



Introduction
Esophageal cancer (EC) ranks eighth globally in terms of incidence and is the sixth leading cause of cancer-related deaths [1]. According to statistics, there were over 600,000 new cases and 540,000 deaths worldwide in 2020. Esophageal squamous cell carcinoma (ESCC) is one of the primary histological subtypes of EC, accounting for 85% of EC cases in the world [2]. The high-risk EC regions of Asia (including East, Central, and South Asia) and Africa (including East and Southern Africa) accounted for about 80% of EC cases, primarily ESCC [1, 3, 4]. The majority of ESCC patients received their diagnosis when the disease was either in the middle or late stages and there was no longer any hope for recovery. In certain countries, the 5-year survival rate is as low as 5%, with less than 20% overall [5, 6]. It has been shown that cancer stem cells (CSCs), a subpopulation of cells with self-renewal, multidirectional differentiation potential, and tumorigenic ability, are the underlying cause of tumorigenesis, progression, recurrence, metastasis, and drug resistance [7]. It is now well established that the poor prognosis of ESCC patients is closely associated with CSCs [8]. Clinical studies have eliminated CSCs by targeting Wnt, NOTCH, Hedgehog, and Hippo signaling pathways [9]. However, there is less consensus on biomarkers for CSCs in ESCC. Therefore, investigating the pathogenic mechanisms of ESCC and looking for more trustworthy targets is imperative considering these situations.
Circular RNA (circRNA) was originally thought to be a byproduct of erroneous splicing, formed by reverse splicing of pre-mRNA into a single-stranded covalently closed RNA molecule [10], and it was resistant to RNA nucleases due to its closed loop structure, which increased the stability and reduced the susceptibility to degradation of their expression [11]. Researchers have recently started to focus on the role of circRNA in various human diseases, including cancer [12–14]. In cancers, the crucial roles of circRNA have been confirmed in gastric cancer [15], colorectal cancer [16], lung cancer [17], breast cancer [18], etc. In certain cases, circRNA functions as a transcription factor, miRNA sponge, protein scaffold, or even a protein translation template [19]. Research on ESCC has focused on the dysregulation of circRNA as a diagnostic biomarker [20] and its role in disease progression [21–23], while the circRNA-miRNA-mRNA regulatory network awaits further exploration. Shen et al. [24] constructed a circRNA-miRNA-mRNA interaction network based on ESCC microarray profiles and public databases, and then identified hub genes, validating the significant function of the circRNA-miRNA-mRNA regulatory network in ESCC. Recently, the studies by Wang et al. [25], Zhou et al. [26], and Song et al. [27] enriched the circRNA-miRNA-mRNA regulatory network in ESCC to some extent. Regrettably, a deeper comprehension of the regulatory mechanisms had not been advanced by these studies. Previous research revealed that ESCC patients with high expression of circRNA6448-14 have poor prognosis, poor differentiation, and high pTNM stage. Cell progression is accelerated by circRNA6448-14. Simultaneously, circRNA6448-14 can act as a sponge for miR-455-3p [28]. This study aimed to define the circRNA-miRNA-mRNA regulatory network and to complete the precise mechanism of this regulatory axis in ESCC progression by investigating the downstream target genes of miR-455-3p.
In this study, we explored the effects of circRNA6448-14 on glycolysis and cell stemness in ESCC cells and, based on this, further explored its downstream molecular regulatory mechanisms. In conclusion, our research added to the understanding of the circRNA-miRNA-mRNA regulatory network in ESCC and offered a novel and solid clue for its treatment.
Materials and Methods
Bioinformatics analysis
We obtained the expression of circRNA6448-14 and miR-455-3p in ESCC tissues from previous studies and verified that circRNA6448-14 could sponge miR-455-3p [28]. After using miRwalk to find the downstream target genes of miR-455-3p and performing a literature review, OTU Domain-Containing Ubiquitin Aldehyde-Binding Protein 2 (OTUB2) was ultimately determined to be the target. OTUB2 differential expression in ESCC was analyzed based on The Cancer Genome Atlas (TCGA)-ESCC-mRNA expression data (normal: 3, tumor: 96).
Cell cultivation
All cells used in this study, human embryonic kidney cells (293T), normal human esophageal epithelial cells (HEEC), and ESCC cell lines (TE1, Eca109, KYSE150) were purchased from BeNa Culture Collection (BNCC, China) and cultured according to the recommended protocol. 293T cells were cultivated in Dulbecco’s modified Eagle’s medium-high glucose (DMEM-H), HEEC cells were cultivated in Eagle’s minimal essential medium (EMEM), TE1 and Eca109 cells were cultivated in RPMI-1640 medium, KYSE-150 cells were cultured in KYSE-150 cell special medium (45% RPMI-1640 + 45% F-12 + 10% fetal bovine serum (FBS)), all media contained 1% penicillin-streptomycin mixture and 10% FBS, and the DMEM-H medium for 293T cells also contained 2 mM L-glutamine [29]. All cells were cultured in the incubator at 37°C with 5% CO2. The media were purchased from BNCC (China), and the reagents were Gibco products from Thermo Fisher Scientific (USA).
Cell transfection
siRNA targeting circRNA6448-14, miR-455-3p inhibitor, miR-455-3p mimic, OTUB2 overexpression vector, and corresponding controls were synthesized by RiboBio (China). According to instructions, the above vectors (1 μg/mL) were transfected into ESCC cells or 293T cells utilizing Lipofectamine™ 2000 (Invitrogen, USA), and the transfection efficiency was detected after 48 hours [30].
qRT-PCR
Total RNA was isolated from cells using RNAsimple Total RNA Kit (TIANGEN, China), and reverse transcription was performed to obtain cDNA using Quantitect Reverse Transcription Kit (Qiagen, Germany). Next, Power Green qPCR Mix (Takara Bio, Japan) used the Applied Biosystems™ 7500 real-time PCR system to conduct the qRT-PCR assay. Finally, relative expression levels were calculated using 2−ΔΔCt with GAPDH or U6 as controls [31]. The primer sequences are in the Table 1.
Table 1. qRT-PCR primer sequences.
| Gene | Sequence |
| hsa_circ RNA6448-14 | Forward: 5′-CCAATGGGGACTGTCATGGA-3′ |
| Reverse: 5′-TCATGCCGTGTTTCAGCTCA-3′ | |
| miR-455-3p | Forward: 5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGTGTAT-3′ |
| Reverse: 5′-ACACTCCAGCTGGGGCAGTCCATGGGCAT-3′ | |
| U6 | Forward: 5′-AACGCTTCACGAATTTGCGT-3′ |
| Reverse: 5′-CTCGCTTCGGCAGCACA-3′ | |
| GAPDH | Forward: 5′-GAAGGTGAAGGTCGGAGTC-3′ |
| Reverse: 5′-GAAGATGGTGATGGGATTTC-3′ |
Western blot (WB)
The collected cells were lysed utilizing radioimmunoprecipitation assay (RIPA) buffer (Beyotime, China) for 30 minutes to obtain the total protein. The bicinchoninic acid (BCA) assay kit (Beyotime, China) was utilized to quantify the protein concentration. The membrane was transferred after SDS-PAGE electrophoresis. Then, the membrane was blocked with 5% BCA for 2 hours and incubated at 4°C overnight with primary rabbit antibodies (Abcam, UK): anti-Nanog (ab109250, 1:1000), anti-OCT4 (ab181557, 1:1000), anti-SOX2 (ab92494, 1:1000), anti-HK2 (ab209847, 1:1000), and anti-LDH4 (ab52488, 1:5000). The next day, after washing with TBST (5 min × 3), the membrane was incubated for 2 hours in the secondary antibody, goat anti-rabbit Immunoglobulin G (IgG) H&L (HRP) (ab6721, 1:2000, Abcam, UK). Finally, Western Blotting Luminol Reagent (Santa Cruz, USA) was used to achieve visualization in the ChemiScope6000 system [32].
MTT assay
The transfected cells were seeded into 96-well plates at 2000 cells per well and cultured until the cells adhered. After 0, 24, 48, and 72 hours of cultivation, each well was added with 20 μl MTT solution (5 mg/mL). After 4 hours of incubation, 150 μl of DMSO was added and shaken for 10 minutes. A microplate reader (MTT and DMSO purchased from Solarbio, China) was used to measure the absorbance at 570 nm wavelength [33].
Stem cell sphere formation assay
6-well ultra-low attachment plates (Corning, USA) were seeded with 4 × 104 cells per well. The DMEM/F-12 medium was used to cultivate the cells, and it was supplemented with β-FGF (10 ng/mL), B27 (20 ng/mL), IGF (20 ng/mL), and human EGF (20 ng/mL). Cells were counted and photographed under an inverted microscope after 14 days of cultivation at 37°C with 5% CO2 [34]. Reagents and culture media were from Invitrogen (USA).
Extracellular acidification rate (ECAR) and oxygen consumption rate (OCR) detection
The Seahorse XFe96 extracellular flux analyzer (Seahorse Bioscience, USA) was used to measure the ECAR and OCR. After cells were harvested and counted, 1 × 104 cells/well were seeded into a SeahorseXF96 cell culture microplate and cultured for 10 hours. Afterwards, the Seahorse XF glycolysis Stress Test Kit and Seahorse XF Cell Mito Stress Test Kit (Agilent, USA) were used to measure ECAR and OCR, respectively. Glucose, and oligomycin or 2-DG should be added at specified times in the determination of ECAR. Oligomycin, and FCCP or antimycin A and Rotenone should be added at specified times in the determination of OCR [35].
Lactate production, glucose consumption, and ATP detection
The Lactate Assay Kit, Glucose Uptake Assay Kit, and Luminescent ATP Detection Assay Kit (Solarbio, China) were used to detect lactate production, glucose consumption, and ATP generation, respectively.
Dual-luciferase reporter assay
First, we constructed pmirGLO-circRNA6448-14-WT (wild-type), pmirGLO-circRNA6448-14-MUT (mutant-type), pmirGLO-OTUB2-MUT, and pmirGLO-OTUB2-MUT plasmids. The 3′UTR regions containing the binding sites of miR-149-3p in circRNA6448-14 or OTUB2 fragments, as well as the 3′UTR regions of circRNA6448-14 or OTUB2 fragments with mutated binding sites, were inserted into the pmirGLO vector. Plasmid and miR-455-3p mimic or NC mimic were co-transfected into 293T cells. As directed by the manufacturer, the Dual-Luciferase Reporter Assay System (Promega, USA) was utilized to measure the luciferase activity.
Statistical analysis
Data were displayed as mean ± standard deviation of a minimum of three independent experiments. The Student’s t-test was used to compare the differences between the two groups. Groups with continuous variables were compared using analysis of variance. P < 0.05 implied a significant difference.
Data availability statement
The data and materials in the current study are available from the corresponding author on reasonable request.
Results
circRNA6448-14 is highly expressed in ESCC
In previous study, we investigated the expression profiles of circRNAs in ESCC and adjacent normal tissues by microarray analysis and selected upregulated circRNA, circRNA6448-14, based on the following criteria: (1) fold change, P-value; (2) the number of miRNA binding sites; (3) presence of target miRNAs associated with ESCC. It has been established that ESCC tissues express circRNA6448-14 at a higher level than normal tissues [28]. This finding led us to investigate the expression level of circRNA6448-14 in ESCC cell lines (TE1, Eca109, KYSE150), and the results suggested that these cell lines also had high expression levels of circRNA6448-14 (Figure 1). In conclusion, circRNA6448-14 was highly expressed in ESCC.
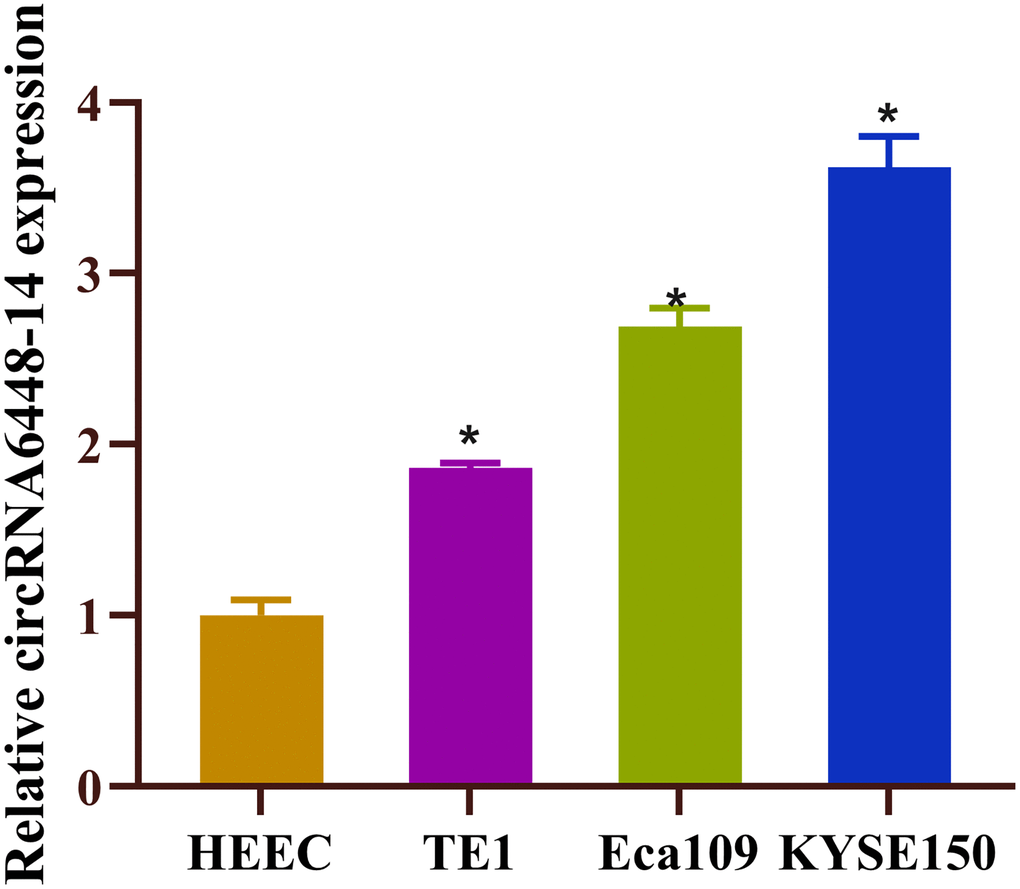
Figure 1. High expression of circRNA6448-14 in ESCC. Expression of circRNA6448-14 in normal esophageal epithelial cell HEEC and ESCC cells TE1, Eca109, KYSE150. *P < 0.05.
circRNA6448-14 stimulates ESCC glycolysis and stemness
It was determined that circRNA6448-14 enhanced the capacity of ESCC cells to migrate and invade and circRNA6448-14 could be enriched in the pathways related to the regulation of tumor protein polysaccharide, according to previous studies [28]. Therefore, we speculated that circRNA6448-14 may stimulate the glycolysis and stemness of ESCC cells. We carried out several experiments to verify this hypothesis. First, si-NC and si-circRNA6448-14 cell groups were constructed, and it was proved that circRNA6448-14 expression was notably reduced in the circRNA6448-14 knockdown group (Figure 2A). MTT assay indicated the inhibitory effect of circRNA6448-14 knockdown for cell viability (Figure 2B). KYSE150 sphere formation ability was markedly suppressed by circRNA6448-14 knockdown in the stem cell sphere formation assay (Figure 2C). After determining the expression of glycolysis rate-limiting enzymes (HK2, LDHA) and stem cell surface markers (Nanog, OCT4, SOX2) in KYSE150 cells with circRNA6448-14 knockdown, it was discovered that these expressions were dramatically lower (Figure 2D, 2E). The circRNA6448-14 knockdown group demonstrated a notable increase in OCR detection and a major decrease in ECAR detection when we performed ECAR and OCR assays (Figure 2F, 2G). The results reflected that knockdown of circrna6448-14 decreased overall glycolytic flux as well as enhanced mitochondrial oxidative respiration in KYSE150 cells. Finally, we determined the effect of circRNA6448-14 knockdown on glycolytic phenotypes including lactic acid production, glucose consumption, and ATP production in KYSE150 cells. We found that knocking down circRNA6448-14 inhibited lactate production, glucose consumption, and ATP production in KYSE150 cells (Figure 2H–2J). In conclusion, circRNA6448-14 promoted ESCC cell proliferation in glycolysis and stemness.
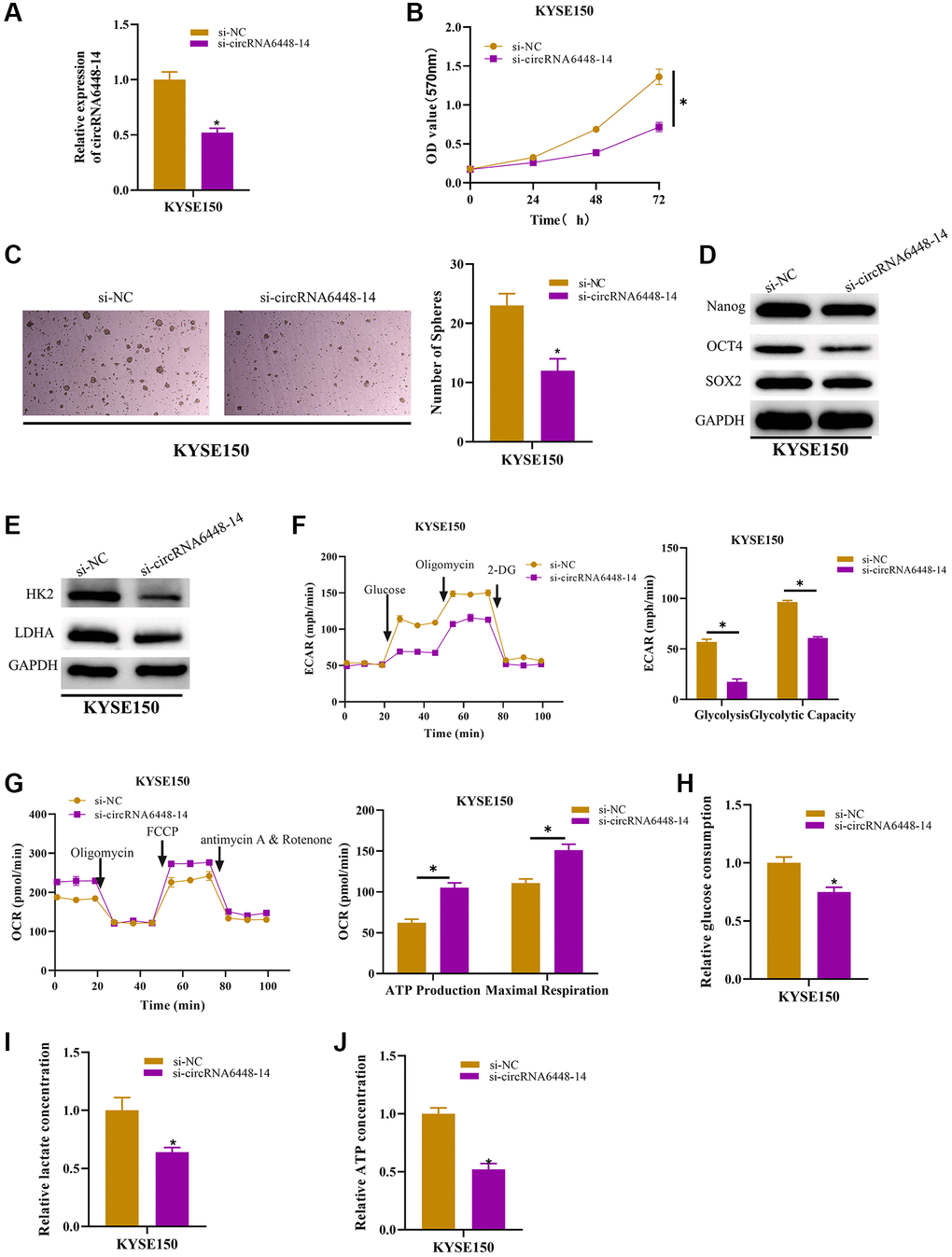
Figure 2. circRNA6448-14 increases stemness and glycolysis in ESCC cells. (A) qRT-PCR detection of circRNA6448-14 expression in circRNA6448-14 knockdown and control cells. (B) MTT assay detection of cell viability in circRNA6448-14 knockdown and control cells. (C) Stem cell sphere formation experiment in circRNA6448-14 knockdown and control cells. (D, E) WB detection of stem cell surface marker proteins Nanog, OCT4, SOX2 expression and glycolysis rate-limiting enzymes HK2, LDHA expression in circRNA6448-14 knockdown and control cells. (F) The ECAR of circRNA6448-14 knockdown and control cells. (G) The OCR of circRNA6448-14 knockdown and control cells. (H–J) The kits detected glucose consumption, lactate production, and ATP generation levels in cells. *P < 0.05.
Functional relationship of circRNA6448-14/miR-455-3p/OTUB2 axis
Recent research suggested that many circRNAs may act as miRNA sponges [36]. The previous study had constructed the circRNA6448-14-miRNA network and demonstrated that circRNA6448-14 could sponge miR-455-3p [28]. miR-455-3p expression was identified and shown to be downregulated in ESCC cell lines (Figure 3A). Furthermore, we performed RIP and dual luciferase assay to validate the binding relationship between circRNA6448-14 and miR-455-3p. According to the RIP results, the group treated with miR-455-3p mimic exhibited significantly higher levels of co-precipitation of AGO2 and circRNA6448-14 in comparison to the IgG group (Figure 3B). According to the dual luciferase assay, transfection of mimic-miR-455-3p significantly reduced the luciferase activity of the WT-circRNA6448-14 group but had no obvious effect on the MUT-circRNA6448-14 group (Figure 3C). Moreover, we used miRwalk to forecast the downstream mRNA regulated by miR-455-3p to enhance the circRNA-miRNA-mRNA network, and ultimately, we chose OTUB2 as the target (binding site shown in Figure 3D). The expression analysis of OTUB2 in TCGA-ESCC-mRNA data revealed its high expression in ESCC tissues (Figure 3E). According to RIP experiments, the group treated with miR-455-3p mimic had a fairly high level of AGO2 enrichment when co-precipitating with OTUB2 than the IgG group (Figure 3F). Meanwhile, in the dual luciferase assay, the WT-OTUB2 group treated with miR-455-3p mimic showed a great decrease in luciferase activity, whereas the MUT-OTUB2 group did not show any huge change (Figure 3G). We discovered that OTUB2 displayed an upregulated trend in ESCC cell lines (Figure 3H). Finally, to find out the relationship between circRNA6448-14/miR-455-3p/OTUB2 axis, we set up si-NC+inhibitor-NC, si-circRNA6448-14+inhibitor-NC, and si-circRNA6448-14+inhibitor-miR-455-3p groups and detected the expression of OTUB2 in each group. It was revealed that OTUB2 expression was inhibited when circRNA6448-14 was knocked down alone but was restored when adding miR-455-3p inhibitor (Figure 3I). In conclusion, circRNA6448-14 promoted the expression of OTUB2 by sponge adsorption of miR-455-3p.
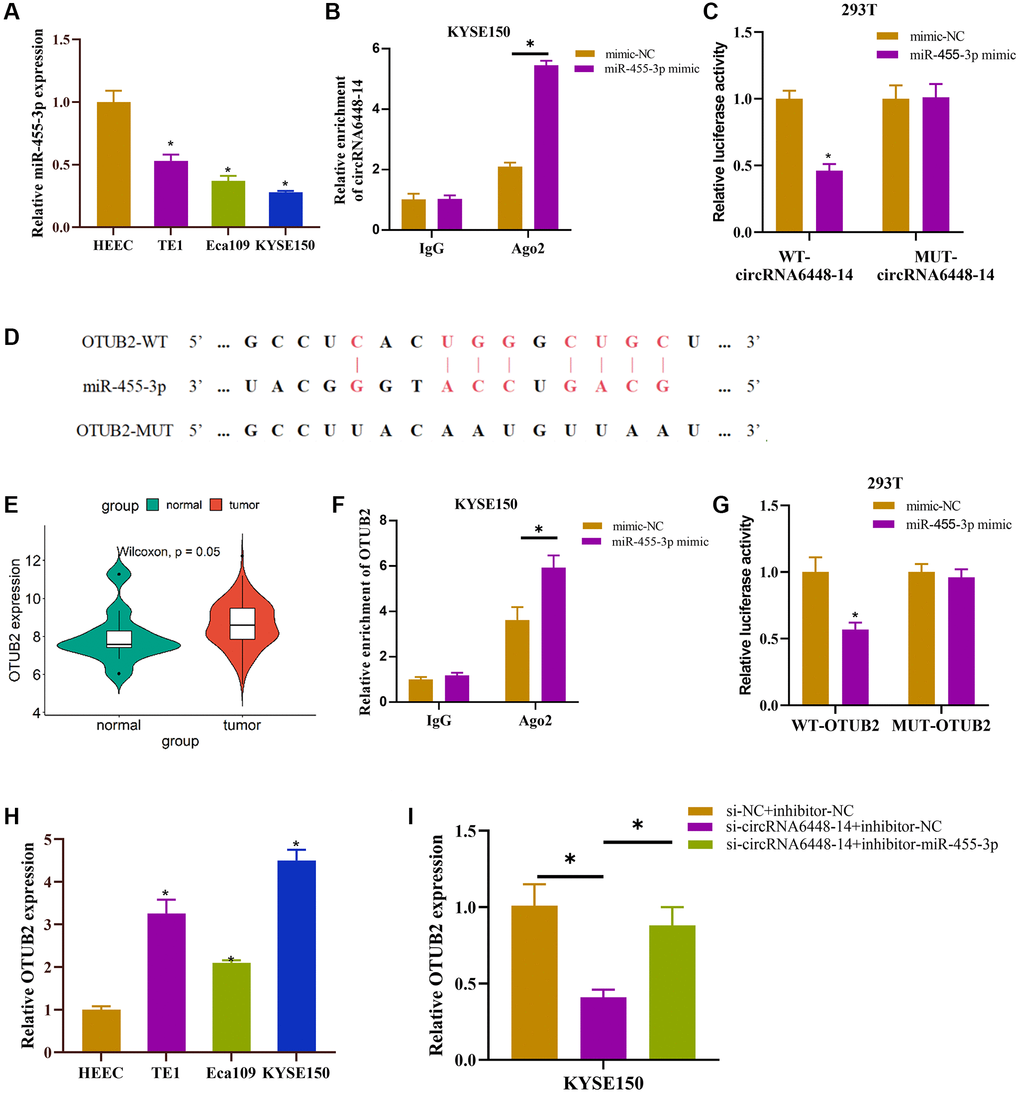
Figure 3. Functional relationship of circRNA6448-14/miR-455-3p/OTUB2 axis. (A) qRT-PCR detection of miR-455-3p expression in normal esophageal epithelial cell HEEC and ESCC cells TE1, Eca109, KYSE150. (B, C) RIP assay and dual luciferase assay were used to verify the binding relationship between circRNA6448-14 and miR-455-3p. (D) Binding site of miR-455-3p and OTUB2. (E) Expression of OTUB2 in ESCC and normal tissues. (F, G) RIP assay and dual luciferase assay were used to verify the binding relationship between miR-455-3p and OTUB2. (H) qRT-PCR detection of OTUB2 expression in normal esophageal epithelial cells HEEC and ESCC cells TE1, Eca109, KYSE150. (I) qRT-PCR detection of OTUB2 expression in different treatment groups. *P < 0.05.
circRNA6448-14/miR-455-3p/OTUB2 axis promotes ESCC cells glycolysis and stemness
After confirming the functional relationship between circRNA6448-14/miR-455-3p/OTUB2 axis, we further investigated the effects of this regulatory axis on ESCC cell glycolysis and their stemness. First, cell groups were constructed for subsequent experiments: NC (si-NC+inhibitor-NC+oe-NC treatment), si-circRNA6448-14 (si-circRNA6448-14+inhibitor-NC+oe-NC treatment), si-circRNA6448-14+miR-455-3p inhibitor (si-circRNA6448-14+miR-455-3p inhibitor+oe-NC treatment), and si-circRNA6448-14+oe-OTUB2 (si-circRNA6448-14+inhibitor-NC+oe-OTUB2 treatment). According to the results, OTUB2 expression in KYSE150 cells was considerably reduced by circRNA6448-14 knockdown alone when compared to the other three groups (Figure 4A). The inhibitory effect of knocking down circRNA6448-14 on cell viability was eliminated by adding miR-455-3p inhibitor or overexpressing OTUB2 (Figure 4B). The stem cell sphere formation assay demonstrated that the inhibitory effect on sphere formation mediated by circRNA6448-14 knockdown could be reversed by inhibiting miR-455-3p or overexpressing OTUB2 (Figure 4C). The expression of cell surface marker proteins and glycolysis rate-limiting enzyme was significantly reduced in the circRNA6448-14 knockdown group and restored upon the addition of miR-455-3p inhibitor or overexpression of OTUB2 (Figure 4D, 4E). Furthermore, knocking down circRNA6448-14 inhibited the ECAR of KYSE150 cells, while inhibiting miR-455-3p or overexpressing OTUB2 restored the ECAR of KYSE150 cells (Figure 4F). In contrast, the knockdown of circRNA6448-14 promoted OCR in KYSE150 cells, and restoration of OCR to control levels was observed by adding miR-455-3p inhibitor or overexpressing OTUB2 (Figure 4G). Finally, the determination of glycolysis phenotype-related indicators showed that the inhibitory effects on lactate production, glucose consumption, and ATP production caused by knocking down circRNA6448-14 could be reversed by miR-455-3p inhibitor or overexpression of OTUB2 (Figure 4H–4J). In conclusion, circRNA6448-14 sponged miR-455-3p, which in turn promoted OTUB2 expression, thereby enhancing glycolysis and stemness of ESCC cells.
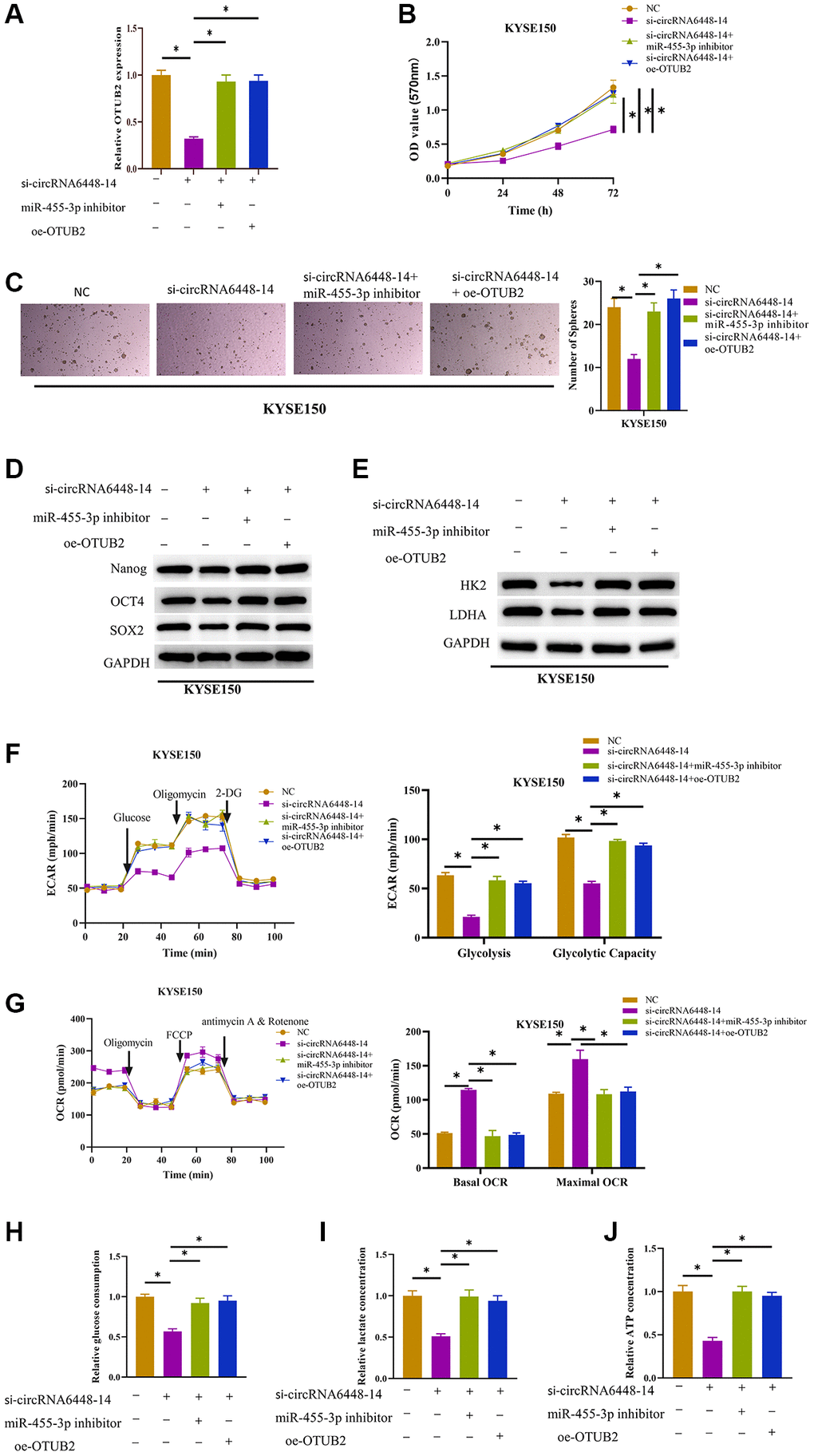
Figure 4. Stimulative effect on cell glycolysis and stemness by circRNA6448-14/miR-455-3p/OTUB2 axis. (A) qRT-PCR detection of OTUB2 expression in different treatment groups. (B) MTT detection of cell viability in different treatment groups. (C) Stem cell sphere formation assay in different treatment groups. (D, E) WB detection of stem cell surface marker proteins Nanog, OCT4, SOX2 expression and glycolysis rate-limiting enzymes HK2, LDHA expression in different treatment groups. (F, G) The ECAR and OCR of different treatment groups. (H–J) Glucose consumption, lactate production, and ATP generation levels in cells of different treatment groups. *P < 0.05.
Discussion
Cancer cells reprogrammed their metabolism to become “glycolysis-dominant” which enables them to meet their energy and macromolecule need and enhances their survival chances. It is also called the “Warburg effect”, which gives cancer cells an advantage in survival and increases the carcinogenic potential of the tumor environment [37]. However, multiple mechanisms contributed to the abnormal glycolysis in tumor cells, including abnormal gene expression [38], abnormal RNA modifications [39, 40], and abnormal post-translational modifications [41]. Recently, the function of circRNA in ESCC cell glycolysis has been evident [42]. For example, circGOT1 adsorbs miR-606 to promote the expression of GOT1, thereby inducing ESCC cell migration, proliferation, aerobic glycolysis, and cisplatin resistance [43]. By absorbing miR-497-5p, CircDUSP16 knockdown controls TKTL1 expression and prevents ESCC cell growth, invasion, and glycolysis [44]. In our study, it was found that knocking down circRNA6448-14 could prevent the glycolysis and stemness of ESCC cells. We identified circRNA6448-14 as a novel oncogene in ESCC that could accelerate the tumor growth by triggering a reprogramming of glycolysis.
In addition, cancer “stemness” is the basis for the existence of cancer, which defines the ability of cancer cells to persist and differentiate indefinitely [45], however, the regulation of cancer cell stemness may also depend on circRNA [46], and circSLC7A11 knockdown can reduce stemness of laryngeal squamous cell carcinoma cells [47]. hsa_circ_0001741 was proved to act as a miRNA sponge and prevent miR-491-5p from inhibiting ESCC cell stemness [48]. In line with earlier research, we discovered that circRNA6448-14 knockdown greatly suppressed the stemness marker Nanog, OCT4, SOX2 in ESCC. Nanog, OCT4, SOX2 are recognized as key stem cell regulators that maintain the ability of stem cells to self-renew, proliferate and differentiate, transforming cancer cells into a stem cell-like phenotype [49–51]. These results demonstrated the importance of circRNA6448-14 as a therapeutic target for ESCC.
circRNA6448-14 had been confirmed to serve as a sponge for miR-455-3 [28]. Through relevant experiments, this study further validated the binding relationship between miR-455-3p and circRNA6448-14. We used miRwalk to predict the downstream target gene (OTUB2) of miR-455-3p and obtained a complete circRNA-miRNA-mRNA regulatory network. The molecular structure of OTUB2 consists of a central five-stranded β-sheet, flanking regions of small helices at the amino terminus (α1, α2), and a large helical region (α3-α8). It is a deubiquitinating enzyme [52]. Zhang et al. [53] measured cancer metastasis by in vivo screening system and demonstrated that OTUB2 is the most indispensable gene that induces metastasis among the screened deubiquitinating enzymes. OTUB2 could make a binding relationship and deubiquitination, thereby stabilizing YAP and TAZ to activate the Hippo pathway, stimulate cancer stem cell characteristics, and maintain tumor cell proliferation and metastasis. Similarly, in non-small cell lung cancer, OTUB2 could bind to U2AF2 and deubiquitinate it, making U2AF2 more stable, promoting the Warburg effect of tumors through the AKT/mTOR signaling pathway [54]. Surprisingly, opposite results were observed in ESCC. Liu et al. [55] believed that OTUB2 is overexpressed in ESCC and can directly stabilize and interact with YAP1/TAZ through SUMO, leading to the stimulation of YAP1/TAZ expression and activation of downstream target genes such as CTGF and CYR61, thereby promoting tumor cell progression. Our results also indicated a high expression of OTUB2 in ESCC. In contrast, Chang et al. [56] discovered that OTUB2 is missing in ESCC tissues, while OTUB2 can deubiquitinate STAT1 and phosphorylate it, thereby activating CALML3 transcription and phospholipid acetylserine synthesis to exert tumor suppressive effects. The dual role of OTUB2 in cancer may be the result of its interaction with different proteins as a deubiquitinating enzyme. Deubiquitination is well known to contribute to the stability of proteins [57, 58]. OTUB2 stimulates the growth of cancer by acting on oncogenic proteins. It has a suppressive effect when it interacts with tumor suppressor proteins, which causes ESCC to react in an entirely different way. Finally, we revealed that circRNA6448-14 promoted OTUB2 expression and enhanced ESCC glycolysis and stemness by competitively binding to miR-455-3p. This is similar to the regulatory mechanism of circRNA-miRNA-mRNA network on glycolysis and stemness in hepatocellular carcinoma reported by Feng et al. [59]. Our findings make a substantial contribution to the understanding of the comparatively sparse circRNA-miRNA-mRNA regulatory network in ESCC.
In summary, this study demonstrated that by competitively binding with miR-455-3p, circRNA6448-14 enhanced OTUB2 expression, ESCC glycolysis, and stemness. Currently, the significance of the circRNA-miRNA-mRNA network in ESCC is still unclear, and our data provided a new circRNA-miRNA-mRNA regulatory network for the ESCC pathogenesis. However, it is unknown whether this regulatory axis plays a carcinogenic role in ESCC by affecting glycolysis and stemness only and has an impact on other pathways or metabolism. Also, this study lacked exploration at clinical and animal levels, but only validated at the cellular level, which was a limitation of this study. We would investigate additional potential effects of this regulatory axis in the upcoming research, which would aid in the creation of novel therapy for ESCC patients.
Author Contributions
(I) Conception and design: Yaowen Zhang, Heming Zhan. (II) Collection and assembly of data: Chenyu Wang, Shasha Cao. (III) Experiments performance: Yaowen Zhang, Heming Zhan. (IV) Data analysis and interpretation: Xinyu Cheng, Linzhi Jin. (V) Manuscript writing: Runchuan Ren, Fuyou Zhou. (VI) Final approval of manuscript: All authors.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
This study is sponsored by Project of Outstanding Young Talents in Young and Middle-aged Health Scientific and Technological Innovation of Henan Province (No. YXKC2021045), Programs for Science and Technology Development of Henan Province (No. 212102310702) and Programs for Science and Technology Development of Anyang City (No. 2021C01SF011).
References
- 1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021; 71:209–49. https://doi.org/10.3322/caac.21660 [PubMed]
- 2. Morgan E, Soerjomataram I, Rumgay H, Coleman HG, Thrift AP, Vignat J, Laversanne M, Ferlay J, Arnold M. The Global Landscape of Esophageal Squamous Cell Carcinoma and Esophageal Adenocarcinoma Incidence and Mortality in 2020 and Projections to 2040: New Estimates From GLOBOCAN 2020. Gastroenterology. 2022; 163:649–58.e2. https://doi.org/10.1053/j.gastro.2022.05.054 [PubMed]
- 3. Lagergren J, Smyth E, Cunningham D, Lagergren P. Oesophageal cancer. Lancet. 2017; 390:2383–96. https://doi.org/10.1016/S0140-6736(17)31462-9 [PubMed]
- 4. Li M, Park JY, Sheikh M, Kayamba V, Rumgay H, Jenab M, Narh CT, Abedi-Ardekani B, Morgan E, de Martel C, McCormack V, Arnold M. Population-based investigation of common and deviating patterns of gastric cancer and oesophageal cancer incidence across populations and time. Gut. 2023; 72:846–54. https://doi.org/10.1136/gutjnl-2022-328233 [PubMed]
- 5. Matsueda K, Ishihara R. Preoperative Diagnosis and Indications for Endoscopic Resection of Superficial Esophageal Squamous Cell Carcinoma. J Clin Med. 2020; 10:13. https://doi.org/10.3390/jcm10010013 [PubMed]
- 6. Yamamoto S, Kato K. JUPITER-06 establishes immune checkpoint inhibitors as essential first-line drugs for the treatment of advanced esophageal squamous cell carcinoma. Cancer Cell. 2022; 40:238–40. https://doi.org/10.1016/j.ccell.2022.02.009 [PubMed]
- 7. Barbato L, Bocchetti M, Di Biase A, Regad T. Cancer Stem Cells and Targeting Strategies. Cells. 2019; 8:926. https://doi.org/10.3390/cells8080926 [PubMed]
- 8. Taniguchi D, Saeki H, Nakashima Y, Kudou K, Nakanishi R, Kubo N, Ando K, Oki E, Oda Y, Maehara Y. CD44v9 is associated with epithelial-mesenchymal transition and poor outcomes in esophageal squamous cell carcinoma. Cancer Med. 2018; 7:6258–68. https://doi.org/10.1002/cam4.1874 [PubMed]
- 9. Clara JA, Monge C, Yang Y, Takebe N. Targeting signalling pathways and the immune microenvironment of cancer stem cells - a clinical update. Nat Rev Clin Oncol. 2020; 17:204–32. https://doi.org/10.1038/s41571-019-0293-2 [PubMed]
- 10. Chen LL. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat Rev Mol Cell Biol. 2020; 21:475–90. https://doi.org/10.1038/s41580-020-0243-y [PubMed]
- 11. Meng F, Zhang X, Wang Y, Lin J, Tang Y, Zhang G, Qiu B, Zeng X, Liu W, He X. Hsa_circ_0021727 (circ-CD44) promotes ESCC progression by targeting miR-23b-5p to activate the TAB1/NFκB pathway. Cell Death Dis. 2023; 14:9. https://doi.org/10.1038/s41419-022-05541-x [PubMed]
- 12. Wang J, Yue BL, Huang YZ, Lan XY, Liu WJ, Chen H. Exosomal RNAs: Novel Potential Biomarkers for Diseases-A Review. Int J Mol Sci. 2022; 23:2461. https://doi.org/10.3390/ijms23052461 [PubMed]
- 13. Min X, Liu DL, Xiong XD. Circular RNAs as Competing Endogenous RNAs in Cardiovascular and Cerebrovascular Diseases: Molecular Mechanisms and Clinical Implications. Front Cardiovasc Med. 2021; 8:682357. https://doi.org/10.3389/fcvm.2021.682357 [PubMed]
- 14. Patil NS, Feng B, Su Z, Castellani CA, Chakrabarti S. Circular RNA mediated gene regulation in chronic diabetic complications. Sci Rep. 2021; 11:23766. https://doi.org/10.1038/s41598-021-02980-y [PubMed]
- 15. Shan C, Zhang Y, Hao X, Gao J, Chen X, Wang K. Biogenesis, functions and clinical significance of circRNAs in gastric cancer. Mol Cancer. 2019; 18:136. https://doi.org/10.1186/s12943-019-1069-0 [PubMed]
- 16. Li X, Wang J, Zhang C, Lin C, Zhang J, Zhang W, Zhang W, Lu Y, Zheng L, Li X. Circular RNA circITGA7 inhibits colorectal cancer growth and metastasis by modulating the Ras pathway and upregulating transcription of its host gene ITGA7. J Pathol. 2018; 246:166–79. https://doi.org/10.1002/path.5125 [PubMed]
- 17. Di X, Jin X, Li R, Zhao M, Wang K. CircRNAs and lung cancer: Biomarkers and master regulators. Life Sci. 2019; 220:177–85. https://doi.org/10.1016/j.lfs.2019.01.055 [PubMed]
- 18. Zhao CH, Qu L, Zhang H, Qu R. Identification of breast cancer-related circRNAs by analysis of microarray and RNA-sequencing data: An observational study. Medicine (Baltimore). 2019; 98:e18042. https://doi.org/10.1097/MD.0000000000018042 [PubMed]
- 19. Patop IL, Wüst S, Kadener S. Past, present, and future of circRNAs. EMBO J. 2019; 38:e100836. https://doi.org/10.15252/embj.2018100836 [PubMed]
- 20. Fan L, Cao Q, Liu J, Zhang J, Li B. Circular RNA profiling and its potential for esophageal squamous cell cancer diagnosis and prognosis. Mol Cancer. 2019; 18:16. https://doi.org/10.1186/s12943-018-0936-4 [PubMed]
- 21. Gu L, Sang Y, Nan X, Zheng Y, Liu F, Meng L, Sang M, Shan B. circCYP24A1 facilitates esophageal squamous cell carcinoma progression through binding PKM2 to regulate NF-κB-induced CCL5 secretion. Mol Cancer. 2022; 21:217. https://doi.org/10.1186/s12943-022-01686-7 [PubMed]
- 22. Hu X, Wu D, He X, Zhao H, He Z, Lin J, Wang K, Wang W, Pan Z, Lin H, Wang M. circGSK3β promotes metastasis in esophageal squamous cell carcinoma by augmenting β-catenin signaling. Mol Cancer. 2019; 18:160. https://doi.org/10.1186/s12943-019-1095-y [PubMed]
- 23. Li J, Song Y, Cai H, Zhou B, Ma J. Roles of circRNA dysregulation in esophageal squamous cell carcinoma tumor microenvironment. Front Oncol. 2023; 13:1153207. https://doi.org/10.3389/fonc.2023.1153207 [PubMed]
- 24. Shen Y, Shao Y, Niu C, Ruan X, Zang Z, Nakyeyune R, Guo X, Liu F. Systematic Identification of circRNA-miRNA-mRNA Regulatory Network in Esophageal Squamous Cell Carcinoma. Front Genet. 2021; 12:580390. https://doi.org/10.3389/fgene.2021.580390 [PubMed]
- 25. Wang C, Zhou M, Zhu P, Ju C, Sheng J, Du D, Wan J, Yin H, Xing Y, Li H, He J, He F. IGF2BP2-induced circRUNX1 facilitates the growth and metastasis of esophageal squamous cell carcinoma through miR-449b-5p/FOXP3 axis. J Exp Clin Cancer Res. 2022; 41:347. https://doi.org/10.1186/s13046-022-02550-8 [PubMed]
- 26. Zhou Q, Lei C, Cui F, Chen H, Cao X. Circ-ATIC regulates esophageal squamous cell carcinoma growth and metastasis through miR-1294/PBX3 pathway. Heliyon. 2023; 9:e12916. https://doi.org/10.1016/j.heliyon.2023.e12916 [PubMed]
- 27. Song D, Ye Z, Chen F, Zhan L, Sun X. circFNDC3B promotes esophageal squamous cell carcinoma progression by targeting MYO5A via miR-370-3p/miR-136-5p. BMC Cancer. 2023; 23:821. https://doi.org/10.1186/s12885-023-11314-2 [PubMed]
- 28. Zhang Y, Yuan X, Yue N, Wang L, Liu J, Dai N, Yang H, Fan R, Zhou F. hsa_circRNA6448-14 promotes carcinogenesis in esophageal squamous cell carcinoma. Aging (Albany NY). 2020; 12:15581–602. https://doi.org/10.18632/aging.103650 [PubMed]
- 29. Zhao R, Chen P, Qu C, Liang J, Cheng Y, Sun Z, Tian H. Circular RNA circTRPS1-2 inhibits the proliferation and migration of esophageal squamous cell carcinoma by reducing the production of ribosomes. Cell Death Discov. 2023; 9:5. https://doi.org/10.1038/s41420-023-01300-9 [PubMed]
- 30. Wang X, Liu Z, Du Y, Hao S, Zhao B. Hsa_circ_0043603 promotes the progression of esophageal squamous cell carcinoma by sponging miR-1178-3p and regulating AADAC expression. Heliyon. 2023; 9:e19807. https://doi.org/10.1016/j.heliyon.2023.e19807 [PubMed]
- 31. Feng Z, Wu J. hsa_circ_0129047 Upregulates LYVE1 to Inhibit Hepatocellular Carcinoma Progression by Sponging miR-492. Dis Markers. 2023; 2023:6978234. https://doi.org/10.1155/2023/6978234 [PubMed]
- 32. Zhao Y, Song J, Dong W, Liu X, Yang C, Wang D, Xue Y, Ruan X, Liu L, Wang P, Zhang M, Liu Y. The MBNL1/circNTRK2/PAX5 pathway regulates aerobic glycolysis in glioblastoma cells by encoding a novel protein NTRK2-243aa. Cell Death Dis. 2022; 13:767. https://doi.org/10.1038/s41419-022-05219-4 [PubMed]
- 33. Fan Q, Lu Q, Wang G, Zhu W, Teng L, Chen W, Bi L. Optimizing component formula suppresses lung cancer by blocking DTL-mediated PDCD4 ubiquitination to regulate the MAPK/JNK pathway. J Ethnopharmacol. 2022; 299:115546. https://doi.org/10.1016/j.jep.2022.115546 [PubMed]
- 34. Tang Q, Chen J, Di Z, Yuan W, Zhou Z, Liu Z, Han S, Liu Y, Ying G, Shu X, Di M. TM4SF1 promotes EMT and cancer stemness via the Wnt/β-catenin/SOX2 pathway in colorectal cancer. J Exp Clin Cancer Res. 2020; 39:232. https://doi.org/10.1186/s13046-020-01690-z [PubMed]
- 35. Li L, Liang Y, Kang L, Liu Y, Gao S, Chen S, Li Y, You W, Dong Q, Hong T, Yan Z, Jin S, Wang T, et al. Transcriptional Regulation of the Warburg Effect in Cancer by SIX1. Cancer Cell. 2018; 33:368–85.e7. https://doi.org/10.1016/j.ccell.2018.01.010 [PubMed]
- 36. Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019; 20:675–91. https://doi.org/10.1038/s41576-019-0158-7 [PubMed]
- 37. Chelakkot C, Chelakkot VS, Shin Y, Song K. Modulating Glycolysis to Improve Cancer Therapy. Int J Mol Sci. 2023; 24:2606. https://doi.org/10.3390/ijms24032606 [PubMed]
- 38. Jiang H, Wei H, Wang H, Wang Z, Li J, Ou Y, Xiao X, Wang W, Chang A, Sun W, Zhao L, Yang S. Zeb1-induced metabolic reprogramming of glycolysis is essential for macrophage polarization in breast cancer. Cell Death Dis. 2022; 13:206. https://doi.org/10.1038/s41419-022-04632-z [PubMed]
- 39. Wu Y, Chen Z, Xie G, Zhang H, Wang Z, Zhou J, Chen F, Li J, Chen L, Niu H, Wang H. RNA m1A methylation regulates glycolysis of cancer cells through modulating ATP5D. Proc Natl Acad Sci U S A. 2022; 119:e2119038119. https://doi.org/10.1073/pnas.2119038119 [PubMed]
- 40. Li Z, Peng Y, Li J, Chen Z, Chen F, Tu J, Lin S, Wang H. N6-methyladenosine regulates glycolysis of cancer cells through PDK4. Nat Commun. 2020; 11:2578. https://doi.org/10.1038/s41467-020-16306-5 [PubMed]
- 41. Zhang Z, Li X, Yang F, Chen C, Liu P, Ren Y, Sun P, Wang Z, You Y, Zeng YX, Li X. DHHC9-mediated GLUT1 S-palmitoylation promotes glioblastoma glycolysis and tumorigenesis. Nat Commun. 2021; 12:5872. https://doi.org/10.1038/s41467-021-26180-4 [PubMed]
- 42. Yu T, Wang Y, Fan Y, Fang N, Wang T, Xu T, Shu Y. CircRNAs in cancer metabolism: a review. J Hematol Oncol. 2019; 12:90. https://doi.org/10.1186/s13045-019-0776-8 [PubMed]
- 43. Zhou S, Guo Z, Lv X, Zhang X. CircGOT1 promotes cell proliferation, mobility, and glycolysis-mediated cisplatin resistance via inhibiting its host gene GOT1 in esophageal squamous cell cancer. Cell Cycle. 2022; 21:247–60. https://doi.org/10.1080/15384101.2021.2015671 [PubMed]
- 44. Ma L, Li H, Lin Y, Wang G, Xu Q, Chen Y, Xiao K, Rao X. CircDUSP16 Contributes to Cell Development in Esophageal Squamous Cell Carcinoma by Regulating miR-497-5p/TKTL1 Axis. J Surg Res. 2021; 260:64–75. https://doi.org/10.1016/j.jss.2020.11.052 [PubMed]
- 45. Alsayed RKM, Sheikhan KSA, Alam MA, Buddenkotte J, Steinhoff M, Uddin S, Ahmad A. Epigenetic programing of cancer stemness by transcription factors-non-coding RNAs interactions. Semin Cancer Biol. 2023; 92:74–83. https://doi.org/10.1016/j.semcancer.2023.04.005 [PubMed]
- 46. Lin Z, Tang X, Wan J, Zhang X, Liu C, Liu T. Functions and mechanisms of circular RNAs in regulating stem cell differentiation. RNA Biol. 2021; 18:2136–49. https://doi.org/10.1080/15476286.2021.1913551 [PubMed]
- 47. Yu W, He X, Zhang C, Huang F. Circular RNA circSLC7A11 contributes to progression and stemness of laryngeal squamous cell carcinoma via sponging miR-877-5p from LASP1. Heliyon. 2023; 9:e18290. https://doi.org/10.1016/j.heliyon.2023.e18290 [PubMed]
- 48. Li L, Lei K, Lyu Y, Tan B, Liang R, Wu D, Wang K, Wang W, Lin H, Wang M. hsa_circ_0001741 promotes esophageal squamous cell carcinoma stemness, invasion and migration by sponging miR-491-5p to upregulate NOTCH3 expression. Am J Cancer Res. 2022; 12:2012–31. [PubMed]
- 49. Vasefifar P, Motafakkerazad R, Maleki LA, Najafi S, Ghrobaninezhad F, Najafzadeh B, Alemohammad H, Amini M, Baghbanzadeh A, Baradaran B. Nanog, as a key cancer stem cell marker in tumor progression. Gene. 2022; 827:146448. https://doi.org/10.1016/j.gene.2022.146448 [PubMed]
- 50. Mohiuddin IS, Wei SJ, Kang MH. Role of OCT4 in cancer stem-like cells and chemotherapy resistance. Biochim Biophys Acta Mol Basis Dis. 2020; 1866:165432. https://doi.org/10.1016/j.bbadis.2019.03.005 [PubMed]
- 51. Novak D, Hüser L, Elton JJ, Umansky V, Altevogt P, Utikal J. SOX2 in development and cancer biology. Semin Cancer Biol. 2020; 67:74–82. https://doi.org/10.1016/j.semcancer.2019.08.007 [PubMed]
- 52. Nanao MH, Tcherniuk SO, Chroboczek J, Dideberg O, Dessen A, Balakirev MY. Crystal structure of human otubain 2. EMBO Rep. 2004; 5:783–8. https://doi.org/10.1038/sj.embor.7400201 [PubMed]
- 53. Zhang Z, Du J, Wang S, Shao L, Jin K, Li F, Wei B, Ding W, Fu P, van Dam H, Wang A, Jin J, Ding C, et al. OTUB2 Promotes Cancer Metastasis via Hippo-Independent Activation of YAP and TAZ. Mol Cell. 2019; 73:7–21.e7. https://doi.org/10.1016/j.molcel.2018.10.030 [PubMed]
- 54. Li J, Cheng D, Zhu M, Yu H, Pan Z, Liu L, Geng Q, Pan H, Yan M, Yao M. OTUB2 stabilizes U2AF2 to promote the Warburg effect and tumorigenesis via the AKT/mTOR signaling pathway in non-small cell lung cancer. Theranostics. 2019; 9:179–95. https://doi.org/10.7150/thno.29545 [PubMed]
- 55. Liu L, Cheng H, Ji M, Su L, Lu Z, Hu X, Guan Y, Xiao J, Ma L, Zhang W, Pu H. OTUB2 Regulates YAP1/TAZ to Promotes the Progression of Esophageal Squamous Cell Carcinoma. Biol Proced Online. 2022; 24:10. https://doi.org/10.1186/s12575-022-00169-9 [PubMed]
- 56. Chang W, Luo Q, Wu X, Nan Y, Zhao P, Zhang L, Luo A, Jiao W, Zhu Q, Fu Y, Liu Z. OTUB2 exerts tumor-suppressive roles via STAT1-mediated CALML3 activation and increased phosphatidylserine synthesis. Cell Rep. 2022; 41:111561. https://doi.org/10.1016/j.celrep.2022.111561 [PubMed]
- 57. Wu X, Wang H, Zhu D, Chai Y, Wang J, Dai W, Xiao Y, Tang W, Li J, Hong L, Pei M, Zhang J, Lin Z, et al. USP3 promotes gastric cancer progression and metastasis by deubiquitination-dependent COL9A3/COL6A5 stabilisation. Cell Death Dis. 2021; 13:10. https://doi.org/10.1038/s41419-021-04460-7 [PubMed]
- 58. Shi J, Zhang Q, Yin X, Ye J, Gao S, Chen C, Yang Y, Wu B, Fu Y, Zhang H, Wang Z, Wang B, Zhu Y, et al. Stabilization of IGF2BP1 by USP10 promotes breast cancer metastasis via CPT1A in an m6A-dependent manner. Int J Biol Sci. 2023; 19:449–64. https://doi.org/10.7150/ijbs.76798 [PubMed]
- 59. Feng Y, Xia S, Hui J, Xu Y. Circular RNA circBNC2 facilitates glycolysis and stemness of hepatocellular carcinoma through the miR-217/high mobility group AT-hook 2 (HMGA2) axis. Heliyon. 2023; 9:e17120. https://doi.org/10.1016/j.heliyon.2023.e17120 [PubMed]