taichman RS. Homing of cancer cells to the bone. Cancer Microenviron. 2011; 4:221–35. https://doi.org/10.1007/s12307-011-0083-6 [PubMed]
- 24. Haddon L, Hugh J. MUC1-mediated motility in breast cancer: a review highlighting the role of the MUC1/ICAM-1/Src signaling triad. Clin Exp Metastasis. 2015; 32:393–403. https://doi.org/10.1007/s10585-015-9711-8 [PubMed]
- 25. Hayes JD, Dinkova-Kostova AT, Tew KD. Oxidative Stress in Cancer. Cancer Cell. 2020; 38:167–97. https://doi.org/10.1016/j.ccell.2020.06.001 [PubMed]
- 26. Gill JG, Piskounova E, Morrison SJ. Cancer, Oxidative Stress, and Metastasis. Cold Spring Harb Symp Quant Biol. 2016; 81:163–75. https://doi.org/10.1101/sqb.2016.81.030791 [PubMed]
- 27. Harris IS, Treloar AE, Inoue S, Sasaki M, Gorrini C, Lee KC, Yung KY, Brenner D, Knobbe-Thomsen CB, Cox MA, Elia A, Berger T, Cescon DW, et al. Glutathione and thioredoxin antioxidant pathways synergize to drive cancer initiation and progression. Cancer Cell. 2015; 27:211–22. https://doi.org/10.1016/j.ccell.2014.11.019 [PubMed]
- 28. Piskounova E, Agathocleous M, Murphy MM, Hu Z, Huddlestun SE, Zhao Z, Leitch AM, Johnson TM, DeBerardinis RJ, Morrison SJ. Oxidative stress inhibits distant metastasis by human melanoma cells. Nature. 2015; 527:186–91. https://doi.org/10.1038/nature15726 [PubMed]
- 29. Gascard P, Tlsty TD. Carcinoma-associated fibroblasts: orchestrating the composition of malignancy. Genes Dev. 2016; 30:1002–19. https://doi.org/10.1101/gad.279737.116 [PubMed]
- 30. Bu L, Baba H, Yoshida N, Miyake K, Yasuda T, Uchihara T, Tan P, Ishimoto T. Biological heterogeneity and versatility of cancer-associated fibroblasts in the tumor microenvironment. Oncogene. 2019; 38:4887–901. https://doi.org/10.1038/s41388-019-0765-y [PubMed]
- 31. Liao J, Li J, Cheng H, Chen Y, Mo Z. CLDN10 single nucleotide polymorphism rs1325774 alters the risk of breast cancer in south Chinese women. Medicine (Baltimore). 2018; 97:e13187. https://doi.org/10.1097/MD.0000000000013187 [PubMed]
- 32. Wu K, Feng J, Lyu F, Xing F, Sharma S, Liu Y, Wu SY, Zhao D, Tyagi A, Deshpande RP, Pei X, Ruiz MG, Takahashi H, et al. Exosomal miR-19a and IBSP cooperate to induce osteolytic bone metastasis of estrogen receptor-positive breast cancer. Nat Commun. 2021; 12:5196. https://doi.org/10.1038/s41467-021-25473-y [PubMed]
- 33. Sayar N, Karahan G, Konu O, Bozkurt B, Bozdogan O, Yulug IG. Transgelin gene is frequently downregulated by promoter DNA hypermethylation in breast cancer. Clin Epigenetics. 2015; 7:104. https://doi.org/10.1186/s13148-015-0138-5 [PubMed]
- 34. Zhang P, Qian B, Liu Z, Wang D, Lv F, Xing Y, Xiao Y. Identification of novel biomarkers of prostate cancer through integrated analysis. Transl Androl Urol. 2021; 10:3239–54. https://doi.org/10.21037/tau-21-401 [PubMed]
- 35. Wang XM, Zhang Z, Pan LH, Cao XC, Xiao C. KRT19 and CEACAM5 mRNA-marked circulated tumor cells indicate unfavorable prognosis of breast cancer patients. Breast Cancer Res Treat. 2019; 174:375–85. https://doi.org/10.1007/s10549-018-05069-9 [PubMed]
- 36. Zeng X, Morgenstern R, Nyström AM. Nanoparticle-directed sub-cellular localization of doxorubicin and the sensitization breast cancer cells by circumventing GST-mediated drug resistance. Biomaterials. 2014; 35:1227–39. https://doi.org/10.1016/j.biomaterials.2013.10.042 [PubMed]
- 37. Kuo MC, Kothari AN, Kuo PC, Mi Z. Cancer stemness in bone marrow micrometastases of human breast cancer. Surgery. 2018; 163:330–5. https://doi.org/10.1016/j.surg.2017.07.027 [PubMed]
- 38. Egeland NG, Austdal M, van Diermen-Hidle B, Rewcastle E, Gudlaugsson EG, Baak JPA, Skaland I, Janssen EA, Jonsdottir K. Validation study of MARCKSL1 as a prognostic factor in lymph node-negative breast cancer patients. PLoS One. 2019; 14:e0212527. https://doi.org/10.1371/journal.pone.0212527 [PubMed]
- 39. Jonsdottir K, Zhang H, Jhagroe D, Skaland I, Slewa A, Björkblom B, Coffey ET, Gudlaugsson E, Smaaland R, Janssen EA, Baak JP. The prognostic value of MARCKS-like 1 in lymph node-negative breast cancer. Breast Cancer Res Treat. 2012; 135:381–90. https://doi.org/10.1007/s10549-012-2155-9 [PubMed]
- 40. Denu RA, Hematti P. Effects of Oxidative Stress on Mesenchymal Stem Cell Biology. Oxid Med Cell Longev. 2016; 2016:2989076. https://doi.org/10.1155/2016/2989076 [PubMed]