


Introduction
Bone fractures are the most common musculoskeletal condition requiring hospitalization in Medicare enrollees aged 65 and older [1–4]. Furthermore, this demographic has been shown to have a decreased capacity to heal from fractures, leading to higher odds than the general population of requiring re-hospitalization due to delayed healing or nonunion of a bone fracture. Complications stemming from inadequate healing can result in more severe and systemic consequences for this population, such as incapacitation [1, 5, 6].
Normal fracture healing consists of 4 stages – hematoma formation, fibrocartilaginous callus formation, bony callus formation, and bone remodeling. Early healing is accomplished by a robust inflammatory response that recruits skeletal tissue progenitor cells that develop into a soft, cartilaginous, and avascular callus [7, 8]. Vascularization initiates the hardening of the soft callus into eventual bone through endochondral ossification by facilitating infiltration of necessary nutrients and cell types such as chondrocytes and osteoblasts [9]. The last step is characterized by remodeling of the callus through alternating osteoclastic and osteoblastic processes to re-establish normal trabecular bone structure and hematopoiesis [10]. Many age-related changes in this process have been associated with impaired and delayed fracture healing in elderly animals, such as a decrease in the proliferation and differentiation capacity of osteochondral stem cells and a delay in the deposition of cartilage matrix [11–14]. Changes that occur during the vascularization step due to aging are also an established contributor to delayed fracture healing in this population. Furthermore, these changes are potentially the most impactful considering angiogenesis is a key rate-limiting factor for bone repair [15, 16]. Indeed, fracture patients with a pre-existing vascular injury or disease can incur delayed healing or non-union at rates as high as 46%, illustrating the importance of angiogenesis in the healing process [17]. Generally, vascular perfusion of the skeleton decreases with age [18], and elderly rats display significantly decreased patency of bone marrow (BM) blood vessels compared to younger rats [19]. In fracture calluses, young mice have a higher surface density of blood vessels compared to elderly mice, which is partially attributed to an earlier induction of vascular endothelial growth factor (VEGF) and hypoxia-inducible factor (HIF)-1α production [20]. While VEGF and HIF-1α are amongst the most well-known angiogenic molecules, other factors such as fibroblast growth factor (FGF), transforming growth factor (TGF)-β, sirtuin 1 (SIRT1), and bone morphogenetic protein (BMP)-2 have also been characterized as angiogenic molecules that play a role in fracture healing and bone regulation [21–26]. Currently, only a few growth factors such as BMP-2, BMP-7, and platelet-derived growth factor (PDGF) are FDA-approved for assisting in bone formation and bone graft substitutions, with each having their own unique advantages and disadvantages [27, 28]. For example, BMP-2 administration at the fracture callus site has been shown to improve both fracture healing time and callus vascularization in mice but increases the risk of notable side effects such as systemic inflammation, ectopic bone formation, wound complications, and an increased risk of developing cancer [27, 29–32]. While there have been no reports that PDGF and BMP-7 demonstrate this exact side effect profile, PDGF’s clinical use has been limited by its short half-life and oncogenicity, while BMP-7’s use has been limited by its cost [33, 34]. Thus, research investigating alternative treatment options for improved fracture healing and vascularization is ongoing. One avenue of novel therapies that has recently been explored is stimulating endogenous megakaryocytes (MKs) with the main MK growth factor, thrombopoietin (TPO) [35], and other cytokines that MKs produce.
Previous studies have shown that mice that displayed an increased number of BM-residing MKs also displayed an increase in cortical bone thickness and trabecular bone number, and had an increased risk of developing osteosclerosis [36–39]. MKs have been shown to exert these effects on bone structure by mechanisms that involve both direct cell-to-cell contact and paracrine secretion of cytokines [25, 40]. MK’s paracrine effects may influence bone growth and homeostasis through a variety of molecular mechanisms such as increasing osteoblast proliferation, suppressing osteoclastogenesis, and increasing the vessel-forming activity of endothelial cells (ECs) [25, 41]. Their effect on EC activity has thus far been attributed to the production and release of VEGF and TGF-β1 [25, 42]. If the production of these cytokines and other angiogenic growth factors were to be altered in MKs due to aging, this could provide a possible mechanism that contributes to the delayed vascularization of fracture calluses that is seen in elderly populations. MKs from older mice have previously been shown to have a decreased capacity to support osteoblast proliferation [43], but the effects of aging on the ability of MKs to stimulate angiogenesis have not yet been characterized.
To investigate the effects of aging on the angiogenic capacity of MKs through paracrine signaling, we measured the difference in activity of murine BM endothelial cells (BMECs) that were supplemented with media cultured with MKs from young (3–4-month-old) and aged (22–24-month-old) mice for 3 days. The activity of BMECs was measured by their capacity to proliferate, develop vessel-like structures, migrate, and express angiogenic factors.
Results
Proliferation of BMECs when cultured with MK CM
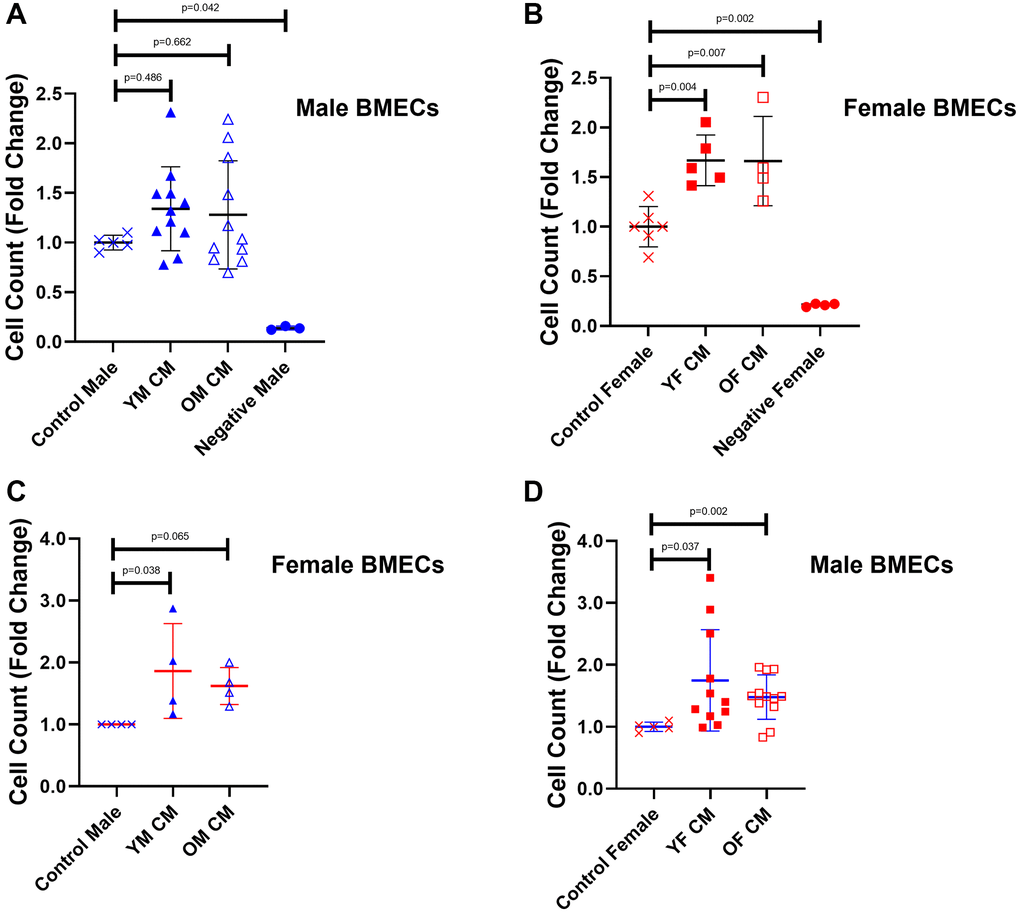
First, we examined cell growth as a proxy for the angiogenic potential of MK conditioned media (CM) on BMECs (Figure 1). While supplementation of young and aged male MK CM did not significantly impact the male BMEC cell number (Figure 1A), female BMEC proliferation was significantly improved with female MK CM, irrespective of age, compared to control female BMECs (Figure 1B). When treated with MK CM generated from mice of the opposite sex, male BMECs exhibited a significant increase in proliferation when cultured with young and old female MK CM (Figure 1D); female BMECs, however, only showed significantly greater proliferation when cultured with young male MK CM (Figure 1C). Together, these results suggest that female MKs secrete factors that increase both male and female BMEC proliferation. Notably, male MK CM generated from young mice significantly increased female BMEC proliferation and male MK CM generated from old mice showed a trending increase in female BMEC, whereas no significant or trending differences were observed when young or aged male MK CM was cultured with male BMECs. It should be noted that throughout this manuscript we report significant and trending differences due to the variability inherent in the use of primary cell cultures.
Effects of MK CM on vessel-like structure formation of BMECs
The effects of young and aged MK CM on the angiogenic potential of BMECs were evaluated by vessel-like structure formation (Figure 2). The criteria for what constitute a vessel-like structure are found in the Methods section. Vessel-like structure formation in the control and treatment groups was assessed and compared by the number of nodes, number of meshes, number of vessel-like structures, and total vessel length. As shown in Figure 2A–2H, treatment with young MK CM significantly improved all parameters in both males and females. Treatment with aged MK CM, irrespective of sex, significantly improved node and mesh numbers (Figure 2A–2D), and aged female MK CM significantly improved the number and length of vessel-like structures (Figure 2F, 2H, 2M–2O). On the other hand, aged male MK CM resulted in a trending, but not statistically significant, increase in the number of vessel-like structures (p = 0.22) and total vessel length (p = 0.14) (Figure 2E, 2G, 2I–2K). When comparing young and aged sex-matched MK CM, a significant difference between young and aged male MK CM on the number of vessel-like structures was detected (p = 0.044), where young male MK CM-treated groups had significantly more vessel-like structures than groups treated with aged male MK CM (Figure 2E). Images of positive control media (5% FBS) cultured with male and female BMECs are found in Figure 2L, 2P, respectively.
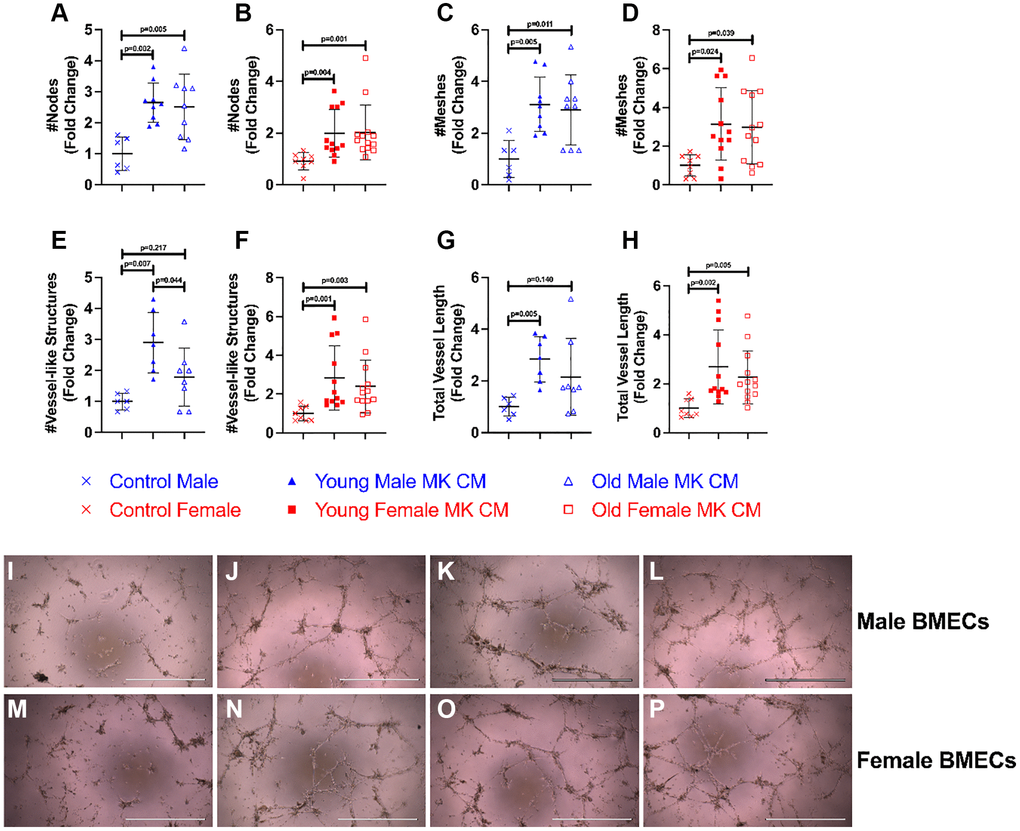
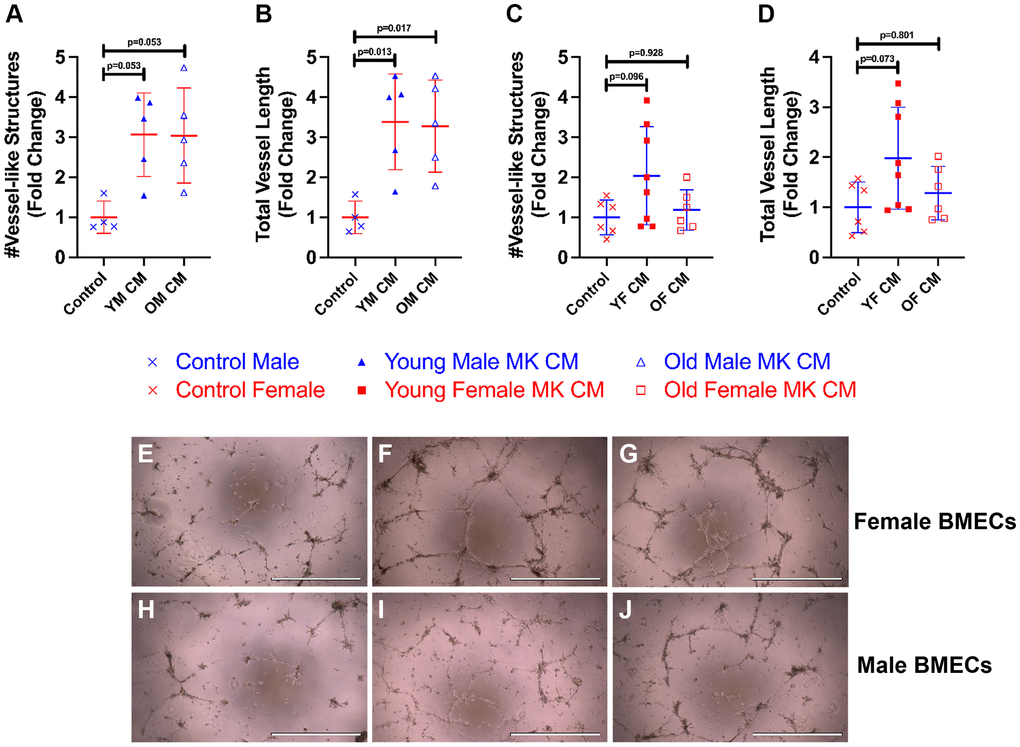
To examine how treatment of MK CM generated from mice of the opposite sex affects male and female BMEC tube formation, we also treated male BMECs with female MK CM from young or old mice and female BMECs with male MK CM from young or old mice (Figure 3). Interestingly, young and old male MK CM increased total vessel length of female BMECs and demonstrated a strong trend toward increasing total number of vessels (Figure 3A, 3B, 3E–3G). However, male BMECs did not respond to old female MK CM but did show a trend toward increased vessel number and length when treated with young female MK CM (Figure 3C, 3D, 3H–3J). Overall, the data in Figures 2 and 3 primarily support the idea that treatment of BMECs with MK CM, especially young MK CM, improves their capacity for vessel-like structure formation irrespective of the sex of the mice from which MKs were generated.
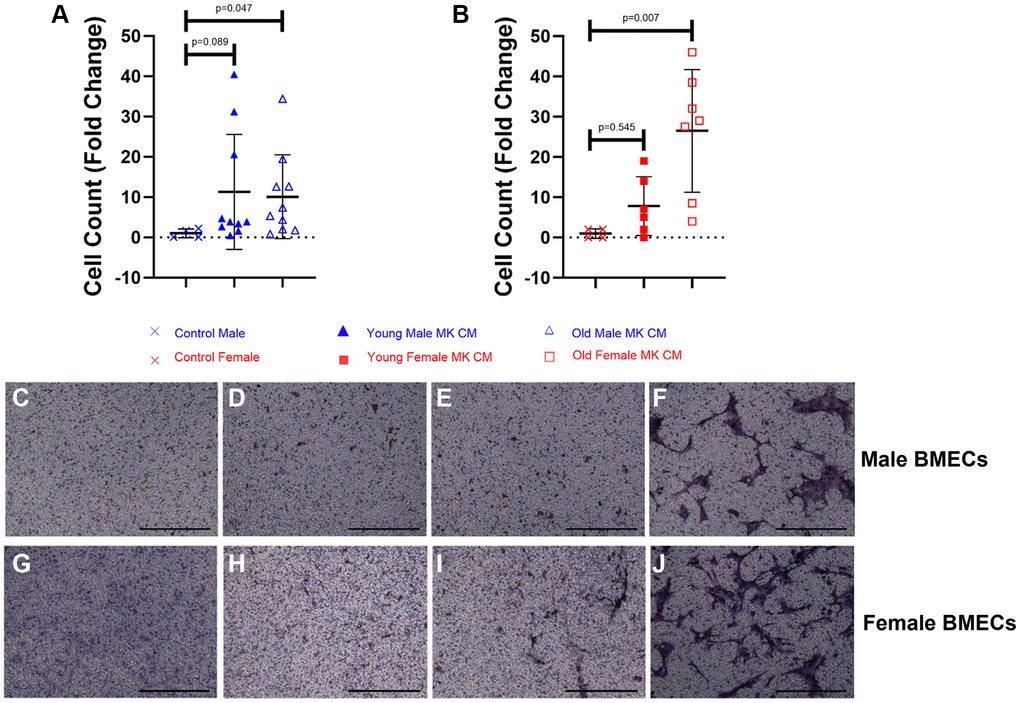
Effects of MK CM on BMEC unidimensional chemotaxis
The impacts of MK CM treatment on BMEC unidimensional chemotaxis were analyzed using transwell migration assays. As shown in Figure 4, the young male MK CM treatment elicited a trending increase in male BMEC migration, and the aged male MK CM significantly increased male BMEC migration (p = 0.047) compared to control male media (Figure 4A, 4C–4E). Similarly, female BMECs that were treated with aged female MK CM showed significantly more migration through the transwell membrane compared to control BMECs (p = 0.007, Figure 4B, 4G–4I). Images of positive control media (5% FBS) for male and female BMECs are found in Figure 4F, 4J, respectively. In general, aged MK CM promoted unidimensional migration of BMECs as compared to that observed with young MK CM.
Effects of MK CM on BMEC two-dimensional motility
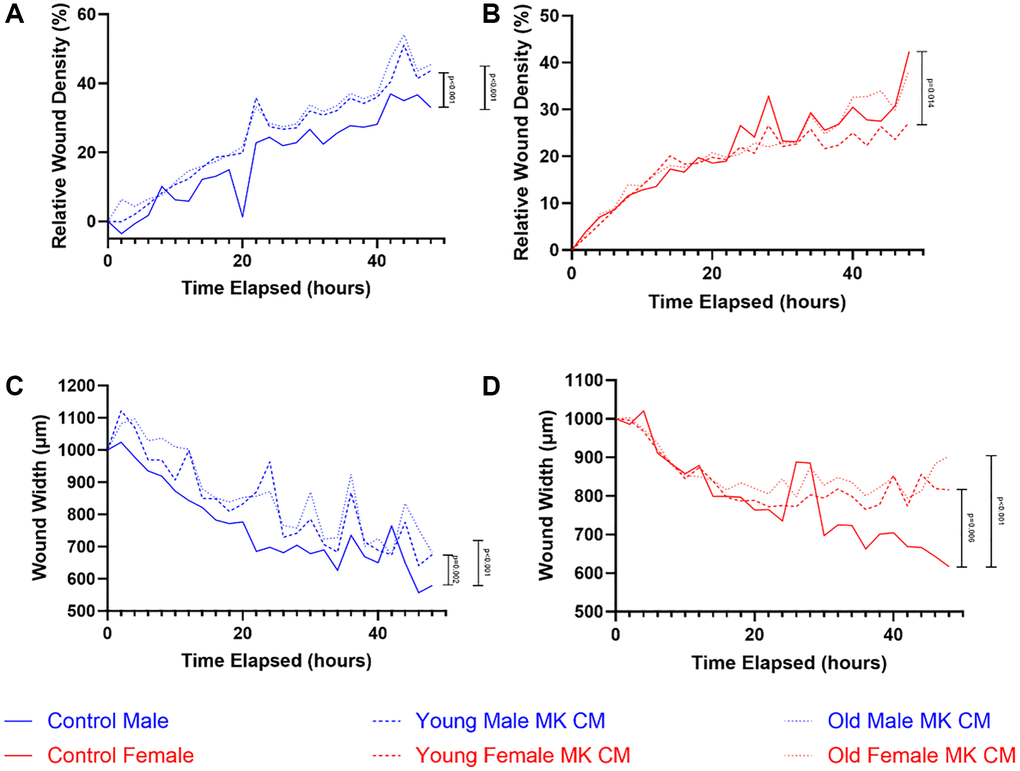
The effects of young and aged MK CM treatment on BMEC two-dimensional motility were examined through a wound migration assay, which compared the treatment and control groups in terms of the relative wound density of cells within the produced wound and the wound width over 48 hours (Figure 5). While relative wound density increased over time for all 3 groups, male MK CM treatment in male BMECs and control treatment in female BMECs led to greater relative wound density when compared by 2-way ANOVA; no significant differences were observed between groups specifically at 12, 24, or 48 hours (Figure 5A, 5B). When wound width was examined, control media resulted in the greatest decrease in both male and female BMECs compared to the MK CM groups, compared by 2-way ANOVA (Figure 5C, 5D). Similarly, no significant differences were observed between the groups specifically at 12, 24, or 48 hours for wound width in males. However, there was a significant difference between aged female and control wound width (p = 0.029) at 48 hours.
MK CM alters gene expression of angiogenic and inflammatory factors in BMECs
Further understanding of how MK CM affects BMECs was pursued. While the previous experiments analyzed BMEC functionality on a cellular level, we sought to examine the molecular changes as well. Eleven genes of receptors present in BMECs were examined for their relative mRNA expression (Figure 6 and Supplementary Figure 1). The selection process was based on the known presence of specific factors in MK CM. These factors may bind to the gene’s encoded receptors and have a downstream effect of changing the degree of transcription, thereby influencing the proangiogenic and antiangiogenic properties of the BMECs [44, 45].
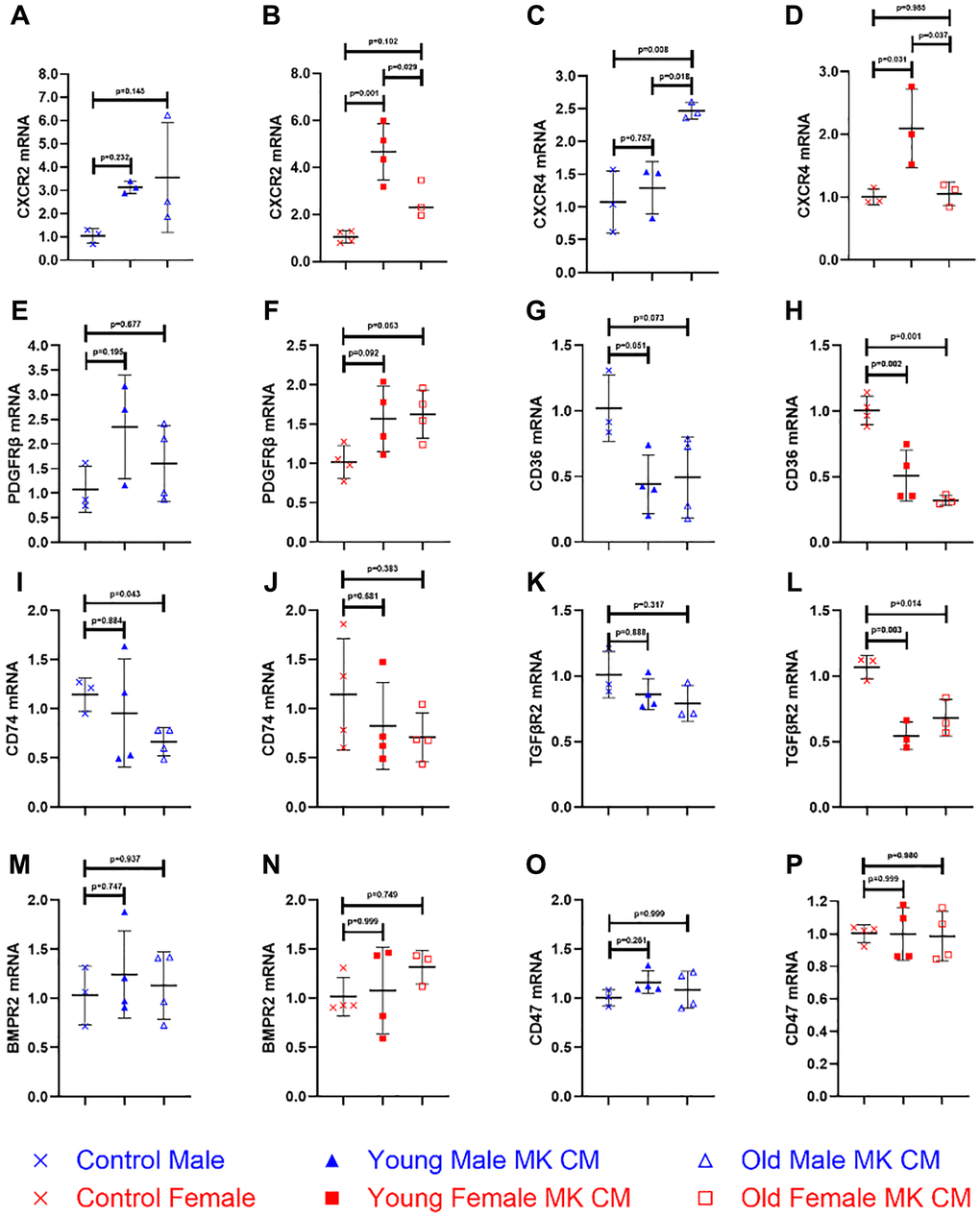
Of the eleven genes examined, five were found to have at least one type of MK CM exhibit significantly altered expression relative to controls. Both young and old male MK CM increased BMEC expression of CXCR2 (Figure 6A) though not significantly. Young female MK CM significantly increased CXCR2 expression, relative to both control and aged female MK CM (Figure 6B). Aged male MK CM significantly increased expression of CXCR4 (Figure 6C) not only relative to control, but also compared to young male MK CM. The opposite was true for CXCR4 with female MK CM, as young female MK CM significantly increased its expression relative to both control and aged female MK CM treatment (Figure 6D). Aged female MK CM increased expression of PDGFRβ to a nonsignificant degree, which was found to have mirroring trends in young female and young male MK CM treatments for this gene (Figure 6E, 6F). Young and aged female MK CM significantly decreased expression of CD36, while male MK CM showed trending decreases (Figure 6G, 6H). Aged male MK CM reduced expression of CD74 to a significant extent, while young male, young female, and old female MK CM treatments showed no significant change in expression of CD74 (Figure 6I, 6J). Female MK CM, both young and aged, decreased expression of TGFβR2 significantly while male MK CM treatments decreased its expression to a nonsignificant degree (Figure 6K, 6L). BMPR2 and CD47 expression remained unchanged (Figure 6M–6P), as did all other genes in Supplementary Figure 1.
Discussion
The risk of bone fractures increases with aging due to a variety of factors including decreased production of androgens and estrogens, decreased dietary intake of calcium and vitamin D, decreased osteoblast activity, and an increasingly sedentary lifestyle [4, 46, 47]. Moreover, bone diseases that are observed in aging populations such as osteoporosis and osteomalacia can lead to increased risk of fracture. MKs may play a role in these consequences of aging as they have been shown to increase in frequency and decrease in functional ability to stimulate bone production with age [43]. MKs and the factors they secrete have been established in recent decades to be prime regulators of the bone marrow microenvironment. While we have previously described in detail their ability to influence osteoblasts, osteoclasts, the hematopoietic niche, and several aging-associated bone disorders, this research is the first attempt at identifying changes that occur in the MK secretome due to aging and sex that contribute to effects on bone vasculature. Future studies are warranted to understand specific factors, signaling pathways, and mechanisms that explain our reported effects [48].
Notably, the relationship between MKs, angiogenesis, and ECs had not been fully described. Although numerous MK-secreted factors are known to influence EC functionality, few studies have directly examined the impact of MKs on the regulation of ECs. Tang et al. [25] conducted a vessel-like formation assay which found increased vessel lengths when ECs were treated with MK CM. Use of a TGF-β inhibitor decreased MK CM-induced vessel lengthening. It was also determined that increasing HIF-1α expression in ECs is one mechanism by which MK CM induces EC expression of VEGF. The effects of aging on MK CM and other aspects of EC functionality have not been examined to the best of our knowledge.
Our study examined BMECs isolated from young male and female mice when treated with young or aged MK CM in vitro. We analyzed the proliferation, formation of vessel-like structures, unidimensional and two-dimensional motility of these cells in response to the different MK CM, and changes observed in mRNA expression of receptors in BMECs in response to the different MK CM. This study aimed to understand the general effects of MK CM on BMECs, identify the differences between young and aged MK CM, and determine if these differences may potentially account for the changes in BMEC function with aging. A secondary goal of this study was to determine if any observable differences between sexes could be discerned. Sex-based differences in bone homeostasis and fracture healing have been well-established. Although the primary discussion regarding these differences typically focuses on the roles of estrogen and other sex hormones, sex-based differences in osteoblasts and BMEC responses to VEGF that result in divergent bone physiology have also been described [49].
The ability of MKs to secrete factors that upregulate BMEC proliferation and angiogenesis has been implicated through its known secretion of VEGF [42]. Moreover, patients with prolonged isolated thrombocytopenia have significantly reduced levels of VEGF secretion coupled with reduced BMEC counts, further suggesting that MK CM affects BMEC growth [50]. Here, we further support the assertions that MKs secrete factors that increase the proliferation of BMECs. Specifically, the BMECs from both sexes treated with young or old female MK CM exhibited significant increases in cell number compared to when they received control treatment (Figure 1B, 1D). When male BMECs were treated with male MK CM, only non-significant increases were observed (Figure 1A); female BMECs, however, proliferated significantly more with young male MK CM treatment and almost reached significance with old male MK CM treatment (Figure 1C). This may suggest that in terms of proliferation, female BMECs are more responsive to MK-secreted factors irrespective of the sex from which MKs were isolated. Furthermore, male MKs may also secrete different factors, or the factors may be generated at different concentrations than those generated by female MKs. Finally, it is also formally possible that the expression of receptors binding MK CM may be altered. Discriminating between these possibilities requires further experimentation.
We also analyzed four parameters in the vessel-like formation assay, also referred to as tube formation assay: nodes, meshes, number of vessel-like structures, and total vessel length. When examining our results, it becomes evident that MK CM stimulates BMEC vessel-like formation to an extent, as expected [25]. Female BMECs treated with young or aged MK CM of both sexes improved all assayed parameters of vessel-like formation (Figures 2 and 3). Young female MK CM caused trending increases in male BMEC vessel length and number (Figure 3C, 3D). Male BMECs treated with aged male MK CM saw only increases in the number of nodes and meshes and had fewer vessel-like structures than the young male MK CM counterparts (Figure 2A, 2C, 2E, 2G). Nodes and meshes are measurements of vessel-like structure interconnectedness, indicating that both young and aged male MK CM may maintain the vasculature complexity of BMECs. However, a significant decrease in the number of vessel-like structures between the young and aged MK CM-treated male groups demonstrates that the capability of MKs to induce BMECs vascular formation may be decreased with aging (Figure 2E). Reduced angiogenesis and vascular density have been characterized as consequences of aging [51]. While further examination is warranted, it is possible that this may be attributed partially to MK CM aging changes, specifically in males.
EC migration is a critical component of angiogenesis, particularly during the fracture healing process [52, 53]. When aged MK CM was used as a chemoattractant, significant increases in BMEC unidimensional or transwell migration were observed, while the young MK CM treatments showed trending increases. On the other hand, the two-dimensional, wound migration assay, identified divergent results. The calculation of relative wound density is the degree of cellular density found in the wound relative to the degree of cellular density found in the undamaged region surrounding the wound. Thus, it accounts for cells found in the wound that may have been present not only due to migration, but due to proliferation as well. There was a significant increase in relative wound density compared to control when male BMECs were treated with young or old male MK CM. Perplexingly, control media in the female BMECs resulted in greater relative wound density than young female MK CM. Wound width was significantly larger in BMECs treated with MK CM regardless of sex or age compared to control groups. This may initially seem to directly contradict the results of the transwell assay; however, the mechanisms of the two assays are important to note. Transwell assay examines BMECs transversing across a membrane toward a chemoattractant found in the underlying media. It is not directly supplemented by MK CM. On the other hand, wound migration assays directly supplement the BMECs with MK CM. The differences between the results of these two assays may be attributed to the variation in migratory behavior of BMECs in unidimensional and two-dimensional environments. Given the designs of each study type, the distribution of the chemotactic agent is different, and this could be a reason for some of the observed disparity in the two results. Overall, the migratory effects of MK CM on BMECs are complex and may hold the key to primary mechanisms by which EC functionality changes with aging [54].
Examining gene expression changes may further elucidate underlying mechanisms. Here we examined eleven genes. Expression patterns for three of the eleven genes analyzed are found in Supplementary Figure 1. FLT-1 and KDR are VEGF receptors 1 and 2, respectively. FLT-1 has shown contrasting results regarding its effect on ECs; while it has been described as antiangiogenic, particularly in embryonic contexts, it has also shown proangiogenic properties [55, 56]. Primarily, soluble FLT-1 functions as a regulator of EC sprouting. Hypoxia reduces soluble FLT-1, which inhibits VEGF, thus creating a directional growth to the hypoxic tissue due to uninhibited VEGF [57]. KDR has also shown different results, although it has not been as clearly characterized as being antiangiogenic [58, 59]. ITGβ1, the integrin β1 receptor, has been shown to inhibit EC proliferation but promote sprouting and migration [60]. While these factors are significant components of EC function, their expression with MK CM treatment remained unchanged. Two other genes of interest, BMPR2 and CD47, found in Figure 6 also lack significant differences across all groups. While BMPs and their receptors are typically thought of as osteoblast and osteoclast regulators, they are also found on ECs and can influence their growth [61]. However, MK CM did not affect the expression of BMPR2, despite the known secretion of BMPs by MK CM [48]. CD47, also known as integrin-associated protein and a possible regulator of EC senescence, showed no differences [62, 63].
CXCR2 and CXCR4 are CXC chemokine receptors and typically regarded to be inducers of angiogenesis [64]. CXCR2 is a receptor of growth-related oncogenes (GRO)-α, β, and γ, as well as neutrophil-activating peptide (NAP)-2, epithelial neutrophil-activating peptide (ENA)-78, and interleukin (IL)-8. These factors have all been found to be promoters of EC growth, migration, and angiogenesis [64–66]. We found a significant increase in CXCR2 expression in young female MK CM treatment, compared to both control and aged female MK CM. CXCR4 is the receptor for stromal-cell derived factor (SDF)-1, known for its proangiogenic properties specifically in BMECs [67]. CXCR4 expression was among the more surprising results: while aged male MK CM significantly increased its expression, young male MK CM showed no change and was in fact significantly reduced when compared to aged male MK CM. The opposite was found in the females, with aged female MK CM showing no differences and young female MK CM significantly increasing BMEC expression of CXCR4. At this time, differences between males and females in their MK secreted factor components has not been fully described. What is known is MKs express estrogen receptors, and estrogen has been shown to promote MK maturation and potentially increase their proliferation [48, 68]. Moreover, estrogen increases MK expression of osteoprotegerin (OPG) and reduces RANKL expression [69]. Despite this, the differences in MK CM effects with regards to CXCR4 and its ligand SDF-1 should be elucidated further.
Some of the genes assessed presented clearer results. PDGFRβ is the receptor for PDGF-BB and is an established proangiogenic receptor [70]. Both young and aged female MK CM and young male MK CM caused trending elevation in its expression. It is interesting to note the correlation between the expression of PDGFRβ and the results of the unidimensional, transwell assay: PDGF-BB is a chemoattractant that induces EC motility, so it may be the primary factor involved in transwell migration of the BMECs [71]. CD36 is a thrombospondin-1 receptor and is typically regarded to be an inhibitor of angiogenesis [72]. Its expression was significantly decreased in both female MK CM treatments and exhibited trending decreases in both male MK CM treatments. The increases in vessel-like formation may be particularly attributed to reductions in CD36 expression, as there were significant improvements in all parameters with exception of aged male MK CM treatment.
CD74 was observed to have trending reductions in its expression with both female MK CM and a significant decrease upon old male MK CM treatments. CD74 is the receptor for the proinflammatory mediator, macrophage migration inhibitory factor (MIF). Although MIF is proangiogenic and most other proinflammatory receptors have shown increases, it is possible that MK secretion of MIF is upregulated to such an extent that it reduces the expression of CD74 through a negative feedback mechanism [45, 73]. TGFβR2 showed similar results, with trending reductions in male BMECs and significant reductions in female BMECs. Just as with CD74, TGFβR2 is a proangiogenic receptor, and its reduced expression typically results in impaired vasculogenesis [74]. Thus, it may be that MKs secrete high levels of TGF-β, and its abundant presence in MK CM may lead to negative feedback inhibition. TGFβR2, in particular, has been shown to be downregulated with increased TGF-β expression [44].
The interpretation of RNA expression, as it currently stands, requires further substantiation. All the genes analyzed herein were those of BMEC receptors, and receptor expression may increase or decrease for different reasons that are often contrasting. CD36 transcription regulation, by example, is contingent on a diverse array of extracellular signaling such as IL-4 and TGF-β, in addition to non-coding RNAs and intracellular mechanisms [75]. The abundance of a factor and its ability to influence the cell’s functionality may result in upregulation or downregulation of its receptor through various feedback mechanisms. This is a logical future direction. Furthermore, comparing the bone vascular niches as well as our gene panel in young and aged male and female mice in vivo may support our findings and elucidate mechanisms behind our observed differences in vitro.
In conclusion, we have reported various similarities and differences in the effects of MK CM on BMEC biology based on age and sex, which aligns with the growing literature that MK secreted factors alter the bone marrow microenvironment (Table 1). Specifically, we found that treatment with all tested categories of MK CM (young, aged, male, female) increased the tendency of BMECs to form vessel-like structures, undergo unidimensional migration, upregulated the expression of the proangiogenic gene CXCR2, and downregulated the expression of CD36. We also found that female MK CM increased the proliferation of BMECs while male MK CM showed only an effect on female BMECs, and MK CM generally decreased the two-dimensional motility of BMECs. It is important to note that this reduction in wound migration found with MK CM treatment, as well as some of the aging and sex-based differences in gene expression, particularly found in CXCR4, should be further examined. An understanding of which factors regulate which mechanisms of EC functionality may allow for isolation of one or a few factors that influence EC migration changes with aging, resulting in the development of targeted therapy to improve EC migration, subsequent angiogenesis, and fracture healing.
Table 1. Results summary.
| Young male MK CM | Old male MK CM | Young female MK CM | Old female MK CM | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Proliferation | − | − | ↑↑ | ↑↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Nodes | ↑↑ | ↑↑ | ↑↑ | ↑↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Meshes | ↑↑ | ↑↑ | ↑↑ | ↑↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| # Vessel-like Structures | ↑↑ | ↑ | ↑↑ | ↑↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total Vessel Length | ↑↑ | ↑ | ↑↑ | ↑↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1-Dimensional Migration | ↑ | ↑↑ | − | ↑↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Relative Wound Density | ↑↑ | ↑↑ | ↓↓ | − | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Wound Width | ↑↑ | ↑↑ | ↑↑ | ↑↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CXCR2 Expression | ↑ | ↑ | ↑↑ | ↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CXCR4 Expression | − | ↑↑ | ↑↑ | - | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PDGFRβ Expression | ↑ | − | ↑ | ↑ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CD36 Expression | ↓ | ↓ | ↓↓ | ↓↓ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CD74 Expression | − | ↓↓ | − | − | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TGFβR2 Expression | − | ↓ | ↓↓ | ↓↓ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BMPR2 Expression | − | − | − | − | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CD47 Expression | − | − | − | − | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ITGβ1 Expression | − | − | − | − | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FLT-1 Expression | − | − | − | − | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KDR Expression | − | − | − | − | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ↑↑/↓↓ = statistically significant. ↑/↓ = trending, not statistically significant. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Materials and Methods
Animals
Young (3–4-month-old) and old (22–24-month-old), male and female C57BL/6J mice were purchased from Jackson Laboratory or were generously provided by the National Institute of Aging (NIA), respectively. The Indiana University School of Medicine Institutional Animal Care and Use Committee approved all described studies.
EC isolation from BM
BMECs were isolated from the BM of femurs, tibiae, and humeri of all mice. Briefly, after euthanasia, skin, muscle, and soft tissue were stripped from each of the bones, after which the distal and proximal epiphyses of the bones were removed. All bones were individually placed into sterile, punctured 0.5 mL tubes that were inserted into 1.5 mL tubes. Each 1.5 mL tube contained 750 μL α-MEM (Gibco, Grand Island, NY, USA) with 10% fetal bovine serum (FBS, Biowest, Riverside, MO, USA). BM was isolated from the bones by centrifugation at 14,000 g for 2 minutes at 4°C. The extracted BM pellets were then resuspended in 1 mL of Complete EC Growth Media (ScienCell, Carlsbad, CA, USA), which contained EC growth supplements, 7.5% FBS, and 1% penicillin/streptomycin (Gibco, Grand Island, NY, USA). The resuspended pellets were plated in a 12-well plate coated with 4 μg/mL fibronectin (Thermo Fisher Scientific, Waltham, MA, USA) containing an additional 1 mL of Complete EC Growth Media. Media changes were performed every two days and BMECs were cultured for 5–7 days prior to use in experimental assays.
MK CM preparation
Murine BM from C57BL/6J mice was extracted and plated in Dulbecco’s Modified Eagle Medium (DMEM; Gibco, Grand Island, NY, USA), supplemented with 5% FBS (Hyclone, Logan, UT, USA), 1% penicillin/streptomycin/glutamine (Gibco, Grand Island, NY, USA), and conditioned medium from a TPO producing cell line [38] until numerous large MKs were observed. MKs were then isolated from the other cell population using a one-step Bovine Serum Albumin (BSA, Millipore-Sigma, Burlington, MA, USA) gradient [38]. The isolated MKs were removed from the bottom of the gradient, washed with Phosphate Buffered Saline (PBS, Gibco, Grand Island, NY, USA), and re-suspended in phenol-free α-MEM at 1 million cells/ml for 3 days. The CM was collected, centrifuged, and stored at −80°C until use.
Proliferation assay
BMECs were seeded in a 96-well plate (Corning, Corning, NY, USA) at 1 × 103 cells/well with 25% MK CM isolated from young or old mice of both sexes (all containing 1.25% FBS), or 25% α-MEM (Control Media) containing 1.25% FBS, or α-MEM with 0% FBS (Negative control) and were incubated for 1 to 2 days to assess proliferation. The BMECs were fixed with 5% neutral buffered formalin (NBF) at room temperature for 15 minutes. The fixed BMECs were stained with 0.05% crystal violet for 20 minutes and were subsequently washed with tap water prior to an overnight drying period. The EVOS FL Cell Imaging System was utilized to image BMEC cultures, and ImageJ.1.52a software was used for counting analysis.
Vessel-like structure formation assay
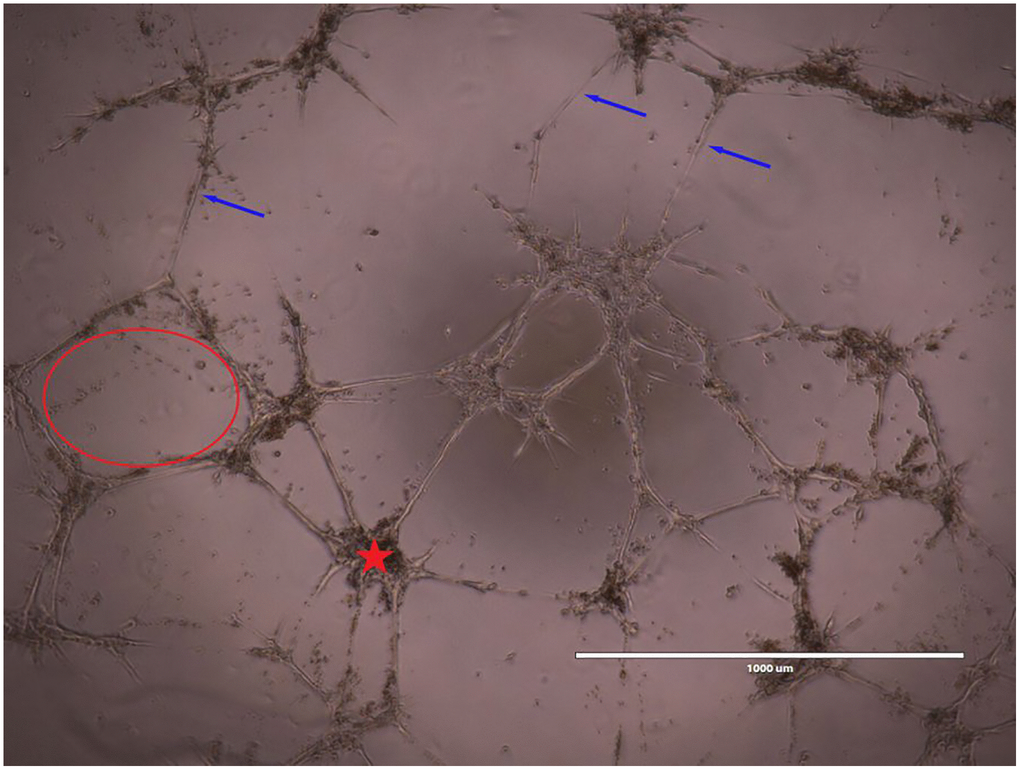
96-well plates (Corning Inc., Corning, NY, USA) were chilled in a −20°C freezer overnight. They were subsequently coated with 50 μL/well of Matrigel basement membrane matrix (Corning, Corning, NY, USA) and incubated 45 minutes at 37°C prior to plating and culturing BMECs. BMECs were seeded at 1 × 104 cells/well with their respective treatments and incubated for 8 hours prior to cell imaging (EVOS FL Cell Imaging System). α-MEM (Control group), 25% MK CM isolated from young or old mice of both sexes (all containing 1.25% FBS), and α-MEM with 5% FBS (positive control) were the designated groups. Vessel-like structure formation was quantified with ImageJ.1.52a software. Images are taken at 10× magnification. The number of nodes, number of meshes, number of complete vessels, and total vessel-like structure length were the parameters analyzed and an example is shown in Figure 7 [30]. Briefly, meshes were defined as areas enclosed by three or more nodes, and nodes were defined as a point where two or more vessels intersect. Simple Neurite Tracer, a plugin of ImageJ.1.52b software, was employed for the manual measurement of the total vessel length and number of vessel-like structures. A blinded analyzer manually measured the number of vessel-like structures as well as vessel length. The automated Angiogenesis Analyzer, a plugin of ImageJ.1.52a software, was used to analyze the number of meshes and nodes.
Transwell assay or unidimensional chemotaxis assay
500 μL of 25% MK CM or 25% α-MEM was added to the bottom of each well in 24-well plates; α-MEM with 5% FBS was used as a positive control. A transwell membrane (8 μm pore size, Corning, Corning, NY, USA) was placed over the media. Cells resuspended in Basal EC Growth Media (ScienCell, Carlsbad, CA, USA containing only 7.5% FBS and 1% penicillin/streptomycin solution. Basal EC Growth Media lacked EC growth supplements.) were seeded on the top of the membrane at a density of 4 × 105 cells/well. The plates were placed in an incubator at 37°C for a period of 24 hours. The membranes were fixed with 5% NBF at room temperature for 15 minutes. The membranes were stained with 0.5% crystal violet for 20 minutes and dried using sterile cotton tip applicators (Puritan Medical Products, Gulliford, ME, USA). The EVOS FL Cell Imaging System was used to image the migration of the BMECs across the membrane at a magnification of 10×. Analysis of images was completed with the ImageJ.1.52a software.
Wound migration assay
96-well plates (Corning, Corning, NY, USA) were seeded with BMECs placed in Complete EC Growth Media at a density of 1 × 105 cells/well and incubated at 37°C for a period of 24 hours to achieve 100% confluence. Complete EC Growth Media was aspirated and replaced with 25% MK CM or 25% α-MEM. The IncuCyte® WoundMaker (Essen BioScience, Ann Arbor, MI, USA) generated a wound in the center of every well. The IncuCyte ZOOM® Live-Cell Analysis System (Essen BioScience, Ann Arbor, MI, USA) took images of each well every 2 hours at 10X magnification for 48 hours. Relative wound density and wound widths were compared across the entire timecourse and specifically at 12, 24, and 48 hours. The IncuCyte™ Scratch Wound Cell Migration Software (Essen BioScience, Ann Arbor, MI, USA) analyzed the images taken.
RNA isolation and gene expression
BMECs were seeded in a 6-well plate at a concentration of 1 × 106 cells/well with 25% MK CM or 25% α-MEM. The plates were incubated at 37°C for 24 hours to achieve no less than 80% confluency. The RNeasy Mini kit (QIAGEN, Hilden, Germany) was used for total RNA isolation. The Transcriptor First Strand cDNA Synthesis Kit (Roche, Basel, Switzerland) was used to prepare cDNA, which was then analyzed through quantitative PCR using the Power SYBR™ Green PCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA) on a CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). GAPDH was used as an internal control. The fold change in gene expression was obtained through the 2−ΔΔCT method. Several genes were analyzed. Table 2 lists the genes and primers used.
Table 2. Primer pairs for gene expression.
| Gene | Forward sequence | Reverse sequence |
| CXCR2 | CTCTATTCTGCCAGATGCTGTCC | ACAAGGCTCAGCAGAGTCACCA |
| CXCR4 | GACTGGCATAGTCGGCAATGGA | CAAAGAGGAGGTCAGCCACTGA |
| PDGFRβ | CATCCGCTCCTTTGATGATCTT | GTGCTCGGGTCATGTTCAAGT |
| CD36 | GGACATTGAGATTCTTTTCCTCTG | GCAAAGGCATTGGCTGGAAGAAC |
| CD74 | CGCGACCTCATCTCTAACCAT | ACAGGTTTGGCAGATTTCGG |
| TGFβR2 | CCTACTCTGTCTGTGGATGACC | GACATCCGTCTGCTTGAACGAC |
| BMPR2 | AGAGACCCAAGTTCCCAGAAGC | TCTCCTCAGCACACTGTGCAGT |
| CD47 | TGCGGTTCAGCTCAACTACTG | GCTTTGCGCCTCCACATTAC |
| ITGβ1 | CTCCAGAAGGTGGCTTTGATGC | GTGAAACCCAGCATCCGTGGAA |
| FLT1 | CCACCTCTCTATCCGCTGG | ACCAATGTGCTAACCGTCTTATT |
| KDR/VEGFR2 | TTTGGCAAATACAACCCTTCAGA | GCTCCAGTATCATTTCCAACCA |
| GAPDH | CGTGGGGCTGCCCAGAACAT | TCTCCAGGCGGCACGTCAGA |
Statistics
Statistical analyses and data normality calculations were performed with GraphPad Prism (ver. 8 and 10, GraphPad Software, La Jolla, CA, USA). Results are displayed in figures as mean ± SD or line graphs. Data in this manuscript comparing 3 or more groups were analyzed by one-way analysis of variance (ANOVA) with Tukey-Kramer post hoc test for parametric data. A Kruskal-Wallis test with a Dunn’s multiple comparison post-hoc test was used for nonparametric data. For the wound healing assay, a two-way ANOVA with a Dunnett’s Multiple comparison test was used to examine the main treatment effects. Specific post-hoc p-values are displayed on the graphs, and a p-value of less than 0.05 was considered significant.
Supplementary Materials
Author Contributions
M.K. Nazzal, H.L. Battina, N.P. Tewari, U.C. Dadwal, and M.A. Kacena designed the research; M.K. Nazzal, H.L. Battina, N.P. Tewari, S.L. Mostardo, R.U. Nagaraj, D. Zhou, O.D. Awosanya, S.K. Majety, S. Samson, R.J. Blosser, U.C. Dadwal, and P.L. Mulcrone performed the research; M.K. Nazzal, H.L. Battina, N.P. Tewari, and M.A. Kacena wrote the paper; M.K. Nazzal, H.L. Battina, N.P. Tewari, S.L. Mostardo, R.U. Nagaraj, D. Zhou, O.D. Awosanya, S.K. Majety, S. Samson, R.J. Blosser, P.L. Mulcrone, U.C. Dadwal, and M.A. Kacena analyzed the data, critically revised the paper, approved the final version of the paper, and take responsibility for the contents of the paper. M.A. Kacena and P.L. Mulcrone take responsibility for the integrity of the data analysis.
Acknowledgments
We would like to thank the Indiana University Angiogenesis Core for aiding with wound migration studies. We would also like to thank the Indiana University In Vitro Therapeutics Cores for providing mice used for these studies.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Ethical Statement
The Indiana University School of Medicine Institutional Animal Care and Use Committee approved all described studies, protocol number is 23133.
Funding
Core facilities are supported in part by the NIH U24DK126127. This project was financed, in part, by the Cooperative Center of Excellence in Hematology (CCEH) Award, funded by the National Institute of Diabetes and Digestive and Kidney Diseases of the NIH under U24DK126127 (MAK, UCD) and U54DK106846 (NPT). The project was also supported, in part, by NIH T32 DK007519 (UCD), NIH T32 AR065971 (UCD), NIH R01 AG060621 (MAK), NIH T32 AR065971 (MKN), NIH F31 AG077931 (ODA), and NIH T32 HL007910 (ODA). In addition, the results of this work were supported with resources and the use of facilities at the Richard L. Roudebush VA Medical Center, Indianapolis, IN: VA Merit #BX003751 and #RX003552 (MAK) and Research Career Scientist #IRX004809A. The views expressed in this article are solely those of the authors and do not necessarily represent the official position or policy of any of the agencies mentioned.
References
- 1. Rose S, Maffulli N. Hip fractures. An epidemiological review. Bull Hosp Jt Dis. 1999; 58:197–201. [PubMed]
- 2. Green E, Lubahn JD, Evans J. Risk factors, treatment, and outcomes associated with nonunion of the midshaft humerus fracture. J Surg Orthop Adv. 2005; 14:64–72. [PubMed]
- 3. Cauley JA, Thompson DE, Ensrud KC, Scott JC, Black D. Risk of mortality following clinical fractures. Osteoporos Int. 2000; 11:556–61. https://doi.org/10.1007/s001980070075 [PubMed]
- 4. Office of the Surgeon General (US). Bone Health and Osteoporosis: A Report of the Surgeon General. Rockville (MD): Office of the Surgeon General (US). 2004. [PubMed]
- 5. Nieminen S, Nurmi M, Satokari K. Healing of femoral neck fractures; influence of fracture reduction and age. Ann Chir Gynaecol. 1981; 70:26–31. [PubMed]
- 6. Geerts WH, Heit JA, Clagett GP, Pineo GF, Colwell CW, Anderson FA
Jr , Wheeler HB. Prevention of venous thromboembolism. Chest. 2001; 119:132S–75. https://doi.org/10.1378/chest.119.1_suppl.132s [PubMed] - 7. Phillips AM. Overview of the fracture healing cascade. Injury. 2005 (Suppl 3); 36:S5–7. https://doi.org/10.1016/j.injury.2005.07.027 [PubMed]
- 8. Hankenson KD, Zimmerman G, Marcucio R. Biological perspectives of delayed fracture healing. Injury. 2014 (Suppl 2); 45:S8–15. https://doi.org/10.1016/j.injury.2014.04.003 [PubMed]
- 9. Kurdy NM, Weiss JB, Bate A. Endothelial stimulating angiogenic factor in early fracture healing. Injury. 1996; 27:143–5. https://doi.org/10.1016/0020-1383(95)00169-7 [PubMed]
- 10. Einhorn TA, Gerstenfeld LC. Fracture healing: mechanisms and interventions. Nat Rev Rheumatol. 2015; 11:45–54. https://doi.org/10.1038/nrrheum.2014.164 [PubMed]
- 11. Lopas LA, Belkin NS, Mutyaba PL, Gray CF, Hankenson KD, Ahn J. Fractures in geriatric mice show decreased callus expansion and bone volume. Clin Orthop Relat Res. 2014; 472:3523–32. https://doi.org/10.1007/s11999-014-3829-x [PubMed]
- 12. Bergman RJ, Gazit D, Kahn AJ, Gruber H, McDougall S, Hahn TJ. Age-related changes in osteogenic stem cells in mice. J Bone Miner Res. 1996; 11:568–77. https://doi.org/10.1002/jbmr.5650110504 [PubMed]
- 13. Gruber R, Koch H, Doll BA, Tegtmeier F, Einhorn TA, Hollinger JO. Fracture healing in the elderly patient. Exp Gerontol. 2006; 41:1080–93. https://doi.org/10.1016/j.exger.2006.09.008 [PubMed]
- 14. Baxter MA, Wynn RF, Jowitt SN, Wraith JE, Fairbairn LJ, Bellantuono I. Study of telomere length reveals rapid aging of human marrow stromal cells following in vitro expansion. Stem Cells. 2004; 22:675–82. https://doi.org/10.1634/stemcells.22-5-675 [PubMed]
- 15. Colnot CI, Helms JA. A molecular analysis of matrix remodeling and angiogenesis during long bone development. Mech Dev. 2001; 100:245–50. https://doi.org/10.1016/s0925-4773(00)00532-3 [PubMed]
- 16. Gerstenfeld LC, Cullinane DM, Barnes GL, Graves DT, Einhorn TA. Fracture healing as a post-natal developmental process: molecular, spatial, and temporal aspects of its regulation. J Cell Biochem. 2003; 88:873–84. https://doi.org/10.1002/jcb.10435 [PubMed]
- 17. Tevlin R, Walmsley GG, Marecic O, Hu MS, Wan DC, Longaker MT. Stem and progenitor cells: advancing bone tissue engineering. Drug Deliv Transl Res. 2016; 6:159–73. https://doi.org/10.1007/s13346-015-0235-1 [PubMed]
- 18. Prisby RD, Ramsey MW, Behnke BJ, Dominguez JM
2nd , Donato AJ, Allen MR, Delp MD. Aging reduces skeletal blood flow, endothelium-dependent vasodilation, and NO bioavailability in rats. J Bone Miner Res. 2007; 22:1280–8. https://doi.org/10.1359/jbmr.070415 [PubMed] - 19. Prisby RD. Bone marrow blood vessel ossification and "microvascular dead space" in rat and human long bone. Bone. 2014; 64:195–203. https://doi.org/10.1016/j.bone.2014.03.041 [PubMed]
- 20. Lu C, Hansen E, Sapozhnikova A, Hu D, Miclau T, Marcucio RS. Effect of age on vascularization during fracture repair. J Orthop Res. 2008; 26:1384–9. https://doi.org/10.1002/jor.20667 [PubMed]
- 21. Ghiasi MS, Chen J, Vaziri A, Rodriguez EK, Nazarian A. Bone fracture healing in mechanobiological modeling: A review of principles and methods. Bone Rep. 2017; 6:87–100. https://doi.org/10.1016/j.bonr.2017.03.002 [PubMed]
- 22. Dadwal UC, Staut CA, Tewari NP, Awosanya OD, Mendenhall SK, Valuch CR, Nagaraj RU, Blosser RJ, Li J, Kacena MA. Effects of diet, BMP-2 treatment, and femoral skeletal injury on endothelial cells derived from the ipsilateral and contralateral limbs. J Orthop Res. 2022; 40:439–48. https://doi.org/10.1002/jor.25033 [PubMed]
- 23. Dadwal UC, Bhatti FUR, Awosanya OD, Staut CA, Nagaraj RU, Perugini AJ
3rd , Tewari NP, Valuch CR, Sun S, Mendenhall SK, Zhou D, Mostardo SL, Blosser RJ, et al. The Effects of SRT1720 Treatment on Endothelial Cells Derived from the Lung and Bone Marrow of Young and Aged, Male and Female Mice. Int J Mol Sci. 2021; 22:11097. https://doi.org/10.3390/ijms222011097 [PubMed] - 24. Ucuzian AA, Gassman AA, East AT, Greisler HP. Molecular mediators of angiogenesis. J Burn Care Res. 2010; 31:158–75. https://doi.org/10.1097/BCR.0b013e3181c7ed82 [PubMed]
- 25. Tang Y, Hu M, Xu Y, Chen F, Chen S, Chen M, Qi Y, Shen M, Wang C, Lu Y, Zhang Z, Zeng H, Quan Y, et al. Megakaryocytes promote bone formation through coupling osteogenesis with angiogenesis by secreting TGF-β1. Theranostics. 2020; 10:2229–42. https://doi.org/10.7150/thno.40559 [PubMed]
- 26. Mulcrone PL, Campbell JP, Clément-Demange L, Anbinder AL, Merkel AR, Brekken RA, Sterling JA, Elefteriou F. Skeletal Colonization by Breast Cancer Cells Is Stimulated by an Osteoblast and β2AR-Dependent Neo-Angiogenic Switch. J Bone Miner Res. 2017; 32:1442–54. https://doi.org/10.1002/jbmr.3133 [PubMed]
- 27. James AW, LaChaud G, Shen J, Asatrian G, Nguyen V, Zhang X, Ting K, Soo C. A Review of the Clinical Side Effects of Bone Morphogenetic Protein-2. Tissue Eng Part B Rev. 2016; 22:284–97. https://doi.org/10.1089/ten.TEB.2015.0357 [PubMed]
- 28. Gillman CE, Jayasuriya AC. FDA-approved bone grafts and bone graft substitute devices in bone regeneration. Mater Sci Eng C Mater Biol Appl. 2021; 130:112466. https://doi.org/10.1016/j.msec.2021.112466 [PubMed]
- 29. Vukicevic S, Oppermann H, Verbanac D, Jankolija M, Popek I, Curak J, Brkljacic J, Pauk M, Erjavec I, Francetic I, Dumic-Cule I, Jelic M, Durdevic D, et al. The clinical use of bone morphogenetic proteins revisited: a novel biocompatible carrier device OSTEOGROW for bone healing. Int Orthop. 2014; 38:635–47. https://doi.org/10.1007/s00264-013-2201-1 [PubMed]
- 30. Bhatti FUR, Dadwal UC, Valuch CR, Tewari NP, Awosanya OD, de Andrade Staut C, Sun S, Mendenhall SK, Perugini AJ
3rd , Nagaraj RU, Battina HL, Nazzal MK, Blosser RJ, et al. The effects of high fat diet, bone healing, and BMP-2 treatment on endothelial cell growth and function. Bone. 2021; 146:115883. https://doi.org/10.1016/j.bone.2021.115883 [PubMed] - 31. Cardier JE, Dempsey J. Thrombopoietin and its receptor, c-mpl, are constitutively expressed by mouse liver endothelial cells: evidence of thrombopoietin as a growth factor for liver endothelial cells. Blood. 1998; 91:923–9. [PubMed]
- 32. Liu KW, Hu B, Cheng SY. Platelet-derived growth factor signaling in human malignancies. Chin J Cancer. 2011; 30:581–4. https://doi.org/10.5732/cjc.011.10300 [PubMed]
- 33. Dahabreh Z, Calori GM, Kanakaris NK, Nikolaou VS, Giannoudis PV. A cost analysis of treatment of tibial fracture nonunion by bone grafting or bone morphogenetic protein-7. Int Orthop. 2009; 33:1407–14. https://doi.org/10.1007/s00264-008-0709-6 [PubMed]
- 34. Bowen-Pope DF, Malpass TW, Foster DM, Ross R. Platelet-derived growth factor in vivo: levels, activity, and rate of clearance. Blood. 1984; 64:458–69. [PubMed]
- 35. Kacena MA, Chu TG, inventors; Osteofuse, Inc., assignee. Use of compounds with thrombopoietic activity to promote bone growth and healing. Indiana University (P0053PCT, 6-3-11). United States patent US 16/392,246. 2019 Apr 23. https://patentimages.storage.googleapis.com/3d/eb/08/04a4c63cdd0242/US20190240291A1.pdf.
- 36. Kacena MA, Gundberg CM, Nelson T, Horowitz MC. Loss of the transcription factor p45 NF-E2 results in a developmental arrest of megakaryocyte differentiation and the onset of a high bone mass phenotype. Bone. 2005; 36:215–23. https://doi.org/10.1016/j.bone.2004.09.024 [PubMed]
- 37. Kacena MA, Shivdasani RA, Wilson K, Xi Y, Troiano N, Nazarian A, Gundberg CM, Bouxsein ML, Lorenzo JA, Horowitz MC. Megakaryocyte-osteoblast interaction revealed in mice deficient in transcription factors GATA-1 and NF-E2. J Bone Miner Res. 2004; 19:652–60. https://doi.org/10.1359/JBMR.0301254 [PubMed]
- 38. Villeval JL, Cohen-Solal K, Tulliez M, Giraudier S, Guichard J, Burstein SA, Cramer EM, Vainchenker W, Wendling F. High thrombopoietin production by hematopoietic cells induces a fatal myeloproliferative syndrome in mice. Blood. 1997; 90:4369–83. [PubMed]
- 39. Yan XQ, Lacey D, Hill D, Chen Y, Fletcher F, Hawley RG, McNiece IK. A model of myelofibrosis and osteosclerosis in mice induced by overexpressing thrombopoietin (mpl ligand): reversal of disease by bone marrow transplantation. Blood. 1996; 88:402–9. [PubMed]
- 40. Cheng YH, Streicher DA, Waning DL, Chitteti BR, Gerard-O'Riley R, Horowitz MC, Bidwell JP, Pavalko FM, Srour EF, Mayo LD, Kacena MA. Signaling pathways involved in megakaryocyte-mediated proliferation of osteoblast lineage cells. J Cell Physiol. 2015; 230:578–86. https://doi.org/10.1002/jcp.24774 [PubMed]
- 41. Lee YS, Kwak MK, Moon SA, Choi YJ, Baek JE, Park SY, Kim BJ, Lee SH, Koh JM. Regulation of bone metabolism by megakaryocytes in a paracrine manner. Sci Rep. 2020; 10:2277. https://doi.org/10.1038/s41598-020-59250-6 [PubMed]
- 42. Möhle R, Green D, Moore MA, Nachman RL, Rafii S. Constitutive production and thrombin-induced release of vascular endothelial growth factor by human megakaryocytes and platelets. Proc Natl Acad Sci U S A. 1997; 94:663–8. https://doi.org/10.1073/pnas.94.2.663 [PubMed]
- 43. Maupin KA, Himes ER, Plett AP, Chua HL, Singh P, Ghosh J, Mohamad SF, Abeysekera I, Fisher A, Sampson C, Hong JM, Childress P, Alvarez M, et al. Aging negatively impacts the ability of megakaryocytes to stimulate osteoblast proliferation and bone mass. Bone. 2019; 127:452–9. https://doi.org/10.1016/j.bone.2019.07.010 [PubMed]
- 44. Kucuksayan H, Akgun S, Ozes ON, Alikanoglu AS, Yildiz M, Dal E, Akca H. TGF-β-SMAD-miR-520e axis regulates NSCLC metastasis through a TGFBR2-mediated negative-feedback loop. Carcinogenesis. 2019; 40:695–705. https://doi.org/10.1093/carcin/bgy166 [PubMed]
- 45. Chesney J, Metz C, Bacher M, Peng T, Meinhardt A, Bucala R. An essential role for macrophage migration inhibitory factor (MIF) in angiogenesis and the growth of a murine lymphoma. Mol Med. 1999; 5:181–91. [PubMed]
- 46. Riggs BL, Melton LJ
3rd . The worldwide problem of osteoporosis: insights afforded by epidemiology. Bone. 1995; 17:505S–11. https://doi.org/10.1016/8756-3282(95)00258-4 [PubMed] - 47. Chrischilles EA, Butler CD, Davis CS, Wallace RB. A model of lifetime osteoporosis impact. Arch Intern Med. 1991; 151:2026–32. [PubMed]
- 48. Karnik SJ, Nazzal MK, Kacena MA, Bruzzaniti A. Megakaryocyte Secreted Factors Regulate Bone Marrow Niche Cells During Skeletal Homeostasis, Aging, and Disease. Calcif Tissue Int. 2023; 113:83–95. https://doi.org/10.1007/s00223-023-01095-y [PubMed]
- 49. Ortona E, Pagano MT, Capossela L, Malorni W. The Role of Sex Differences in Bone Health and Healing. Biology (Basel). 2023; 12:993. https://doi.org/10.3390/biology12070993 [PubMed]
- 50. Kong Y, Hu Y, Zhang XH, Wang YZ, Mo XD, Zhang YY, Wang Y, Han W, Xu LP, Chang YJ, Huang XJ. Association between an impaired bone marrow vascular microenvironment and prolonged isolated thrombocytopenia after allogeneic hematopoietic stem cell transplantation. Biol Blood Marrow Transplant. 2014; 20:1190–7. https://doi.org/10.1016/j.bbmt.2014.04.015 [PubMed]
- 51. Rivard A, Fabre JE, Silver M, Chen D, Murohara T, Kearney M, Magner M, Asahara T, Isner JM. Age-dependent impairment of angiogenesis. Circulation. 1999; 99:111–20. https://doi.org/10.1161/01.cir.99.1.111 [PubMed]
- 52. Lamalice L, Le Boeuf F, Huot J. Endothelial cell migration during angiogenesis. Circ Res. 2007; 100:782–94. https://doi.org/10.1161/01.RES.0000259593.07661.1e [PubMed]
- 53. Guo S, Lok J, Liu Y, Hayakawa K, Leung W, Xing C, Ji X, Lo EH. Assays to examine endothelial cell migration, tube formation, and gene expression profiles. Methods Mol Biol. 2014; 1135:393–402. https://doi.org/10.1007/978-1-4939-0320-7_32 [PubMed]
- 54. Lähteenvuo J, Rosenzweig A. Effects of aging on angiogenesis. Circ Res. 2012; 110:1252–64. https://doi.org/10.1161/CIRCRESAHA.111.246116 [PubMed]
- 55. Lee HK, Chauhan SK, Kay E, Dana R. Flt-1 regulates vascular endothelial cell migration via a protein tyrosine kinase-7-dependent pathway. Blood. 2011; 117:5762–71. https://doi.org/10.1182/blood-2010-09-306928 [PubMed]
- 56. Krueger J, Liu D, Scholz K, Zimmer A, Shi Y, Klein C, Siekmann A, Schulte-Merker S, Cudmore M, Ahmed A, le Noble F. Flt1 acts as a negative regulator of tip cell formation and branching morphogenesis in the zebrafish embryo. Development. 2011; 138:2111–20. https://doi.org/10.1242/dev.063933 [PubMed]
- 57. Ikeda T, Sun L, Tsuruoka N, Ishigaki Y, Yoshitomi Y, Yoshitake Y, Yonekura H. Hypoxia down-regulates sFlt-1 (sVEGFR-1) expression in human microvascular endothelial cells by a mechanism involving mRNA alternative processing. Biochem J. 2011; 436:399–407. https://doi.org/10.1042/BJ20101490 [PubMed]
- 58. Katoh O, Tauchi H, Kawaishi K, Kimura A, Satow Y. Expression of the vascular endothelial growth factor (VEGF) receptor gene, KDR, in hematopoietic cells and inhibitory effect of VEGF on apoptotic cell death caused by ionizing radiation. Cancer Res. 1995; 55:5687–92. [PubMed]
- 59. Gerber HP, Condorelli F, Park J, Ferrara N. Differential transcriptional regulation of the two vascular endothelial growth factor receptor genes. Flt-1, but not Flk-1/KDR, is up-regulated by hypoxia. J Biol Chem. 1997; 272:23659–67. https://doi.org/10.1074/jbc.272.38.23659 [PubMed]
- 60. Yamamoto H, Ehling M, Kato K, Kanai K, van Lessen M, Frye M, Zeuschner D, Nakayama M, Vestweber D, Adams RH. Integrin β1 controls VE-cadherin localization and blood vessel stability. Nat Commun. 2015; 6:6429. https://doi.org/10.1038/ncomms7429 [PubMed]
- 61. Theilmann AL, Hawke LG, Hilton LR, Whitford MKM, Cole DV, Mackeil JL, Dunham-Snary KJ, Mewburn J, James PD, Maurice DH, Archer SL, Ormiston ML. Endothelial BMPR2 Loss Drives a Proliferative Response to BMP (Bone Morphogenetic Protein) 9 via Prolonged Canonical Signaling. Arterioscler Thromb Vasc Biol. 2020; 40:2605–18. https://doi.org/10.1161/ATVBAHA.119.313357 [PubMed]
- 62. Zhao W, Shen B, Cheng Q, Zhou Y, Chen K. Roles of TSP1-CD47 signaling pathway in senescence of endothelial cells: cell cycle, inflammation and metabolism. Mol Biol Rep. 2023; 50:4579–85. https://doi.org/10.1007/s11033-023-08357-w [PubMed]
- 63. Zhang X, Wang Y, Fan J, Chen W, Luan J, Mei X, Wang S, Li Y, Ye L, Li S, Tian W, Yin K, Ju D. Blocking CD47 efficiently potentiated therapeutic effects of anti-angiogenic therapy in non-small cell lung cancer. J Immunother Cancer. 2019; 7:346. https://doi.org/10.1186/s40425-019-0812-9 [PubMed]
- 64. Salcedo R, Oppenheim JJ. Role of chemokines in angiogenesis: CXCL12/SDF-1 and CXCR4 interaction, a key regulator of endothelial cell responses. Microcirculation. 2003; 10:359–70. https://doi.org/10.1038/sj.mn.7800200 [PubMed]
- 65. Keglowich L, Roth M, Philippova M, Resink T, Tjin G, Oliver B, Lardinois D, Dessus-Babus S, Gosens R, Hostettler Haack K, Tamm M, Borger P. Bronchial smooth muscle cells of asthmatics promote angiogenesis through elevated secretion of CXC-chemokines (ENA-78, GRO-α, and IL-8). PLoS One. 2013; 8:e81494. https://doi.org/10.1371/journal.pone.0081494 [PubMed]
- 66. Rizzo M, Varnier L, Pezzicoli G, Pirovano M, Cosmai L, Porta C. IL-8 and its role as a potential biomarker of resistance to anti-angiogenic agents and immune checkpoint inhibitors in metastatic renal cell carcinoma. Front Oncol. 2022; 12:990568. https://doi.org/10.3389/fonc.2022.990568 [PubMed]
- 67. Yun HJ, Jo DY. Production of stromal cell-derived factor-1 (SDF-1)and expression of CXCR4 in human bone marrow endothelial cells. J Korean Med Sci. 2003; 18:679–85. https://doi.org/10.3346/jkms.2003.18.5.679 [PubMed]
- 68. Du C, Xu Y, Yang K, Chen S, Wang X, Wang S, Wang C, Shen M, Chen F, Chen M, Zeng D, Li F, Wang T, et al. Estrogen promotes megakaryocyte polyploidization via estrogen receptor beta-mediated transcription of GATA1. Leukemia. 2017; 31:945–56. https://doi.org/10.1038/leu.2016.285 [PubMed]
- 69. Bord S, Frith E, Ireland DC, Scott MA, Craig JI, Compston JE. Synthesis of osteoprotegerin and RANKL by megakaryocytes is modulated by oestrogen. Br J Haematol. 2004; 126:244–51. https://doi.org/10.1111/j.1365-2141.2004.05024.x [PubMed]
- 70. Cui Z, Wu H, Xiao Y, Xu T, Jia J, Lin H, Lin R, Chen K, Lin Y, Li K, Wu X, Li C, Yu B. Endothelial PDGF-BB/PDGFR-β signaling promotes osteoarthritis by enhancing angiogenesis-dependent abnormal subchondral bone formation. Bone Res. 2022; 10:58. https://doi.org/10.1038/s41413-022-00229-6 [PubMed]
- 71. Heldin CH, Westermark B. Mechanism of action and in vivo role of platelet-derived growth factor. Physiol Rev. 1999; 79:1283–316. https://doi.org/10.1152/physrev.1999.79.4.1283 [PubMed]
- 72. Silverstein RL, Febbraio M. CD36, a scavenger receptor involved in immunity, metabolism, angiogenesis, and behavior. Sci Signal. 2009; 2:re3. https://doi.org/10.1126/scisignal.272re3 [PubMed]
- 73. Kanzler I, Tuchscheerer N, Steffens G, Simsekyilmaz S, Konschalla S, Kroh A, Simons D, Asare Y, Schober A, Bucala R, Weber C, Bernhagen J, Liehn EA. Differential roles of angiogenic chemokines in endothelial progenitor cell-induced angiogenesis. Basic Res Cardiol. 2013; 108:310. https://doi.org/10.1007/s00395-012-0310-4 [PubMed]
- 74. Bertolino P, Deckers M, Lebrin F, ten Dijke P. Transforming growth factor-beta signal transduction in angiogenesis and vascular disorders. Chest. 2005; 128:585S–90. https://doi.org/10.1378/chest.128.6_suppl.585S [PubMed]
- 75. Wang J, Li Y. CD36 tango in cancer: signaling pathways and functions. Theranostics. 2019; 9:4893–908. https://doi.org/10.7150/thno.36037 [PubMed]