



Introduction
Cancer, which is currently the leading cause of death in most of the world’s population, has been considered an important factor hindering the increase of human life expectancy [1]. With an ageing and rapidly growing population, as well as accelerating socioeconomic development, the burden of cancer continues to increase in both developed and developing countries [2]. The most recent data estimates that approximately 160 cancer cases will be diagnosed for every 1,889,000 Americans [3]. By 2021, 608,570 Americans will die from cancer, which is equivalent to more than 1,600 deaths per day [3]. Data for China, the world’s most populous country with an estimated 1.42 billion people, suggested that approximately 4.51 million cancer cases and 3.04 million cancer-related deaths had occurred in 2020 [4]. Despite the differing levels of social or economic development, an increase in cancer incidence or mortality represents a great threat to individual health and a significant economic burden for any country and society.
Therefore, a better approach to cancer prevention and detection is clearly needed. Tumour biomarkers have a wide range of promising clinical applications. They can be used for cancer risk assessment, screening, surveillance, diagnosis, predicting treatment response and monitoring disease progression and recurrence [5, 6], while having the potential to become an important component of precision cancer management [7].
Apart from its critical role in female sexual development and reproductive processes, estrogen is also extensively involved in physiological and pathophysiological processes across different tissues in both sexes [8]. It exerts significant influence in carcinogenesis by regulating cell apoptosis, proliferation, and the cell cycle [9–11]. Additionally, it interacts with various cell types within the tumor microenvironment, including fibroblasts, immune cells, and adipocytes [11]. Among the recognized estrogen receptors (ERs), namely ERα, ERβ, and G-protein-coupled estrogen receptor 1 (GPER1), the first two are classical estrogen receptors. GPER1 serves as a receptor for mediating rapid estrogen effects. Encoded by the GPER1 gene, GPER1 is widely expressed in the human body across multiple systems such as reproductive, digestive, cardiovascular, respiratory, nervous, and hematopoietic systems [12, 13]. This receptor binds to estrogen and activates multiple downstream signaling pathways, mediating rapid non-genomic estrogen signaling events. It exerts diverse biological effects in tumor cell proliferation, apoptosis, migration, tumor initiation, and metastasis across various cancers [14]. Recent reports highlight a significant correlation between GPER1 and the progression of diverse cancers. Furthermore, GPER1 is considered a potential therapeutic target for cancer treatment [13, 15]. Despite comprehensive pan-cancer analyses exploring ERs as prognostic markers and therapeutic targets across different cancers [16], a comprehensive pan-cancer analysis of GPER1 is lacking to date. Consequently, this study aims to investigate the diagnostic and prognostic significance of GPER1 expression in the context of pan-cancer.
The expression of GPER1 mRNA and protein in various tissues and organs throughout the body was explored using the Human Protein Atlas (HPA) database. Subsequently, GPER1 expression in tumour tissues was assessed and compared to than in normal and paracancerous tissues using three databases, The Cancer Genome Atlas (TCGA), University of California, Santa Cruz Xena (UCSC XENA) and Genotype-Tissue Expression (GTEx) (https://gtexportal.org/). Promoter methylation of GPER1 was explored using the MethSurv and The University of Alabama at Birmingham CANcer data analysis Portal (UALCAN) databases. Genetic alterations and their associated survival analysis were evaluated via the cBioPortal. Differentially expressed genes (DEGs) related to GPER1 expression, Protein–Protein Interaction (PPI), functional enrichment and Gene Set Enrichment Analysis (GSEA) of DEGs were also explored. We further investigated the relationship between GPER1 and tumour-infiltrating lymphocytes (TILs), immunoinhibitors, immunostimulators, major histocompatibility complex (MHC) molecules, chemokines and chemokine receptors via the TISDB.
Results
Expression landscape and pan-cancer expression of GPER1
According to the results obtained from the HPA database, GPER1 mRNA and protein are widely expressed in various organs and tissues throughout the body (Figure 1A). GPER1 mRNA is expressed primarily in the lungs, stomach, liver, thyroid, adipose tissue, placenta, basal ganglia, amygdala, seminal vesicles, breast, cerebral cortex and adrenal glands (Figure 1B, 1C).
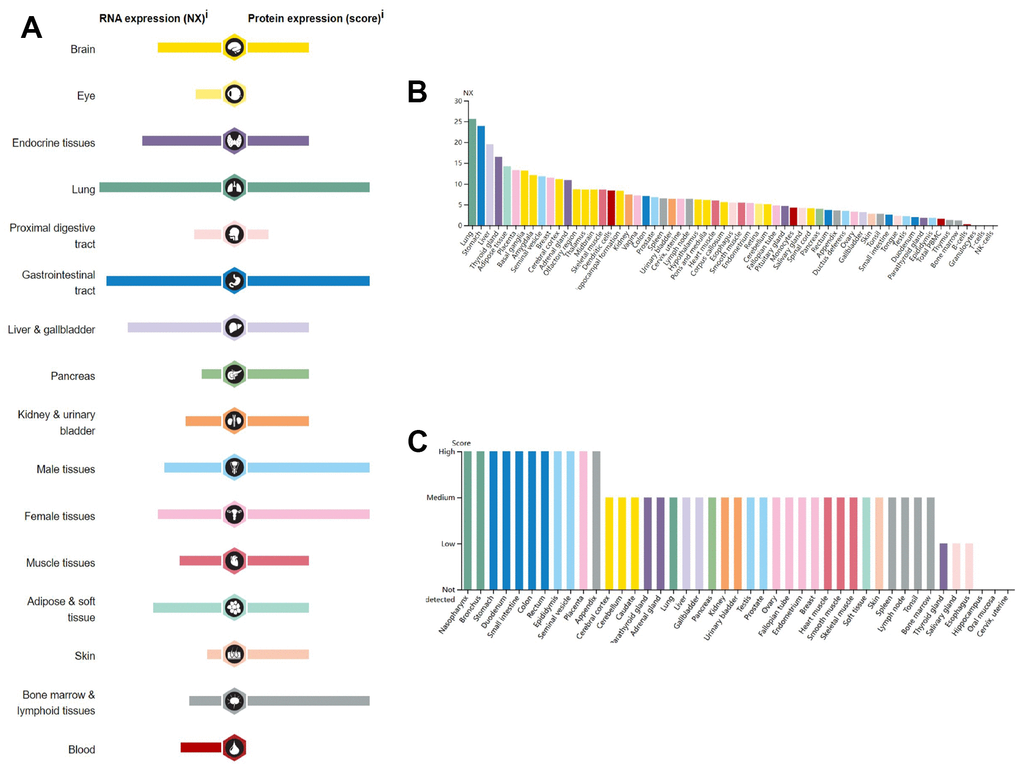
Figure 1. RNA and protein expression profile for GPER1 in different human organs and tissues present by HPA. (A) GPER1 RNA and protein expression summary in different human organs and tissues; Summary of RNA and protein expression information produced within the Human Protein Atlas initiative. Examined tissues are categorized into groups with color-coded distinctions based on shared functional attributes. (B) GPER1 RNA expression summary in different human organs and tissues based on consensus dataset; The unified dataset comprises normalized expression (nTPM) levels for 55 distinct tissue types, achieved through the integration of HPA and GTEx transcriptomics datasets via an internal normalization process. The utilization of color codes corresponds to tissue groupings, with each group comprising tissues sharing common functional attributes. (C) GPER1 protein expression summary in different human organs and tissues. For every one of the 44 tissues, protein expression information is displayed. Color classification is rooted in tissue groups, where each group is composed of tissues that share common functional characteristics.
GPER1 mRNA expression was evaluated in pan-cancer including Adrenocortical carcinoma (ACC), Bladder Urothelial Carcinoma (BLCA), Breast invasive carcinoma (BRCA), Cervical squamous cell carcinoma and endocervical adenocarcinoma (CESC), Cholangiocarcinoma (CHOL), Colon adenocarcinoma (COAD), Lymphoid Neoplasm Diffuse Large B-cell Lymphoma (DLBC), Esophageal carcinoma (ESCA), Glioblastoma multiforme (GBM), Head and Neck squamous cell carcinoma (HNSC), Kidney Chromophobe (KICH), Kidney renal clear cell carcinoma (KIRC), Kidney renal papillary cell carcinoma (KIRP), Acute Myeloid Leukemia (LAML), Brain Lower Grade Glioma (LGG), Liver hepatocellular carcinoma (LIHC), Lung adenocarcinoma (LUAD), Lung squamous cell carcinoma (LUSC), Mesothelioma (MESO), Ovarian serous cystadenocarcinoma (OV), Pancreatic adenocarcinoma (PAAD), Pheochromocytoma and Paraganglioma (PCPG), Prostate adenocarcinoma (PRAD), Rectum adenocarcinoma (READ), Sarcoma (SARC), Skin Cutaneous Melanoma (SKCM), Stomach adenocarcinoma (STAD), Testicular Germ Cell Tumors (TGCT), Thyroid carcinoma (THCA), Thymoma (THYM), Uterine Corpus Endometrial Carcinoma (UCEC), Uterine Carcinosarcoma (UCS), and Uveal Melanoma (UVM). As shown in Figure 2A, unpaired sample analysis found that compared to normal samples, GPER1 mRNA expression was higher in GBM (P = 0.009), LGG (P = 0.002), HNSC, KIRC, LAML and PAAD (all P < 0.001) and lower in ACC, BLCA, BRCA, CESC, CHOL, COAD, ESCA, KICH, LUAD, LUSC, OV, PRAD, READ, PAN-CNACER, STAD, TGCT, THCA, UCEC, UCS (all P < 0.001) and PCPG (P = 0.015). MESO and UVM could not be analysed due to insufficient normal samples. Compared to paracancerous tissue, GPER1 mRNA expressed was significant higher in HNSC and KIRC (both P < 0.001) and significant lower in BLCA, BRCA, CHOL, COAD, ESCA, KICH, LUAD, LUSC, PRAD, READ, STAD, THCA, UCEC (all P < 0.001), CESC (P = 0.023) and PCPG (P = 0.015) (Figure 2B). ACC, DLBC, LAML, LGG, OV, TGCT, UCS, MESO and UVM could not be analysed due to insufficient paracancerous samples. Among the paired sample analyses that can be performed, GPER1 mRNA expression was increased in HNSC and KIRC (both P < 0.001) but was decreased in BLCA, BRCA, COAD, KICH, LUAD, LUSC, PRAD, STAD, THCA, UCEC (all P < 0.001), CHOL (P = 0.004), ESCA (P = 0.023) and READ (P = 0.004) (Figure 2C).
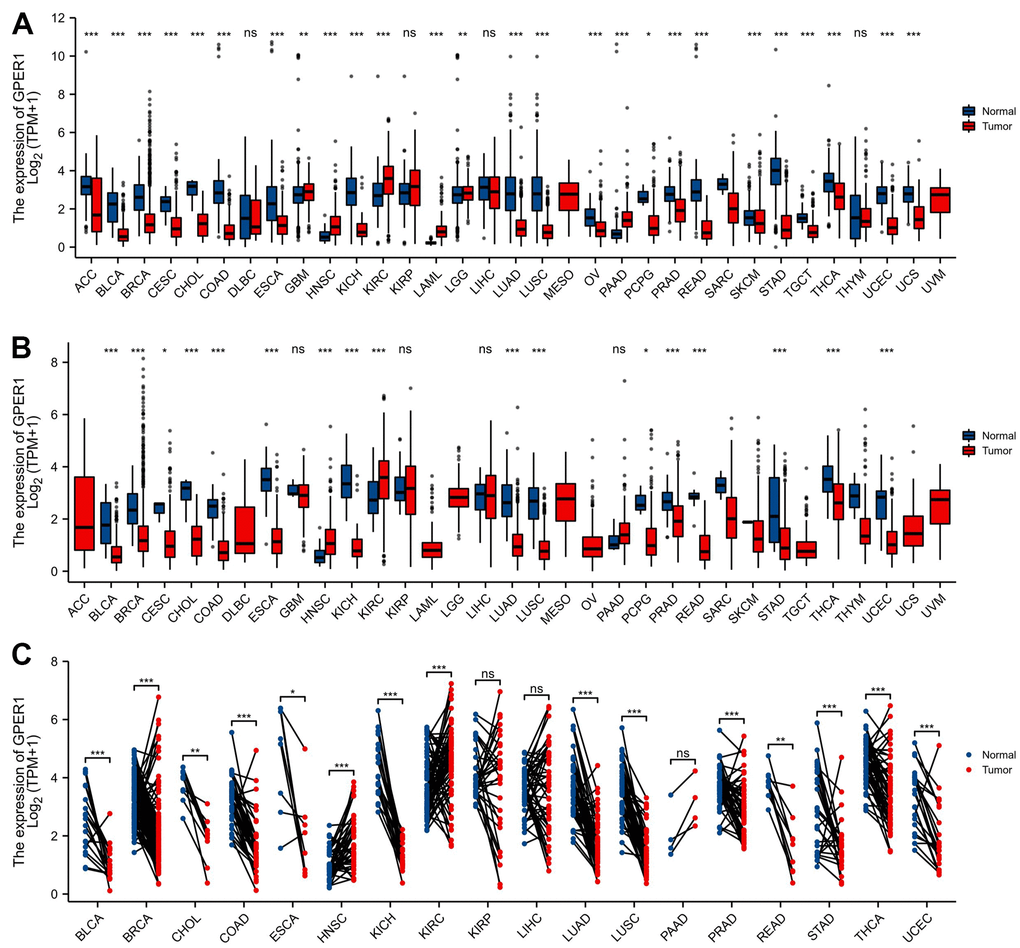
Figure 2. The expression of GPER1 mRNA in pan-cancer. (A) Pan-cancer expression of GPER1 between tumor and normal tissues in unpaired sample analysis; (B) Pan-cancer expression of GPER1 between tumor and paracancerous tissue in unpaired sample analysis; Based on publicly available data, molecular distinctions across diverse pan-cancer datasets are directly analyzed to perform comparative analysis between the tumor group and the normal (adjacent) group. (C) Paired sample analysis of GPER1 between tumor and normal tissues in BLCA, BRCA, CHOL, COAD, ESCA, HNSC, KICH, KIRC, KIRP, LIHC, LUAD, LUSC, PAAD, PRAD, READ, STAD, THCA and UCEC. Each line represents a paired sample, namely the normal (adjacent) versus tumor samples selected from the available public data. The more consistent and inclined the trend direction of the lines, the more pronounced the differences between the two groups. Wilcoxon rank sum test * p < 0:05, ** p < 0:01, *** p < 0:001.
The diagnostic value of GPER1 in pan-cancer
As shown in Figure 3, GPER1 had good diagnostic value in various cancers. Its Area under Curve (AUC) was greater than 0.7 in 21 cancers and even exceeded 0.9 in 9 cancers, including CHOL, COAD, KICH, LAML, LUAD, LUSC, READ, STAD and THYM (Supplementary Table 1), which had high diagnostic value.
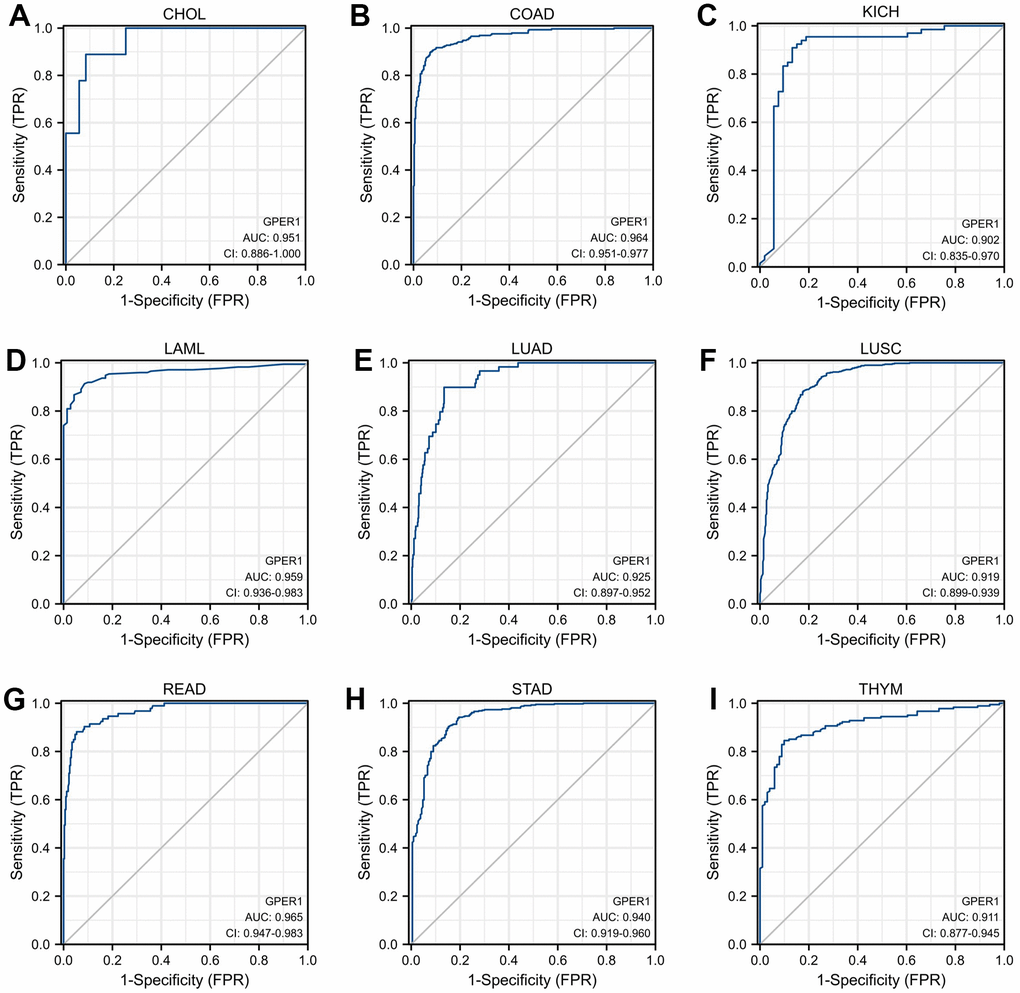
Figure 3. Receiver operator characteristic (ROC) curve of GPER1 in cancers. Cancers with AUC > 0.9 for GPER1: (A) CHOL; (B) COAD; (C) KICH; (D) LAML; (E) LUAD; (F) LUSC; (G) READ; (H) STAD; (I) THYM.
Survival analysis of GPER1 in pan-cancer
For the purpose of evaluating the prognostic value of GPER1 in pan-cancer, Kaplan–Meier (K-M) analysis was conducted. Cox regression analysis of 35 cancers showed that GPER1 expression in 11 cancers was significantly associated with OS (Supplementary Table 2). Our results found that the high GPER1 group had significantly better overall survival (OS) than the low GPER1 group in BRCA (Hazard ratio (HR) 0.69, 95% Confidence Interval (CI): 0.50–0.97; p = 0.03), DLBC (HR 0.09, 95% CI: 0.01–0.79; p = 0.029), ESCA (HR 0.46, 95% CI: 0.28–0.77; p = 0.003), HNSC (HR 0.75, 95% CI: 0.57–0.99; p = 0.042), KIRC (HR 0.59, 95% CI: 0.44–0.80; p = 0.001), KIRP (HR 0.45, 95% CI: 0.25–0.81; p = 0.008), LUAD (HR 0.71, 95% CI: 0.52–0.98; p = 0.036), PAAD (HR 0.59, 95% CI: 0.38–0.93; p = 0.022), SARC (HR 0.38, 95% CI: 0.20–0.70; p = 0.002) and UCEC (HR 0.53, 95% CI: 0.34–0.80; p = 0.003) (Figures 4, 5A–5J). However, the low GPER1 group showed significantly better OS than the high GPER1 group in STAD (HR 1.50, 95% CI: 1.06–2.12; p = 0.023) (Figures 4, 5K).
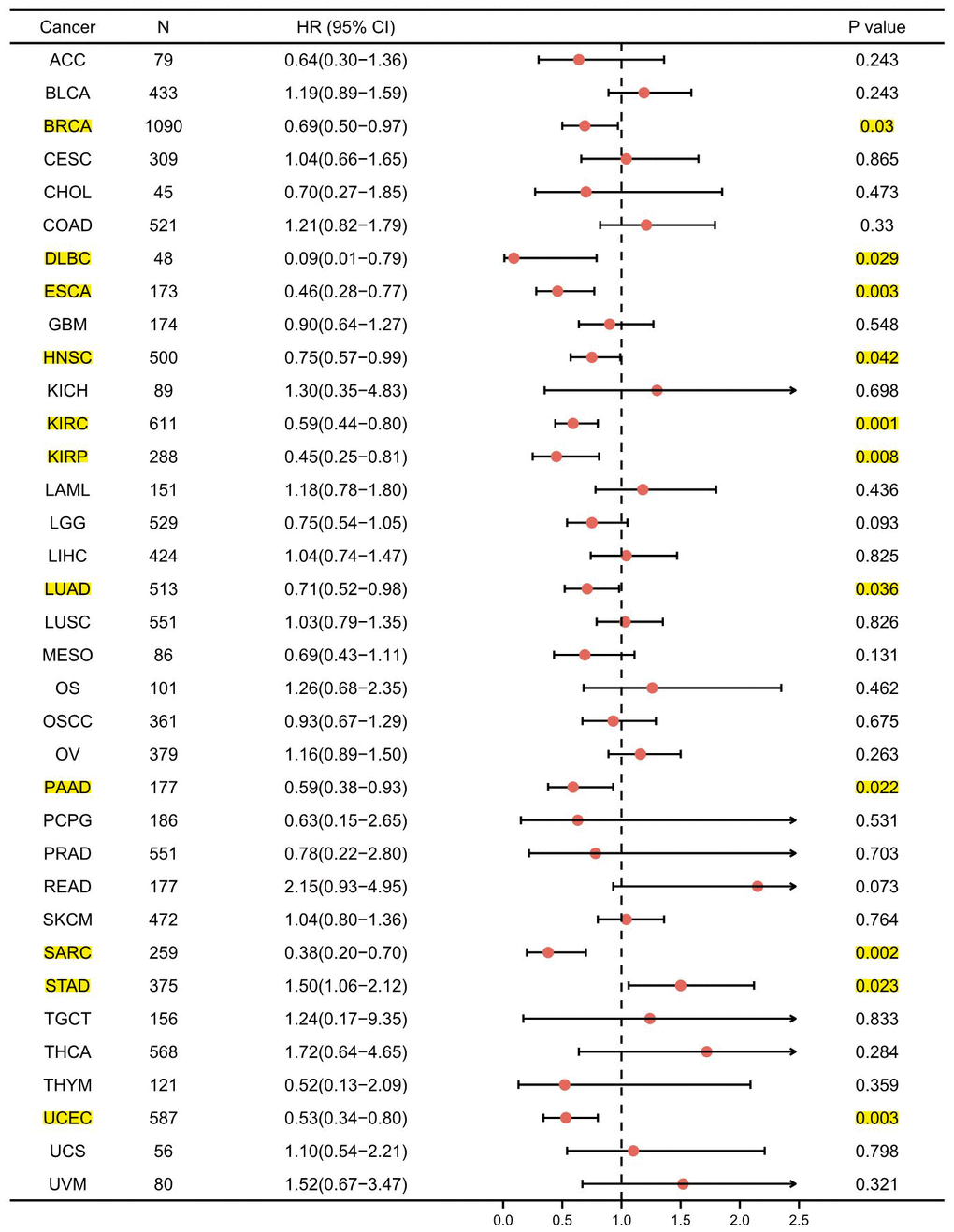
Figure 4. Forest plot of GPER1 OS in 35 cancer types. The marked yellow cancer species indicated that the p-value of prognostic K-M analysis for high- and low- GPER1 gene expression in the cancer species (BRCA, DLBC, ESCA, HNSC, KIRC, KIRP, LUAD, PAAD, SARC, STAD, UCEC) were less than 0.05.
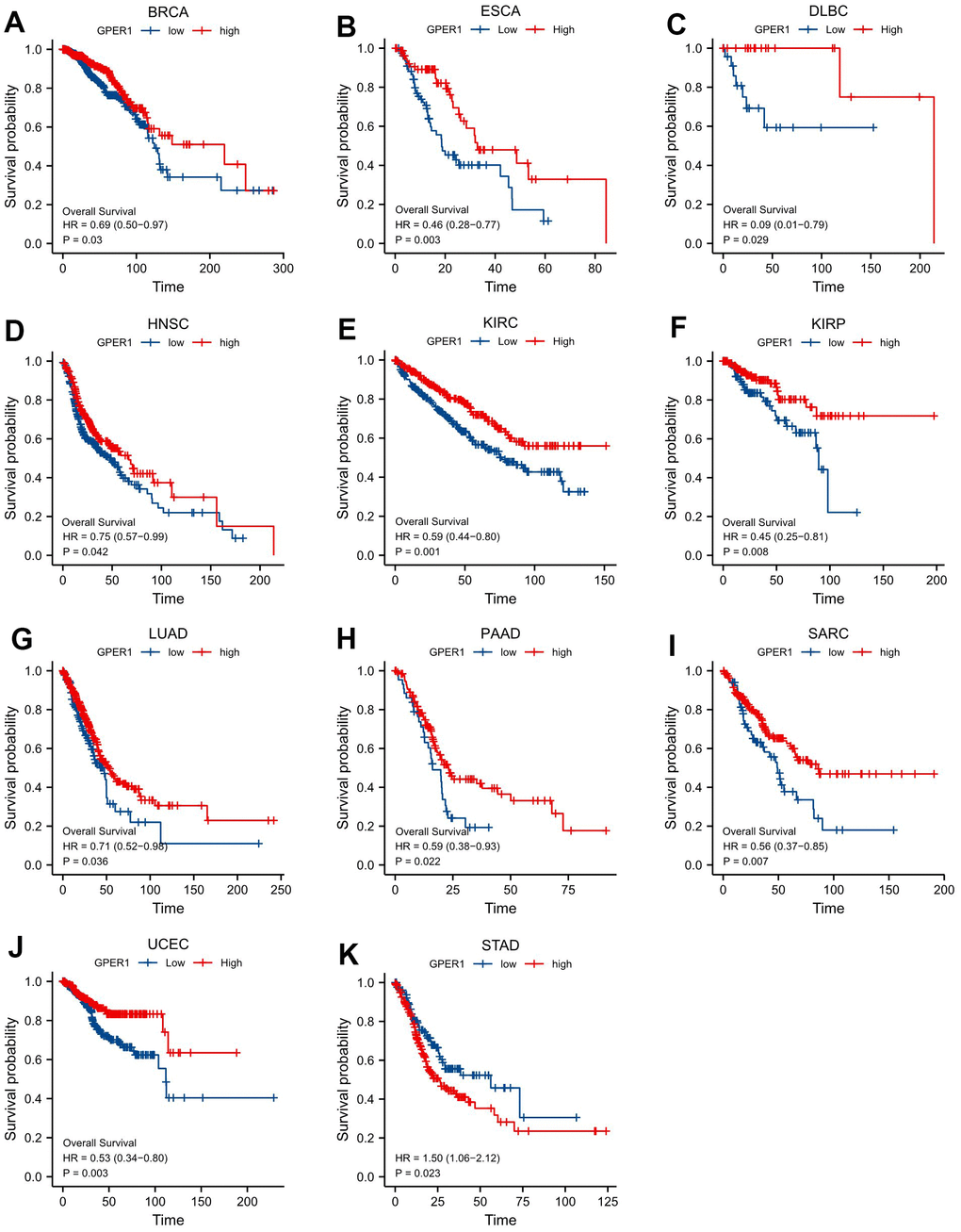
Figure 5. Correlations between GPER1 and prognosis in 11 cancer types. OS K-M curve for GPER1 11 cancer types. The unit of X-axis is month. (A) BRCA, (B) ESCA, (C) DLBC, (D) HNSC, (E) KIRC, (F) KIRP, (G) LUAD, (H) PAAD, (I) SARC, (J) UCEC, (K) STAD.
Genetic alteration of GPER1 in pan-cancer
This study analysed genetic mutations of GPER1 in pan-cancer using the cBioPortal online tool. Based on TCGA, GPER1 mutations were most commonly seen in ESCA, STAD, LUAD, SKCM, DLBC, BLCA and UCEC (Figure 6A). The mutation rate of GPER1 genes was 2.2%, with the most predominant mutation types being Amplification, Deep Deletion and Missense mutation (Figure 6B). The correlation between genetic mutations and prognosis of pan-cancer patients was further explored. Accordingly, GPER1 genetic mutations promoted a significant decrease in OS (Figure 6C), disease-free survival (DFS) (Figure 6D), disease-specific survival (DSS) (Figure 6E) and progression-free survival (PFS) (Figure 6F) (all p < 0.001) in pan-cancer.
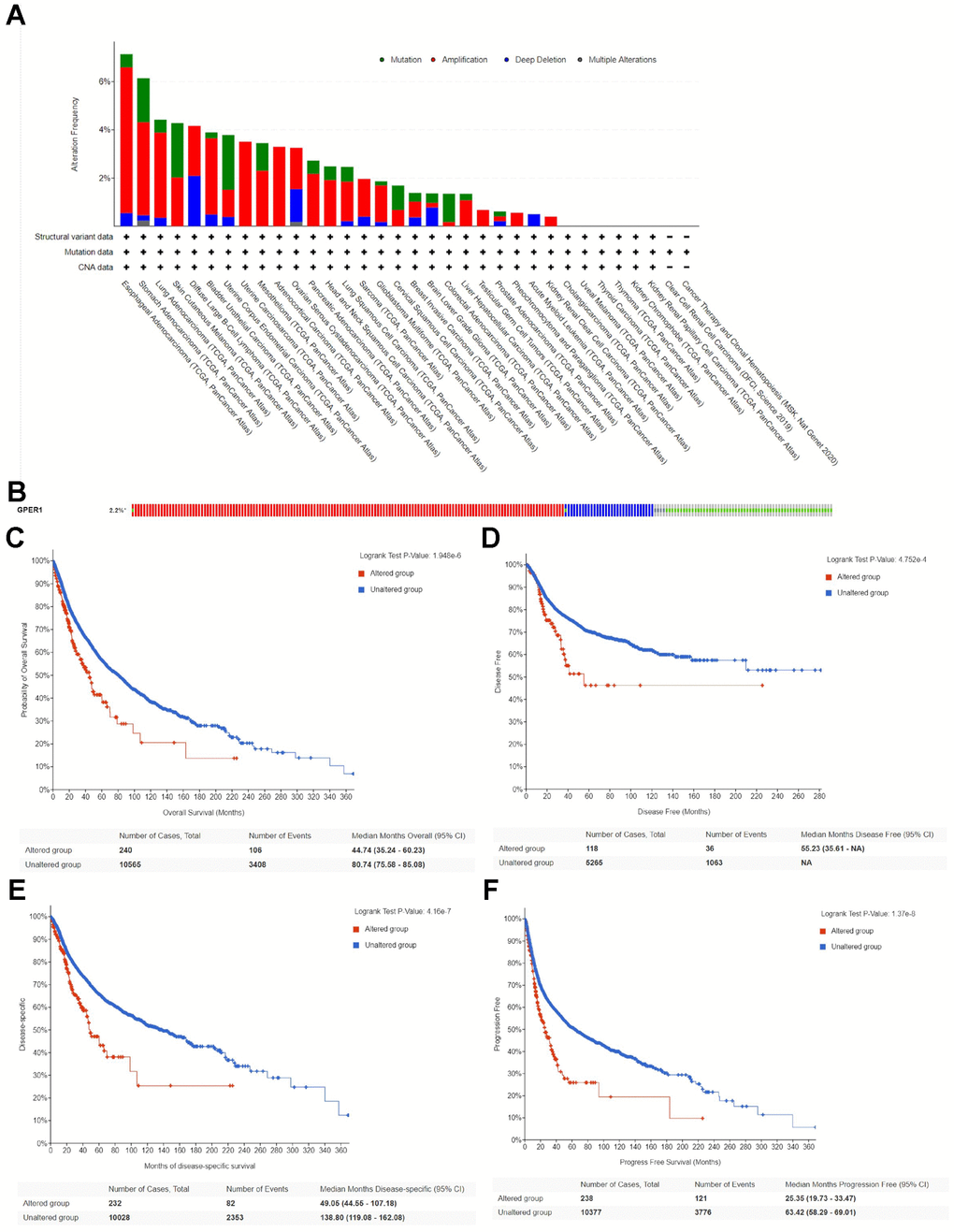
Figure 6. Genetic alteration of GPER1 in pan-cancer. (A) Bar chart of GPER1 mutation in pan-cancer based on TCGA database. (B) The alteration frequency with different types of GPER1 gene mutations in pan-cancer. Kaplan-Meier curve of (C) OS, (D) DSS, (E) DFS, (F) PFS in pan-cancer patients with altered (red) and unaltered (blue) mRNA expression of the GPER1 gene.
DEGs, PPI, functional enrichment and gene set enrichment of GPER1 in cancers
Earlier, we showed that GPER1 expression affected the OS from 11 cancers. To evaluate the biological function of GPER1 in specific cancers, we then analysed differential genes for high and low expression of GPER1 in these cancers and constructed PPI networks with the top 30 up- or downregulated DEGs, as well as performing functional enrichment and gene set enrichment analyses. By analyzing the standardized DEGs in each specific cancer type, the counts of DEGs identified were as follows: BRCA (1117), DLBC (739), ESCA (671), HNSC (837), KIRC (4876), KIRP (2380), LUAD (3960), PAAD (470), SARC (1309), STAD (2228), UCEC (1020). All the DEGs were list in Supplementary File 1.
Our results were presented in a form similar to that in BRCA (Figures 7, 8). The top 30 up- or downregulated DEGs of GPER1 in BRCA are summarised in Figure 7A, 7B, respectively (Supplementary Table 3). The DEGs of GPER1 in BRCA were presented as volcano plots (Figure 7C). Figure 7D presents the PPI networks of the top 30 up- or downregulated DEGs (Supplementary Table 4).
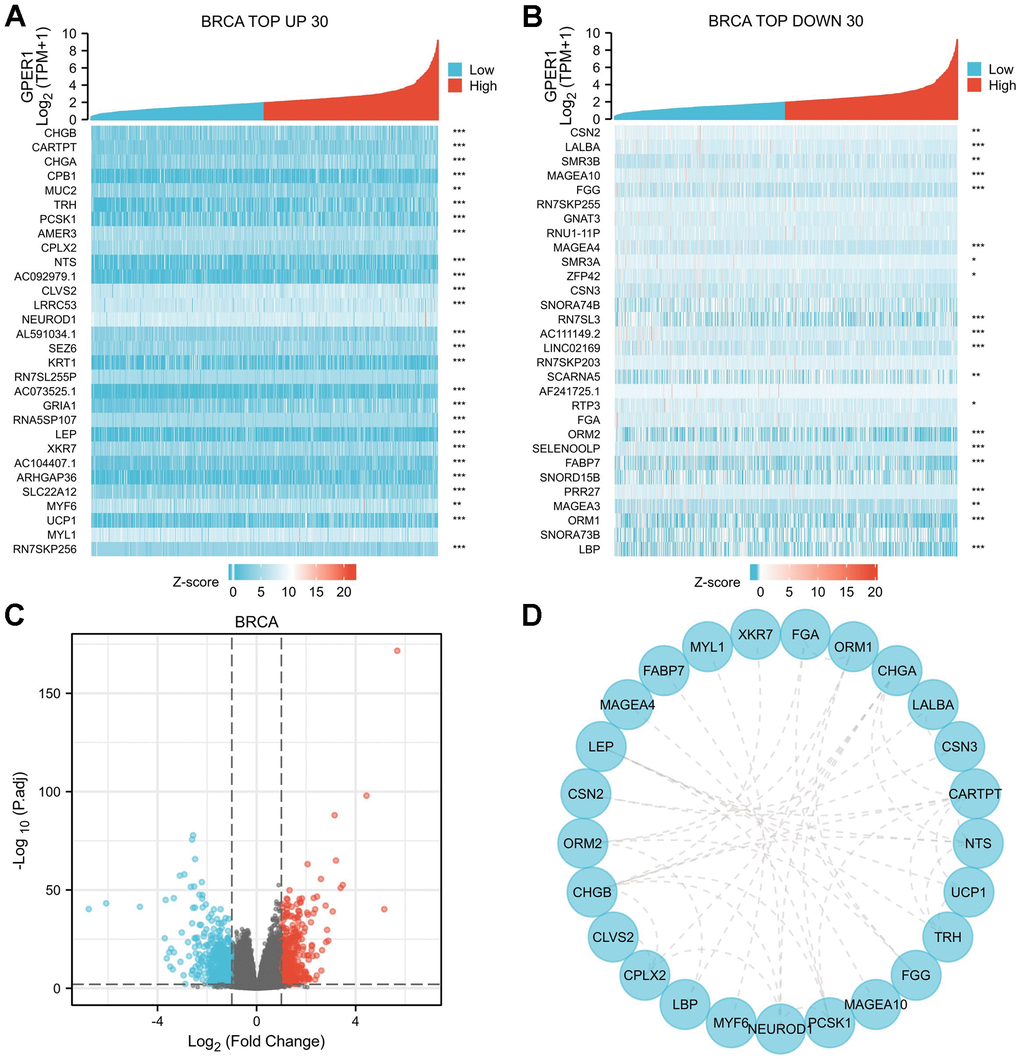
Figure 7. DEGs of high and low GPER1 expression in BRCA and PPI network of DEGs. (A) The heatmap of top 30 up-regulated DEGs, (B) The heatmap of top 30 down-regulated DEGs. Each square represents the expression value of other molecules after undergoing Z-score transformation across various samples (Z-score involves subtracting the mean expression value of each molecule in individual samples from its mean expression value across all samples and then dividing by the standard deviation), with color intensity indicating the absolute value of the expression level. (C) The volcano plots of DEGs between high and low GPER1 expression groups, (D) PPI network of DEGs of high and low GPER1 expression in BRCA. * p < 0.05, ** p < 0.01, *** p < 0.001. The “p” value represents the p-value obtained from the Spearman test conducted to calculate the correlation coefficient between GPER1 and the top 30 up- and downregulated genes.
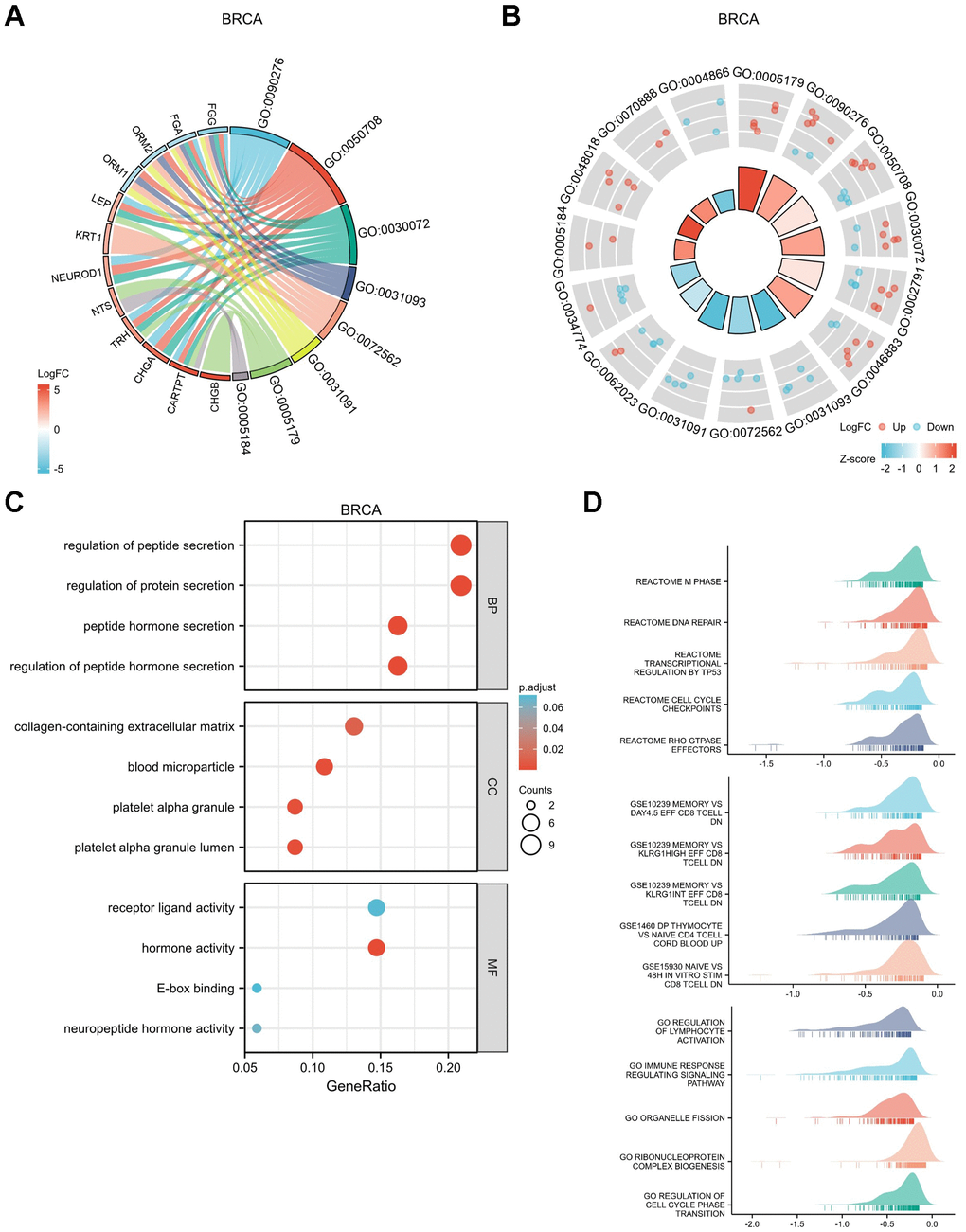
Figure 8. Functional enrichment analysis for DEGs between High and Low expression of GPER1 expression in BRCA. (A) GO/KEGG pathway enrichment joint logFC for DEGs between High and -Low expression of GPER1 expression in BRCA presented as string graph. The left half of the figure shows the gene blocks, and the different colors of the blocks represent the corresponding logFC values. The right half of the graph shows the entry blocks, the size of the blocks represents the corresponding Counts, and the lines (strings) between the blocks on the left and right half of the graph represent the molecules contained in the entry, the presence of the lines means that the entry contains the corresponding molecules. (B) GO/KEGG joint logFC results presented as circle graph. The circle diagram can be divided into two parts: the inner circle and the outer circle. Each bar in the inner circle corresponds to an entry, and the height is the relative size of the p.adj. The higher the bar, the smaller the p.adj of the ID. The color of the corresponding filled column represents the Zscore value of the entry. (C) GO/KEGG pathway enrichment presented as bubble chart, (D) GSEA of the signaling pathways associated with DEGs of GPER1 expression in BRCA.
The top 30 up- or downregulated DEGs were used to perform Gene Ontology (GO) / Kyoto Encyclopedia of Genes and Genomes (KEGG) joint logFC analyses (Supplementary Table 5). The results were presented as string (Figure 8A) and circle graphs (Figure 8B). The different RNA functions of DEGs can be divided into three categories: biological process (BP), molecular function (MF) and cellular component (CC). The top three GO terms for the BP in BRCA included regulation of peptide hormone secretion, regulation of protein secretion and peptide hormone secretion; those for CC included platelet alpha granule lumen, blood microparticle and platelet alpha granule; and those for MF included hormone activity and neuropeptide hormone activity. As shown in the bubble chart demonstrating the results for GO/KEGG analyses (Figure 8C and Supplementary Table 6), the top BP terms in BRCA included regulation of peptide hormone secretion, peptide hormone secretion, regulation of hormone secretion and signal release; those for CC included mast cell granule, axon terminus and neuron projection terminus; and those for MF included hormone activity and neuropeptide hormone activity.
Figure 8D shows the GSEA results for DEGs in BRCA. The top five enrichments in biological pathways were REACTOME M PHASE, REACTOME DNA REPAIR, REACTOME TRANSCRIPTIONAL REGULATION BY TP53, REACTOME CELL CYCLE CHECKPOINTS and REACTOME RHO GTPASE EFFECTORS. The top five enrichments in GO were REGULATION OF LYMPHOCYTE ACTIVATION, IMMUNE RESPONSE REGULATING SIGNALLING PATHWAY, ORGANELLE FISSION, RIBONUCLEOPROTEIN COMPLEX BIOGENESIS and REGULATION OF CELL CYCLE PHASE TRANSITION. The top five enrichments in immunologic signatures were GSE10239 MEMORY VS DAY4.5 EFF, CD8 TCELL DN, GSE10239 MEMORY VS KLRG1HIGH EFF CD8 TCELL DN, GSE10239 MEMORY VS KLRG1INT EFF CD8 TCELL DN, GSE1460 DP THYMOCYTE VS NAIVE CD4 TCELL CORD BLOOD UP and GSE15930 NAIVE VS 48H IN VITRO STIM CD8 TCELL DN.
The results for other cancers, including DLBC (Supplementary Figures 1, 2), ESCA (Supplementary Figures 3, 4), HNSC (Supplementary Figures 5, 6), KIRC (Supplementary Figures 7, 8), KIRP (Supplementary Figures 9, 10), LUAD (Supplementary Figures 11, 12), PAAD (Supplementary Figures 13, 14), SARC (Supplementary Figures 15, 16), STAD (Supplementary Figures 17, 18) and UCEC (Supplementary Figures 19, 20), were presented in a form similar to that for BRCA.
Methylation level of GPER1 in cancers
Gene methylation is closely associated with the development and progression of several cancers. This study obtained the methylation data of GPER1, which have been considered significant in the survival analysis with corresponding normal tissues, for 10 cancers using the MethSurv database. The database does not include information on the methylation of GPER1 in DLBC. The methylation information of GPER1 for 10 cancers were presented as heatmaps in Supplementary Figure 21A–21J.
We further compared the GPER1 methylation levels among the identified cancers. Except for DLBC, methylation data of GPER1 for 10 cancers and normal tissues were obtained. Our findings showed that the methylation level of GPER1 was significantly higher in BRCA (p < 0.001), ESCA (p < 0.001), HNSC (p < 0.05), LUAD (p < 0.005) and UCEC (p < 0.001) compared to that in normal tissues. In contrast, the methylation level of GPER1 was significantly lower in KIRC (p < 0.001), KIRP (p < 0.001) and PAAD (p < 0.05) compared to that in normal tissues. No differences were found between SARC and STAD tissues and corresponding normal tissues (Figure 9A–9J).
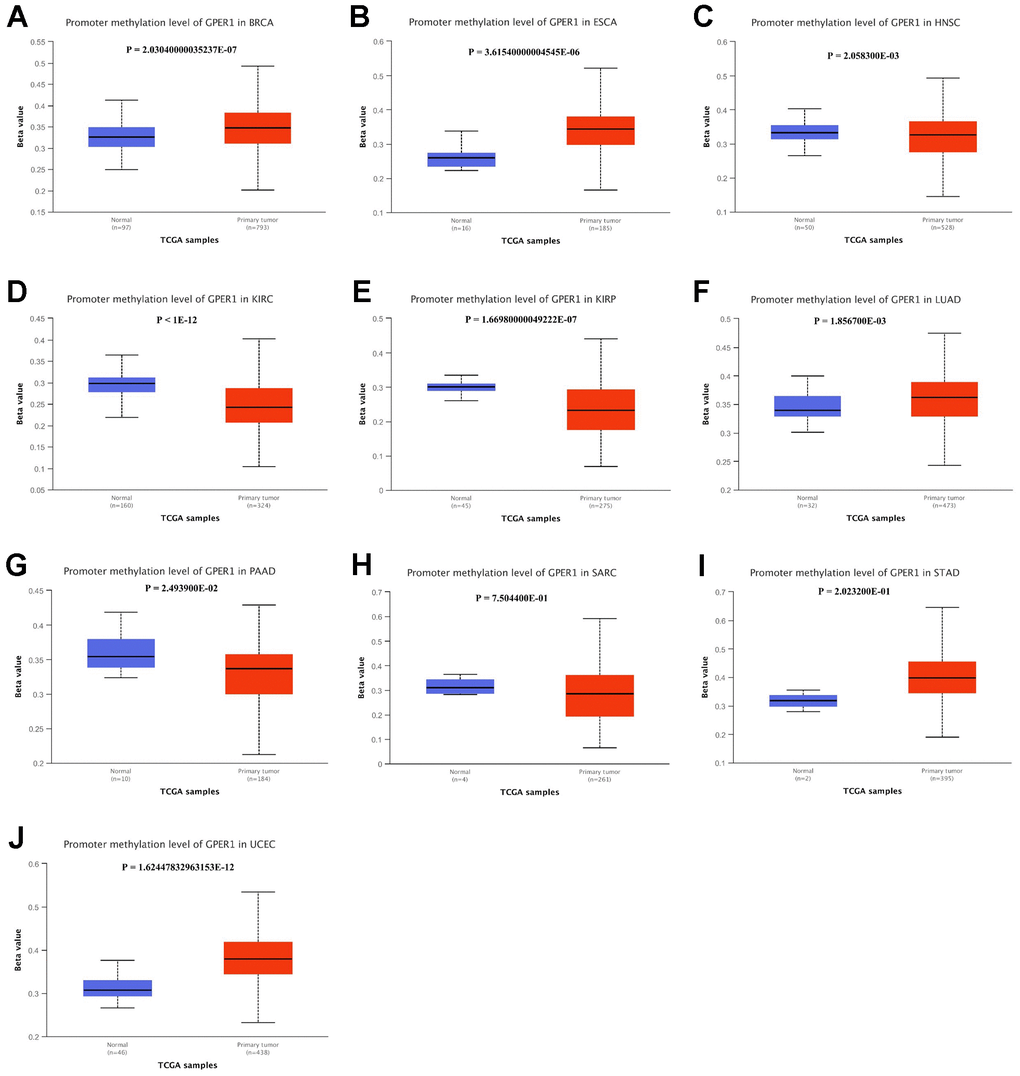
Figure 9. Promoter methylation level of GPER1 between 10 types of cancer and normal tissue. (A) BRCA, (B) ESCA, (C) HNSC, (D) KIRC, (E) KIRP, (F) LUAD, (G) PAAD, (H) SARC, (I) STAD, (J) UCEC.
Pan-cancer immunogenomic analyses of GPER1
The correlation between GPER1 expression level and immune components in pan-cancer were inferred via the TISIDB database. The relationship between GPER1 and TILs (Figure 10A), immunoinhibitors (Figure 10B), immunostimulators (Figure 10C), MHC molecules (Figure 10D), chemokines (Figure 10E) and chemokine receptors (Figure 10F), as well as that between methylation level of GPER1 and immune components (Figure 11A–11F), were presented as heatmaps.
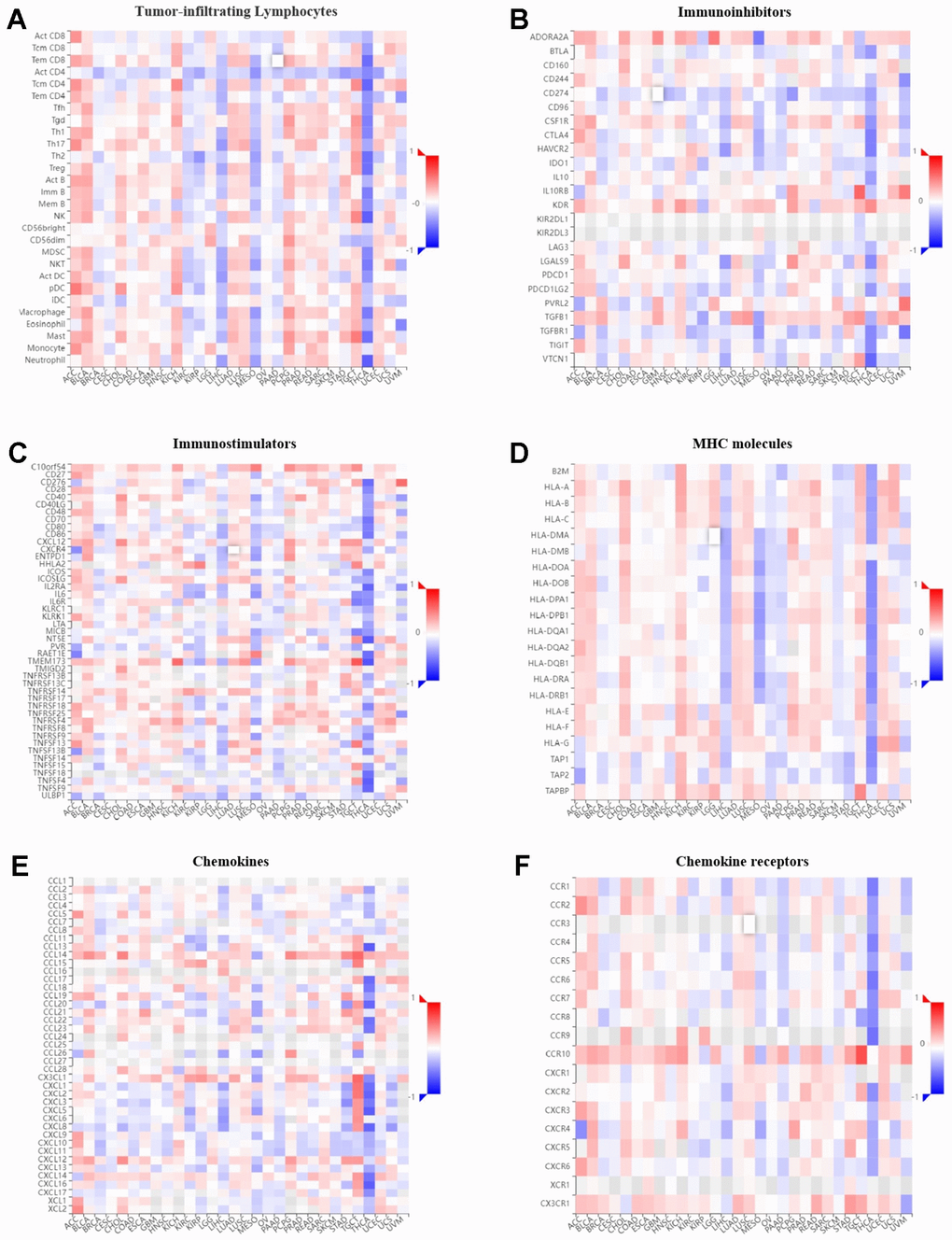
Figure 10. Correlation of GPER1 with TILs and immunoregulation-related genes in pan-cancers. Correlations between GPER1 and (A) TILs, (B) immunoinhibitors, (C) immunostimulators, (D) MHC molecules, (E) Chemokines, (F) Chemokine receptors.
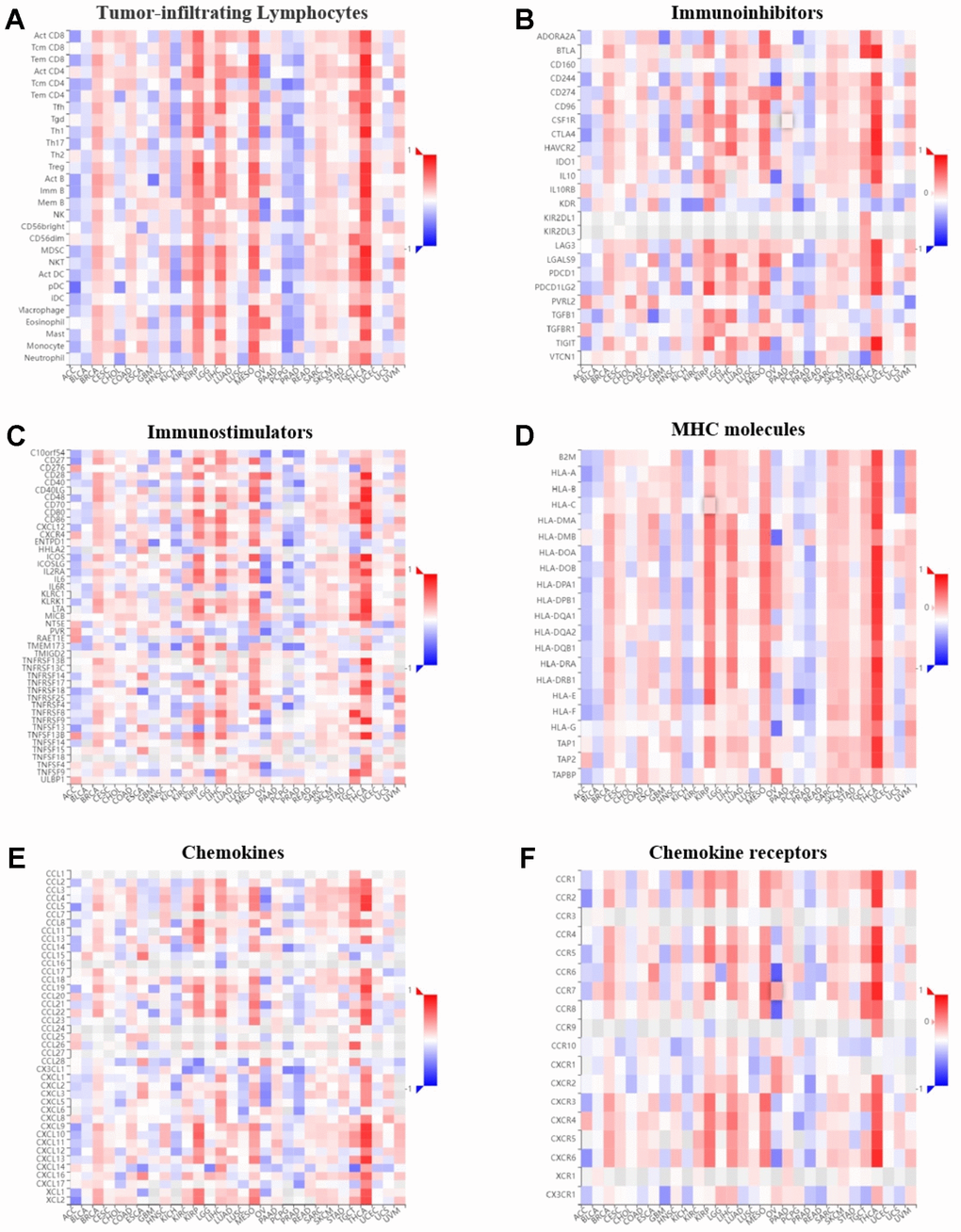
Figure 11. Correlation of GPER1 DNA methylation with TILs and immunoregulation-related genes in pan-cancers. Correlations between GPER1 DNA methylation and (A) TILs, (B) immunoinhibitors, (C) immunostimulators, (D) MHC molecules, (E) Chemokines, (F) Chemokine receptors.
Our findings showed that GPER1 expression was positively correlated with TILs in majority cancers. However, we found that GPER1 expression was negatively correlated with TILs in KIRC, KIRP, LIHC, MESO, PAAD, SKCM and THCA. Of interest was the negative correlation between GPER1 and act CD4 in most cancers. Our results also found that GPER1 expression was negatively correlated with TILs, immunoinhibitors, immunostimulators, MHC molecules, chemokines and chemokine receptors in LIHC, MESO, PAAD and THCA. In particular, the negative correlation in THCA was greater than that in other cancers. The correlation between GPER1 methylation and immune components differed from that between GPER1 and immune components. The positive correlation in ACC, BLCA, KICH, PCPG and PRAD turned out to be a negative correlation, whereas the negative correlation in KIRP, LIHC, MESO and THCA turned out to be a positive correlation.
Discussion
Cancer, a leading cause of global mortality, demands improved prevention and treatment. Tumor biomarkers offer versatile clinical applications. Estrogen’s roles extend beyond reproduction, impacting physiology and carcinogenesis through regulating apoptosis, proliferation, and tumor microenvironment interactions. GPER1, alongside classical estrogen receptors, is expressed widely, mediating rapid estrogen effects across various systems. It affects tumor processes, highlighting its therapeutic potential in cancer. However, comprehensive pan-cancer analysis of GPER1 remains lacking. This study comprehensively investigated the multifaceted role of GPER1 across various cancers. The study delved into GPER1’s expression, diagnostic potential, survival implications, epigenetic regulation, genetic alterations, functional significance, and immunogenomic interactions. GPER1’s mRNA and protein were found to be widely expressed in diverse tissues, forming the basis for pan-cancer analyses. These analyses unveiled its diagnostic potential, as elevated expression was observed in specific cancers such as GBM, LGG, and HNSC, while decreased expression characterized others. Interestingly, GPER1 expression correlated with diverse overall survival outcomes in distinct cancer types. Furthermore, the study explored GPER1’s epigenetic landscape, highlighting methylation patterns that exhibited heterogeneity across cancers, contributing to its regulatory complexity. Genetic mutations, encompassing amplifications and deletions, were associated with distinct survival profiles. Functional analyses provided insights into GPER1’s potential roles in various pathways. The investigation extended to GPER1’s intricate relationship with the immune microenvironment, revealing both positive and negative correlations with immune components in different cancers.
Timely diagnosis holds paramount importance in the realm of cancer prevention and treatment. Furthermore, the correlation between early diagnosis and prompt therapeutic intervention has been well-established, demonstrating a substantial enhancement in the survival rates across various malignancies [6]. Therefore, identifying tumour markers with high diagnostic value is imperative. Our results showed that GPER1 is widely expressed in various organs and tissues, both in mRNA and protein. Furthermore, the expression of GPER1 differs significantly between normal or paracancerous tissues in numerous cancers, providing easy access to samples for clinical diagnosis. Our findings also showed that the AUC of GPER1 Receiver operator characteristic (ROC) was more than 0.7 in 21 cancers, indicating that GPER1 had a wide diagnostic efficacy in cancer. Notably, its AUC exceeded 0.9 in nine cancers, suggesting that GPER1 has great detective ability and reliable efficiency, allowing its use as a diagnostic biomarker in these cancers.
GPER1 is involved in regulating various tumours, such as breast, pancreatic, oesophageal, endometrial, ovarian, cervical, prostate and testicular cancers, as well as lung, liver, thyroid, colorectal and kidney cancers [13]. Although an increasing number of studies have focused on the role of GPER1 in different types of cancers, it remains controversial whether GPER1 plays a pro- or anti-cancer role in tumours. Several studies have shown that activation of GPER1 can promote carcinogenesis, whereas others have shown that its activation can suppress tumours (reviewed in [14]). Neither ex vivo nor in vivo experiments have so far led to definitive conclusions. Our results showed that low GPER1 expression was associated with poor prognosis in BRCA, DLBC, ESCA, HNSC, KIRC, KIRP, LUAD, PAAD, SARC and UCEC, whereas high GPER1 expression was associated with poor prognosis in STAD. Evidence has suggested that GPER1 may be a prognostic predictive marker for these cancers. Although our results had been derived from the aggregation of multiple studies’ samples in the TCGA database, the limited number of individual study samples may explain the inconsistency within our results derived from individual studies. Moreover, the results of some experiments had been derived from cell lines that may differ from the results of the primary tumour.
GPER1 plays an important biological role in regulating oestrogenic responses in breast malignancies and has been associated with increased tumour size, increased risk of recurrence and metastasis, decreased survival and therapy resistance in breast cancer patients [17]. However, additional studies have reported that GPER1 inhibits breast cancer proliferation, progression and tumour angiogenesis [18–20]. Study concerning endometrial cancer had found that GPER1 expression was reduced in endometrial cancer cell lines, which is consistent with our results [21]. The GPER1 agonist G1 dose-dependently inhibited the growth of GPER1-positive cell lines RL95-2 and HEC-1A, whereas the GPER1-negative cell line HEC-1B was not affected [21]. This indicates that G1 requires only a moderate amount of GPER1 to exert growth inhibitory effects. This also suggests that the effects of GPER1 on tumours may not depend on the amount of expression but rather in the activation. In gastric cancer, GPER1 promotes gastric cancer proliferation, migration and invasion through PI3K/akt-mediated EMT [22]. This is consistent with our results showing that high GPER1 levels are associated with poor prognosis in STAD. GPER1 agonist G1 increased the number of tumour nodules, tumour grade and tumour index in urethane-induced lung adenocarcinoma models [23]. However, another study reported that G1 can mediate anti-proliferative and pro-apoptotic effects of oxidants and antioxidant molecules on A549 cells [24] and that GPER1 activation can also inhibit migration of human NSCLC cells by suppressing IKK-β/NF-κB signalling [25]. Furthermore, the activation of GPER1 had also been found to inhibit the migration and invasion of osteosarcoma cells through FBXL5-mediated post-translational downregulation of Snail [26]. In pancreatic cancer, high GPER1 expression has been associated with improved survival [27], and GPER1 activation leads to peritumoral mesenchymal remodelling in PDAC, reducing fibrous tissue proliferation, inflammation and immunosuppression [28].
Methylation of gene promoter regions can lead to gene transcriptional repression, and aberrant gene methylation may contribute to oncogenic transformation [29]. Our study findings demonstrate that across the 10 studied cancer types, GPER1 exhibits high methylation in 5 cancers (BRCA, ESCA, HNSC, LUAD, and UCEC) and low methylation in 3 cancers (KIRC, KIRP, and PAAD). We observe decreased GPER1 expression in BRCA, ESCA, LUAD, and UCEC compared to normal or adjacent tissues, while it is elevated in KIRC, KIRP, and PAAD, consistent with our methylation analysis results. In HNSC, GPER1 expression results do not align with methylation outcomes. Methylation analysis reveals minimal differences in median BETA values between HNSC and LUAD, suggesting uncertain biological significance despite potential statistical disparities. This might elucidate the discrepancy between elevated GPER1 expression and unexpectedly “higher” DNA methylation in HNSC compared to normal tissue. GPER1 shows no significant differences in methylation compared to normal tissue in two cancers (SARC and STAD). GPER1 exhibits lower expression in STAD, contradictory to survival analysis indicating better OS in the low GPER1 subgroup of STAD. This complexity might suggest intricate epigenetic regulation of GPER1, with DNA methylation potentially holding a dominant role while histone modifications might also play a crucial role. This also implies the intricate involvement of GPER1 in cancer development. Whether GPER1 promotes or suppresses cancer lacks a definite conclusion, as GPER1 methylation involves a series of subsequent changes, encompassing downstream signaling pathways and immune modulation [13].
Gene mutations affect not only cancer development but also cancer progression. Loss of homozygous ancestral genotype GG is more common in two polymorphisms (rs3808350 and rs3808351) in the GPER1 promoter region of spermatocytomas but not in non-seminomas [30]. The T allele of the GPER1 gene SNP rs11544331 triggers the expression of the P16L variant, which promotes the migration of breast cancer cells [31]. Our study found that GPER1 gene mutations were associated with poor prognosis in patients with tumours. In all samples from patients with tumour, GPER1 gene mutations decreased OS, DFS, DSS and PFS, suggesting that GPER1 gene alterations play an important role in cancer progression and that the associated changes in GPER1 expression levels could provide prognostic value for patients with tumour. However, this also implies that GPER1 itself plays a key role in the development and progression of several cancers and that mutations can cause failure or alteration of this role.
Genes and proteins associated with differential expression of target genes may be associated with specific biological functions or pathways. Analysis of the associated genes or proteins can help us better determine the mechanism of action of the target genes in the disease. Among the 11 cancers selected for analysis, high or low GPER1 expression showed a significant effect on OS. Unlike other pan-cancer analyses, our study analysed the target genes in each specific cancer species. Thus, the biological pathways obtained during subsequent enrichment analysis were closely associated with that specific cancer species.
Immune components, including TILs, immune activators, immunosuppressors, MHC, chemokines and chemokine receptors, are important components of tumour immunity. Our results found that GPER1 was associated with gene expression of these immune components various cancers. In some cancers, GPER1 was correlated with immune components with some consistency. For instance, GPER1 was negatively correlated with immune components in LIHC, MESO and THCA, suggesting its involvement in the immune infiltration of these tumours and the composition of the tumour microenvironment. However, this also indicates the complexity of the role played by GPER1. The negative or positive correlation between GPER1 and both immunosuppressive and immunostimulatory factors may explain why no conclusive conclusion can be reached on whether GPER1 is cancer-promoting or -suppressing despite the numerous studies. This complexity is also compounded by differences in the genetic correlation between GPER1 and the same immune component among various tumours, suggesting that GPER1 has different effects on tumour immunity in different cancers. Our results also found that the correlation between GPER1 and immune component-related genes was altered after methylation. For instance, the negative correlation in LIHC, MESO and THCA changed to a positive correlation, further suggesting that GPER1 is closely associated with the tumour immune microenvironment and ligand-receptor interactions between lymphocytes and malignant tumour cells, potentially influencing tumour progression and prognosis.
Recent years of research accumulation have progressively unveiled the multifaceted association between GPER1 and various aspects of cancer pathogenesis, further accentuating its potential as a therapeutic target for cancer. Consequently, the development of cancer treatment strategies targeting GPER1 has garnered significant attention [32]. The study by Wegnera et al. [33] revealed that GPER1 overexpression reduces proliferation and mitochondrial activity in MCF-7 breast cancer cells, concurrently inducing autophagy. However, this also diminishes MCF-7 cell sensitivity to doxorubicin while augmenting the cytotoxic effects of cyclophosphamide. Additionally, the application of fumaric acid ester further enhances the cytotoxic impact of these substances on GPER1-overexpressing cells. On a different note, research by Sathya et al. [19] indicated that under low oxygen conditions, estrogen suppresses breast cancer growth via the GPER1/ROS/p38 MAPK/p21 signaling pathway. Weißenborn’s findings [34, 35] demonstrated that the GPER1-specific agonist G-1 activates GPER1 in a concentration-dependent manner, effectively inhibiting breast cancer cell growth. This suggests that cell surface-expressed GPER1 holds promise as a potential therapeutic target for non-triple-negative and triple-negative breast cancers. Recent meta-analysis results [36] correlate elevated GPER1 mRNA expression with improved survival rates in breast cancer patients. Furthermore, studies have indicated that GPER1 can inhibit tumor formation and metastasis in cervical cancer cells; reducing GPER1 expression may strengthen cervical cancer cell stemness and migration/invasion capabilities [37]. Moreover, Xu et al.’s research [22] demonstrated that silencing the GPER1 gene can inhibit gastric cancer cell proliferation, migration, and invasion by suppressing the PI3K/AKT-mediated EMT process. These research outcomes align with our findings of elevated GPER1 expression being correlated with adverse prognosis in STAD, indicating its potential as a therapeutic target for gastric cancer treatment [22].
Conclusions
In summary, pan-cancer analysis of GPER1 in our study showed that it was widely expressed in human tissues and organs and that its expression differs from normal tissue in various cancers. The methylation, mutation and mutation-related prognosis of GPER1 in cancers, the associated pathways in specific cancers and its extensive correlation with immune components suggest that GPER1 may have a bright future in the diagnosis, and prognosis of multiple tumours, providing new concepts for precise and personalised anti-tumour strategies.
Materials and Methods
GPER1 expression and datasets obtained
A summary on GPER1 RNA and protein expression in humans was obtained from HPA (https://www.proteinatlas.org/). GPER1 RNA expression was presented as consensus datasets created by combining data from the three transcriptomics datasets (HPA, GTEx and FANTOM5) using the internal normalisation pipeline.
GPER1 mRNA expression of tumour samples and corresponding paracancer samples were determined using TCGA (https://cancergenome.nih.gov), UCSC XENA (https://xenabrowser.net/datapages/) and GTEx (https://gtexportal.org/). Samples with ‘0’ values for gene expression were excluded. The analysis involves the direct examination of molecular distinctions in various pan-cancer datasets, enabling a comparative analysis between tumor and normal (adjacent) groups. The Wilcoxon rank sum test is employed for statistical analysis. In cases where a group consists of fewer than three observations or exhibits a standard deviation greater than zero, said groups will be excluded from the statistical analysis.
Paired samples were retained for paired sample analysis. The analysis is conducted by directly assessing molecular variations across diverse pan-cancer datasets, specifically targeting the differential analysis between tumor samples and adjacent or normal tissue groups within samples exhibiting paired relationships. Statistically, it is stipulated that each group of samples must comprise a minimum of three observations and possess a non-zero variance; failure to meet these conditions will result in the exclusion of said groups from the statistical analysis.
RNA sequencing data in Fragments Per Kilobase per million format were converted and normalised as transcripts per million reads using the Toil process and log2 transformed for further analysis [38]. The statistical analysis employs the Wilcoxon rank sum test. R software was used to perform statistical analyses in this study (version 3.6.3). The ‘ggplot2’ package was used to present GPER1 gene expression as bar graphs in patients with pan-cancer.
ROC curve of GPER1 in pan-cancer
ROC curves were used to estimate the diagnostic value of GPER1 in pan-cancer. ROC curves were calculated using the package ‘pROC’ (version 1.17.0.1) of R software and plotted by package ‘ggplot2’ (v3.3.3). The AUC, cutoff, sensitivity, specificity, positive predictive value, negative predictive value and Youden’s index (YI) were also calculated [39]. An AUC closer to 1 indicates better diagnostic value. Accordingly, an AUC of 0.5 to 0.7, 0.7 to 0.9 and 0.9 or more indicates low, good and high accuracy, respectively [40]. YI indicates the total ability of the screening method to detect real patients from non-patients, with a larger index indicating a more valid and true screening method [41, 42].
Survival analysis of GPER1 in pan-cancer
The ‘survival’ package was used to conduct K-M analysis. The patients with corresponding cancers in the TCGA database were divided into “high” and “low” expression level groups based on the median expression level of GPER1. The OS rates in the high and low GPER1 gene expression groups were compared across 35 cancer types. The p value was determined using Cox regression analysis. The forest plots plotted the HR and 95% CI, and the p values of the survival curves were calculated and visualised using ‘survminer’ and ‘ggplot2’ (version 3.3.3) package.
Genetic alteration analysis of GPER1
Genetic Alteration Analysis of GPER1 was performed using cBioPortal (https://www.cbioportal.org/) [43]. The ‘OncoPrint’ module was used to explore genetic alterations of GPER1. The somatic mutation frequency and genomic information of GPER1 mutations in pan-cancer were explored using the ‘cancer types summary and mutations’ module. The prognostic value of GPER1 for pan-cancer was investigated using the ‘Comparison/Survival’ module.
DEGs analysis between high and low GPER1 expression
According to the expression value of GPER1, patients were divided into high and low GPER1 expression groups, and the DEGs of the two groups were analyzed. These DEGs will be utilized for subsequent analyses involving functional enrichment and gene set enrichment.
During the detection of DEGs, it is necessary to perform differential statistical tests individually on thousands of genes within a single cancer type. This process involves multiple comparisons, which can potentially lead to false positive results, necessitating the implementation of a multiple hypothesis testing correction. We utilize the Benjamini-Hochberg (BH) method for False Discovery Rate (FDR) correction [44]. FDR is a widely employed technique for correcting multiple hypothesis testing, designed to control the proportion of erroneous rejections of null hypotheses. Compared to the conventional Bonferroni correction, the FDR method demonstrates increased applicability, particularly when facing a substantial number of hypothesis tests.
‘DESeq2’ analysis was performed in R to identify DEGs between pan-cancer patients related to GPER1 expression using unpaired Student’s t-test, with the thresholds set at an adjusted P < 0.01 and absolute log-fold change >1. Identified genes were analysed and presented as volcano plots. The top 30 up- and downregulated genes were presented as heat maps. The correlation between GPER1 and the top 30 up- and downregulated genes was assessed using the Spearman non-parametric correlation test. This method evaluates the presence of correlation between two sets by analyzing their rank orders. Each square represents the expression value of other molecules after undergoing z-score transformation among various samples, with the color intensity reflecting the absolute magnitude of the values. Z-score transformation is a commonly employed data conversion method in generating heatmap visualizations, utilized to mitigate expression value discrepancies across diverse molecules within a dataset. This approach involves subtracting the mean value of each molecule’s expression in individual samples from its global mean across all samples, followed by division by the standard deviation. Consequently, the data is endowed with similar scales and distributions across different molecules. This aids in diminishing the impact of extreme expression values on heatmap visualization while retaining the depiction of molecular differences among samples. Visualisation of all data was achieved using the ‘ggplot2’ package in R.
PPI network analyses of GPER1
To collect and integrate potential protein interactions with GPER1 in cancer patients whose OS was significantly associated with GPER1 expression, the top 30 up- and downregulated DEGs for individual types of cancers were used to search the STRING database (https://string-db.org/) [45] and conducted PPI network analysis. Through this apo reach, the PPI network was strongly associated with specific cancers. A confidence score >0.7 was set as the significance threshold.
Gene set enrichment analysis
GSEA was performed via the ‘clusterProfiler’ package to determine the biological pathway, GO and immunologic signature differences between the high and low GPER1 groups. Pathways with a false discovery rate <0.25 and adjusted p value <0.05 were considered to have remarkably changed. Gene set permutation was performed 1,000 times for each analysis. The top five entries of the enrichment analysis were presented as a mountain map. DEGs not enriched to the relevant entry were not presented. GSEA results were presented using the ‘ggplot2’ package in R.
Promoter methylation level of GPER1 in cancers
Heatmaps of the DNA methylation of GPER1 in cancers were obtained from the MethSurv database (https://biit.cs.ut.ee/methsurv/) [46]. Promoter methylation level of GPER1 in cancers whose OS was significantly associated with GPER1 expression was explored. GPER1 methylation levels in cancers and corresponding adjacent tissues were determined from TCGA and presented via the UALCAN database [47] (http://ualcan.path.uab.edu/analysis.html). Student’s t-test was used to determine whether differences were significant. A p value of <0.05 indicated statistical significance.
Pan-cancer immunogenomic analyses of GPER1
Pan-cancer immunogenomic analyses of GPER1 was performed via the TISIDB online tool (http://cis.hku.hk/TISIDB/index.php) [48]. Correlations between expression and methylation level of GPER1 and immune components, such as TILs, immunoinhibitors, immunostimulators, MHC molecules, chemokines and chemokine receptors in pan-cancer, were presented as heatmaps. A p value <0.05 indicated statistical significance.
Data availability
The datasets for this study can be found in the HPA(https://www.proteinatlas.org/), TCGA Research Network (https://www.cancer.gov/tcga), GTEx (http://commonfund.nih.gov/GTEx/), UCSC Xena (http://xena.ucsc.edu/), MethSurv (https://biit.cs.ut.ee/methsurv/), UALCAN (http://ualcan.path.uab.edu/), cBioPortal (http://www.cbioportal.org/), STRING (https://string-db.org/) and TISIDB (http://cis.hku.hk/TISIDB/) databases.
Supplementary Materials
Author Contributions
YC F and WW: writing the article, analysis and interpretation, data mining. HW Z and XF L: conception and design, critical revision of the article. All authors contributed to the article and approved the submitted version.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
HW Z is supported by the Science and Technology Program of Sichuan Province (Grant No. 2020YFS0417).
References
- 1. Bray F, Laversanne M, Weiderpass E, Soerjomataram I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer. 2021; 127:3029–30. https://doi.org/10.1002/cncr.33587 [PubMed]
- 2. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021; 71:209–49. https://doi.org/10.3322/caac.21660 [PubMed]
- 3. Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer Statistics, 2021. CA Cancer J Clin. 2021; 71:7–33. https://doi.org/10.3322/caac.21654 [PubMed]
- 4. Feng RM, Zong YN, Cao SM, Xu RH. Current cancer situation in China: good or bad news from the 2018 Global Cancer Statistics? Cancer Commun (Lond). 2019; 39:22. https://doi.org/10.1186/s40880-019-0368-6 [PubMed]
- 5. Biomarkers Definitions Working Group. Biomarkers and surrogate endpoints: preferred definitions and conceptual framework. Clin Pharmacol Ther. 2001; 69:89–95. https://doi.org/10.1067/mcp.2001.113989 [PubMed]
- 6. Elmore LW, Greer SF, Daniels EC, Saxe CC, Melner MH, Krawiec GM, Cance WG, Phelps WC. Blueprint for cancer research: Critical gaps and opportunities. CA Cancer J Clin. 2021; 71:107–39. https://doi.org/10.3322/caac.21652 [PubMed]
- 7. Wender RC, Brawley OW, Fedewa SA, Gansler T, Smith RA. A blueprint for cancer screening and early detection: Advancing screening’s contribution to cancer control. CA Cancer J Clin. 2019; 69:50–79. https://doi.org/10.3322/caac.21550 [PubMed]
- 8. Trenti A, Tedesco S, Boscaro C, Trevisi L, Bolego C, Cignarella A. Estrogen, Angiogenesis, Immunity and Cell Metabolism: Solving the Puzzle. Int J Mol Sci. 2018; 19:859. https://doi.org/10.3390/ijms19030859 [PubMed]
- 9. Paterni I, Granchi C, Katzenellenbogen JA, Minutolo F. Estrogen receptors alpha (ERα) and beta (ERβ): subtype-selective ligands and clinical potential. Steroids. 2014; 90:13–29. https://doi.org/10.1016/j.steroids.2014.06.012 [PubMed]
- 10. Prossnitz ER, Barton M. Estrogen biology: new insights into GPER function and clinical opportunities. Mol Cell Endocrinol. 2014; 389:71–83. https://doi.org/10.1016/j.mce.2014.02.002 [PubMed]
- 11. Somasundaram A, Rothenberger NJ, Stabile LP. The Impact of Estrogen in the Tumor Microenvironment. Adv Exp Med Biol. 2020; 1277:33–52. https://doi.org/10.1007/978-3-030-50224-9_2 [PubMed]
- 12. Hasbi A, O’Dowd BF, George SR. A G protein-coupled receptor for estrogen: the end of the search? Mol Interv. 2005; 5:158–61. https://doi.org/10.1124/mi.5.3.5 [PubMed]
- 13. Xu S, Yu S, Dong D, Lee LT. G Protein-Coupled Estrogen Receptor: A Potential Therapeutic Target in Cancer. Front Endocrinol (Lausanne). 2019; 10:725. https://doi.org/10.3389/fendo.2019.00725 [PubMed]
- 14. Feldman RD, Limbird LE. GPER (GPR30): A Nongenomic Receptor (GPCR) for Steroid Hormones with Implications for Cardiovascular Disease and Cancer. Annu Rev Pharmacol Toxicol. 2017; 57:567–84 https://doi.org/10.1146/annurev-pharmtox-010716-104651 [PubMed]
- 15. Pepermans RA, Sharma G, Prossnitz ER. G Protein-Coupled Estrogen Receptor in Cancer and Stromal Cells: Functions and Novel Therapeutic Perspectives. Cells. 2021; 10:672. https://doi.org/10.3390/cells10030672 [PubMed]
- 16. Shen YT, Huang X, Zhang G, Jiang B, Li CJ, Wu ZS. Pan-Cancer Prognostic Role and Targeting Potential of the Estrogen-Progesterone Axis. Front Oncol. 2021; 11:636365. https://doi.org/10.3389/fonc.2021.636365 [PubMed]
- 17. Hernández-Silva CD, Villegas-Pineda JC, Pereira-Suárez AL. Expression and Role of the G Protein-Coupled Estrogen Receptor (GPR30/GPER) in the Development and Immune Response in Female Reproductive Cancers. Front Endocrinol (Lausanne). 2020; 11:544. https://doi.org/10.3389/fendo.2020.00544 [PubMed]
- 18. Liang S, Chen Z, Jiang G, Zhou Y, Liu Q, Su Q, Wei W, Du J, Wang H. Activation of GPER suppresses migration and angiogenesis of triple negative breast cancer via inhibition of NF-κB/IL-6 signals. Cancer Lett. 2017; 386:12–23. https://doi.org/10.1016/j.canlet.2016.11.003 [PubMed]
- 19. Sathya S, Sudhagar S, Lakshmi BS. Estrogen suppresses breast cancer proliferation through GPER / p38 MAPK axis during hypoxia. Mol Cell Endocrinol. 2015; 417:200–10. https://doi.org/10.1016/j.mce.2015.09.032 [PubMed]
- 20. Wei W, Chen ZJ, Zhang KS, Yang XL, Wu YM, Chen XH, Huang HB, Liu HL, Cai SH, Du J, Wang HS. The activation of G protein-coupled receptor 30 (GPR30) inhibits proliferation of estrogen receptor-negative breast cancer cells in vitro and in vivo. Cell Death Dis. 2014; 5:e1428. https://doi.org/10.1038/cddis.2014.398 [PubMed]
- 21. Skrzypczak M, Schüler S, Lattrich C, Ignatov A, Ortmann O, Treeck O. G protein-coupled estrogen receptor (GPER) expression in endometrial adenocarcinoma and effect of agonist G-1 on growth of endometrial adenocarcinoma cell lines. Steroids. 2013; 78:1087–91. https://doi.org/10.1016/j.steroids.2013.07.007 [PubMed]
- 22. Xu E, Xia X, Jiang C, Li Z, Yang Z, Zheng C, Wang X, Du S, Miao J, Wang F, Wang Y, Lu X, Guan W. GPER1 Silencing Suppresses the Proliferation, Migration, and Invasion of Gastric Cancer Cells by Inhibiting PI3K/AKT-Mediated EMT. Front Cell Dev Biol. 2020; 8:591239. https://doi.org/10.3389/fcell.2020.591239 [PubMed]
- 23. Liu C, Liao Y, Fan S, Tang H, Jiang Z, Zhou B, Xiong J, Zhou S, Zou M, Wang J. G protein-coupled estrogen receptor (GPER) mediates NSCLC progression induced by 17β-estradiol (E2) and selective agonist G1. Med Oncol. 2015; 32:104. https://doi.org/10.1007/s12032-015-0558-2 [PubMed]
- 24. Kurt AH, Çelik A, Kelleci BM. Oxidative/antioxidative enzyme-mediated antiproliferative and proapoptotic effects of the GPER1 agonist G-1 on lung cancer cells. Oncol Lett. 2015; 10:3177–82. https://doi.org/10.3892/ol.2015.3711 [PubMed]
- 25. Zhu G, Huang Y, Wu C, Wei D, Shi Y. Activation of G-Protein-Coupled Estrogen Receptor Inhibits the Migration of Human Nonsmall Cell Lung Cancer Cells via IKK-β/NF-κB Signals. DNA Cell Biol. 2016; 35:434–42. https://doi.org/10.1089/dna.2016.3235 [PubMed]
- 26. Wang Z, Chen X, Zhao Y, Jin Y, Zheng J. G-protein-coupled estrogen receptor suppresses the migration of osteosarcoma cells via post-translational regulation of Snail. J Cancer Res Clin Oncol. 2019; 145:87–96. https://doi.org/10.1007/s00432-018-2768-4 [PubMed]
- 27. Natale CA, Li J, Pitarresi JR, Norgard RJ, Dentchev T, Capell BC, Seykora JT, Stanger BZ, Ridky TW. Pharmacologic Activation of the G Protein-Coupled Estrogen Receptor Inhibits Pancreatic Ductal Adenocarcinoma. Cell Mol Gastroenterol Hepatol. 2020; 10:868–80.e1. https://doi.org/10.1016/j.jcmgh.2020.04.016 [PubMed]
- 28. Cortes E, Sarper M, Robinson B, Lachowski D, Chronopoulos A, Thorpe SD, Lee DA, Del Río Hernández AE. GPER is a mechanoregulator of pancreatic stellate cells and the tumor microenvironment. EMBO Rep. 2019; 20:e46556. https://doi.org/10.15252/embr.201846556 [PubMed]
- 29. Saghafinia S, Mina M, Riggi N, Hanahan D, Ciriello G. Pan-Cancer Landscape of Aberrant DNA Methylation across Human Tumors. Cell Rep. 2018; 25:1066–80.e8. https://doi.org/10.1016/j.celrep.2018.09.082 [PubMed]
- 30. Chevalier N, Paul-Bellon R, Camparo P, Michiels JF, Chevallier D, Fénichel P. Genetic variants of GPER/GPR30, a novel estrogen-related G protein receptor, are associated with human seminoma. Int J Mol Sci. 2014; 15:1574–89. https://doi.org/10.3390/ijms15011574 [PubMed]
- 31. Pupo M, Bodmer A, Berto M, Maggiolini M, Dietrich PY, Picard D. A genetic polymorphism repurposes the G-protein coupled and membrane-associated estrogen receptor GPER to a transcription factor-like molecule promoting paracrine signaling between stroma and breast carcinoma cells. Oncotarget. 2017; 8:46728–44. https://doi.org/10.18632/oncotarget.18156 [PubMed]
- 32. Rouhimoghadam M, Lu AS, Salem AK, Filardo EJ. Therapeutic Perspectives on the Modulation of G-Protein Coupled Estrogen Receptor, GPER, Function. Front Endocrinol (Lausanne). 2020; 11:591217. https://doi.org/10.3389/fendo.2020.591217 [PubMed]
- 33. Wegner MS, Gruber L, Schömel N, Trautmann S, Brachtendorf S, Fuhrmann D, Schreiber Y, Olesch C, Brüne B, Geisslinger G, Grösch S. GPER1 influences cellular homeostasis and cytostatic drug resistance via influencing long chain ceramide synthesis in breast cancer cells. Int J Biochem Cell Biol. 2019; 112:95–106. https://doi.org/10.1016/j.biocel.2019.05.002 [PubMed]
- 34. Weißenborn C, Ignatov T, Poehlmann A, Wege AK, Costa SD, Zenclussen AC, Ignatov A. GPER functions as a tumor suppressor in MCF-7 and SK-BR-3 breast cancer cells. J Cancer Res Clin Oncol. 2014; 140:663–71. https://doi.org/10.1007/s00432-014-1598-2 [PubMed]
- 35. Weißenborn C, Ignatov T, Ochel HJ, Costa SD, Zenclussen AC, Ignatova Z, Ignatov A. GPER functions as a tumor suppressor in triple-negative breast cancer cells. J Cancer Res Clin Oncol. 2014; 140:713–23. https://doi.org/10.1007/s00432-014-1620-8 [PubMed]
- 36. Zhang D, Wang J, Chen H, Yan S. Cytoplasmic G Protein-Coupled Estrogen Receptor 1 as a Prognostic Indicator of Breast Cancer: A Meta-Analysis. Technol Cancer Res Treat. 2022; 21:15330338221131664. https://doi.org/10.1177/15330338221131664 [PubMed]
- 37. Ruckriegl S, Loris J, Wert K, Bauerschmitz G, Gallwas J, Gründker C. Knockdown of G Protein-coupled Estrogen Receptor 1 (GPER1) Enhances Tumor-supportive Properties in Cervical Carcinoma Cells. Cancer Genomics Proteomics. 2023; 20:281–97. https://doi.org/10.21873/cgp.20381 [PubMed]
- 38. Vivian J, Rao AA, Nothaft FA, Ketchum C, Armstrong J, Novak A, Pfeil J, Narkizian J, Deran AD, Musselman-Brown A, Schmidt H, Amstutz P, Craft B, et al. Toil enables reproducible, open source, big biomedical data analyses. Nat Biotechnol. 2017; 35:314–6. https://doi.org/10.1038/nbt.3772 [PubMed]
- 39. Youden WJ. Index for rating diagnostic tests. Cancer. 1950; 3:32–5. https://doi.org/10.1002/1097-0142(1950)3:1<32::aid-cncr2820030106>3.0.co;2-3 [PubMed]
- 40. Nahm FS. Receiver operating characteristic curve: overview and practical use for clinicians. Korean J Anesthesiol. 2022; 75:25–36. https://doi.org/10.4097/kja.21209 [PubMed]
- 41. Perkins NJ, Schisterman EF. The inconsistency of “optimal” cutpoints obtained using two criteria based on the receiver operating characteristic curve. Am J Epidemiol. 2006; 163:670–5. https://doi.org/10.1093/aje/kwj063 [PubMed]
- 42. Greiner M, Pfeiffer D, Smith RD. Principles and practical application of the receiver-operating characteristic analysis for diagnostic tests. Prev Vet Med. 2000; 45:23–41. https://doi.org/10.1016/s0167-5877(00)00115-x [PubMed]
- 43. Gao J, Aksoy BA, Dogrusoz U, Dresdner G, Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, Cerami E, Sander C, Schultz N. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 2013; 6:pl1. https://doi.org/10.1126/scisignal.2004088 [PubMed]
- 44. Glickman ME, Rao SR, Schultz MR. False discovery rate control is a recommended alternative to Bonferroni-type adjustments in health studies. J Clin Epidemiol. 2014; 67:850–7. https://doi.org/10.1016/j.jclinepi.2014.03.012 [PubMed]
- 45. Szklarczyk D, Gable AL, Nastou KC, Lyon D, Kirsch R, Pyysalo S, Doncheva NT, Legeay M, Fang T, Bork P, Jensen LJ, von Mering C. The STRING database in 2021: customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021; 49:D605–12. https://doi.org/10.1093/nar/gkaa1074 [PubMed]
- 46. Modhukur V, Iljasenko T, Metsalu T, Lokk K, Laisk-Podar T, Vilo J. MethSurv: a web tool to perform multivariable survival analysis using DNA methylation data. Epigenomics. 2018; 10:277–88. https://doi.org/10.2217/epi-2017-0118 [PubMed]
- 47. Chandrashekar DS, Bashel B, Balasubramanya SA, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BV, Varambally S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia. 2017; 19:649–58. https://doi.org/10.1016/j.neo.2017.05.002 [PubMed]
- 48. Ru B, Wong CN, Tong Y, Zhong JY, Zhong SS, Wu WC, Chu KC, Wong CY, Lau CY, Chen I, Chan NW, Zhang J. TISIDB: an integrated repository portal for tumor-immune system interactions. Bioinformatics. 2019; 35:4200–2. https://doi.org/10.1093/bioinformatics/btz210 [PubMed]