



Introduction
Polycystic ovary syndrome (PCOS) is a common endocrine disease among women of reproductive age, which is reported in approximately 5–10% women worldwide [1]. The etiology of PCOS remains unclear but may be associated with genetic and environmental factors. According to the Rotterdam consensus, women who present with at least two of the following symptoms should be diagnosed as having PCOS: clinical or biochemical hyperandrogenism, oligo-ovulation or anovulation and the presence of polycystic ovaries on ultrasonography [2]. However, the diagnosis and treatment of PCOS remain a great challenge owing to the great heterogeneity of this systemic disease. Therefore, understanding the pathogenesis of PCOS is important for developing new diagnostic and treatment methods.
Angiogenesis refers to the formation of new blood vessels from existing vessels, which dominantly entails the activation, proliferation, and migration of endothelial cells (ECs) and is counterbalanced by proangiogenic and antiangiogenic factors [3]. In sexually mature female mammals, the vasculature of reproductive system presents periodical regeneration and degeneration. The periodical formation and regression of ovarian blood vessels provide necessary nutrients and hormones for follicular growth, ovulation, luteum formation and secretion of ovarian hormones [4]. Interestingly, ovarian angiogenesis is abnormal in PCOS patients, manifested as increased ovarian stromal vascularization [5, 6]. In addition, the expression of angiogenic factors such as VEGF in PCOS women is altered [7]. VEGF is one of the most significant angiogenic factors in PCOS, which has been reported to be upregulated in the ovaries, serum, and follicular fluid of patients with PCOS [8–10]. Besides, serum VEGF levels in obese PCOS patients are significantly higher than in non-obese PCOS patients [11]. Moreover, a positive correlation is found between the levels of serum VEGF and ovarian vascularization [12]. Nevertheless, the specific mechanisms leading to the increase in VEGF levels and ovarian stromal vascularization in PCOS patients remain unclear.
Approximately 50% patients with PCOS are obese [13]. Obesity, which is characterized by the deposition of adipose tissue (AT), aggravates endocrine and metabolic disorder in PCOS patients. As the major constituent of AT, adipocytes secret adipokines (such as leptin, adiponectin and resistin) that are involved in the development of PCOS-related weight gain, insulin resistance, inflammation, and granulosa apoptosis, which denotes obesity modifies PCOS manifestations by adipocytes [14]. Exosomes are extracellular vesicles (EVs) with a diameter of 30–200 nm, which are responsible for cell–cell communication. Donor cells can regulate the biological function of recipient cells by delivering the contents of exosomes, such as proteins, lipids, mRNAs and noncoding RNAs (ncRNAs) [15]. As energy-saving and endocrine cells, adipocytes exert great metabolic regulatory effects [16]. Notably, studies have indicated that adipocyte-derived exosomes participate in the regulation of glucose metabolism, lipid synthesis and endothelial injury as a new type of adipokines, providing a new explanation for the pathogenesis of obesity-related diseases [17–19]. However, to the best of our knowledge, no study has explored the role of adipocyte-derived exosomes in the pathogenesis of PCOS.
MicroRNAs (miRNAs) are short endogenous ncRNAs that have been widely studied as the functional contents of exosomes owing to their strong regulatory effects on gene expression. AT is an important source of exosomal miRNAs in the circulatory system [20]. As a member of the miR-30 family, miR-30c-5p is usually recognized as a regulator of adipogenesis, pathological cardiac remodeling, tumour invasion and angiogenesis [21, 22]. The expression of miR-30c-5p was upregulated during the adipogenesis of human multipotent adipose-derived stem cells. Besides, miR-30c-5p overexpression boosted adipocyte marker gene expression and triglyceride accumulation [23]. A previous study found miR-30c-5p promoted tube formation in human umbilical vein endothelial cells (HUVECs) in vitro, and downregulation of miR-30c-5p in zebrafish significantly inhibited angiogenesis in vivo [24]. Moreover, the serum levels of miR-30c are higher in PCOS patients [25]. Taken together, we hypothesized that adipocyte-derived exosomes might be involved in the pathogenesis of PCOS by delivering miR-30c-5p to promote ovarian angiogenesis and verified this hypothesis by performing experiments described below.
Results
Adipocyte-derived exosomes were internalized by HOMECs
Adipogenic differentiation of 3T3-L1 cells was induced to imitate mature adipocytes in vivo. Differentiated 3T3-L1 cells exhibited typical adipocyte phenotypes as indicated by oil red O staining (Figure 1A). Exosomes from differentiated adipocytes were isolated via ultracentrifugation and identified via transmission electron microscopy (TEM), nanoparticle tracking analysis (NTA) and western blotting. TEM revealed the presence of vesicles with a bi-layer membrane structure ranging from 30 to 200 nm (Figure 1B). The average diameter of the vesicles was 136.3 nm detected by NTA (Figure 1C). Western blotting revealed the expression of exosome markers, namely, CD9 and CD63, in isolated vesicles. Fatty acid binding protein 4 (FABP4) is a specific marker of adipocyte-derived exosomes [26], which was detected in differentiated 3T3-L1 cells and vesicles (Figure 1D). Above results indicated we successfully isolated adipocyte-derived exosomes. To assess whether HOMCEs can internalize adipocyte-derived exosomes, HOMECs were incubated with PKH-26 labelled exosomes, while PBS was added as the control (CON). After 6 h, PKH26-labelled exosomes were observed in HOMECs (Figure 1E). Altogether, these results indicated that adipocyte-derived exosomes were internalized by HOMECs.
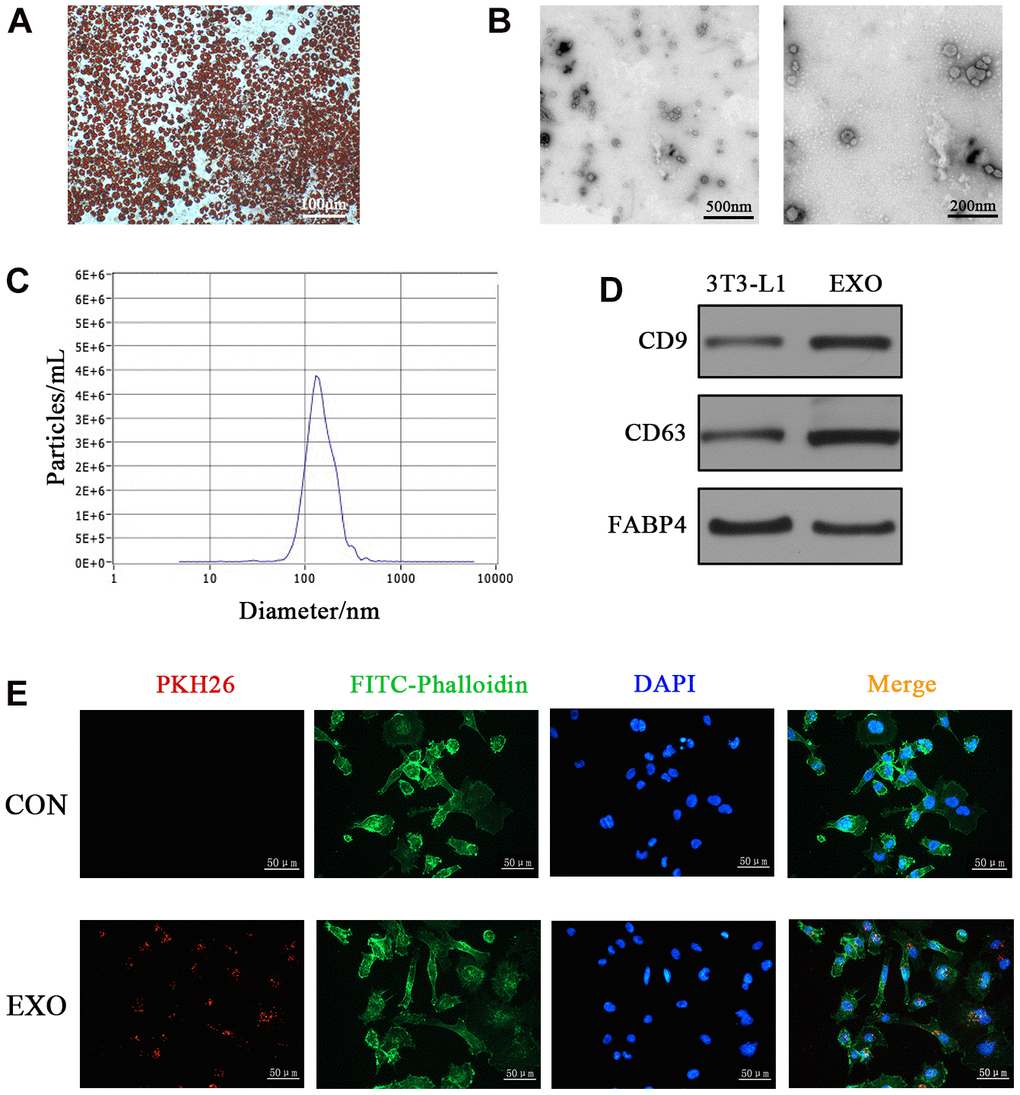
Figure 1. Characterization of adipocyte-derived exosomes. (A) Oil red O staining of differentiated 3T3-L1 cells. (B) TEM analyses of exosomes secreted by adipocytes. (C) NTA of exosomes. (D) Western blotting analyses of exosomal markers (CD9, CD63 and FABP4) in 3T3-L1 cells and adipocyte-derived exosomes. (E) Fluorescence images of HOMECs incubated with PKH26-labelled exosomes (red). The cytoskeleton was stained with phalloidin-FITC (green), and nuclei were stained with DAPI (blue).
Adipocyte-derived exosomes promoted proliferation, migration, and tube formation in HOMECs
To assess the effects of adipocyte-derived exosomes on the biological function of HOMECs, exosomes at different concentrations were added to HOMECs according to the exosome concentrations reported in previous studies [27, 28]. The results of cell counting kit-8 (CCK-8) assay demonstrated that the proliferation rate of HOMECs treated with exosomes was significantly higher than that in the control group (Figure 2A). In addition, the protein expression of Ki67 and proliferating cell nuclear antigen (PCNA) was upregulated in HOMECs treated with exosomes, indicating that adipocyte-derived exosomes strengthened HOMEC proliferation (Figure 2B). Compared with PBS treatment, exosome treatment promoted the migration of HOMECs as indicated by an increased number of migrated cells, upregulation of Vimentin and downregulation of E-cadherin (Figure 2C, 2D). Furthermore, tube formation assay indicated that exosomes enhanced the capillary-like structure formation capacity of HOMECs as indicated by the increased total tube length and number of branches (Figure 2E). Western blotting verified that exosomes enhanced the expression of VEGFA in HOMECs (Figure 2F). Altogether, these results showed that adipocyte-derived exosomes promoted proliferation, migration, and tube formation in HOMECs. Because 20 μg/mL exosomes exerted greater effects on the biological function and protein expression in HOMECs than 10 μg/mL exosomes, the exosome concentration of 20 μg/mL was selected for subsequent cell experiments.
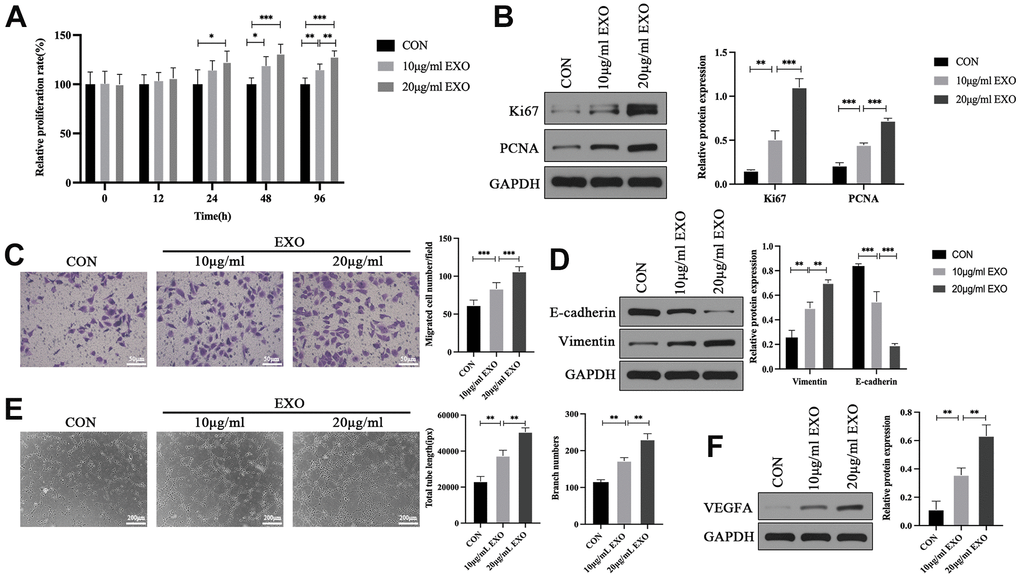
Figure 2. Adipocyte-derived exosomes promote proliferation, migration, and tube formation in HOMECs. (A) Detection of the effect of adipocyte-derived exosomes on the cell proliferation by the CCK-8 assay. (B) Western blotting analyses of Ki67 and PCNA expression in HOMECs. (C) Detection of the effect of adipocyte-derived exosomes on the cell migration by the transwell assay. (D) Western blotting analyses of E-cadherin and Vimentin expression in HOMECs. (E) Detection of the effect of adipocyte-derived exosomes on the cell tube formation by the tube formation assay. (F) Western blotting analyses of VEGFA expression in HOMECs. Data are presented as mean ± SD for three independent experiments. *P<0.05, **P<0.01, ***P<0.001.
miR-30c-5p was delivered by adipocyte-derived exosomes and promoted proliferation, migration, and tube formation in HOMECs
A coculture system was used to verify that adipocyte-derived exosomes delivered miR-30c-5p to HOMECs (Figure 3A). The levels of miR-30c-5p were increased in HOMECs co-cultured with differentiated 3T3-L1 cells but were decreased after adipocytes were treated with the exosome inhibitor GW4869 (Figure 3B). These results indicated that miR-30c-5p was delivered to HOMECs by exosomes. Furthermore, miR-30c-5p mimics/inhibitors/negative controls (NCs) were transfected into HOMECs to investigate the effects of miR-30c-5p on the biological function of HOMECs. The miR-30c-5p levels in HOMECs were significantly increased after transfection of miR-30c-5p mimics while they were decreased after transfection of miR-30c-5p inhibitors (Figure 3C). Subsequently, the role of miR-30c-5p in the proliferation, migration, and tube formation of HOMECs was examined. The results of CCK-8 assay demonstrated that the miR-30c-5p overexpression enhanced the proliferation rate of HOMECs whereas miR-30c-5p inhibition suppressed HOMEC proliferation (Figure 3D). The number of migrated HOMECs was increased after the transfection of miR-30c-5p mimics but decreased after the transfection of miR-30c-5p inhibitors (Figure 3E, 3F). In addition, tube formation assays suggested miR-30c-5p enhanced capillary-like structure formation capacity of HOMECs (Figure 3G, 3H).
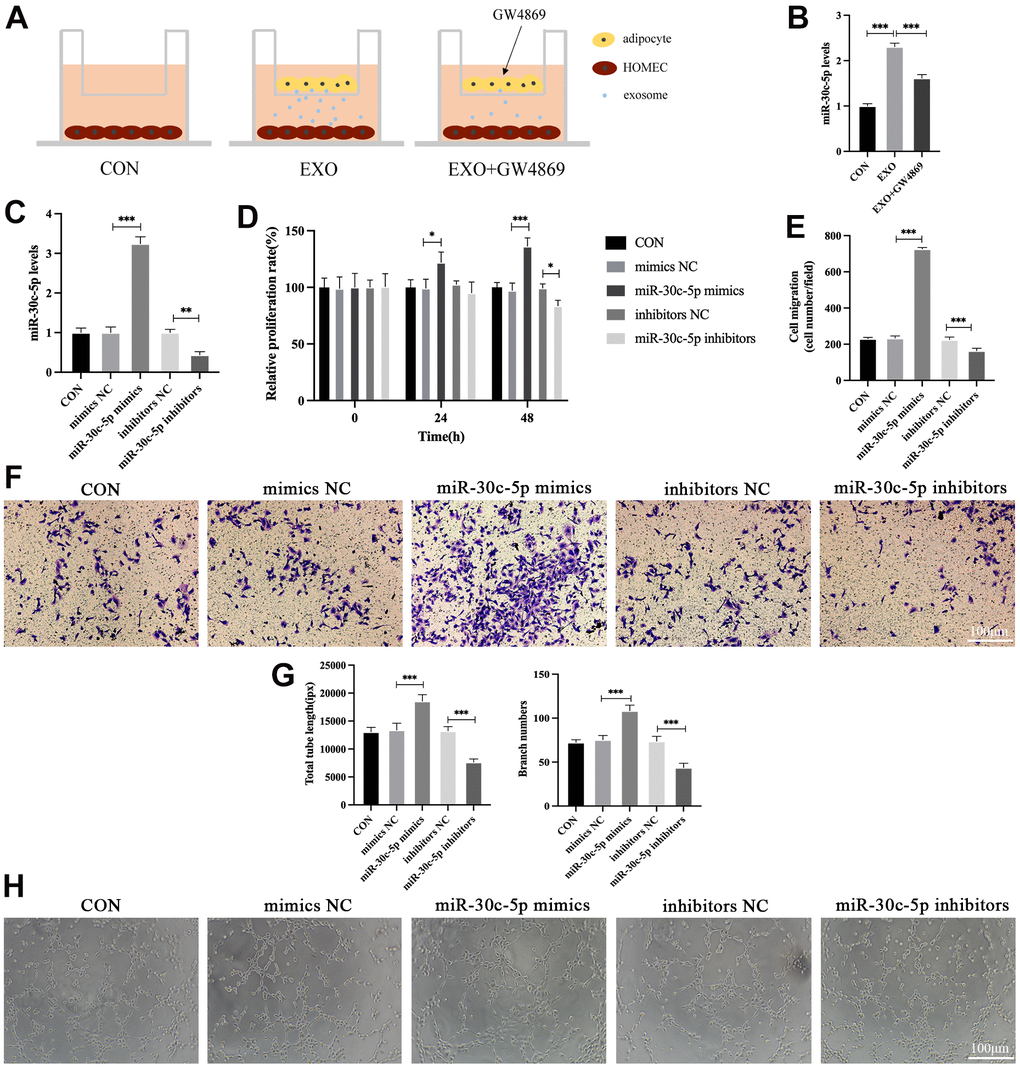
Figure 3. miR-30c-5p is delivered by adipocyte-derived exosomes and promotes proliferation, migration, and tube formation in HOMECs. (A) Coculture of HOMECs and adipocytes treated with or without GW4869. (B) qRT-PCR detection of miR-30c-5p levels in HOMECs in the coculture system. (C) qRT-PCR detection of miR-30c-5p levels after cell transfection. (D) Detection of the effect of miR-30c-5p on the cell proliferation by the CCK-8 assay. (E, F) Detection of the effect of miR-30c-5p on the cell migration by the transwell assay. (G, H) Detection of the effect of miR-30c-5p on the cell tube formation capacity by the tube formation assay. Data are presented as mean ± SD for three independent experiments. *P<0.05, **P<0.01, ***P<0.001.
miR-30c-5p activated the STAT3/VEGFA pathway by targeting SOCS3
To determine the mechanism of miR-30c-5p regulating biological function of HOMECs, the target genes of miR-30c-5p were predicted via bioinformatic analysis. The results indicated that SOCS3 was one of the 12 target genes of miR-30c-5p jointly predicted by TargetScan, miRDB and Starbase (Figure 4A, 4B). TargetScan predicted that the 3’-UTR of the SOCS3 mRNA was the conserved binding site of miR-30c-5p (Figure 4C). Furthermore, the results of dual luciferase reporter assay showed that the transfection of miR-30c-5p mimics reduced the luciferase activity in HOMECs transfected with pmirGLO-SOCS3-WT instead of pmirGLO-SOCS3-MUT (Figure 4D). These results indicated that miR-30c-5p inhibited the transcription of SOCS3 by directly binding to the 3’-UTR of SOCS3 mRNA.
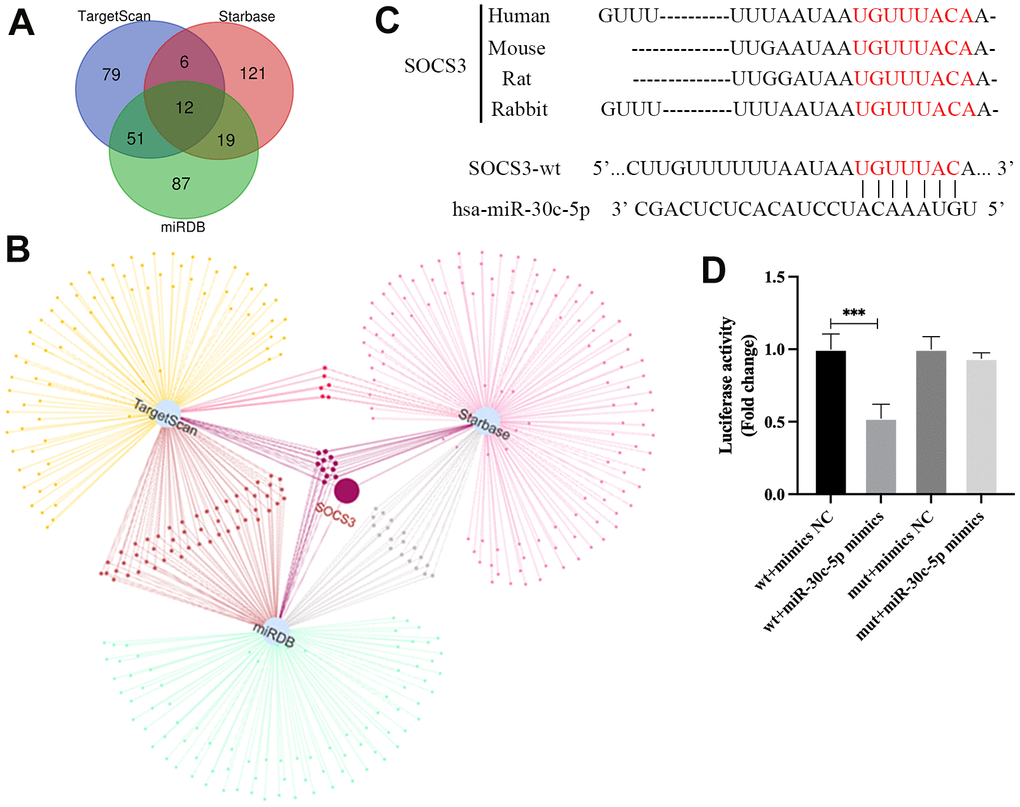
Figure 4. SOCS3 is a target gene of miR-30c-5p. (A, B) Bioinformatic analyses of target genes of miR-30c-5p. (C) Prediction of the binding site of miR-30c-5p in the 3’-UTR of SOCS3 mRNA by TargetScan. (D) Confirmation of the targeting relationship between miR-30c-5p and SOCS3 by the dual luciferase reporter assay. Data are presented as mean ± SD for three independent experiments. ***P<0.001.
SOCS3 can indirectly inhibit the transcription of VEGFA by inhibiting STAT3 [29, 30]. Therefore, the effects of miR-30c-5p on the SOCS3/STAT3/VEGFA pathway were further investigated. Western blotting showed that overexpression of miR-30c-5p suppressed the expression of SOCS3 and promoted the expression of p-STAT3 and VEGFA, whereas miR-30c-5p inhibition significantly upregulated the expression of SOCS3 and inhibited the expression of p-STAT3 and VEGFA (Figure 5A). Furthermore, HOMECs were co-transfected with miR-30c-5p mimics and SOCS3 plasmid to investigate the effects of the miR-30c-5p/SOCS3 axis on the STAT3/VEGFA pathway. The results showed transfection of only miR-30c-5p mimics led to downregulation of SOCS3 and upregulation of p-STAT3 and VEGFA, while this effect was reversed after co-transfection with the SOCS3 plasmid (Figure 5B). Therefore, these results suggest that miR-30c-5p activates the STAT3/VEGFA pathway by targeting SOCS3.
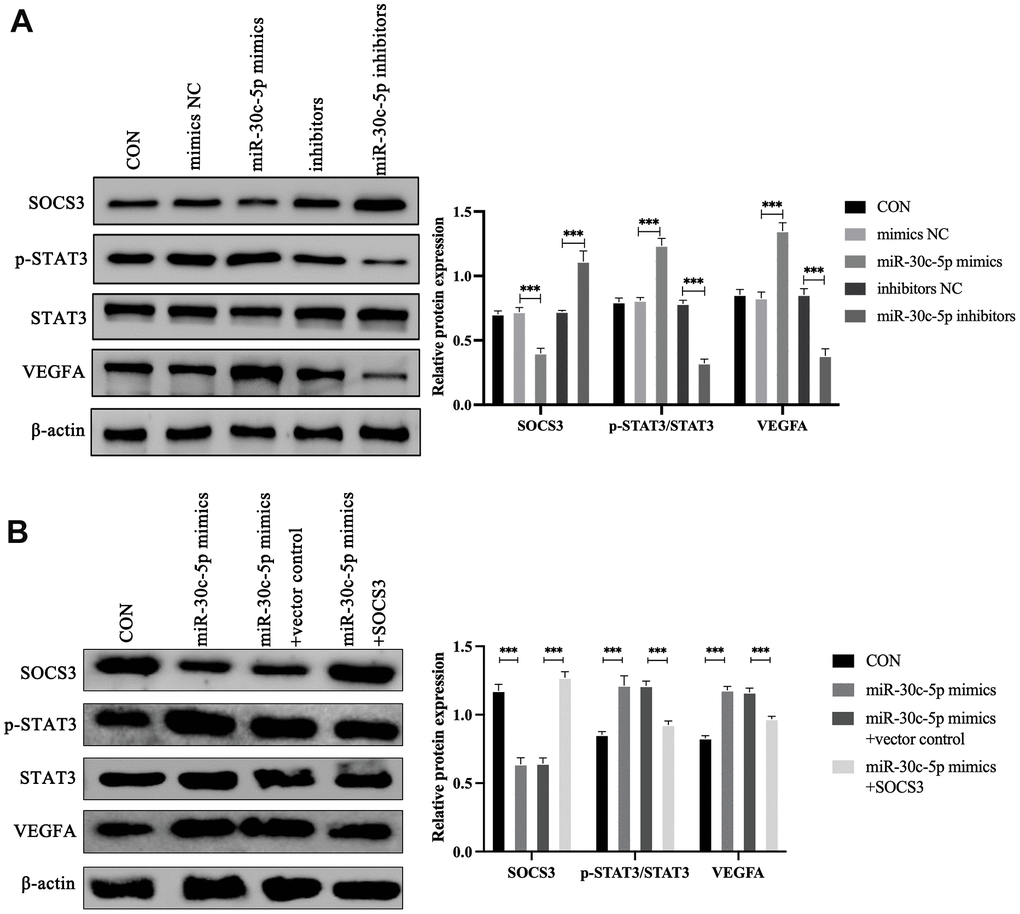
Figure 5. miR-30c-5p regulates the expression of the SOCS3/STAT3/VEGFA pathway. (A) Western blotting analyses of the effect of miR-30c-5p on SOCS3/STAT3/VEGFA pathway expression in HOMECs. (B) Western blotting analyses of the effect of SOCS3 on STAT3/VEGFA pathway expression in HOMECs. Data are presented as mean ± SD for three independent experiments. ***P<0.001.
Adipocyte-derived exosomal miR-30c-5p promoted proliferation, migration, and tube formation in HOMECs and activating the SOCS3/STAT3/VEGFA pathway
To verify whether miR-30c-5p mediates the effects of adipocyte-derived exosomes on the biological function of HOMECs, HOMECs were co-treated with miR-30c-5p inhibitors and adipocyte-derived exosomes. qRT-PCR showed that exosome treatment enhanced the levels of miR-30c-5p in HOMECs, whereas this effect was mitigated after co-treatment with miR-30c-5p inhibitors (Figure 6A). Furthermore, the effects of exosomal miR-30c-5p on the proliferation of HOMECs were examined, and the results revealed that miR-30c-5p inhibitors reversed the stimulative effects of exosomes on the proliferation of HOMECs (Figure 6B). In addition, wound healing assay indicated that miR-30c-5p inhibitors reversed the stimulative effects of exosomes on the migration of HOMECs (Figure 6C). Moreover, tube formation ability of HOMECs simultaneously treated with exosomes and miR-30c-5p inhibitors was weaker than that of HOMECs treated with only exosomes (Figure 6D). Western blotting revealed that exosomes stimulated p-STAT3 and VEGFA expression and suppressed SOCS3 expression in HOMECs, while these effects were alleviated after miR-30c-5p inhibition (Figure 6E).
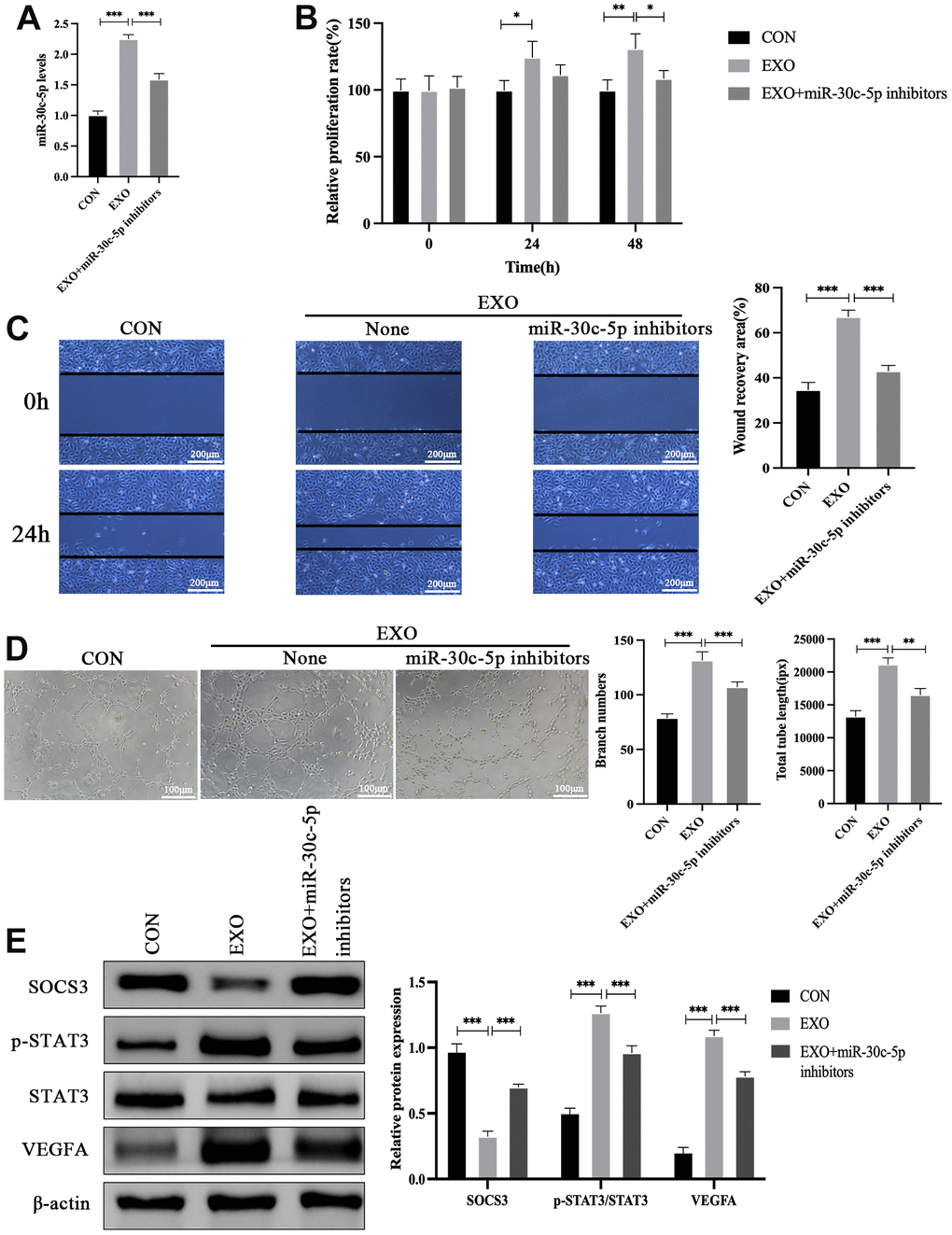
Figure 6. miR-30c-5p mediates the effects of adipocyte-derived exosomes on the biological function of HOMECs and the SOCS3/STAT3/VEGFA pathway. (A) qRT-PCR detection of the effect of exosomes combined with or without miR-30c-5p inhibitors on miR-30c-5p levels in HOMECs. (B) Detection of the effect of exosomal miR-30c-5p on the cell proliferation by the CCK-8 assay. (C) Detection of the effect of exosomal miR-30c-5p on the cell migration by the wound healing assay. (D) Detection of the effect of exosomal miR-30c-5p on the cell tube formation capacity by the tube formation assay. (E) Western blotting analyses of the effect of exosomal miR-30c-5p on SOCS3/STAT3/VEGFA pathway expression in HOMECs. Data are presented as mean ± SD for three independent experiments. *P<0.05, ** P<0.01, ***P<0.001.
Adipocyte-derived exosomal miR-30c-5p exacerbated endocrine and metabolic disorders in mice with PCOS
To verify the abovementioned findings in vivo, dehydroepiandrosterone (DHEA) was used to induce PCOS in mice, and mice with PCOS were treated with adipocyte-derived exosomes and miR-30c-5p antagomir via tail vein injection (Figure 7A). DHEA-treated mice exhibited a disordered estrous cycle and weight gain, with the formation of cystic follicles (CFs) and deterioration of corpus luteum (CL) in the ovaries, indicating the establishment of PCOS models (Supplementary Figure 1A–1C). Immunofluorescence (IF) analyses of the ovaries showed that exosomes in blood circulation were taken up by the ovaries (Figure 7B). miR-30c-5p levels of the ovaries were increased in mice with PCOS. In the exosome-treated groups, exosome treatment alone increased miR-30c-5p levels in the ovaries of PCOS mice, but this effect was mitigated after co-treatment with miR-30c-5p antagomir. (Figure 7C). Notably, exosome treatment combined with or without miR-30c-5p antagomir had no significant effect on the estrous cycle of PCOS mice (Figure 7D). Additionally, PCOS mice gained weight after exosome treatment for 2 weeks, while miR-30c-5p antagomir alleviated this effect (Figure 7E). Furthermore, the glucometabolic state of mice in different groups was evaluated. The results demonstrated that miR-30c-5p antagomir mitigated the effects of exosomes on aggravating insulin resistance in PCOS mice (Figure 7F–7H). Exosome treatment downregulated serum follicle-stimulating hormone (FSH) levels and upregulated serum estradiol (E2), testosterone (T) and VEGFA levels in PCOS mice. However, miR-30c-5p antagomir attenuated these changes (Figure 7I–7L). These results indicate that adipocyte-derived exosomes exacerbate endocrine and metabolic disorders in PCOS via miR-30c-5p.
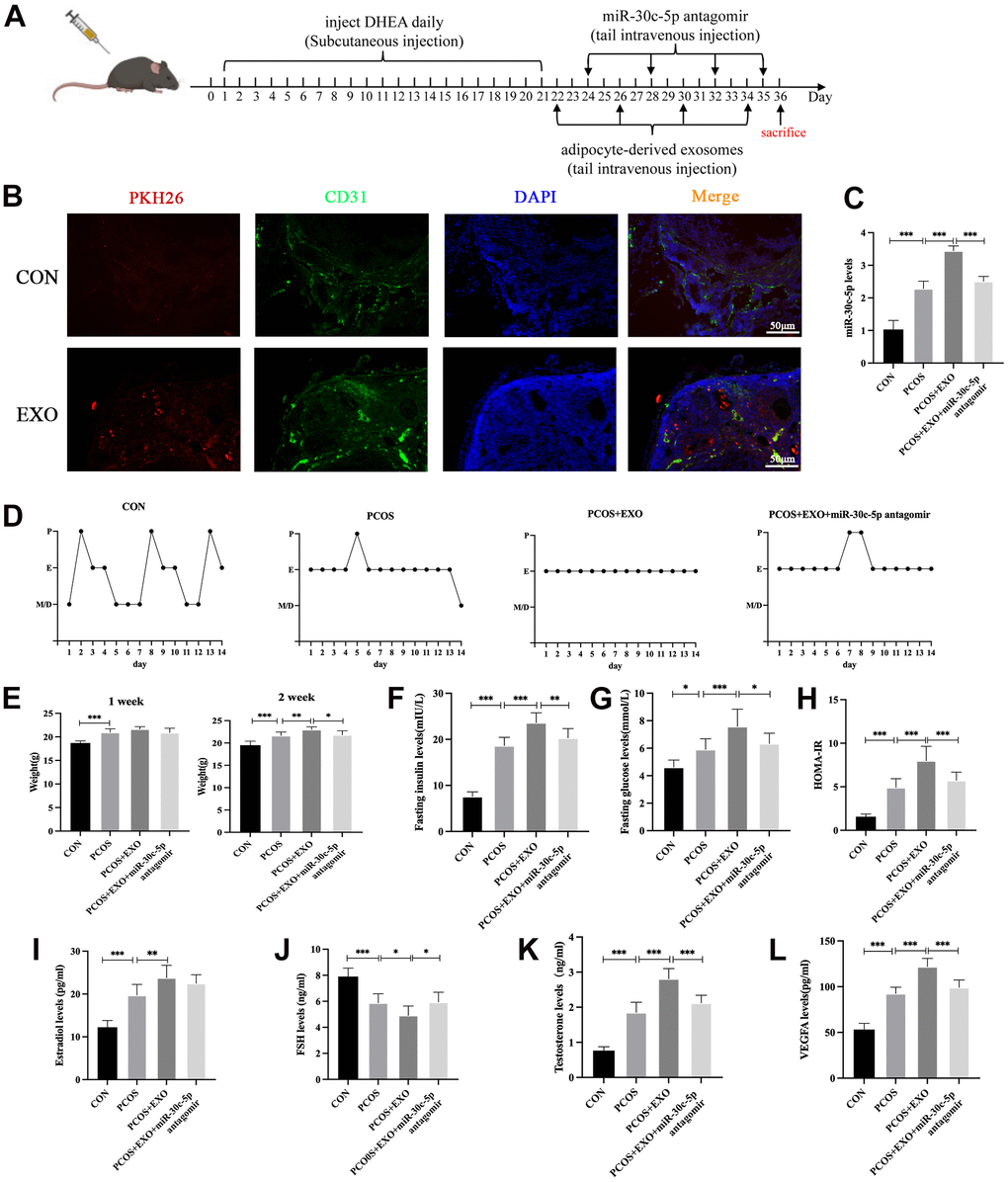
Figure 7. Adipocyte-derived exosomal miR-30c-5p exacerbates endocrine and metabolic disorders in mice with PCOS. (A) Procedure of animal experiments. (B) Visualization of uptake of exosomes by the ovaries in vivo. (C) qRT-PCR detection of miR-30c-5p levels in the ovaries of mice in the indicated groups. (D) Estrous cycle analyses of mice in each group via vaginal smears. (E) Weight change in the indicated groups after administration of exosomes and miR-30c-5p antagomir for 1 and 2 weeks. (F) Fasting insulin levels in the indicated groups. (G) Fasting glucose levels in the indicated groups. (H) HOMA-IR index in the indicated groups. HOMA-IR = fasting insulin × fasting glucose/22.5. (I–L) ELISA detection of serum E2, FSH, T and VEGFA levels in the indicated groups. Proestrus (P), estrus (E), metestrus (M) and diestrus (D). Data are presented as mean ± SD for three independent experiments. *P<0.05, ** P<0.01, ***P<0.001.
Adipocyte-derived exosomal miR-30c-5p mediated ovarian angiogenesis through the SOCS3/STAT3/VEGFA pathway
In in vivo experiments, exosome treatment increased the number of CFs and deteriorated CL in the ovaries of PCOS mice. Nevertheless, miR-30c-5p antagomir attenuated these changes (Figure 8A). Ovarian angiogenesis was determined via immunohistochemical (IHC) analyses, which suggested that adipocyte-derived exosomes facilitated ovarian angiogenesis as shown by the downregulation of SOCS3 and upregulation of CD31 and VEGFA expression in ovaries. In addition, compared with PCOS mice treated with only exosomes, ovarian angiogenesis was inhibited in mice simultaneously treated with exosomes and miR-30c-5p antagomir (Figure 8B). Western blotting indicated adipocyte-derived exosomes suppressed SOCS3 expression and upregulated p-STAT3 and VEGFA expression in the ovaries of PCOS mice. Moreover, miR-30c-5p antagomir partially reversed the effects of adipocyte-derived exosomes on the expression of the SOCS3/STAT3/VEGFA pathway in ovaries (Figure 8C).
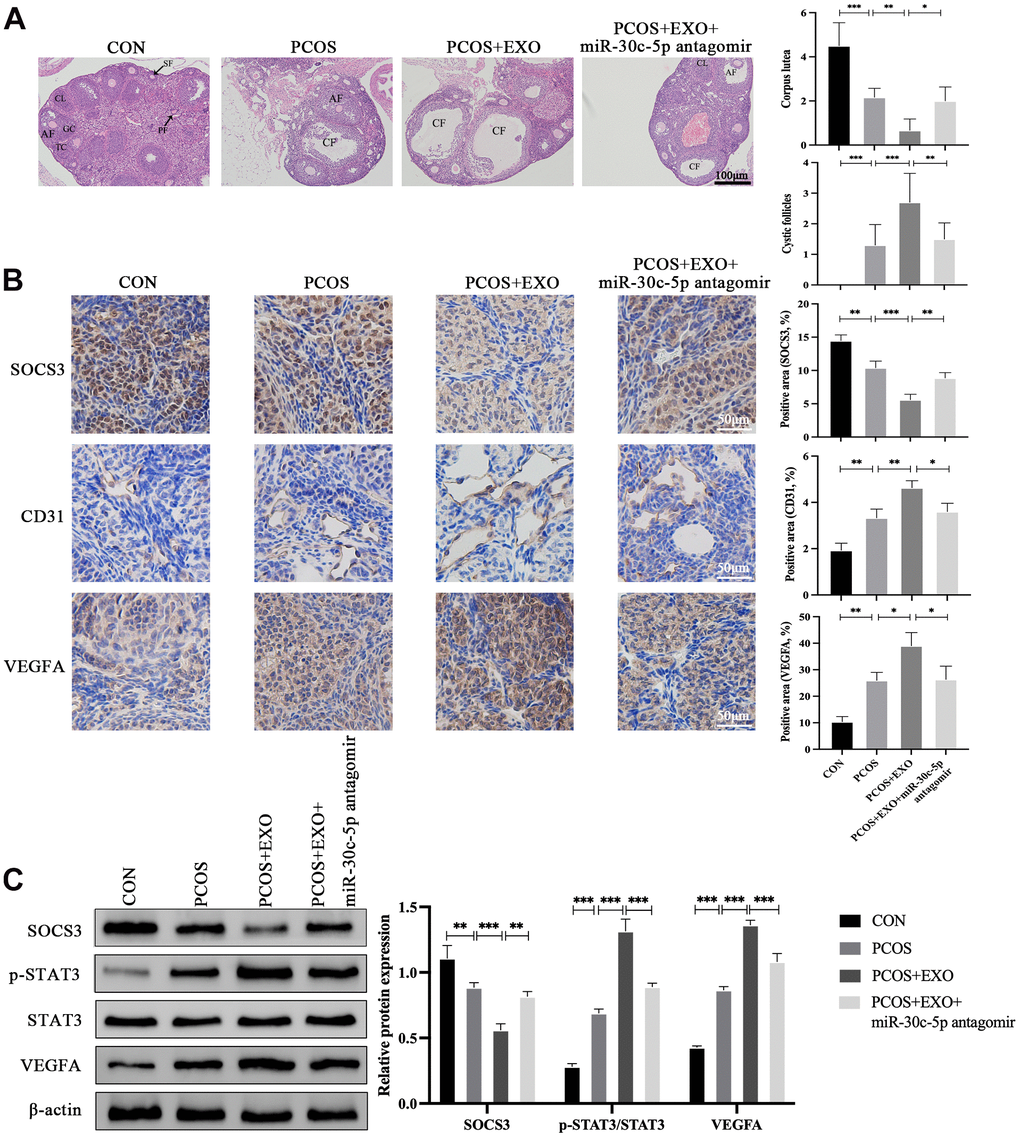
Figure 8. Adipocyte-derived exosomal miR-30c-5p mediates ovarian angiogenesis through the SOCS3/STAT3/VEGFA pathway. (A) Visualization of the ovarian morphology in the indicated groups by HE staining. (B) Detection of SOCS3, CD31 and VEGFA expression in the ovaries in the indicated groups by IHC staining. (C) Western blotting analyses of the SOCS3/STAT3/VEGFA pathway expression in the indicated groups. Primary follicles (PFs), granulosa cells (GCs), theca cells (TCs), secondary follicles (SFs), antral follicles (AFs), corpus luteum (CL), and cystic follicles (CFs). Data are presented as mean ± SD for three independent experiments. *P<0.05, ** P<0.01, ***P<0.001.
Discussion
Balanced ovarian angiogenesis is essential for normal ovarian function. Therefore, elucidating the pathogenesis of abnormal ovarian angiogenesis and VEGF expression in PCOS is of great significance. In the present study, we found adipocyte-derived exosomes not only promoted proliferation, migration, and tube formation in HOMECs but also enhanced VEGFA expression. In addition, miR-30c-5p mediated the effects of adipocyte-derived exosomes on regulating the biological function of HOMECs and activating the STAT3/VEGFA pathway by targeting SOCS3. Furthermore, adipocyte-derived exosomal miR-30c-5p also participated in the development of endocrine and metabolic disorders, ovarian angiogenesis and VEGFA upregulation in PCOS mice. Overall, this study provides novel insights into the aetiology and pathology of PCOS.
An extracellular vesicle axis has been reported between ECs and adipocytes. ECs transfer caveolin 1-containing EVs to neighbouring adipocytes, whereas adipocytes reciprocate by releasing EVs to ECs. This reciprocal communication indicates that adipocytes and ECs can mutually regulate their functions through EVs [31]. Adipocyte-derived exosomal small nucleolar RNA host gene 9 (SNHG9) alleviated inflammation and apoptosis in ECs by targeting TNF receptor-associated death domain (TRADD), thus participating in the development of obesity-related endothelial cell dysfunction [19]. Moreover, insulin-resistant adipocyte-derived exosomes enhanced capillary-like structure formation capacity in HUVECs and promote vasa vasorum angiogenesis in vivo via Shh [27]. Our study showed that adipocyte-derived exosomes promoted proliferation, migration, and tube formation in HOMECs. VEGFA (also referred to as VEGF) is the main promoter of angiogenesis, which promotes EC proliferation and migration and vascular permeability by binding to its receptor [32]. In this study, adipocyte-derived exosomes enhanced VEGFA expression both in HOMECs and ovaries. Therefore, we suggest adipocyte-derived exosomes are involved in ovarian angiogenesis in PCOS.
Insulin resistance and obesity are common complications of PCOS. Previous studies have reported that adipocyte-derived exosomes can induce insulin resistance and energy metabolism dysfunction by regulating the function and phenotypic transformation of skeletal muscle, macrophages and pro-opiomelanocortin neurons [17, 33, 34]. In this study, adipocyte-derived exosomes exacerbated insulin resistance and energy metabolism disorder in PCOS mice as indicated by the upregulation of fasting blood glucose levels, fasting insulin, homeostasis model assessment of insulin resistance (HOMA-IR) index, and weight. Studies have shown that exosomes are involved in the development of follicles. For instance, follicular fluid-derived exosomes have been reported to regulate follicular dysplasia in PCOS by modulating glycolysis in granulosa cells [35]. Our research showed that the ovaries of PCOS mice treated with adipocyte-derived exosomes had an increased number of CFs and deteriorated CL, indicating that exosomes from adipocytes affect follicle growth and development. In addition, DHEA-induced PCOS mice had increased serum T and E2 levels and decreased FSH levels, and adipocyte-derived exosomes further exacerbated these changes in sex hormones. However, the exact mechanism remains unknown and needs more research.
As a highly conserved miRNA, miR-30c is considered a multifunctional regulator of cell proliferation, differentiation, and metabolism [36]. A previous study indicated that mesenchymal stem cell-derived exosomes promote angiogenesis by transferring miR-30c [37]. In our research, miR-30c-5p levels were higher in ovaries of PCOS mice. Inhibition of miR-30c-5p alleviated the effects of adipocyte-derived exosomes on the biological behaviour and VEGFA expression in HOMECs as well as endocrine alterations and ovarian angiogenesis in PCOS mice, indicating adipocyte-derived exosomes exert the above effects through miR-30c-5p. However, miR-30c-5p did not significantly affect the estrous cycle of PCOS mice, possibly because there exist other mechanisms regulating estrous cycle.
SOCS is a protein family of eight members (SOCS1–7 and cytokine-induced SH2 protein) that inhibit STAT activation by suppressing the Janus kinase (JAK)/STAT pathway [38]. After cytokines such as interleukin-6 (IL-6) bind to their receptors, receptor-associated JAK proteins are activated via transphosphorylation. JAK activation triggers phosphorylation of tyrosine on the cytoplasmic tail subunit of docking site receptor of the STAT protein. This phosphorylation results in phosphorylation and dimerization of STAT proteins and translocation of dimers to the nucleus, where they integrate with DNA and activate the transcription of response genes such as cyclins, antiapoptotic proteins, VEGF, matrix metalloproteinases and immunosuppressive proteins [39–41]. As a negative regulator of angiogenesis, SOCS3 contains a short N-terminal kinase inhibitory region like that of JAK substrates, which enables them to inhibit signalling by directly inhibiting the catalytic activity of JAKs [42, 43] (Figure 9). Previous studies have revealed that SOCS3 knockdown results in greater phosphorylation and activation of STAT3, thereby upregulating VEGFA and promoting angiogenesis [29, 30]. In addition, a study has reported downregulation of SOCS3 and upregulation of p-STAT3 in ovaries of PCOS rats, which is consistent with our findings in PCOS mice [44]. Besides, our study also showed that adipocyte-derived exosomal miR-30c-5p activated the STAT3/VEGFA pathway in vivo and in vitro by targeting SOCS3. Altogether, we believe adipocyte-derived exosomal miR-30c-5p regulates ovarian angiogenesis in PCOS via the SOCS3/STAT3/VEGFA pathway.
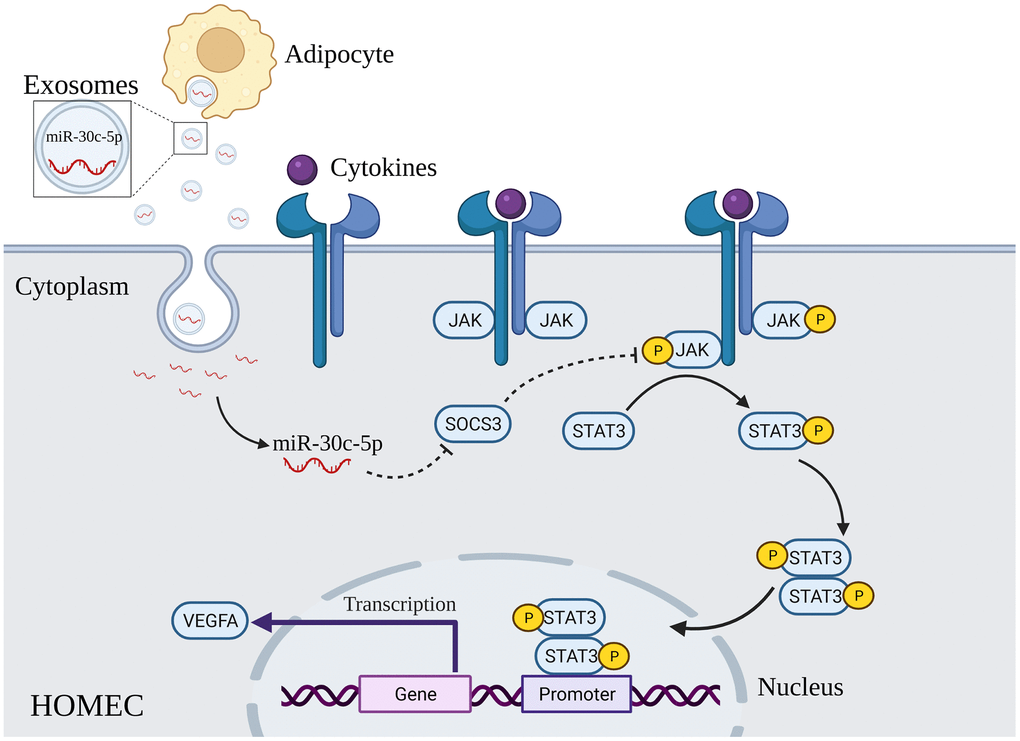
Figure 9. Schematic illustration of adipocyte-derived exosomal miR-30c-5p activating the STAT3/VEGFA pathway by targeting SOCS3 in HOMECs. Exosomes derived from adipocytes promote the activation of the STAT3/VEGFA pathway in HOMECs by transporting miR-30c-5p to directly inhibit SOCS3 expression at the post-transcriptional level.
However, this study has some limitations. First, cells used in this study were obtained from cell lines, which cannot completely simulate the state of specific cells in patients with PCOS. Second, we did not examine the effects of adipocyte-derived exosomes on healthy mice. Third, there might be other exosomal contents mediating the biological effects of adipocyte-derived exosomes. Nevertheless, to the best of our knowledge, this study is the first to examine the role of adipocyte-derived exosomes in the development of PCOS. Hence, this study is of great significance and provides novel insights into the role of exosomes in PCOS. In the future, we plan to conduct more studies to overcome the abovementioned limitations and discover more detailed mechanisms of adipocyte-derived exosomes participating in the development of PCOS.
In conclusion, adipocyte-derived exosomal miR-30c-5p promotes angiogenesis and activates the STAT3/VEGFA pathway by targeting SOCS3, thus contributing to abnormal ovarian angiogenesis in PCOS. This intercellular communication regulated by exosomes may provide new ideas for developing individualized diagnostic and therapeutic strategies for PCOS.
Materials and Methods
Cell line and culture
Preadipocyte 3T3-L1 cells were obtained from the American Type Culture Collection (ATCC) and cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco) containing 10% fetal bovine serum (FBS, 10099141c, Gibco) and 1% penicillin–streptomycin (Biosharp). HOMECs were obtained from Zhongqiaoxinzhou Biotechnology Co. and cultured in Endothelial Cell Medium (1001, Zhongqiaoxinzhou Biotechnology Co.) containing 5% fetal bovine serum, 1% endothelial cell growth supplement and 1% penicillin–streptomycin. All cells were cultivated in a 5% CO2 incubator at 37° C.
Adipogenic differentiation
For adipogenic differentiation, 2 days after confluence (day 0), 3T3-L1 cells were incubated in DMEM containing 10% FBS, 1 μM dexamethasone (D4902, Sigma-Aldrich), 0.5 mM 1-methyl-3-isobutylxanthine (I7018, Sigma-Aldrich) and 10 μg/mL insulin (I5500, Sigma-Aldrich) for 4 days. On day 4, the medium was replaced with DMEM containing 10% FBS and 10 μg/mL insulin for 2 days. Afterwards, the cells were maintained in DMEM containing 10% FBS until day 12. All medium was changed every 2 days.
On day 12, differentiated 3T3-L1 cells were washed twice with PBS and fixed in 4% paraformaldehyde for 20 min. Subsequently, the cells were stained with 60% oil red O (3:2, oil red O saturated solution in isopropanol:distilled water) for 30 min at room temperature. The stained cells were photographed using an inverted microscope (IX51, Olympus).
Exosome isolation and identification
On day 12 of adipogenic differentiation, the medium was replaced with DMEM containing 10% exosome-depleted FBS (EXO-FBS-50A-1, System Biosciences), where differentiated 3T3-L1 cells were cultured for 48 h. The supernatant was collected and centrifuged at 1,000 ×g for 10 min, 3,000 ×g for 30 min and 10,000×g for 20 min at 4° C to separate cell components and fragments. The supernatant from the final centrifugation was ultracentrifuged at 100,000×g for 1 h at 4° C, and then the exosome pellet was resuspended in PBS and centrifuged at 100,000×g for another 1 h at 4° C [45]. Thereafter, exosomes were resuspended in PBS for further use.
For exosome identification, TEM (HT77000, Hitachi) was applied to determine the morphological features of exosomes. In addition, western blotting was used for detecting the specific exosome markers CD9, CD63 and FABP4. Meanwhile, NTA was used for measuring the particle size and concentration of exosomes using the ZetaView system (PMX 110, Particle Metrix).
In vitro exosome uptake assay
Exosomes were labelled with the red fluorescence dye PKH26 (MX4021, MKbio) according to the manufacturer’s instructions. HOMECs were cultured with PKH26-labelled exosomes for 6 h in shade and fixed in 4% paraformaldehyde for 30 min. The cytoskeletons of cells were stained with fluorescein isothiocyanate (FITC)-labelled phalloidin (40735ES75, YEASEN), and nuclei were stained with 4’,6-diamidino-2-phenylindole (DAPI, AS1075, ASPEN). The images were captured using an inverted fluorescent microscope (IX51, Olympus).
Cell counting kit-8 assay
Cells in the logarithmic growth phase were suspended in serum-free medium, seeded in 96-well plates at a density of 1×103 cells/well and cultured overnight. The serum-free medium was replaced with a medium containing 10% exosome-depleted FBS on the following day, and the cells were incubated for 0, 12, 24, 48 and 96 h. Subsequently, the CCK-8 (C0038, Beyotime) reagent was added, and then the cells were placed in an incubator at 37° C for 2 h. The optical density value was recorded on a microplate reader (Ensight, Perkin Elmer) at a wavelength of 450 nm.
Transwell assay
Transwell assay was performed in transwell chambers with a pore size of 8.0 μm (3422, Corning). Briefly, 2 × 104 HOMECs suspended in 200 μL of serum-free medium were added to the upper chamber, and 600 μL of medium containing 10% exosome-depleted FBS was added to the lower chamber. The cells were incubated at 37° C for 24 h. Subsequently, the medium was removed, and 4% paraformaldehyde was used to fix the cells for 30 min. The cells were stained with 0.1% (w/v) crystal violet for 10 min, and the migrated cells were visualized and imaged using an optical microscope.
Wound healing assay
For wound healing assay, 2 × 105 cells/well were seeded in a 6-well plate and grown to confluence. The monolayer was scratched using a pipette tip, and then the detached cells were removed using serum-free medium. Thereafter, the cells were cultured in medium supplemented with 10% exosome-depleted FBS at 37° C for 24 h and photographed at 0 h and 24 h after the scratch. Wound closure was calculated as follows: wound recovery area (%) = (A0 – A1)/A0 × 100, where A0 represents the initial wound area, and A1 represents the wound area at the time of measurement.
Tube formation assay
Tube formation assay was performed for determining the capillary-like structure formation capacity of HOMECs. Thawed Matrigel matrix (354248, Corning) was added to a 96-well plate and incubated at 37° C for 1 h. Thereafter, the cells were seeded in a 96-well plate at a density of 2 × 104 cells/well and incubated at 37° C. After 6 h, capillary-like structure formation was observed using an inverted microscope.
Coculture
A transwell system (3412, Corning) was used for coculture of differentiated adipocytes and HOMECs. In the coculture experiments, differentiated adipocytes were cultured in the upper chambers, and GW4869 (MedChemExpress) was added to block the secretion of exosomes from adipocytes. Then HOMECs were seeded in the bottom chambers and cocultured with adipocytes for 48 h. qRT-PCR was used to detect the levels of miR-30c-5p in HOMECs.
Cell transfection
miR-30c-5p mimics, inhibitors and the corresponding NCs were obtained from RiboBio Co.. Cells were cultured in a 6-well plate until 70% confluence was achieved. The transfection reagent was prepared by adding the miRNA mimics, inhibitors, or NCs and Lipofectamine 2000 (Lipo 2000, Invitrogen) to serum-free medium, which was incubated at room temperature for 15 min. Subsequently, the reagent was added to each group of cells and incubated at 37° C for 24 h. Transfected cells were treated with fresh medium for 4–6 h before further analyses [46].
The SOCS3 plasmid (pEnCMV-SOCS3[human]-6×His-SV40-Neo) and control vector (pEnCMV-MCS-6×His-SV40-Neo) were obtained from MiaoLing Biotechnology Co.. The transfection method mentioned above was also applied for plasmid transfection and co-transfection of miR-30c-5p mimics with the SOCS3 plasmid.
Bioinformatic analysis
The target genes of miR-30c-5p were jointly predicted using TargetScan (http://www.targetscan.org/vert_71/), Starbase (https://starbase.sysu.Edu.cn/index.php) and miRDB (http://mirdb.org/). Evenn (http://www.ehbio.com/test/venn/#/) was used to combine the data and determine potential target genes. TargetScan was used to identify the seed sequences and regions of potential base-pairing of the target gene.
Dual luciferase reporter assay
The 3’-UTR of the SOCS3 gene was cloned into a dual luciferase reporter vector (Genepharma), referred to as pmirGLO-SOCS3-WT, and a mutant type of the SOCS3 3’-UTR reporter plasmid (pmirGLO-SOCS3-MUT) was also generated by altering the seed region. pmirGLO-SOCS3-WT/MUT was co-transfected with miR-30c-5p mimics into HOMECs using Lipofectamine 2000 for 48 h. Finally, the dual luciferase reporter gene assay kit (Genepharma) was used to measure the binding activity of miR-30c-5p based on firefly/renilla luciferase activity.
Animals and experimental protocol
Female C57BL/6J mice (N=42, 21 days old) were raised at a controlled temperature between 22 and 24° C, with free access to food and water. The PCOS model group (N=32) was subcutaneously injected with dehydroepiandrosterone (DHEA) (6 mg/100 g weight in sesame oil) daily for 21 days to induce PCOS [47]. The control group (CON, N=10) was injected with sesame oil. The establishment of PCOS models was determined by weight, estrous cycle, and ovarian morphology. DHEA-induced PCOS mice were divided into the following three groups: PCOS control group(N=10), PCOS+EXO group (N=12) and PCOS+EXO+miR-30c-5p antagomir group (N=10). Exosomes (30 μg in 200 μL PBS) and miR-30c-5p antagomir (10 mg/kg) were injected into PCOS mice twice a week via the tail vein for 2 weeks [48, 49]. During the treatment period, the mice were weighed weekly, and the estrous cycle was assessed daily via vaginal smears from the 10th day of the first DHEA injection until the end of the experiment. At the end of the treatment period, fasting blood glucose levels were measured via the tail vein after 12 h of fasting using a blood glucose meter (Sinocare). Thereafter, the blood of all mice was collected from the heart using a 1 mL syringe under isoflurane anaesthesia. Serum samples were collected via centrifugation and stored at −80° C for further analyses, and the ovaries were removed for conducting additional experiments. All protocols and experiments were approved by the Animal Care and Use Committee of Renmin Hospital of Wuhan University.
In vivo exosome uptake assay
For verification of adipocyte-derived exosomes being taken up by ovaries, PKH26-labeled exosomes were injected into PCOS mice via tail vein. After 24h, the mice were sacrificed, and the ovaries were collected for further IF analyses. For IF staining, the harvested sections were incubated with the primary antibody CD31 (sc-376764, Santa Cruz, 1:100). Subsequently, the sections were incubated with Alexa Fluor 488-conjugated secondary antibody and stained with DAPI for visualization. The fluorescence expression of CD31, DAPI and PKH26-labelled exosomes was determined using a fluorescence microscope.
Ovarian morphology
The ovaries were incubated with paraformaldehyde, dehydrated with ethanol of different concentrations, and embedded with paraffin wax. The maximum cross-section of the ovaries was cut and stained with haematoxylin and eosin (HE). The follicles were classified as primary follicles (PFs) with one layer of cubical granulosa cells (GCs), secondary follicles (SFs) with two or more layers of GCs and antral follicles (AFs) based on the presence or absence of an antrum and CL. CFs contain an attenuated GC layer and thickened theca interna cell layer with a large antrum or a large fluid-filled structure [50].
Elisa
The levels of VEGFA, E2, T, FSH and insulin in the serum were measured using sandwich ELISA kits (Servicebio, Wuhan, China) according to the manufacturer’s instructions. Each sample was measured in duplicate.
Immunohistochemical
For IHC staining, tissue sections were deparaffinized and rehydrated with xylene and ethanol. The endogenous peroxidase activity was blocked using H2O2 in PBS, and non-specific binding was blocked using 5% BSA. Subsequently, the sections were incubated with biotinylated IgG (ASPEN), followed by incubation with the primary antibodies CD31 (#92841, CST, 1:200), VEGFA (ab52917, Abcam, 1:100) and SOCS3 (ab280884, Abcam, 1:1000). Finally, the sections were visualized using the chromogenic substrate diaminobenzidine, and an optical microscope (BX51, Olympus) was used to capture images of the stained sections.
qRT-PCR analysis
Total RNA in cells and the ovaries was isolated using the TRIzol reagent (Life Technology), and cDNA for miRNAs was synthesised using the miRNA First Strand cDNA Synthesis kit (Sangon) according to the manufacturer’s instructions. qRT-PCR was performed using a miRNA qPCR Kit (Sangon) with the forward primer and the universal reverse primer of miR-30c-5p (Sangon). Gene expression was evaluated using the 2-ΔΔCT method [51], and U6 was used to normalize the relative expression of miR-30c-5p. The primer sequences used are as follows: miR-30c-5p forward primer, 5'-GCTGCGTGTAAACATCCTACACT-3'; reverse primer, 5'-AGTGCAGGGTCCGAGGTATT-3' and U6 forward primer, 5'- CTCGCTTCGGCAGCACA-3'; reverse primer, 5'- AACGCTTCACGAATTTGCGT -3'.
Western blotting analysis
Cell and tissue proteins were extracted using Radio Immunoprecipitation Assay (RIPA) buffer (ASPEN). The concentrations of proteins were detected by a BCA Protein Assay Kit (P0010, Beyotime) according to the manufactures’ instructions. Protein extracts were separated via sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene fluoride (PVDF) membranes. The membranes were blocked with 5% milk in Tris-buffered saline containing 0.1% Tween-20 for 1 h at room temperature. Subsequently, the membranes were incubated with primary antibodies at 4° C overnight, followed by incubation with horseradish peroxidase (HRP)-labelled secondary antibodies at 37° C for 1 h. The antibodies included rabbit anti-CD9 (ab92726, Abcam, 1:1000), mouse anti-CD63 (sc-5275, Santa Cruz, 1:500), rabbit anti-FABP4 (ab92501, Abcam, 1:2000), rabbit anti-Ki67 (ab16667, Abcam, 1:500), rabbit anti-PCNA (#13110, CST, 1:1000), rabbit anti-Vimentin (#5741, CST, 1:500), mouse anti-E-cadherin (#14472, CST, 1:500), rabbit anti-VEGFA (ab46154, Abcam, 1:1000), rabbit anti-SOCS3 (ab280884, Abcam, 1:1000), rabbit anti-STAT3 (sc-8019, Santa Cruz, 1:500), rabbit anti-p-STAT3 (sc-8059, Santa Cruz, 1:500), HRP-labelled goat anti-rabbit (ASPEN) and HRP-labelled goat anti-mouse (ASPEN) antibodies. Rabbit anti-GAPDH (ab37168, Abcam, 1:10000) and rabbit anti-β-actin (ab8227, Abcam, 1:5000) were used as the internal reference. The immunoreactive bands were visualized on an ECL-Plus western blotting detection system (Thermo Fisher Scientific). The ratio of the grey value of the target band to the internal reference band was used to assess the relative expression of the protein.
Statistical analysis
All data were analyzed using the SPSS Statistics (version 26.0) (IBM) and Prism 9 (GraphPad) software. Data were expressed as mean ± standard deviation (SD) of at least three independent experiments. Student’s t-test was used to compare data between two groups, and one-way ANOVA followed by Tukey’s multiple comparison test was used to compare data among multiple groups. Statistical significance was established at *P < 0.05 versus the indicated group.
Data availability
All data generated or analyzed during this study are included in this published article.
Supplementary Materials
Author Contributions
JH and FYL designed the study. JH and FYL performed the experiments and drafted the manuscript. JH, FYL, YCY and YJS participated in data analysis. JH, FYL, ZNX and WMX were involved in the discussion and interpretation of results. ZNX and WMX organized the whole project. All authors have read and approved the final manuscript.
Acknowledgments
We thank all participants of this study. Our deepest gratitude also goes to the editor and anonymous reviewers for their careful work and thoughtful suggestions that have helped improve this paper substantially.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement and Consent
All animal protocols were approved (approval no. WDRM-20210612) by the Animal Care and Use Committee of Renmin Hospital of Wuhan University and conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Funding
This study was supported by the National Natural Science Foundation of China (Grant No.: 81471455 and 81100418).
References
- 1. Escobar-Morreale HF. Polycystic ovary syndrome: definition, aetiology, diagnosis and treatment. Nat Rev Endocrinol. 2018; 14:270–84. https://doi.org/10.1038/nrendo.2018.24 [PubMed]
- 2. Rotterdam ESHRE/ASRM-Sponsored PCOS consensus workshop group. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome (PCOS). Hum Reprod. 2004; 19:41–7. https://doi.org/10.1093/humrep/deh098 [PubMed]
- 3. Stapor P, Wang X, Goveia J, Moens S, Carmeliet P. Angiogenesis revisited - role and therapeutic potential of targeting endothelial metabolism. J Cell Sci. 2014; 127:4331–41. https://doi.org/10.1242/jcs.153908 [PubMed]
- 4. Duncan WC, Nio-Kobayashi J. Targeting angiogenesis in the pathological ovary. Reprod Fertil Dev. 2013; 25:362–71. https://doi.org/10.1071/RD12112 [PubMed]
- 5. Dwivedi AN, Ganesh V, Shukla RC, Jain M, Kumar I. Colour Doppler evaluation of uterine and ovarian blood flow in patients of polycystic ovarian disease and post-treatment changes. Clin Radiol. 2020; 75:772–9. https://doi.org/10.1016/j.crad.2020.05.023 [PubMed]
- 6. Alcázar JL, Kudla MJ. Ovarian stromal vessels assessed by spatiotemporal image correlation-high definition flow in women with polycystic ovary syndrome: a case-control study. Ultrasound Obstet Gynecol. 2012; 40:470–5. https://doi.org/10.1002/uog.11187 [PubMed]
- 7. Di Pietro M, Pascuali N, Parborell F, Abramovich D. Ovarian angiogenesis in polycystic ovary syndrome. Reproduction. 2018; 155:R199–209. https://doi.org/10.1530/REP-17-0597 [PubMed]
- 8. Kamat BR, Brown LF, Manseau EJ, Senger DR, Dvorak HF. Expression of vascular permeability factor/vascular endothelial growth factor by human granulosa and theca lutein cells. Role in corpus luteum development. Am J Pathol. 1995; 146:157–65. [PubMed]
- 9. Dambala K, Vavilis D, Bili E, Goulis DG, Tarlatzis BC. Serum visfatin, vascular endothelial growth factor and matrix metalloproteinase-9 in women with polycystic ovary syndrome. Gynecol Endocrinol. 2017; 33:529–33. https://doi.org/10.1080/09513590.2017.1296425 [PubMed]
- 10. Artini PG, Monti M, Matteucci C, Valentino V, Cristello F, Genazzani AR. Vascular endothelial growth factor and basic fibroblast growth factor in polycystic ovary syndrome during controlled ovarian hyperstimulation. Gynecol Endocrinol. 2006; 22:465–70. https://doi.org/10.1080/09513590600906607 [PubMed]
- 11. Elci E, Kaya C, Cim N, Yildizhan R, Elci GG. Evaluation of cardiac risk marker levels in obese and non-obese patients with polycystic ovaries. Gynecol Endocrinol. 2017; 33:43–7. https://doi.org/10.1080/09513590.2016.1203893 [PubMed]
- 12. Agrawal R, Conway G, Sladkevicius P, Tan SL, Engmann L, Payne N, Bekir J, Campbell S, Jacobs H. Serum vascular endothelial growth factor and Doppler blood flow velocities in in vitro fertilization: relevance to ovarian hyperstimulation syndrome and polycystic ovaries. Fertil Steril. 1998; 70:651–8. https://doi.org/10.1016/s0015-0282(98)00249-0 [PubMed]
- 13. Rashid R, Mir SA, Kareem O, Ali T, Ara R, Malik A, Amin F, Bader GN. Polycystic ovarian syndrome-current pharmacotherapy and clinical implications. Taiwan J Obstet Gynecol. 2022; 61:40–50. https://doi.org/10.1016/j.tjog.2021.11.009 [PubMed]
- 14. de Medeiros SF, Rodgers RJ, Norman RJ. Adipocyte and steroidogenic cell cross-talk in polycystic ovary syndrome. Hum Reprod Update. 2021; 27:771–96. https://doi.org/10.1093/humupd/dmab004 [PubMed]
- 15. Pegtel DM, Gould SJ. Exosomes. Annu Rev Biochem. 2019; 88:487–514. https://doi.org/10.1146/annurev-biochem-013118-111902 [PubMed]
- 16. Lanthier N, Leclercq IA. Adipose tissues as endocrine target organs. Best Pract Res Clin Gastroenterol. 2014; 28:545–58. https://doi.org/10.1016/j.bpg.2014.07.002 [PubMed]
- 17. Yu Y, Du H, Wei S, Feng L, Li J, Yao F, Zhang M, Hatch GM, Chen L. Adipocyte-Derived Exosomal MiR-27a Induces Insulin Resistance in Skeletal Muscle Through Repression of PPARγ. Theranostics. 2018; 8:2171–88. https://doi.org/10.7150/thno.22565 [PubMed]
- 18. Sano S, Izumi Y, Yamaguchi T, Yamazaki T, Tanaka M, Shiota M, Osada-Oka M, Nakamura Y, Wei M, Wanibuchi H, Iwao H, Yoshiyama M. Lipid synthesis is promoted by hypoxic adipocyte-derived exosomes in 3T3-L1 cells. Biochem Biophys Res Commun. 2014; 445:327–33. https://doi.org/10.1016/j.bbrc.2014.01.183 [PubMed]
- 19. Song Y, Li H, Ren X, Li H, Feng C. SNHG9, delivered by adipocyte-derived exosomes, alleviates inflammation and apoptosis of endothelial cells through suppressing TRADD expression. Eur J Pharmacol. 2020; 872:172977. https://doi.org/10.1016/j.ejphar.2020.172977 [PubMed]
- 20. Thomou T, Mori MA, Dreyfuss JM, Konishi M, Sakaguchi M, Wolfrum C, Rao TN, Winnay JN, Garcia-Martin R, Grinspoon SK, Gorden P, Kahn CR. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature. 2017; 542:450–5. https://doi.org/10.1038/nature21365 [PubMed]
- 21. Irani S, Hussain MM. Role of microRNA-30c in lipid metabolism, adipogenesis, cardiac remodeling and cancer. Curr Opin Lipidol. 2015; 26:139–46. https://doi.org/10.1097/MOL.0000000000000162 [PubMed]
- 22. Moghiman T, Barghchi B, Esmaeili SA, Shabestari MM, Tabaee SS, Momtazi-Borojeni AA. Therapeutic angiogenesis with exosomal microRNAs: an effectual approach for the treatment of myocardial ischemia. Heart Fail Rev. 2021; 26:205–13. https://doi.org/10.1007/s10741-020-10001-9 [PubMed]
- 23. Karbiener M, Neuhold C, Opriessnig P, Prokesch A, Bogner-Strauss JG, Scheideler M. MicroRNA-30c promotes human adipocyte differentiation and co-represses PAI-1 and ALK2. RNA Biol. 2011. https://doi.org/10.4161/rna.8.5.16153 [PubMed]
- 24. Bridge G, Monteiro R, Henderson S, Emuss V, Lagos D, Georgopoulou D, Patient R, Boshoff C. The microRNA-30 family targets DLL4 to modulate endothelial cell behavior during angiogenesis. Blood. 2012; 120:5063–72. https://doi.org/10.1182/blood-2012-04-423004 [PubMed]
- 25. Long W, Zhao C, Ji C, Ding H, Cui Y, Guo X, Shen R, Liu J. Characterization of serum microRNAs profile of PCOS and identification of novel non-invasive biomarkers. Cell Physiol Biochem. 2014; 33:1304–15. https://doi.org/10.1159/000358698 [PubMed]
- 26. Ferrante SC, Nadler EP, Pillai DK, Hubal MJ, Wang Z, Wang JM, Gordish-Dressman H, Koeck E, Sevilla S, Wiles AA, Freishtat RJ. Adipocyte-derived exosomal miRNAs: a novel mechanism for obesity-related disease. Pediatr Res. 2015; 77:447–54. https://doi.org/10.1038/pr.2014.202 [PubMed]
- 27. Wang F, Chen FF, Shang YY, Li Y, Wang ZH, Han L, Li YH, Zhang L, Ti Y, Zhang W, Zhong M. Insulin resistance adipocyte-derived exosomes aggravate atherosclerosis by increasing vasa vasorum angiogenesis in diabetic ApoE-/- mice. Int J Cardiol. 2018; 265:181–7. https://doi.org/10.1016/j.ijcard.2018.04.028 [PubMed]
- 28. Gao XR, Ge J, Li WY, Zhou WC, Xu L, Geng DQ. miR-34a carried by adipocyte exosomes inhibits the polarization of M1 macrophages in mouse osteolysis model. J Biomed Mater Res A. 2021; 109:994–1003. https://doi.org/10.1002/jbm.a.37088 [PubMed]
- 29. Sun Y, Ju M, Lin Z, Fredrick TW, Evans LP, Tian KT, Saba NJ, Morss PC, Pu WT, Chen J, Stahl A, Joyal JS, Smith LE. SOCS3 in retinal neurons and glial cells suppresses VEGF signaling to prevent pathological neovascular growth. Sci Signal. 2015; 8:ra94. https://doi.org/10.1126/scisignal.aaa8695 [PubMed]
- 30. Yan Z, Hong S, Song Y, Bi M. microR-4449 Promotes Colorectal Cancer Cell Proliferation via Regulation of SOCS3 and Activation of STAT3 Signaling. Cancer Manag Res. 2021; 13:3029–39. https://doi.org/10.2147/CMAR.S266153 [PubMed]
- 31. Crewe C, Joffin N, Rutkowski JM, Kim M, Zhang F, Towler DA, Gordillo R, Scherer PE. An Endothelial-to-Adipocyte Extracellular Vesicle Axis Governed by Metabolic State. Cell. 2018; 175:695–708.e13. https://doi.org/10.1016/j.cell.2018.09.005 [PubMed]
- 32. Apte RS, Chen DS, Ferrara N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell. 2019; 176:1248–64. https://doi.org/10.1016/j.cell.2019.01.021 [PubMed]
- 33. Song M, Han L, Chen FF, Wang D, Wang F, Zhang L, Wang ZH, Zhong M, Tang MX, Zhang W. Adipocyte-Derived Exosomes Carrying Sonic Hedgehog Mediate M1 Macrophage Polarization-Induced Insulin Resistance via Ptch and PI3K Pathways. Cell Physiol Biochem. 2018; 48:1416–32. https://doi.org/10.1159/000492252 [PubMed]
- 34. Gao J, Li X, Wang Y, Cao Y, Yao D, Sun L, Qin L, Qiu H, Zhan X. Adipocyte-derived extracellular vesicles modulate appetite and weight through mTOR signalling in the hypothalamus. Acta Physiol (Oxf). 2020; 228:e13339. https://doi.org/10.1111/apha.13339 [PubMed]
- 35. Cao J, Huo P, Cui K, Wei H, Cao J, Wang J, Liu Q, Lei X, Zhang S. Follicular fluid-derived exosomal miR-143-3p/miR-155-5p regulate follicular dysplasia by modulating glycolysis in granulosa cells in polycystic ovary syndrome. Cell Commun Signal. 2022; 20:61. https://doi.org/10.1186/s12964-022-00876-6 [PubMed]
- 36. Han W, Cui H, Liang J, Su X. Role of MicroRNA-30c in cancer progression. J Cancer. 2020; 11:2593–601. https://doi.org/10.7150/jca.38449 [PubMed]
- 37. Gong M, Yu B, Wang J, Wang Y, Liu M, Paul C, Millard RW, Xiao DS, Ashraf M, Xu M. Mesenchymal stem cells release exosomes that transfer miRNAs to endothelial cells and promote angiogenesis. Oncotarget. 2017; 8:45200–12. https://doi.org/10.18632/oncotarget.16778 [PubMed]
- 38. Carow B, Rottenberg ME. SOCS3, a Major Regulator of Infection and Inflammation. Front Immunol. 2014; 5:58. https://doi.org/10.3389/fimmu.2014.00058 [PubMed]
- 39. Sims NA. The JAK1/STAT3/SOCS3 axis in bone development, physiology, and pathology. Exp Mol Med. 2020; 52:1185–97. https://doi.org/10.1038/s12276-020-0445-6 [PubMed]
- 40. Gao Y, Zhao H, Wang P, Wang J, Zou L. The roles of SOCS3 and STAT3 in bacterial infection and inflammatory diseases. Scand J Immunol. 2018; 88:e12727. https://doi.org/10.1111/sji.12727 [PubMed]
- 41. Zou S, Tong Q, Liu B, Huang W, Tian Y, Fu X. Targeting STAT3 in Cancer Immunotherapy. Mol Cancer. 2020; 19:145. https://doi.org/10.1186/s12943-020-01258-7 [PubMed]
- 42. Rao N, Lee YF, Ge R. Novel endogenous angiogenesis inhibitors and their therapeutic potential. Acta Pharmacol Sin. 2015; 36:1177–90. https://doi.org/10.1038/aps.2015.73 [PubMed]
- 43. Sasaki A, Yasukawa H, Suzuki A, Kamizono S, Syoda T, Kinjyo I, Sasaki M, Johnston JA, Yoshimura A. Cytokine-inducible SH2 protein-3 (CIS3/SOCS3) inhibits Janus tyrosine kinase by binding through the N-terminal kinase inhibitory region as well as SH2 domain. Genes Cells. 1999; 4:339–51. https://doi.org/10.1046/j.1365-2443.1999.00263.x [PubMed]
- 44. Zhou Y, Lv L, Liu Q, Song J. Total flavonoids extracted from Nervilia Fordii function in polycystic ovary syndrome through IL-6 mediated JAK2/STAT3 signaling pathway. Biosci Rep. 2019; 39:BSR20181380. https://doi.org/10.1042/BSR20181380 [PubMed]
- 45. Wang J, Wu Y, Guo J, Fei X, Yu L, Ma S. Adipocyte-derived exosomes promote lung cancer metastasis by increasing MMP9 activity via transferring MMP3 to lung cancer cells. Oncotarget. 2017; 8:81880–91. https://doi.org/10.18632/oncotarget.18737 [PubMed]
- 46. Lin FY, Han ST, Yu WM, Rao T, Ruan Y, Yuan R, Li HY, Ning JZ, Xia YQ, Xie JN, Qi YC, Zhou XJ, Cheng F. microRNA-486-5p is implicated in the cisplatin-induced apoptosis and acute inflammation response of renal tubular epithelial cells by targeting HAT1. J Biochem Mol Toxicol. 2022; 36:e23039. https://doi.org/10.1002/jbt.23039 [PubMed]
- 47. Xie Q, Xiong X, Xiao N, He K, Chen M, Peng J, Su X, Mei H, Dai Y, Wei D, Lin G, Cheng L. Mesenchymal Stem Cells Alleviate DHEA-Induced Polycystic Ovary Syndrome (PCOS) by Inhibiting Inflammation in Mice. Stem Cells Int. 2019; 2019:9782373. https://doi.org/10.1155/2019/9782373 [PubMed]
- 48. Deng ZB, Poliakov A, Hardy RW, Clements R, Liu C, Liu Y, Wang J, Xiang X, Zhang S, Zhuang X, Shah SV, Sun D, Michalek S, et al. Adipose tissue exosome-like vesicles mediate activation of macrophage-induced insulin resistance. Diabetes. 2009; 58:2498–505. https://doi.org/10.2337/db09-0216 [PubMed]
- 49. Wang X, Guo B, Li Q, Peng J, Yang Z, Wang A, Li D, Hou Z, Lv K, Kan G, Cao H, Wu H, Song J, et al. miR-214 targets ATF4 to inhibit bone formation. Nat Med. 2013; 19:93–100. https://doi.org/10.1038/nm.3026 [PubMed]
- 50. Mao Z, Yang L, Lu X, Tan A, Wang Y, Ding F, Xiao L, Qi X, Yu Y. C1QTNF3 in the murine ovary and its function in folliculogenesis. Reproduction. 2018; 155:333–46. https://doi.org/10.1530/REP-17-0783 [PubMed]
- 51. Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008; 3:1101–8. https://doi.org/10.1038/nprot.2008.73 [PubMed]