



Introduction
In theory, living organisms can avoid age-related death for a potentially unlimited period of time [1,2]. This is because from the point of view of thermodynamics living organisms are open self-organizing systems, i.e. they can use exogenous energy to resist a progressive increase in entropy and the resulting molecular damage and disorder as they age [1,2]. Yet, it is well known that organismal lifespan 1) has a limit that is unique to each species; and 2) varies drastically between different species [3–6]. Since late XIX century, numerous evolutionary theories of aging have been proposed in an attempt to resolve this paradox [1–6]. Theories of programmed aging postulate that the evolutionary force actively restricts organismal lifespan at a certain age distinctive for each species [2,5–17], whereas theories of non-programmed aging assume that lack of such evolutionary force passively restrains organismal lifespan at a species-specific age [2,4–6,15–17]. These two groups of evolutionary aging theories are discussed below.
The first evolutionary theory of aging, known as the theory of programmed death, was developed by August Weismann. According to this theory, natural selection resulted in the preferential reproduction of those members of a particular species that are able to die when they reach a certain age, which is unique to this species [5,6,17,18]. By undergoing a ″programmed″ death at such species-specific age, older members of this species are eliminated from a competition with their younger counterparts for natural resources [5,6,17,18]. In the programmed death theory, the evolutionary advantage to having a limited lifespan at a species-specific age consists in providing a benefit to survival of a group of individuals by creating a disadvantage to those individuals within the group that has reached such an age [5,6,17,18].
Recent advances in the understanding of molecular mechanisms underlying cellular aging and organismal longevity marked a Renaissance period in developing evolutionary theories of programmed aging and age-related death [1,2,4–17]. These relatively recently developed theories include the following: 1) group selection theory [5,6,15,17]; 2) kin selection theory [5,6,15,17,19]; 3) evolvability theory [5,6,15,17,20]; 4) phenoptosis theory [6–9,21]; and 5) altruistic aging theory [12–14,22–25]. Akin to the theory of programmed death developed by August Weismann [5,6,17,18], all these contemporary evolutionary theories of programmed aging are based on the notion that natural selection resulted in preferential reproduction of those members of various species that have evolved certain active mechanisms for limiting their lifespans in a species-specific fashion and upon reaching a species-specific age [5–9,12,15,17,19–21].
Recent studies have provided evidence favoring evolutionary theories of programmed aging and age-related death. In particular, it has been shown that cellular aging can be delayed and organismal longevity can be extended by some genetic, dietary and pharmacological interventions that attenuate certain pro-aging signaling pathways that control the rate of aging [26–59]. These pro-aging signaling pathways operate as active mechanisms that (according to evolutionary theories of programmed aging) can limit organismal lifespan at a specific age. It is conceivable therefore that these pathways have evolved to restrict organismal lifespan at a particular age characteristic of each group of evolutionarily distant organisms.
One of the key features of all contemporary evolutionary theories of programmed aging and age-related death is that longevity-extending genetic traits attenuating different pro-aging signaling pathways may or may not reduce early-life fitness; is has been proposed that early-life fitness can only be decreased by those genetic traits that impair the pro-aging signaling pathways essential for the development of fitness early in life [12,60–73]. Early-life fitness is known to include the following features: 1) metabolic rate under various environmental conditions; 2) growth rate and, in yeast, the ability to utilize alternative carbon sources; 3) physical activity; 4) fecundity - i.e. the efficacies of mating and reproduction (including sporulation in yeast); 5) resistance to fluctuations in temperature, light, humidity and other environmental factors (such as osmolarity fluctuations in yeast); and 6) susceptibility to environmental toxins [12,60–73]. Until now the effects of various longevity-extending genetic traits on early-life fitness have been analyzed mainly under laboratory conditions in which long-lived mutants of a certain species were growing and undergoing aging alone, in the absence of ″wild-type (WT)″ individuals of the same species; these WT individuals do not carry any longevity-extending genetic traits and thus do not have lifespan extended beyond a species-specific age [26–51,53–55,74,75]. However, these laboratory conditions do not mimic the process of natural selection within a mixed population of individuals of the same species. Under such conditions of natural selection, different individuals within the population 1) possess different longevity-defining genetic backgrounds; 2) have lifespans at a species-specific age and above it; and 3) compete for nutrients and other environmental resources [73,76–81].
Unlike the evolutionary theories of programmed aging and age-related death, all evolutionary theories of non-programmed aging posit that organismal lifespan is limited at an age characteristic of each species due to lack of the evolutionary force [2,4–6,15–17]. These theories include the following: 1) the mutation accumulation theory [5,6,15,17,82,83] and its modified version known as the late-life mortality plateau theory [5,6,15,17,84]; and 2) the antagonistic pleiotropy theory [5,6,15,17,85] and its contemporary version called the disposable soma theory [5,6,15,17,86–88]. Both, the mutation accumulation theory and the late-life mortality plateau theory, postulate that natural selection favours alleles of a gene that are beneficial early in organismal life over alleles of the same gene that provide an advantage late in life of this organism [5,6,15,17,82–84]. Thus, by eliminating gene alleles that are beneficial late in life, natural selection will diminish its power with age of an organism and will limit its lifespan at an age that is unique to each species [5,6,15,17,82–84]. In contrast, the antagonistic pleiotropy theory and the disposable soma theory assume that alleles of certain genes that are beneficial in early life of an organism exhibit detrimental effects in its late life [5,6,15,17,85–88]. Because different alleles of these genes display age-related antagonistic effects on several fitness-defining traits of an organism, these genes are called pleiotropic genes. According to both the antagonistic pleiotropy theory and the disposable soma theory, natural selection limits organismal lifespan at an age unique to each species by actively retaining only those alleles of pleiotropic genes that increase early-life fitness and thus reduce fitness at old age [5,6,15,17,85–88].
Noteworthy, contemporary evolutionary theories of programmed aging and age-related death postulate that organisms of all species possess mechanisms that have been evolved to actively limit their lifespans at a species-specific age [5–9,12,15,17,19–21]. In contrast, evolutionary theories of non-programmed aging assume that such mechanisms cannot exist, just because organismal lifespan is limited at a species-specific age passively - i.e. due to lack of the evolutionary force [5,6,15,17,82–88]. It was therefore concluded that the demonstrated ability of certain genetic, dietary and pharmacological interventions to extend lifespan in evolutionarily distant species by targeting mechanisms that actively limit organismal lifespan at a species-specific age [26–59] validates evolutionary theories of programmed aging and invalidates evolutionary theories of non-programmed aging [5–17]. However, in all these cases the ability of genetic, dietary and pharmacological interventions to prolong organismal lifespan has been revealed under laboratory conditions. As discussed above, these conditions do not imitate the process of natural selection within a mixed population of same-species individuals having different longevity-defining genetic backgrounds [73,76–81]. But none of the evolutionary theories of non-programmed aging assumes that in the absence of natural selection (i.e. under laboratory conditions) longevity-extending mutant gene alleles decreasing early-life fitness cannot exist; all these theories only proclaim that such mutant gene alleles will be eliminated from the gene pool of a species under the pressure of natural selection (i.e. in the wild or under field-like laboratory conditions) [5,6,15,17,82–88]. Furthermore, it seems impossible in the wild or under field-like laboratory conditions to impose any of the currently known longevity-extending dietary or pharmacological interventions (such as caloric restriction [CR], dietary restriction [DR] or aging-delaying chemical compounds) only on some individuals of the same species; thus, it is unlikely that such non-genetic interventions can be used for empirical verification of evolutionary theories of programmed or non-programmed aging.
We have recently conducted the experimental evolution of long-lived yeast species by a lasting exposure to exogenous lithocholic bile acid (LCA) (Gomez-Perez et al., submitted). We selected 3 long-lived mutants capable of sustaining their greatly extended chronological lifespans (CLS) after numerous passages in medium without LCA (Gomez-Perez et al., submitted). The extended longevity of each of these yeast mutants is a dominant polygenic trait caused by mutations in more than two genes (Gomez-Perez et al., submitted). The objective of this study was to use these long-lived yeast mutants for the empirical verification of evolutionary theories of programmed or non-programmed aging. To attain this objective, we investigated if the dominant polygenic trait extending longevity of each of these mutants affects such key features of early-life fitness as the exponential growth rate, efficacy of post-exponential growth, fecundity, and resistance to apoptotic and liponecrotic forms of programmed cell death. We also examined if any of these long-lived mutants can be forced out of an ecosystem by the parental WT strain exhibiting shorter lifespan; these experiments were carried out under laboratory conditions mimicking the process of natural selection within an ecosystem composed of yeast cells having different longevity-defining genetic backgrounds.
Results
Dominant polygenic trait extending longevity of each of the 3 long-lived yeast mutants does not affect some key features of early-life fitness and enhance other such features
To empirically verify evolutionary theories of programmed or non-programmed aging, we elucidated if the dominant polygenic trait that extends longevity of each of the 3 selected long-lived yeast mutants affects early-life fitness when each mutant grows and ages alone – i.e. in the absence of a parental WT strain. The following key features of early-life fitness were measured: the exponential growth rate and efficacy of post-exponential growth, fecundity, and resistance to apoptotic and liponecrotic forms of programmed cell death.
We first assessed if the long-lived mutant strains 3, 5 and/or 12 exhibit altered exponential growth rate and/or efficacy of post-exponential growth in media containing 1) a fermentable carbon source - i.e. glucose at the initial concentration of 0.2% [CR conditions] or 2% [non-CR conditions]; and 2) a non-fermentable carbon source - i.e. ethanol at the initial concentration of 1% or glycerol at the initial concentration of 3%. In these experiments, we used the single-gene-deletion mutant strains rpp2BΔ and dbp3Δ as controls. Each of these mutant strains is known to exhibit extended replicative lifespan (RLS) and reduced growth rate on 2% glucose [73]. dbp3Δ is also known to have prolonged CLS [89]. rpp2BΔ lacks a gene encoding ribosomal protein P2 beta, whereas dbp3Δ lacks a gene encoding a DEAD-box family protein involved in ribosomal biogenesis [73]. By monitoring the OD600 of cell cultures recovered at different time points as a measure of cell growth, we found that the long-lived mutant strains 3, 5 and 12 do not differ from the parental WT strain BY4742 in the exponential growth rates and post-exponential growth efficacies in medium initially containing 0.2% glucose, 2% glucose, 1% ethanol or 3% glycerol (Figures 1A, 1B, 1C and 1D, respectively). Of note, the control strain rpp2BΔ exhibited a reduced growth rate in medium initially containing any of these four carbon sources, whereas the control strain dbp3Δ displayed a decreased growth rate in medium initially containing 0.2% glucose or 2.0% glucose (Figure 1). Moreover, the control strain rpp2BΔ exhibited a significantly reduced efficiency of post-exponential growth in medium initially containing 3% glycerol (Figure 1D).
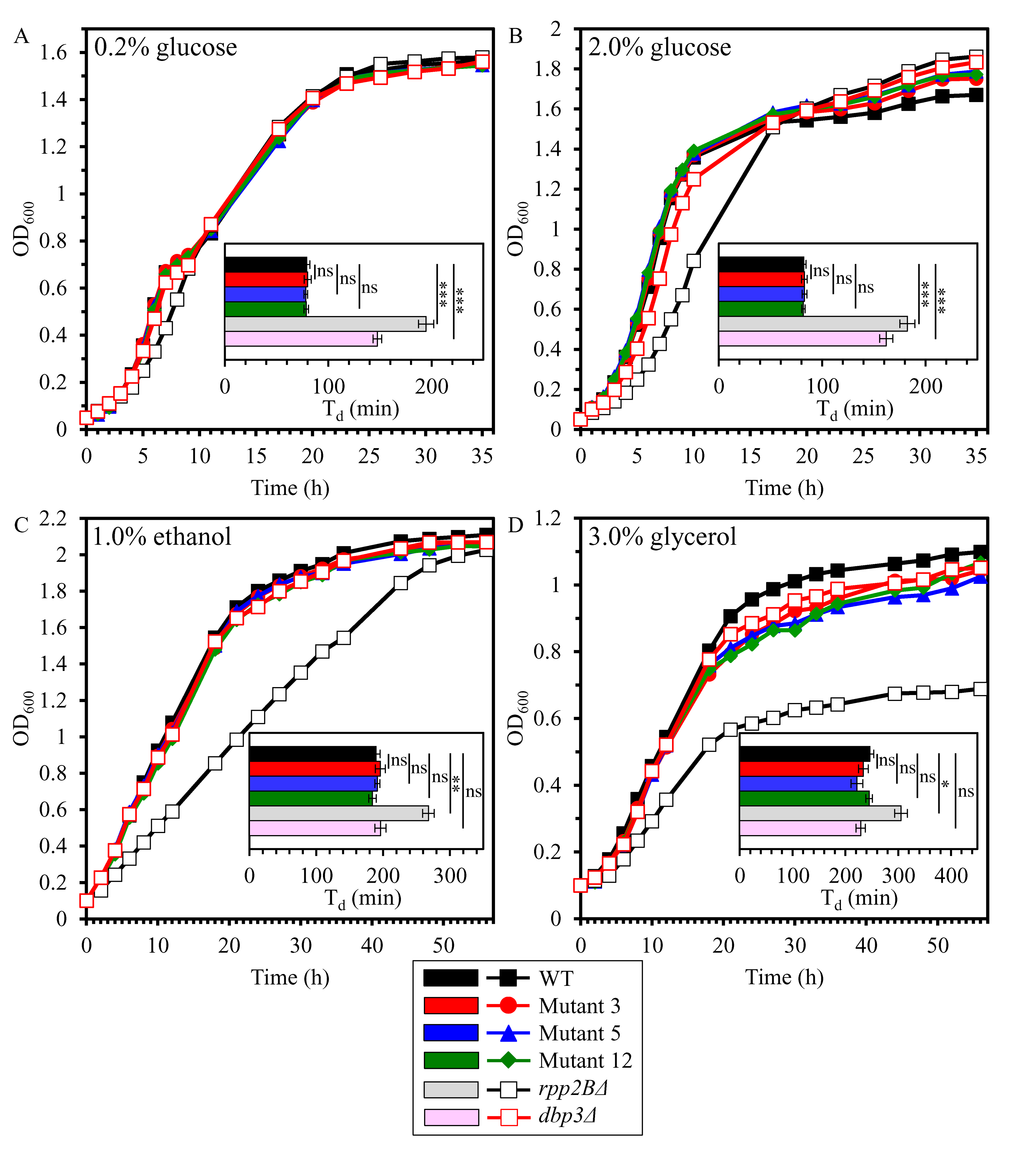
Figure 1. The long-lived mutant strains 3, 5 and 12 do not differ from the parental WT strain in the exponential growth rates and post-exponential growth efficacies in medium initially containing fermentable or non-fermentable carbon source. The parental haploid WT strain BY4742, long-lived mutant strains 3, 5 and 12 (each in the BY4742 genetic background), and the single-gene-deletion mutant strains rpp2BΔ and dbp3Δ (each in the BY4742 genetic background) were cultured in YP medium initially containing 0.2% glucose (A), 2.0% glucose (B), 1.0% ethanol (C) or 3.0% glycerol (D). The OD600 of cell cultures recovered at different time points was measured. Growth curves are shown; data for growth curves are presented as means (n = 3). For each strain, a doubling time (min) was calculated as Td = (t2 – t1) × log 2/log (OD2/OD1), where: t2 = a given time point; t1 = an earlier time point; OD2 = OD600 at a given time point; OD1 = OD600 at an earlier time point. Data for the values of Td are presented as means ± SEM (n = 3; ns, not significant; *p < 0.05; **p < 0.01; ***p < 0.001).
We then elucidated if the long-lived mutant strains 3, 5 and/or 12 exhibit altered efficacy of their sexual reproduction by mating, one of the measures of fecundity. In these experiments, yeast cells of mating type MATa (i.e. the haploid WT strain BY4741) and mating type MATα (i.e. the haploid WT strain BY4742 or the selected long-lived haploid mutant strains 3, 5 or 12, all in the BY4742 genetic background) were pre-grown separately to mid-logarithmic phase in YP medium initially containing 0.2% glucose or 1% ethanol. The efficiency of mating was measured as described in the ″Materials and methods″ section; it was calculated as the number of colonies of MATa/MATα diploids divided by the sum of MATa/MATα diploids plus haploid colonies. Crosses between two WT strains of opposite mating types (i.e. the haploid strain BY4741 [MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0]) and the haploid strain BY4742 [MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0]) were used as controls. We found that the long-lived mutant strains 3, 5 and 12 do not differ from the parental WT strain BY4742 in efficacy of their sexual reproduction by mating if pre-grown in medium initially containing 0.2% glucose or 1% ethanol (Figures 2A and 2B, respectively).
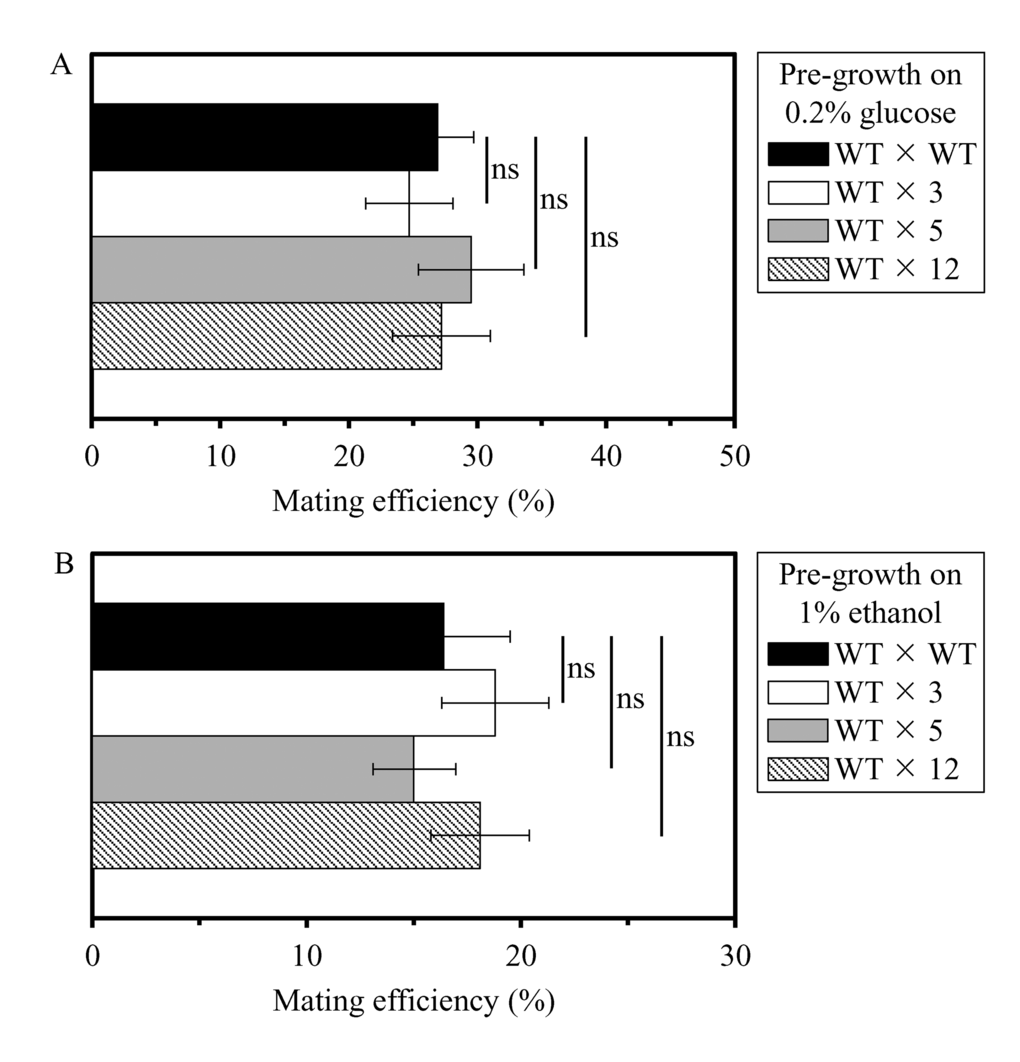
Figure 2. The long-lived mutant strains 3, 5 and 12 do not differ from the parental WT strain in efficacy of their sexual reproduction by mating, a measure of fecundity and a key trait of early-life fitness. Yeast cells of mating type MATa (i.e. the haploid WT strain BY4741) and mating type MATα (i.e. the haploid WT strain BY4742 or the selected long-lived haploid mutant strains 3, 5 or 12, each in the BY4742 genetic background) were pre-grown separately to mid-logarithmic phase in YP medium initially containing 0.2% glucose (a fermentable carbon source; CR conditions) (A) or 1% ethanol (a non-fermentable carbon source) (B). The efficiency of mating was measured as described in the ″Materials and methods″ section; it was calculated as the number of colonies of MATa/MATα diploids divided by the sum of MATa/MATα diploids plus haploid colonies. Data are presented as means ± SEM (n = 3; ns, not significant difference).
We then investigated if the long-lived mutant strains 3, 5 and/or 12 display altered efficacy of their sexual reproduction by sporulation, another measure of fecundity. In these experiments, each of the four diploid strains formed between cells of the haploid WT strain BY4741 (MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0) and cells of the haploid WT strain BY4742 (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0) or cells of each of the selected long-lived haploid mutant strains 3, 5 or 12 (each in the BY4742 genetic background) were pre-grown to mid-logarithmic phase in YP medium initially containing 0.2% glucose or 1% ethanol. The efficiency of sporulation of each of the four diploid strains was then measured at various time points since the beginning of a sporulation assay as described in the ″Materials and methods″ section; it was calculated as the percentage of tetrads and dyads produced by a diploid strain, relative to the total number of cells. We found that the long-lived mutant strains 3, 5 and 12 do not differ from the parental WT strain BY4742 in efficacy of their sexual reproduction by sporulation when cells of the hybrid each of them formed with the haploid WT strain BY4741 of opposite mating type were pre-grown in medium initially containing 0.2% glucose or 1% ethanol (Figures 3A and 3B, respectively).
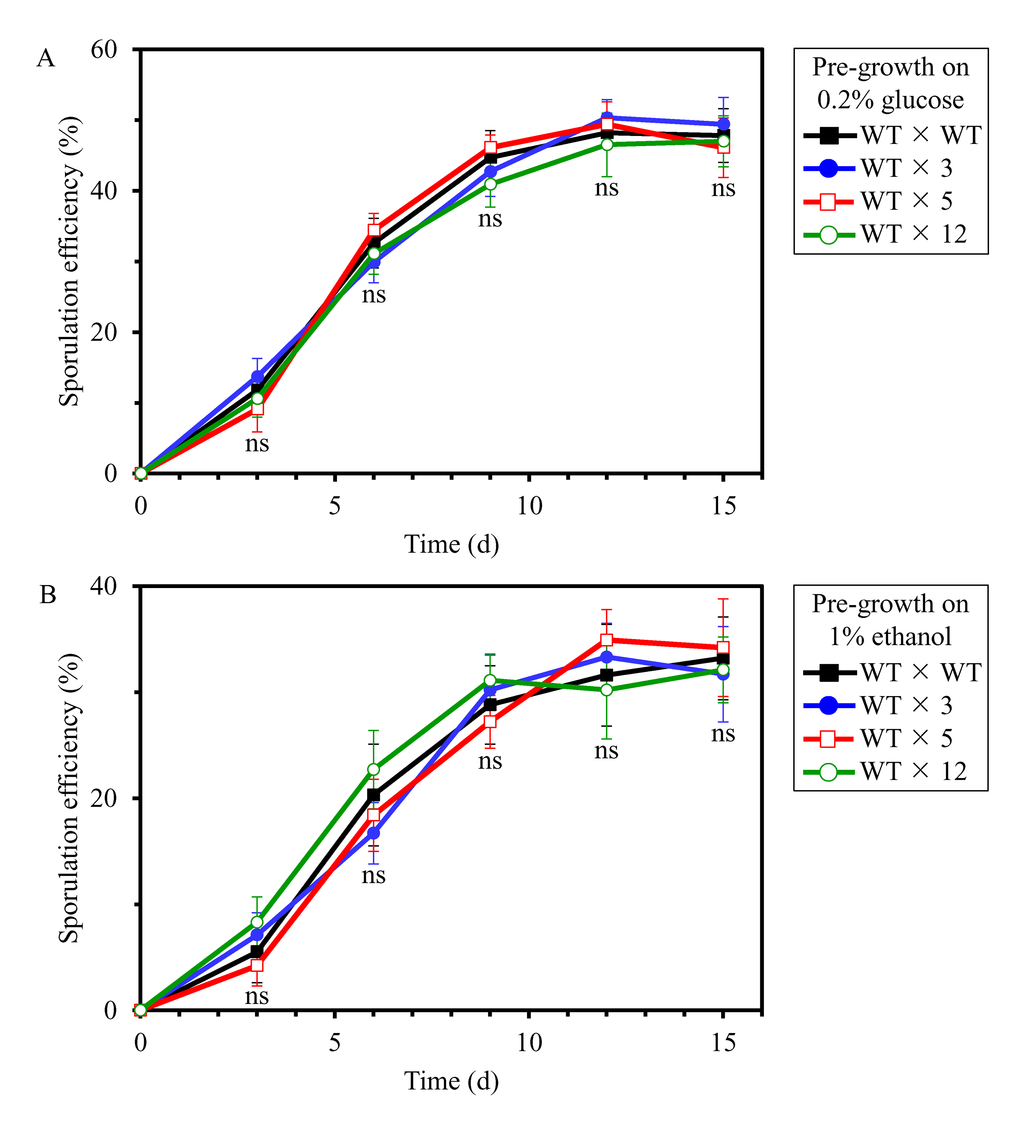
Figure 3. The long-lived mutant strains 3, 5 and 12 do not differ from the parental WT strain in efficacy of their sexual reproduction by sporulation, a measure of fecundity and a key trait of early-life fitness. Each of the four diploid strains formed between cells of the haploid WT strain BY4741 (MATa) and cells of the haploid WT strain BY4742 (MATα) or cells of each of the selected long-lived haploid mutant strains 3, 5 or 12 (each MATα in the BY4742 genetic background) were pre-grown to mid-logarithmic phase in YP medium initially containing 0.2% glucose (a fermentable carbon source; CR conditions) (A) or 1% ethanol (a non-fermentable carbon) (B). The efficiency of sporulation of each of the four diploid strains was measured at various time points since the beginning of a sporulation assay as described in the ″Materials and methods″ section; it was calculated as the percentage of tetrads and dyads produced by a diploid strain, relative to the total number of cells. Data are presented as means ± SEM (n = 3; ns, not significant difference). At each time point, sporulation efficiencies of the WT × 3, WT × 5 and WT × 12 diploid strains were statistically insignificant in comparison with sporulation efficiency of the WT × WT diploid strain.
We also assessed if the dominant polygenic trait that extends longevity of each of the 3 long-lived mutant strains affects two other essential aspects of early-life fitness, namely 1) cell susceptibility to a mitochondria-controlled apoptotic form of death triggered by a brief exposure to exogenous hydrogen peroxide [48,90–99]; and 2) cell susceptibility to a ″liponecrotic″ form of death elicited by a short-term exposure to exogenous palmitoleic acid [48,100–103]. We found that the long-lived mutant strains 3, 5 and 12 exhibit enhanced (as compared to the parental WT strain BY4742) susceptibilities to 1) mitochondria-controlled apoptotic death of yeast cells pre-grown in media initially containing 0.2% glucose or 1% ethanol (Figures 4A and 4B, respectively); and 2) liponecrotic death of yeast cells pre-grown in media initially containing 0.2% glucose or 1% ethanol (Figures 5A and 5B, respectively).
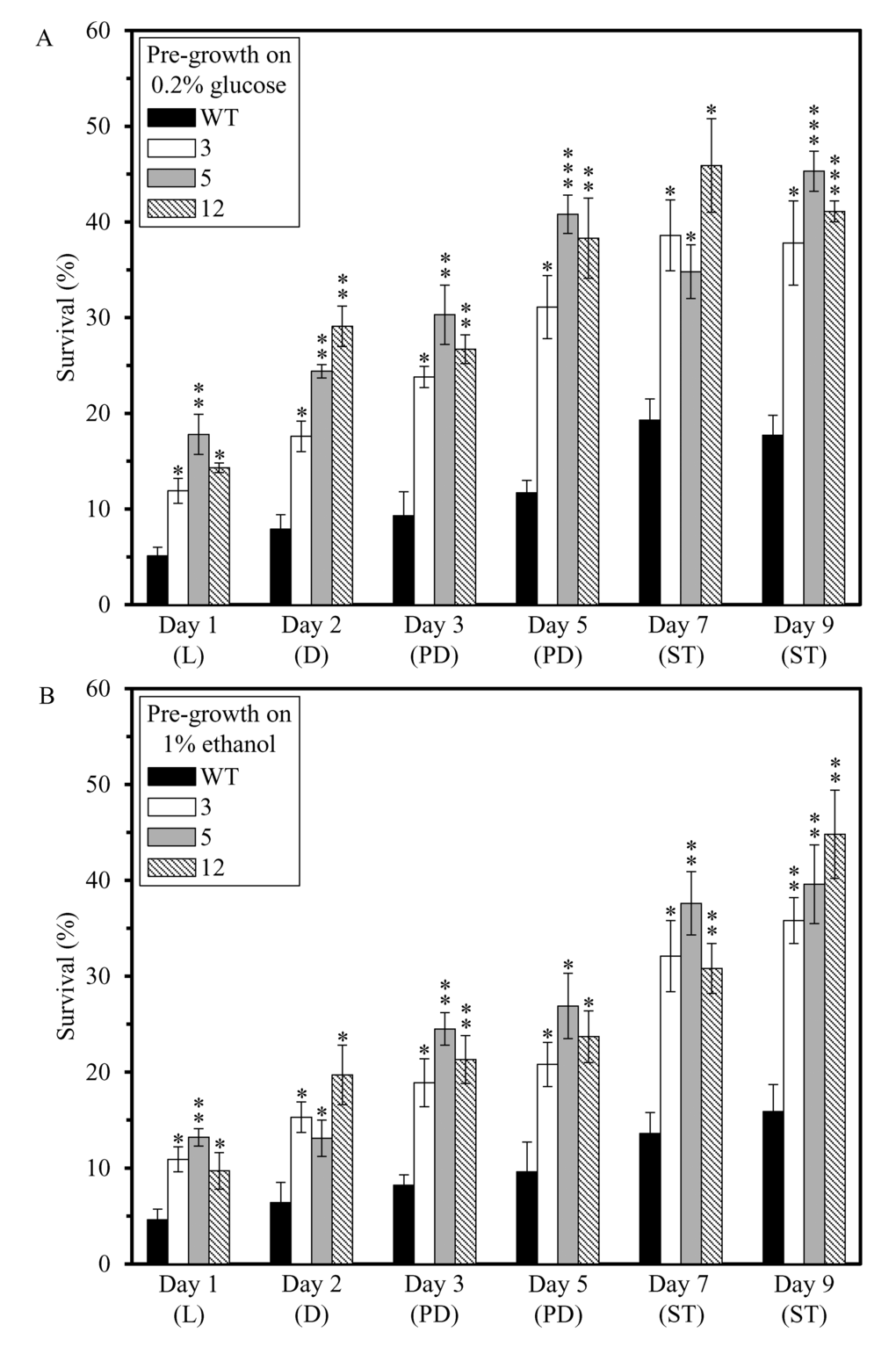
Figure 4. The long-lived mutant strains 3, 5 and 12 exhibit enhanced (as compared to the parental WT strain) susceptibilities to a mitochondria-controlled apoptotic form of cell death, one of the traits of early-life fitness. The parental WT strain BY4742 and long-lived mutant strains 3, 5 and 12 (each in the BY4742 genetic background) were cultured in YP medium initially containing 0.2% glucose (a fermentable carbon source; CR conditions) (A) or 1% ethanol (a non-fermentable carbon source) (B). Cell aliquots were recovered from various growth phases and then treated for 2 h with 2.5 mM hydrogen peroxide to induce mitochondria-controlled apoptosis. The % of viable cells was calculated as described in in the ″Materials and methods″ section. D, diauxic growth phase; L, logarithmic growth phase; PD, post-diauxic growth phase; ST, stationary growth phase. Data originate are presented as means ± SEM (n = 3; *p < 0.05; **p < 0.01; ***p < 0.001).
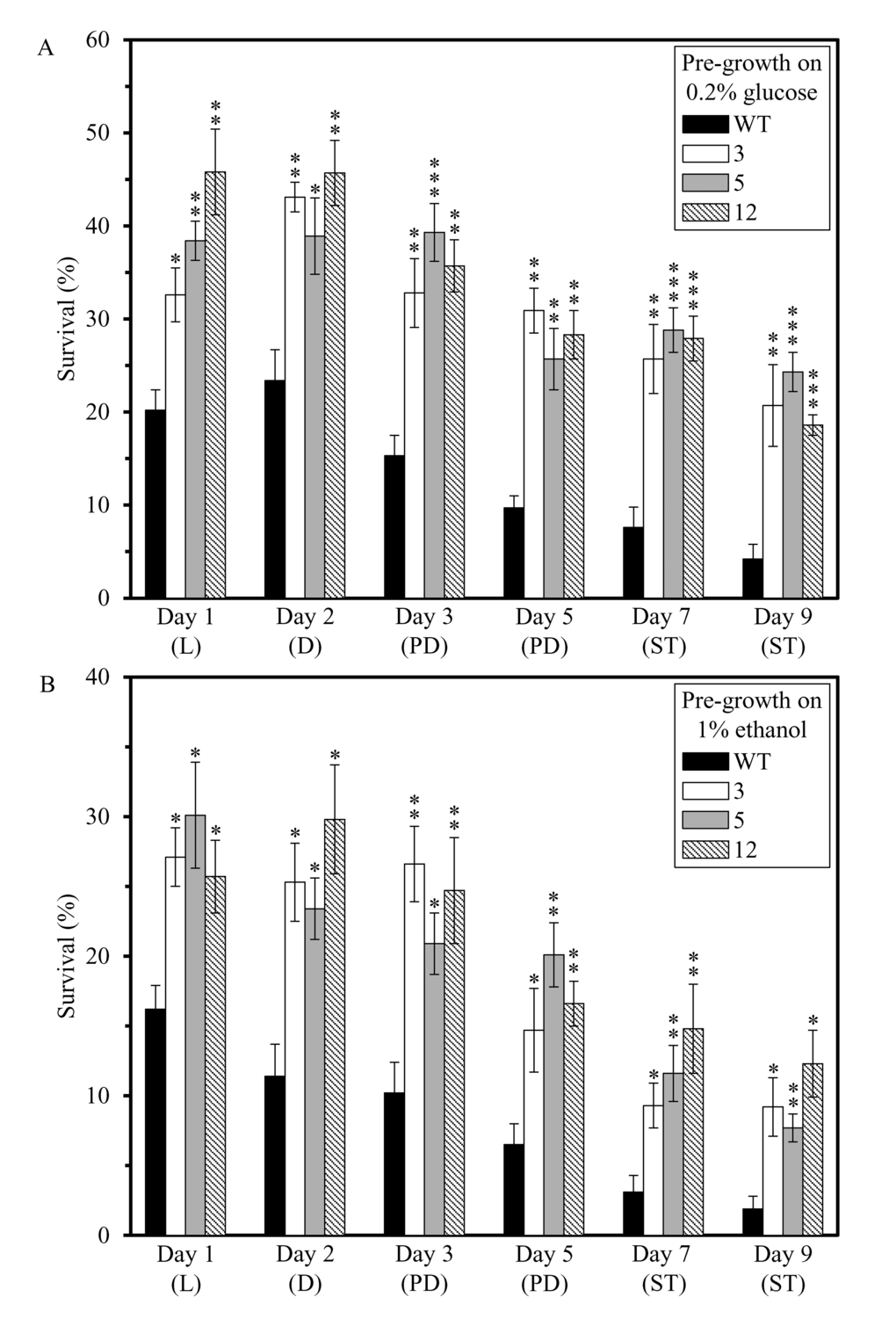
Figure 5. The long-lived mutant strains 3, 5 and 12 exhibit enhanced (as compared to the parental WT strain) susceptibilities to a liponecrotic form of cell death, one of the traits of early-life fitness. The parental WT strain BY4742 and long-lived mutant strains 3, 5 and 12 (each in the BY4742 genetic background) were cultured in YP medium initially containing 0.2% glucose (a fermentable carbon source; CR conditions) (A) or 1% ethanol (a non-fermentable carbon source) (B). Cell aliquots were recovered from various growth phases and then exposed for 2 h to 0.2 mM palmitoleic acid to induce liponecrosis. The % of viable cells was calculated as described in in the ″Materials and methods″ section. D, diauxic growth phase; L, logarithmic growth phase; PD, post-diauxic growth phase; ST, stationary growth phase. Data originate are presented as means ± SEM (n = 3; *p < 0.05; **p < 0.01; ***p < 0.001).
Of note, our recent study revealed that the long-lived mutant strains 3, 5 and 12 show enhanced resistance to chronic oxidative, thermal and osmotic stresses (Gomez-Perez et al., submitted). Akin to cell susceptibility to apoptotic and liponecrotic forms of cell death, such resistance to acute stresses is one of the key traits of early-life fitness [12,48,58,61,65,99,100,104–109].
In sum, findings presented in this section and elsewhere (Gomez-Perez et al., submitted) imply that the dominant polygenic traits extending longevities of the long-lived mutant strains 3, 5 and 12 do not affect such key features of early-life fitness as the exponential growth rate, efficacy of post-exponential growth and fecundity. Moreover, these longevity-extending polygenic traits enhance such features of early-life fitness as susceptibility to chronic exogenous stresses, and the resistance to apoptotic and liponecrotic forms of programmed cell death.
Development and validation of a quantitative assay for assessing the relative fitness of a long-lived mutant strain that competes for nutrients with a parental WT strain
To investigate if the dominant polygenic trait that extends longevity of each of the 3 selected long-lived yeast mutants influences the relative fitness of the mutant when it competes for nutrients and other environmental resources with a parental WT strain, we developed a direct competition assay. In this assay (Figure 6), the WT strains BY4739 (MATα leu2Δ0 lys2Δ0 ura3Δ0) and BY4742 (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0), the single-gene-deletion mutant strain dbp3Δ (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0 dbp3Δ::kanMX4) in the BY4742 genetic background, and the long-lived mutant strains 3, 5 and 12 (each in the BY4742 genetic background) were grown separately in the liquid nutrient-rich YP medium initially containing 0.2% glucose, 2% glucose or 1% ethanol as carbon source until mid-exponential phase. The single-gene-deletion mutant strain dbp3Δ was used as a control mutant strain because it is known to exhibit 1) extended CLS [89] and RLS [73]; 2) a decreased growth rate on 0.2% glucose (see above), 2% glucose [73] and 1% ethanol (see above); and 3) a reduced relative fitness when it is co-cultured with a parental WT strain in medium initially containing 2% glucose [73,89]. Cells of the WT strain BY4739 were mixed with the same number of cells of BY4742, dbp3Δ or a selected long-lived mutant strain (i.e. mutant strain 3, 5 or 12) in liquid YP medium initially containing 0.2% glucose, 2% glucose or 1% ethanol as carbon source. After culturing the cell mixture for 7 days, an aliquot of cell suspension was diluted and plated on solid YP medium supplemented with 2% glucose. Following 2 days of incubation, colonies on each plate were replicated onto plates with the synthetic minimal YNB medium without amino acids and nucleotides supplemented with 2% glucose. One of these plates contained leucine, lysine, uracil and histidine (hereafter it is called a ″His+″ plate), whereas the other plate contained leucine, lysine and uracil (hereafter it is called a ″His-″ plate). After 2 days of incubation at 30oC, the number of CFU on ″His+″ and ″His-″ plates was counted. The relative fitness of each His+ strain (i.e. the BY4742, dbp3Δ, 3, 5 or 12 strain) in direct competition with the His- strain BY4739 was calculated as log2 [(CFUxmutant/CFUxWT/(CFU0mutant/CFU0WT)], where: CFUx is the colony count at the end of week x, whereas CFU0 is the colony count at initial inoculation of a mixed culture (Figure 6). In every experiment for measuring relative fitness, the direct competition step of culturing a cell mixture for 7 days in liquid YP medium was repeated 6 times.
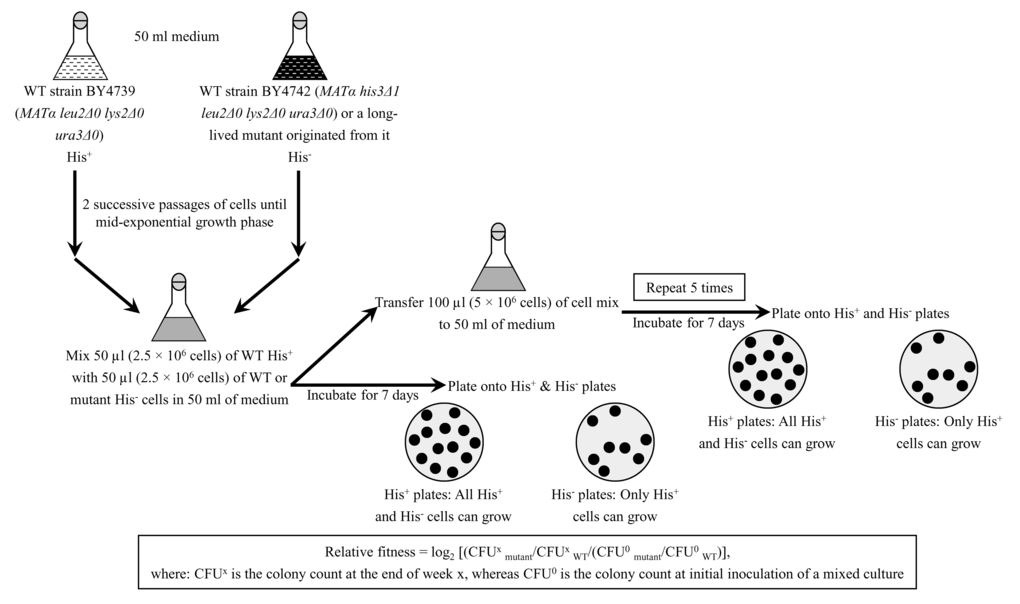
Figure 6. Quantifying the relative fitness of a long-lived mutant strain in a direct competition assay with a parental WT strain. His+ and His- strains used in the direct fitness competition experiment are first cultured separately in the complete YP medium rich in amino acids, nucleotides and other nutrients until mid-exponential phase. Cells of the His+ strain are then mixed with the same number of cells of the His- strain in liquid YP medium. After culturing the cell mixture for 7 days, an aliquot of cell suspension is diluted and plated on a solid YP medium. Following 2 days of incubation, colonies on each plate are replicated onto plates with the synthetic minimal YNB medium without amino acids and nucleotides. One of these plates contains leucine, lysine, uracil and histidine (it is called a His+ plate), whereas the other plate contains leucine, lysine and uracil (it is called a ″His-″ plate). After 2 days of incubation at 30oC, the number of CFU on ″His+″ and ″His-″ plates is counted. The relative fitness of each His+ strain in a direct competition with the His- is calculated as log2 [(CFUxmutant/CFUxWT/(CFU0mutant/CFU0WT)], where: CFUx is the colony count at the end of week x, whereas CFU0 is the colony count at initial inoculation of a mixed culture. The direct competition step of culturing a cell mixture for 7 days in liquid YP medium was repeated 6 times.
To validate this assay in a control experiment, we compared the fitness of the WT strain BY4742 (His-; the parental strain of the long-lived mutant strains 3, 5 and 12) to that of the WT strain BY4739 (His+, but otherwise isogenic to BY4742). We found that even after 6 consecutive 7-days incubations BY4742 (His-) exhibits similar relative fitness in a direct competition assay with BY4739 (His+) co-cultured in YP medium initially containing the following carbon source: 1) 0.2% glucose, after cell transfer from 0.2% glucose (Figure 7A); 2) 2% glucose, after cell transfer from 2% glucose (Figure 7B); 3) 1% ethanol, after cell transfer from 0.2% glucose (Figure 7C); 4) 1% ethanol, after cell transfer from 2% glucose (Figure 7D); or 5) 1% ethanol, after cell transfer from 1% ethanol (Figure 7E). Based on these findings, we concluded that the developed direct competition assay outlined in Figure 6 accurately reproduces the expected equal fitness of each of the two WT strains used, i.e. BY4739 (His+) and BY4742 (His-). Moreover, this assay also accurately reproduces the reduced fitness [73,89] of the mutant strain dbp3Δ (which is isogenic to the WT strain BY4742) in direct competition with the parental WT strain BY4739 (His+) (Figure 7).
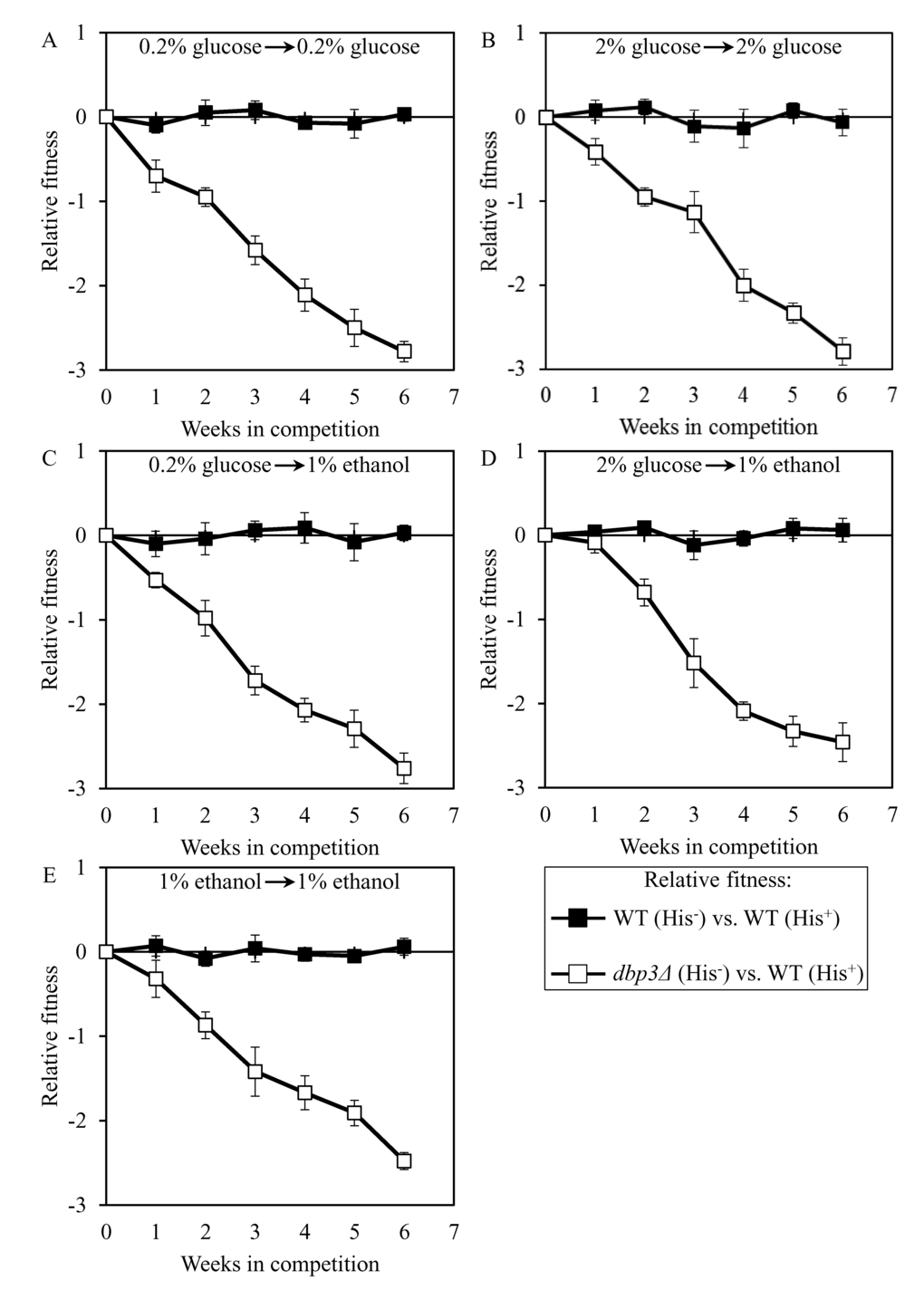
Figure 7. Validation of the developed assay for quantifying the relative fitness of a long-lived mutant strain in direct competition with a parental WT strain. The WT strains BY4742 (His-) and BY4739 (His+, but otherwise isogenic to BY4742) were cultured separately in the complete YP medium containing 0.2% glucose, 2% glucose or 1% ethanol glucose until mid-exponential phase. Another pair of strains whose relative fitness was measured, namely the long-lived mutant strain dbp3Δ (His-; is isogenic to BY4742) and the WT strain BY4739 (His+), was also cultured separately in YP medium containing 0.2% glucose 2% glucose or 1% ethanol glucose until mid-exponential phase. Cells of the His+ strain were mixed with the same number of cells of the His- strain and then co-cultured for 7 days in liquid YP medium initially containing different carbon sources. Cells of the His- and His+ strains pre-cultured separately on 0.2% glucose were subjected to direct fitness competition by being cultured together on 0.2% glucose (A) or 1% ethanol (C). Cells of the His- and His+ strains pre-cultured separately on 2% glucose were subjected to direct fitness competition by being cultured together on 2% glucose (B) or 1% ethanol (D). Cells of the His- and His+ strains pre-cultured separately on 1% ethanol were subjected to direct fitness competition by being cultured together on 1% ethanol (E). After culturing the cell mixture for 7 days, an aliquot of cell suspension was used to measure the relative fitness of the His+ strain in direct competition with the His- strain (as described in ″Materials and Methods″). The direct fitness competition step of culturing a cell mixture for 7 days in liquid YP medium was repeated 6 times.
Dominant polygenic trait extending longevity of each of the 3 long-lived yeast mutants decreases its relative fitness under some laboratory conditions
We used the developed direct competition assay to measure the relative fitness of the long-lived mutant strain 3, 5 or 12 in direct competition with a parental WT strain. Cells of each of these mutant strains were first cultured separately in liquid YP medium containing different concentrations of glucose or ethanol. Cells of each mutant strain were then mixed with the same number of cells of the WT strain BY4739 (His+, but otherwise isogenic to the parental WT strain BY4742) and underwent 6 consecutive 7-days incubations together. We found that the dominant polygenic trait extending longevity of the long-lived mutant strain 3, 5 or 12 does not alter its relative fitness in a direct competition assay with the parental WT strain co-cultured in medium initially containing one of the following carbon sources: 1) 0.2% glucose, after cell transfer from 0.2% glucose (Figures 8A, 9A and 10A, respectively); or 2) 2% glucose, after cell transfer from 2% glucose (Figures 8B, 9B and 10B, respectively). In contrast, the dominant polygenic trait extending longevity of the long-lived mutant strain 3, 5 or 12 decreased its relative fitness in a direct competition assay with the parental WT strain co-cultured in medium initially containing 1% ethanol, after cell transfer from any of the following carbon sources: 1) 0.2% glucose (Figures 8C, 9C and 10C, respectively); 2) 2% glucose (Figures 8D, 9D and 10D, respectively); or 3) 1% ethanol (Figures 8E, 9E and 10E, respectively).
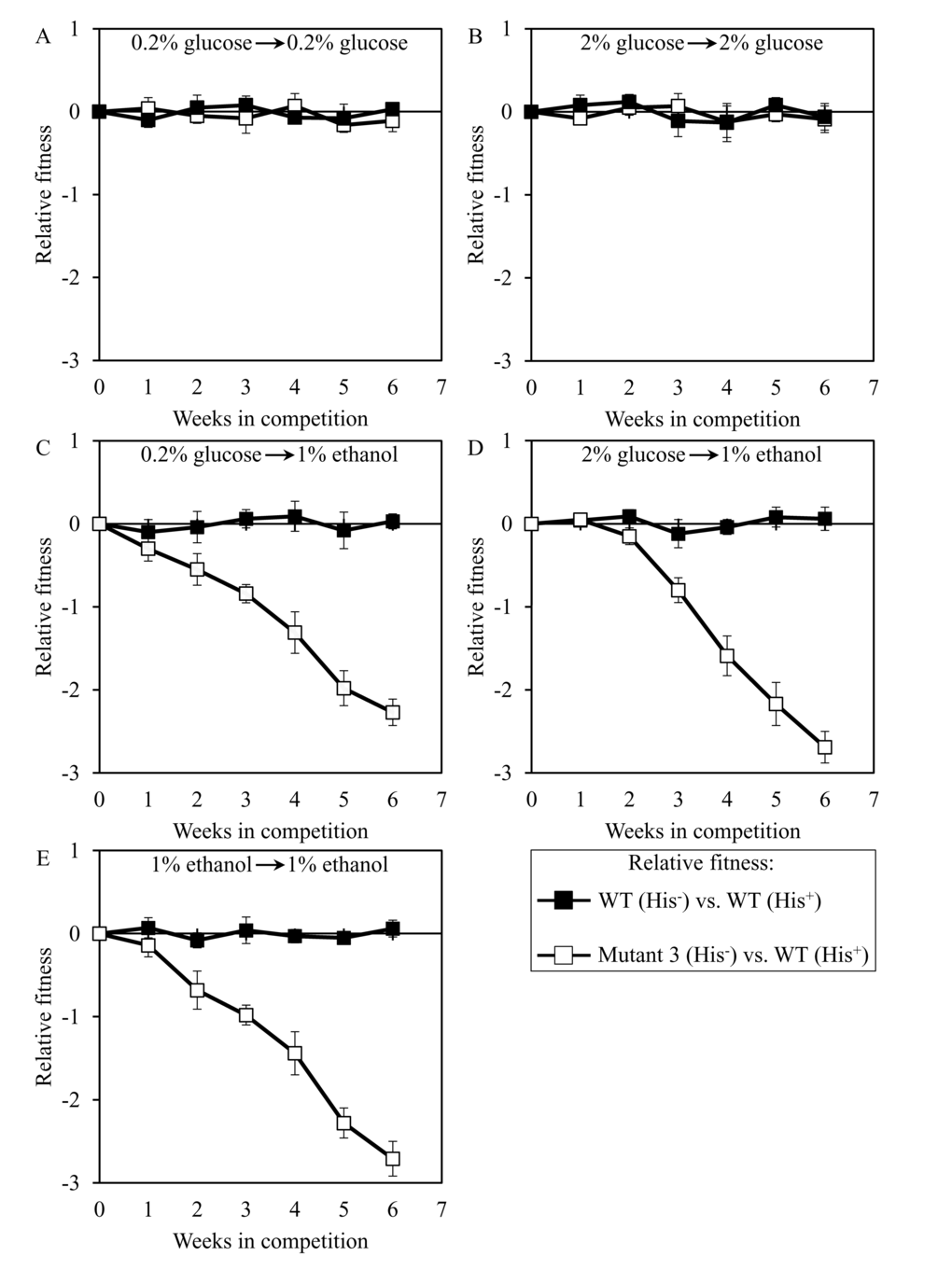
Figure 8. Dominant polygenic trait extending longevity of the long-lived yeast mutant 3 decreases its relative fitness in medium initially containing 1% ethanol. The WT strains BY4742 (His-) and BY4739 (His+, but otherwise isogenic to BY4742) were cultured separately in the complete YP medium containing 0.2% glucose, 2% glucose or 1% ethanol glucose until mid-exponential phase. Another pair of strains whose relative fitness was measured, namely the long-lived mutant strain 3 (His-; selected during lasting exposure of BY4742 to LCA) and the WT strain BY4739 (His+), was also cultured separately in YP medium containing 0.2% glucose 2% glucose or 1% ethanol glucose until mid-exponential phase. Cells of the His+ strain were mixed with the same number of cells of the His- strain and then co-cultured for 7 days in liquid YP medium initially containing different carbon sources. Cells of the His- and His+ strains pre-cultured separately on 0.2% glucose were subjected to direct fitness competition by being cultured together on 0.2% glucose (A) or 1% ethanol (C). Cells of the His- and His+ strains pre-cultured separately on 2% glucose were subjected to direct fitness competition by being cultured together on 2% glucose (B) or 1% ethanol (D). Cells of the His- and His+ strains pre-cultured separately on 1% ethanol were subjected to direct fitness competition by being cultured together on 1% ethanol (E). After culturing the cell mixture for 7 days, an aliquot of cell suspension was used to measure the relative fitness of the His+ strain in a direct competition with the His- strain (as described in ″Materials and Methods″). The direct fitness competition step of culturing a cell mixture for 7 days in liquid YP medium was repeated 6 times.
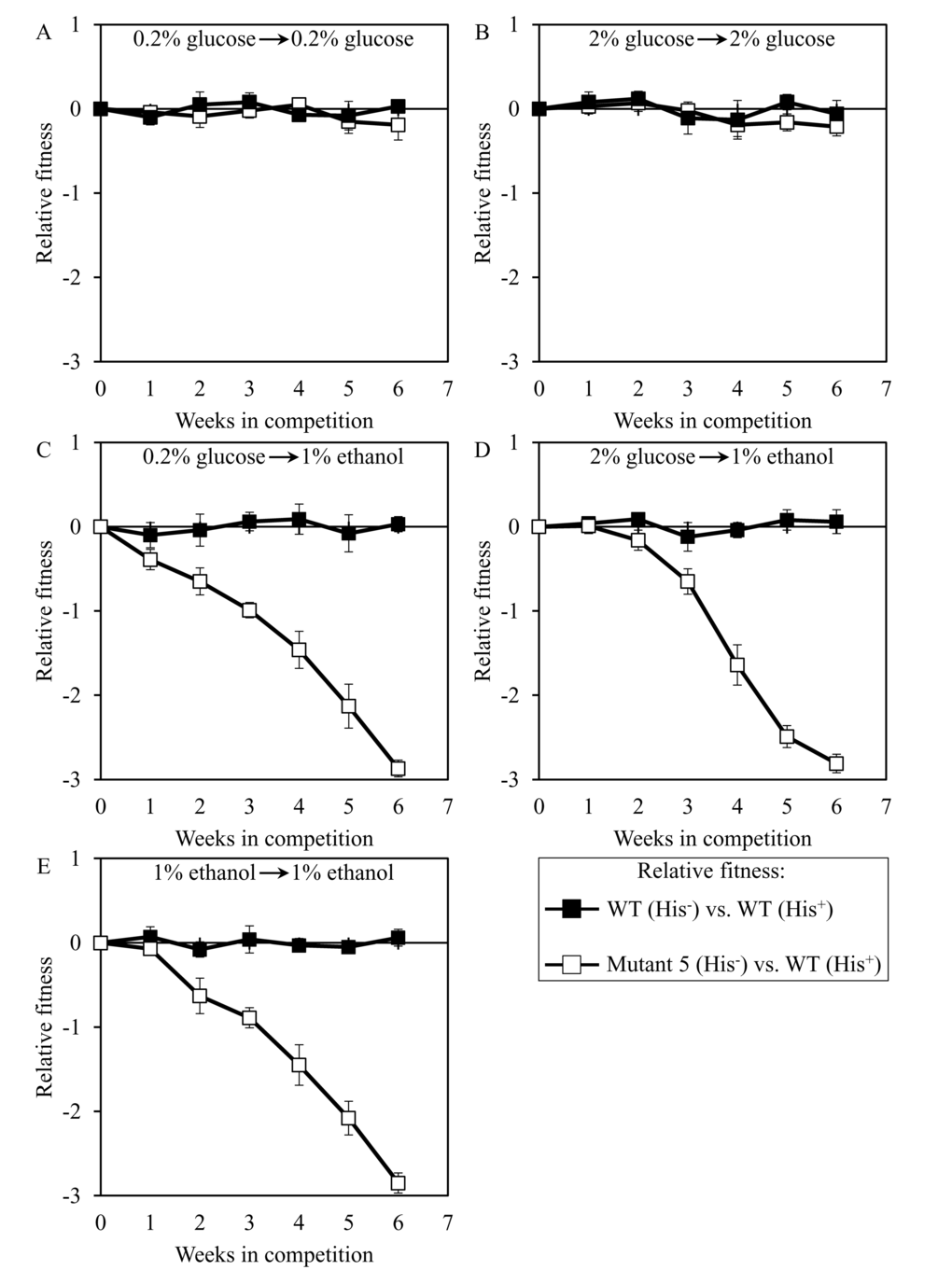
Figure 9. Dominant polygenic trait extending longevity of the long-lived yeast mutant 5 decreases its relative fitness in medium initially containing 1% ethanol. The WT strains BY4742 (His-) and BY4739 (His+, but otherwise isogenic to BY4742) were cultured separately in the complete YP medium containing 0.2% glucose, 2% glucose or 1% ethanol glucose until mid-exponential phase. Another pair of strains whose relative fitness was measured, namely the long-lived mutant strain 5 (His-; selected during lasting exposure of BY4742 to LCA) and the WT strain BY4739 (His+), was also cultured separately in YP medium containing 0.2% glucose 2% glucose or 1% ethanol glucose until mid-exponential phase. Cells of the His+ strain were mixed with the same number of cells of the His- strain and then co-cultured for 7 days in liquid YP medium initially containing different carbon sources. Cells of the His- and His+ strains pre-cultured separately on 0.2% glucose were subjected to direct fitness competition by being cultured together on 0.2% glucose (A) or 1% ethanol (C). Cells of the His- and His+ strains pre-cultured separately on 2% glucose were subjected to direct fitness competition by being cultured together on 2% glucose (B) or 1% ethanol (D). Cells of the His- and His+ strains pre-cultured separately on 1% ethanol were subjected to direct fitness competition by being cultured together on 1% ethanol (E). After culturing the cell mixture for 7 days, an aliquot of cell suspension was used to measure the relative fitness of the His+ strain in a direct competition with the His- strain (as described in ″Materials and Methods″). The direct fitness competition step of culturing a cell mixture for 7 days in liquid YP medium was repeated 6 times.
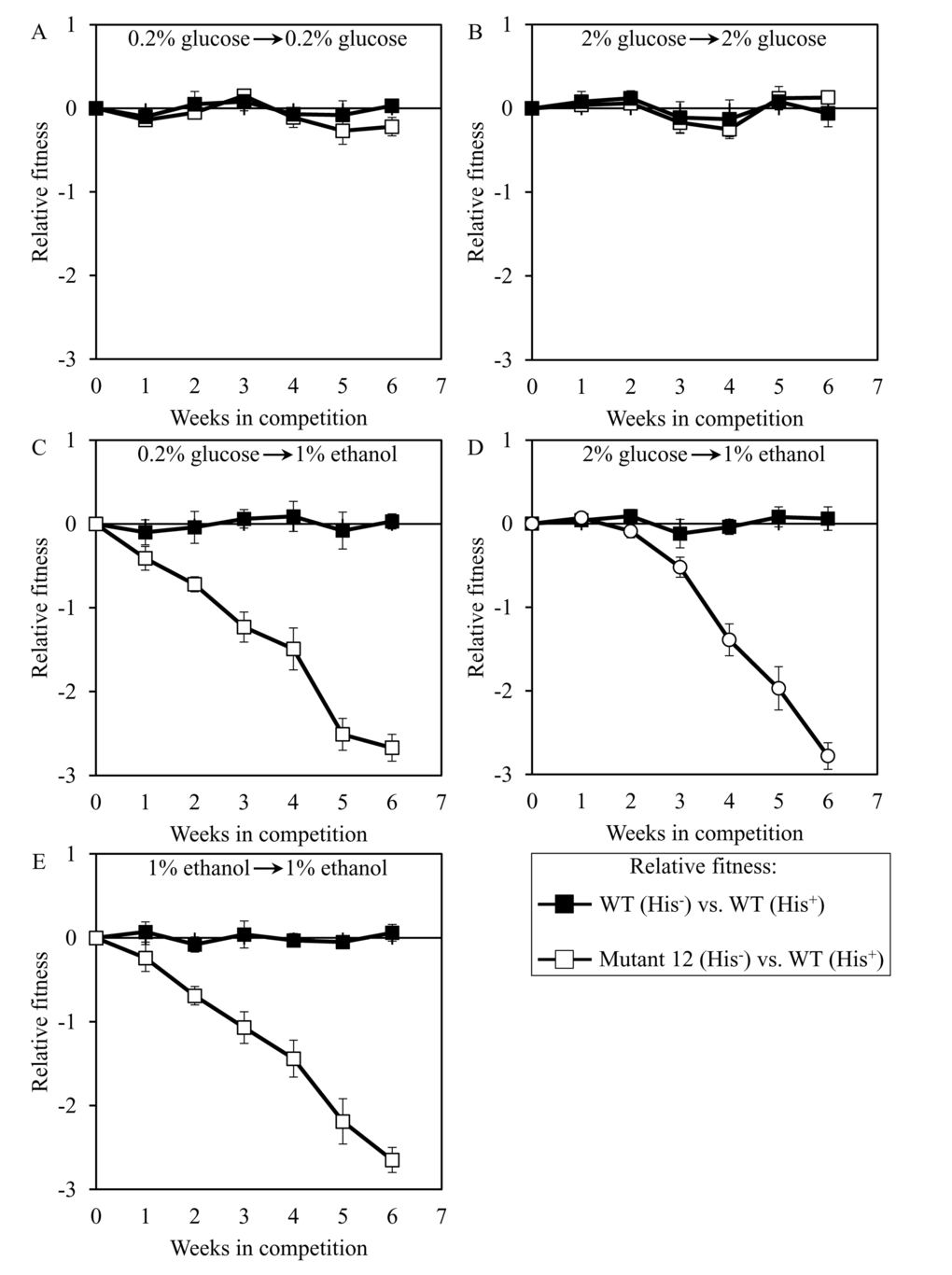
Figure 10. Dominant polygenic trait extending longevity of the long-lived yeast mutant 12 decreases its relative fitness in medium initially containing 1% ethanol. The WT strains BY4742 (His-) and BY4739 (His+, but otherwise isogenic to BY4742) were cultured separately in the complete YP medium containing 0.2% glucose, 2% glucose or 1% ethanol glucose until mid-exponential phase. Another pair of strains whose relative fitness was measured, namely the long-lived mutant strain 12 (His-; selected during lasting exposure of BY4742 to LCA) and the WT strain BY4739 (His+), was also cultured separately in YP medium containing 0.2% glucose 2% glucose or 1% ethanol glucose until mid-exponential phase. Cells of the His+ strain were mixed with the same number of cells of the His- strain and then co-cultured for 7 days in liquid YP medium initially containing different carbon sources. Cells of the His- and His+ strains pre-cultured separately on 0.2% glucose were subjected to direct fitness competition by being cultured together on 0.2% glucose (A) or 1% ethanol (C). Cells of the His- and His+ strains pre-cultured separately on 2% glucose were subjected to direct fitness competition by being cultured together on 2% glucose (B) or 1% ethanol (D). Cells of the His- and His+ strains pre-cultured separately on 1% ethanol were subjected to direct fitness competition by being cultured together on 1% ethanol (E). After culturing the cell mixture for 7 days, an aliquot of cell suspension was used to measure the relative fitness of the His+ strain in a direct competition with the His- strain (as described in ″Materials and Methods″). The direct fitness competition step of culturing a cell mixture for 7 days in liquid YP medium was repeated 6 times.
Our findings revealed that the conditions of pre-culturing of any of the 3 long-lived mutant strains do not influence the extent of its decreased relative fitness during the subsequent co-culturing with the parental WT strain in medium initially containing 1% ethanol (Figures 8C-8E, 9C-9E, 10C-10E). We therefore concluded that none of these long-lived mutant strains keeps a ″memory″ of conditions under which it has been grown prior to being mixed with the parental WT strain in medium supplemented with 1% ethanol for fitness competition.
Discussion
Using the 3 long-lived mutant strains selected during experimental evolution under laboratory conditions [89], in this study we empirically verified evolutionary theories programmed or non-programmed aging. We demonstrate that the dominant polygenic trait extending longevity of each of these mutants does not affect such key features of early-life fitness as the exponential growth rate, efficacy of post-exponential growth, and fecundity (which was assessed by measuring the efficacies of mating and sporulation). These findings provide evidence in support of evolutionary theories of programmed aging and invalidate evolutionary theories of non-programmed aging and age-related death. Indeed, all evolutionary theories of non-programmed aging and age-related death predict that any longevity-extending genetic trait must decrease early-life fitness of an organism if it grows and ages alone, in the absence of WT individuals of the same species; these WT individuals do not carry longevity-extending mutations and thus do not have lifespan extended beyond a species-specific age [2,4–6,15–17,82–88].
This study and our recent findings (Gomez-Perez et al., submitted) show for the first time that a longevity-extending genetic trait can enhance such features of early-life fitness as susceptibility to chronic exogenous stresses, and the resistance to apoptotic and liponecrotic forms of programmed cell death. We have observed this enhancement of some early-life fitness features in the 3 long-lived mutant strains when each of them was growing and undergoing chronological aging in the absence of the parental WT yeast strain (i.e. in the absence of natural selection).
In this study, we also developed and validated a direct competition assay for the measurement of relative fitness under laboratory conditions. This assay mimics the process of natural selection within a mixed population of yeast cells that 1) exhibit different longevity-defining genetic backgrounds; 2) differ in their lifespans if grow as a genetically homogenous cell population; and 3) compete for nutrients and other environmental resources. Using this assay, we found that in a population of mixed cells grown on 1% ethanol the dominant polygenic trait extending longevity of each of the 3 long-lived yeast mutants decreases the relative fitness of the mutant strain in direct competition with the parental WT strain BY4742. These findings imply that under laboratory conditions that imitate the process of natural selection within an ecosystem composed of yeast cells having different longevity-defining genetic backgrounds, each of the 3 long-lived mutants is forced out of the ecosystem by the parental WT strain exhibiting shorter lifespan. It seems conceivable therefore that 1) yeast cells have evolved some mechanisms for limiting their lifespan upon reaching a certain chronological age; and 2) these mechanisms drive the evolution of yeast longevity towards maintaining a finite yeast lifespan within ecosystems. We speculate that these mechanisms may consist in the ability of the parental WT strain to secrete into growth medium certain compounds (small molecules and/or proteins) that can slow down growth and/or kill long-lived yeast mutants. The challenge for the near future is to identify these compounds responsible for the maintenance a finite yeast lifespan within ecosystems.
Materials and Methods
Yeast strains and growth conditions
The haploid WT strains BY4741 (MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0) and BY4742 (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0) of the yeast S. cerevisiae, the long-lived mutant strains 3, 5 and 12 (each in the BY4742 genetic background), as well as the single-gene-deletion mutant strains rpp2BΔ (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0 rpp2BΔ::kanMX4) and dbp3Δ (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0 dbp3Δ::kanMX4) (each in the BY4742 genetic background) were used in this study. All strains were from Open Biosystems. Cells were grown in YP medium (1% yeast extract, 2% peptone) initially containing 0.2% glucose (a fermentable carbon source; CR conditions), 2% glucose (a fermentable carbon source; non-CR conditions), 1% ethanol (a non-fermentable carbon source) or 3% glycerol (a non-fermentable carbon source). Cells were cultured at 30oC with rotational shaking at 200 rpm in Erlenmeyer flasks at a ″flask volume/medium volume″ ratio of 5:1.
Quantitative mating assay
Cultures of mating type MATa (i.e. the haploid WT strain BY4741 [MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0]) and mating type MATα (i.e. the haploid WT strain BY4742 [MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0] or the selected long-lived haploid mutant strains 3, 5 or 12) were grown separately to mid-logarithmic phase in YP medium (1% yeast extract, 2% peptone) initially containing 0.2% glucose (a fermentable carbon source; CR conditions) or 1% ethanol (a non-fermentable carbon source). Equal numbers (5 × 106) of cells of each mating type were mixed and then collected on a 0.45-µm pore, 25-mm diameter nitrocellulose (NC) filter. The filters were placed on the surface of a YEPD (1% yeast extract, 2% peptone, 2% glucose, 2% agar) plate and incubated at 30oC for 5 hours. The filters were then transferred to Eppendorf tubes and resuspended in 1 ml of a liquid synthetic minimal YNB medium (0.67% Yeast Nitrogen Base without Amino Acids) with 2% glucose. The suspensions were used for making serial 10-fold dilutions. 100-μl aliquots of each dilution were spread on 1) a synthetic minimal YNB medium plate (0.67% Yeast Nitrogen Base without Amino Acids, 2% glucose, 2% agar) without supplements; and 2) a synthetic minimal YNB medium plate supplemented with 20 mg/l L-histidine, 30 mg/l L-leucine and 20 mg/l uracil. These plates were incubated at 30oC for 2 days. The numbers of diploid cells (Nd) were counted on synthetic minimal YNB medium plates without supplements, whereas the total numbers of cells (Nt) were counted on synthetic minimal YNB medium plates supplemented with 20 mg/l L-histidine, 30 mg/l L-leucine and 20 mg/l uracil. The efficiency of mating was calculated as the number of colonies of MATa/MATα diploids (Nd) divided by the sum of MATa/MATα diploids plus haploid colonies (Nt). Crosses between two WT strains of opposite mating types (i.e. the haploid strain BY4741 [MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0]) and the haploid strain BY4742 [MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0]) were used as controls. All tests were carried out in triplicate in 3 independent experiments.
Quantitative sporulation assay
A small patch of cells of the haploid WT strain BY4741 (MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0) was applied to the surface of a master YEPD (1% yeast extract, 2% peptone, 2% glucose, 2% agar) plate. 106 cells of mating type MATα (i.e. the haploid WT strain BY4742 [MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0] or the selected long-lived haploid mutant strains 3, 5 or 12) were spread on the surface of a separate crossing plate with YEPD medium. The master plate was replica plated onto a lawn of cells on each of the four crossing plates; different velvet was used for each crossing plate. The crossing plates were incubated overnight at 30oC. Each of the four crossing plates was then replica plated onto a synthetic minimal YNB medium plate (0.67% Yeast Nitrogen Base without Amino Acids, 2% glucose, 2% agar) supplemented with 20 mg/l L-histidine, 30 mg/l L-leucine and 20 mg/l uracil. These plates were incubated overnight at 30oC. A positive mating reaction between cells of the haploid WT strain BY4741 (MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0) and cells of the haploid WT strain BY4742 (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0) or cells of each of the selected long-lived haploid mutant strains 3, 5 or 12 resulted in confluent growth of diploid cells on a YNB plate (supplemented with L-histidine, L-leucine and uracil) at the position of a patch of haploid BY4741 cells. To measure sporulation efficiency, cells of each of the four recovered diploid strains were first grown to mid-logarithmic phase in YP medium (1% yeast extract, 2% peptone) initially containing 0.2% glucose (a fermentable carbon source; CR conditions) or 1% ethanol (a non-fermentable carbon source). The cell cycle of these cells was then synchronized by growing them in YPA medium (1% yeast extract, 2% peptone, 2% potassium acetate) from a starting optical density at 600 nm (OD600) of 0.2 to final OD600 of 1.0; cells were cultured at 30oC with rotational shaking at 200 rpm in Erlenmeyer flasks at a ″flask volume/medium volume″ ratio of 10:1. 2 × 107 of cells from this synchronized culture were then incubated in liquid SPO (0.1% yeast extract, 1% potassium acetate, 0.05% glucose) medium supplemented with 20 mg/l L-histidine, 30 mg/l L-leucine and 20 mg/l uracil at 30oC for the duration of experiment. At various time points, aliquots of cells were examined for sporulation efficiency by differential interference contrast (DIC) microscopy with an Olympus BX microscope with a ☓ 100 oil immersion objective. Sporulation efficiency was measured as the percentage of tetrads and dyads produced by a strain, relative to the total number of cells. All tests were carried out in triplicate in 3 independent experiments.
Cell viability assay for monitoring the susceptibility of yeast to an apoptotic mode of cell death induced by hydrogen peroxide
A sample of cells was taken from a culture at a certain time-point. A fraction of the sample was diluted in order to determine the total number of cells using a hemacytometer. 2 × 107 cells were harvested by centrifugation for 1 min at 21,000 × g at room temperature and resuspended in 2 ml of YP medium containing 0.2% glucose as carbon source. Each cell suspension was divided into 2 equal aliquots. One aliquot was supplemented with hydrogen peroxide to the final concentration of 2.5 mM, whereas other aliquot remained untreated. Both aliquots were then incubated for 2 h at 30oC on a Labquake rotator set for 360o rotation. Serial dilutions of cells were plated in duplicate onto plates containing YP medium with 2% glucose as carbon source. After 2 d of incubation at 30oC, the number of colony forming units (CFU) per plate was counted. The number of CFU was defined as the number of viable cells in a sample. For each aliquot of cells exposed to hydrogen peroxide, the % of viable cells was calculated as follows: (number of viable cells per ml in the aliquot exposed to hydrogen peroxide/number of viable cells per ml in the control aliquot that was not exposed to hydrogen peroxide) × 100.
Cell viability assay for monitoring the susceptibility of yeast to a liponecrotic mode of cell death induced by palmitoleic acid
A sample of cells was taken from a culture at a certain time-point. A fraction of the sample was diluted in order to determine the total number of cells using a hemacytometer. 2 × 107 cells were harvested by centrifugation for 1 min at 21,000 × g at room temperature and resuspended in 2 ml of YP medium containing 0.2% glucose as carbon source. Each cell suspension was divided into 2 equal aliquots. One aliquot was supplemented with palmitoleic acid (#P9417; Sigma) from a 50 mM stock solution (in 10% chloroform, 45% hexane and 45% ethanol); the final concentration of palmitoleic acid was 0.15 mM (in 0.03% chloroform, 0.135% hexane and 0.135% ethanol). Other aliquot was supplemented with chloroform, hexane and ethanol added to the final concentrations of 0.03%, 0.135% and 0.135%, respectively. Both aliquots were then incubated for 2 h at 30oC on a Labquake rotator set for 360o rotation. Serial dilutions of cells were plated in duplicate onto plates containing YP medium with 2% glucose as carbon source. After 2 d of incubation at 30oC, the number of CFU per plate was counted. The number of CFU was defined as the number of viable cells in a sample. For each aliquot of cells exposed to palmitoleic acid, the % of viable cells was calculated as follows: (number of viable cells per ml in the aliquot exposed to palmitoleic acid/number of viable cells per ml in the control aliquot that was not exposed to palmitoleic acid) × 100.
Quantifying the relative fitness of a long-lived mutant strain in a direct competition assay with a parental WT strain
The WT strains BY4739 (MATα leu2Δ0 lys2Δ0 ura3Δ0) and BY4742 (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0), the single-gene-deletion mutant strain dbp3Δ (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0 dbp3Δ::kanMX4) in the BY4742 genetic background (all from Open Biosystems), and the long-lived mutant strains 3, 5 and 12 (all 3 in the BY4742 genetic background) were grown separately in YP medium (1% yeast extract, 2% peptone) initially containing 0.2% glucose, 2% glucose or 1% ethanol as carbon source until mid-exponential phase. Cells were cultured at 30oC with rotational shaking at 200 rpm in Erlenmeyer flasks at a ″flask volume/medium volume″ ratio of 5:1. The single-gene-deletion mutant strain dbp3Δ lacks a gene encoding a DEAD-box family protein involved in ribosomal biogenesis [73]. dbp3Δ was used as a control mutant strain because it is known to exhibit 1) an extended replicative lifespan (as compared to the parental WT strain BY4742) [73]; 2) a reduced growth rate on 0.2% glucose (this study), 2% glucose [73] and 1% ethanol (this study); and 3) a reduced relative fitness when is co-cultured with a parental WT strain in medium initially containing 2% glucose [73]. 2.5 × 106 cells of the WT strain BY4739 (MATα leu2Δ0 lys2Δ0 ura3Δ0) were mixed with the same number of cells of the BY4742 (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0), dbp3Δ (MATα his3Δ1 leu2Δ0 lys2Δ0 ura3Δ0 dbp3Δ::kanMX4), 3, 5 or 12 strain in 50 ml of YP medium initially containing 0.2% glucose, 2% glucose or 1% ethanol as carbon source. After culturing the cell mixture at 30oC for 7 days, an aliquot of cell suspension was diluted and plated on a solid YP medium supplemented with 2% glucose. Following 2 days of incubation at 30oC, colonies on each plate were replicated onto 2 plates with solid YNB (Yeast Nitrogen Base) medium without amino acids supplemented with 2% glucose; one of these plates contained leucine, lysine, uracil and histidine [hereafter called a ″His+″ plate], whereas the other plate contained leucine, lysine and uracil [hereafter called a ″His-″ plate]. After 2 days of incubation at 30oC, the number of CFU on ″His+″ and ″His-″ plates was counted. The relative fitness of each His+ strain (relative to the His- strain BY4739 [MATα leu2Δ0 lys2Δ0 ura3Δ0]) was calculated as log2 [(CFUxmutant/CFUxWT/(CFU0mutant/CFU0WT)], where: CFUx is the colony count at the end of week x, whereas CFU0 is the colony count at initial inoculation of a mixed culture.
Statistical analysis
Statistical analysis was performed using Microsoft Excel’s (2010) Analysis ToolPack-VBA. All data on cell survival are presented as mean ± SEM. The p values for comparing the means of two groups (using an unpaired two-tailed t test) were calculated with the help of the GraphPad Prism statistics software.
Acknowledgements
We are grateful to current and former members of the Titorenko laboratory for discussions. We acknowledge the Centre for Structural and Functional Genomics at Concordia University for outstanding service.
Conflicts of Interest
The authors have no conflict of interests to declare.
Funding
This study was supported by grants from the Natural Sciences and Engineering Research Council (NSERC) of Canada and Concordia University Chair Fund. P.K. was supported by the Fonds québécois de la recherche sur la nature et les technologies Doctoral Research Fellowship Award. V.I.T. is a Concordia University Research Chair in Genomics, Cell Biology and Aging.
References
- 1. Mitteldorf J. Aging is not a process of wear and tear. Rejuvenation Res. 2010; 13:322–26. https://doi.org/10.1089/rej.2009.0967 [PubMed]
- 2. Trindade LS, Aigaki T, Peixoto AA, Balduino A, Mânica da Cruz IB, Heddle JG. A novel classification system for evolutionary aging theories. Front Genet. 2013; 4:25. https://doi.org/10.3389/fgene.2013.00025 [PubMed]
- 3. Kirkwood TB. Comparative life spans of species: why do species have the life spans they do? Am J Clin Nutr. 1992 (Suppl ); 55:1191S–95S. [PubMed]
- 4. Ljubuncic P, Reznick AZ. The evolutionary theories of aging revisited--a mini-review. Gerontology. 2009; 55:205–16. https://doi.org/10.1159/000200772 [PubMed]
- 5. Goldsmith T. Aging by Design. Azinet Press, Annapolis, 2011; ISBN 0-9788709-3-X.
- 6. Kowald A, Kirkwood TB. Can aging be programmed? A critical literature review. Aging Cell. 2016; Epub ahead of print. https://doi.org/10.1111/acel.12510 [PubMed]
- 7. Skulachev VP. Phenoptosis: programmed death of an organism. Biochemistry (Mosc). 1999; 64:1418–26. [PubMed]
- 8. Skulachev VP. Mitochondrial physiology and pathology; concepts of programmed death of organelles, cells and organisms. Mol Aspects Med. 1999; 20:139–84. https://doi.org/10.1016/S0098-2997(99)00008-4 [PubMed]
- 9. Skulachev VP. The programmed death phenomena, aging, and the Samurai law of biology. Exp Gerontol. 2001; 36:995–1024. https://doi.org/10.1016/S0531-5565(01)00109-7 [PubMed]
- 10. Skulachev VP. Programmed death phenomena: from organelle to organism. Ann N Y Acad Sci. 2002; 959:214–37. https://doi.org/10.1111/j.1749-6632.2002.tb02095.x [PubMed]
- 11. Skulachev VP. Programmed death in yeast as adaptation? FEBS Lett. 2002; 528:23–26. https://doi.org/10.1016/S0014-5793(02)03319-7 [PubMed]
- 12. Longo VD, Mitteldorf J, Skulachev VP. Programmed and altruistic ageing. Nat Rev Genet. 2005; 6:866–72. https://doi.org/10.1038/nrg1706 [PubMed]
- 13. Skulachev VP, Longo VD. Aging as a mitochondria-mediated atavistic program: can aging be switched off? Ann N Y Acad Sci. 2005; 1057:145–64. https://doi.org/10.1196/annals.1356.009 [PubMed]
- 14. Severin FF, Meer MV, Smirnova EA, Knorre DA, Skulachev VP. Natural causes of programmed death of yeast Saccharomyces cerevisiae. Biochim Biophys Acta. 2008; 1783:1350–53. https://doi.org/10.1016/j.bbamcr.2008.02.001 [PubMed]
- 15. Goldsmith TC. On the programmed/non-programmed aging controversy. Biochemistry (Mosc). 2012; 77:729–32. https://doi.org/10.1134/S000629791207005X [PubMed]
- 16. Mitteldorf JJ. Adaptive aging in the context of evolutionary theory. Biochemistry (Mosc). 2012; 77:716–25. https://doi.org/10.1134/S0006297912070036 [PubMed]
- 17. Goldsmith T. The Evolution of Aging 3 ed. Azinet Press, Annapolis, 2013; ISBN: 0978870905.
- 18. Weismann A. Über die Dauer des Lebens. Fisher, Jena, 1882.
- 19. Libertini G. An adaptive theory of increasing mortality with increasing chronological age in populations in the wild. J Theor Biol. 1988; 132:145–62. https://doi.org/10.1016/S0022-5193(88)80153-X [PubMed]
- 20. Goldsmith TC. Aging, evolvability, and the individual benefit requirement; medical implications of aging theory controversies. J Theor Biol. 2008; 252:764–68. https://doi.org/10.1016/j.jtbi.2008.02.035 [PubMed]
- 21. Skulachev VP. Aging is a specific biological function rather than the result of a disorder in complex living systems: biochemical evidence in support of Weismann’s hypothesis. Biochemistry (Mosc). 1997; 62:1191–95. [PubMed]
- 22. Fabrizio P, Battistella L, Vardavas R, Gattazzo C, Liou LL, Diaspro A, Dossen JW, Gralla EB, Longo VD. Superoxide is a mediator of an altruistic aging program in Saccharomyces cerevisiae.. J Cell Biol. 2004; 166:1055–67. https://doi.org/10.1083/jcb.200404002 [PubMed]
- 23. Herker E, Jungwirth H, Lehmann KA, Maldener C, Fröhlich KU, Wissing S, Büttner S, Fehr M, Sigrist S, Madeo F. Chronological aging leads to apoptosis in yeast. J Cell Biol. 2004; 164:501–07. https://doi.org/10.1083/jcb.200310014 [PubMed]
- 24. Váchová L, Palková Z. Physiological regulation of yeast cell death in multicellular colonies is triggered by ammonia. J Cell Biol. 2005; 169:711–17. https://doi.org/10.1083/jcb.200410064 [PubMed]
- 25. Büttner S, Eisenberg T, Herker E, Carmona-Gutierrez D, Kroemer G, Madeo F. Why yeast cells can undergo apoptosis: death in times of peace, love, and war. J Cell Biol. 2006; 175:521–25. https://doi.org/10.1083/jcb.200608098 [PubMed]
- 26. Howitz KT, Bitterman KJ, Cohen HY, Lamming DW, Lavu S, Wood JG, Zipkin RE, Chung P, Kisielewski A, Zhang LL, Scherer B, Sinclair DA. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature. 2003; 425:191–96. https://doi.org/10.1038/nature01960 [PubMed]
- 27. Baur JA, Sinclair DA. Therapeutic potential of resveratrol: the in vivo evidence. Nat Rev Drug Discov. 2006; 5:493–506. https://doi.org/10.1038/nrd2060 [PubMed]
- 28. Powers RW
3rd , Kaeberlein M, Caldwell SD, Kennedy BK, Fields S. Extension of chronological life span in yeast by decreased TOR pathway signaling. Genes Dev. 2006; 20:174–84. https://doi.org/10.1101/gad.1381406 [PubMed] - 29. Dasgupta B, Milbrandt J. Resveratrol stimulates AMP kinase activity in neurons. Proc Natl Acad Sci USA. 2007; 104:7217–22. https://doi.org/10.1073/pnas.0610068104 [PubMed]
- 30. Giorgio M, Trinei M, Migliaccio E, Pelicci PG. Hydrogen peroxide: a metabolic by-product or a common mediator of ageing signals? Nat Rev Mol Cell Biol. 2007; 8:722–28. https://doi.org/10.1038/nrm2240 [PubMed]
- 31. Greer EL, Brunet A. Signaling networks in aging. J Cell Sci. 2008; 121:407–12. https://doi.org/10.1242/jcs.021519 [PubMed]
- 32. Mair W, Dillin A. Aging and survival: the genetics of life span extension by dietary restriction. Annu Rev Biochem. 2008; 77:727–54. https://doi.org/10.1146/annurev.biochem.77.061206.171059 [PubMed]
- 33. Wanke V, Cameroni E, Uotila A, Piccolis M, Urban J, Loewith R, De Virgilio C. Caffeine extends yeast lifespan by targeting TORC1. Mol Microbiol. 2008; 69:277–85. https://doi.org/10.1111/j.1365-2958.2008.06292.x [PubMed]
- 34. Armour SM, Baur JA, Hsieh SN, Land-Bracha A, Thomas SM, Sinclair DA. Inhibition of mammalian S6 kinase by resveratrol suppresses autophagy. Aging (Albany NY). 2009; 1:515–28. https://doi.org/10.18632/aging.100056 [PubMed]
- 35. Blagosklonny MV. Inhibition of S6K by resveratrol: in search of the purpose. Aging (Albany NY). 2009; 1:511–14. https://doi.org/10.18632/aging.100059 [PubMed]
- 36. Colman RJ, Anderson RM, Johnson SC, Kastman EK, Kosmatka KJ, Beasley TM, Allison DB, Cruzen C, Simmons HA, Kemnitz JW, Weindruch R. Caloric restriction delays disease onset and mortality in rhesus monkeys. Science. 2009; 325:201–04. https://doi.org/10.1126/science.1173635 [PubMed]
- 37. Demidenko ZN, Blagosklonny MV. At concentrations that inhibit mTOR, resveratrol suppresses cellular senescence. Cell Cycle. 2009; 8:1901–04. https://doi.org/10.4161/cc.8.12.8810 [PubMed]
- 38. Eisenberg T, Knauer H, Schauer A, Büttner S, Ruckenstuhl C, Carmona-Gutierrez D, Ring J, Schroeder S, Magnes C, Antonacci L, Fussi H, Deszcz L, Hartl R, et al. Induction of autophagy by spermidine promotes longevity. Nat Cell Biol. 2009; 11:1305–14. https://doi.org/10.1038/ncb1975 [PubMed]
- 39. Finley LW, Haigis MC. The coordination of nuclear and mitochondrial communication during aging and calorie restriction. Ageing Res Rev. 2009; 8:173–88. https://doi.org/10.1016/j.arr.2009.03.003 [PubMed]
- 40. Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandez E, Miller RA. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009; 460:392–95. [PubMed]
- 41. Laplante M, Sabatini DM. mTOR signaling at a glance. J Cell Sci. 2009; 122:3589–94. https://doi.org/10.1242/jcs.051011 [PubMed]
- 42. Narasimhan SD, Yen K, Tissenbaum HA. Converging pathways in lifespan regulation. Curr Biol. 2009; 19:R657–66. https://doi.org/10.1016/j.cub.2009.06.013 [PubMed]
- 43. Skulachev VP, Anisimov VN, Antonenko YN, Bakeeva LE, Chernyak BV, Erichev VP, Filenko OF, Kalinina NI, Kapelko VI, Kolosova NG, Kopnin BP, Korshunova GA, Lichinitser MR, et al. An attempt to prevent senescence: a mitochondrial approach. Biochim Biophys Acta. 2009; 1787:437–61. https://doi.org/10.1016/j.bbabio.2008.12.008 [PubMed]
- 44. Anderson RM, Weindruch R. Metabolic reprogramming, caloric restriction and aging. Trends Endocrinol Metab. 2010; 21:134–41. https://doi.org/10.1016/j.tem.2009.11.005 [PubMed]
- 45. Bjedov I, Toivonen JM, Kerr F, Slack C, Jacobson J, Foley A, Partridge L. Mechanisms of life span extension by rapamycin in the fruit fly Drosophila melanogaster. Cell Metab. 2010; 11:35–46. https://doi.org/10.1016/j.cmet.2009.11.010 [PubMed]
- 46. Blagosklonny MV. Rapamycin and quasi-programmed aging: four years later. Cell Cycle. 2010; 9:1859–62. https://doi.org/10.4161/cc.9.10.11872 [PubMed]
- 47. Fontana L, Partridge L, Longo VD. Extending healthy life span--from yeast to humans. Science. 2010; 328:321–26. https://doi.org/10.1126/science.1172539 [PubMed]
- 48. Goldberg AA, Richard VR, Kyryakov P, Bourque SD, Beach A, Burstein MT, Glebov A, Koupaki O, Boukh-Viner T, Gregg C, Juneau M, English AM, Thomas DY, Titorenko VI. Chemical genetic screen identifies lithocholic acid as an anti-aging compound that extends yeast chronological life span in a TOR-independent manner, by modulating housekeeping longevity assurance processes. Aging (Albany NY). 2010; 2:393–414. https://doi.org/10.18632/aging.100168 [PubMed]
- 49. Kaeberlein M. Lessons on longevity from budding yeast. Nature. 2010; 464:513–19. https://doi.org/10.1038/nature08981 [PubMed]
- 50. Kenyon CJ. The genetics of ageing. Nature. 2010; 464:504–12. https://doi.org/10.1038/nature08980 [PubMed]
- 51. Anisimov VN, Berstein LM, Popovich IG, Zabezhinski MA, Egormin PA, Piskunova TS, Semenchenko AV, Tyndyk ML, Yurova MN, Kovalenko IG, Poroshina TE. If started early in life, metformin treatment increases life span and postpones tumors in female SHR mice. Aging (Albany NY). 2011; 3:148–57. https://doi.org/10.18632/aging.100273 [PubMed]
- 52. Titorenko VI, Terlecky SR. Peroxisome metabolism and cellular aging. Traffic. 2011; 12:252–59. https://doi.org/10.1111/j.1600-0854.2010.01144.x [PubMed]
- 53. Longo VD, Shadel GS, Kaeberlein M, Kennedy B. Replicative and chronological aging in Saccharomyces cerevisiae. Cell Metab. 2012; 16:18–31. https://doi.org/10.1016/j.cmet.2012.06.002 [PubMed]
- 54. Smoliga JM, Vang O, Baur JA. Challenges of translating basic research into therapeutics: resveratrol as an example. J Gerontol A Biol Sci Med Sci. 2012; 67:158–67. https://doi.org/10.1093/gerona/glr062 [PubMed]
- 55. Gems D, Partridge L. Genetics of longevity in model organisms: debates and paradigm shifts. Annu Rev Physiol. 2013; 75:621–44. https://doi.org/10.1146/annurev-physiol-030212-183712 [PubMed]
- 56. Leonov A, Titorenko VI. A network of interorganellar communications underlies cellular aging. IUBMB Life. 2013; 65:665–74. https://doi.org/10.1002/iub.1183 [PubMed]
- 57. Arlia-Ciommo A, Leonov A, Piano A, Svistkova V, Titorenko VI. Cell-autonomous mechanisms of chronological aging in the yeast Saccharomyces cerevisiae. Microb Cell. 2014; 1:164–78. https://doi.org/10.15698/mic2014.06.152
- 58. Leonov A, Arlia-Ciommo A, Piano A, Svistkova V, Lutchman V, Medkour Y, Titorenko VI. Longevity extension by phytochemicals. Molecules. 2015; 20:6544–72. https://doi.org/10.3390/molecules20046544 [PubMed]
- 59. Lutchman V, Dakik P, McAuley M, Cortes B, Ferraye G, Gontmacher L, Graziano D, Moukhariq FZ, Simard É, Titorenko VI. Six plant extracts delay yeast chronological aging through different signaling pathways. Oncotarget. 2016; •••:7; Epub ahead of print. [PubMed]
- 60. Kenyon C, Chang J, Gensch E, Rudner A, Tabtiang R. A C. elegans mutant that lives twice as long as wild type. Nature. 1993; 366:461–64. https://doi.org/10.1038/366461a0 [PubMed]
- 61. Parsons PA. Inherited stress resistance and longevity: a stress theory of ageing. Heredity (Edinb). 1995; 75:216–21. https://doi.org/10.1038/hdy.1995.126 [PubMed]
- 62. Zwaan BJ, Bijlsma R, Hoekstra RF. Direct selection on lifespan in Drosophila melanogaster. Evolution. 1995; 49:649–59. https://doi.org/10.2307/2410318
- 63. Brown-Borg HM, Borg KE, Meliska CJ, Bartke A. Dwarf mice and the ageing process. Nature. 1996; 384:33. https://doi.org/10.1038/384033a0 [PubMed]
- 64. Partridge L, Prowse N, Pignatelli P. Another set of responses and correlated responses to selection on age at reproduction in Drosophila melanogaster. Proc Biol Sci. 1999; 266:255–61. https://doi.org/10.1098/rspb.1999.0630 [PubMed]
- 65. Buck S, Vettraino J, Force AG, Arking R. Extended longevity in Drosophila is consistently associated with a decrease in developmental viability. J Gerontol A Biol Sci Med Sci. 2000; 55:B292–301. https://doi.org/10.1093/gerona/55.6.B292 [PubMed]
- 66. Stearns SC, Ackermann M, Doebeli M, Kaiser M. Experimental evolution of aging, growth, and reproduction in fruitflies. Proc Natl Acad Sci USA. 2000; 97:3309–13. https://doi.org/10.1073/pnas.97.7.3309 [PubMed]
- 67. Clancy DJ, Gems D, Harshman LG, Oldham S, Stocker H, Hafen E, Leevers SJ, Partridge L. Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein. Science. 2001; 292:104–06. https://doi.org/10.1126/science.1057991 [PubMed]
- 68. Tatar M, Kopelman A, Epstein D, Tu MP, Yin CM, Garofalo RS. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science. 2001; 292:107–10. https://doi.org/10.1126/science.1057987 [PubMed]
- 69. Marden JH, Rogina B, Montooth KL, Helfand SL. Conditional tradeoffs between aging and organismal performance of Indy long-lived mutant flies. Proc Natl Acad Sci USA. 2003; 100:3369–73. https://doi.org/10.1073/pnas.0634985100 [PubMed]
- 70. Liu X, Jiang N, Hughes B, Bigras E, Shoubridge E, Hekimi S. Evolutionary conservation of the clk-1-dependent mechanism of longevity: loss of mclk1 increases cellular fitness and lifespan in mice. Genes Dev. 2005; 19:2424–34. https://doi.org/10.1101/gad.1352905 [PubMed]
- 71. Partridge L, Gems D. Beyond the evolutionary theory of ageing, from functional genomics to evo-gero. Trends Ecol Evol. 2006; 21:334–40. https://doi.org/10.1016/j.tree.2006.02.008 [PubMed]
- 72. Giannakou ME, Goss M, Jacobson J, Vinti G, Leevers SJ, Partridge L. Dynamics of the action of dFOXO on adult mortality in Drosophila. Aging Cell. 2007; 6:429–38. https://doi.org/10.1111/j.1474-9726.2007.00290.x [PubMed]
- 73. Delaney JR, Murakami CJ, Olsen B, Kennedy BK, Kaeberlein M. Quantitative evidence for early life fitness defects from 32 longevity-associated alleles in yeast. Cell Cycle. 2011; 10:156–65. https://doi.org/10.4161/cc.10.1.14457 [PubMed]
- 74. Beach A, Titorenko VI. In search of housekeeping pathways that regulate longevity. Cell Cycle. 2011; 10:3042–44. https://doi.org/10.4161/cc.10.18.16947 [PubMed]
- 75. Kyryakov P, Beach A, Richard VR, Burstein MT, Leonov A, Levy S, Titorenko VI. Caloric restriction extends yeast chronological lifespan by altering a pattern of age-related changes in trehalose concentration. Front Physiol. 2012; 3:256. https://doi.org/10.3389/fphys.2012.00256 [PubMed]
- 76. McColl G, Jenkins NL, Walker DW, Lithgow GJ. Testing evolutionary theories of aging. Ann N Y Acad Sci. 2000; 908:319–20. https://doi.org/10.1111/j.1749-6632.2000.tb06663.x [PubMed]
- 77. Walker DW, McColl G, Jenkins NL, Harris J, Lithgow GJ. Evolution of lifespan in C. elegans. Nature. 2000; 405:296–97. https://doi.org/10.1038/35012693 [PubMed]
- 78. Marden JH, Rogina B, Montooth KL, Helfand SL. Conditional tradeoffs between aging and organismal performance of Indy long-lived mutant flies. Proc Natl Acad Sci USA. 2003; 100:3369–73. https://doi.org/10.1073/pnas.0634985100 [PubMed]
- 79. Jenkins NL, McColl G, Lithgow GJ. Fitness cost of extended lifespan in Caenorhabditis elegans. Proc Biol Sci. 2004; 271:2523–26. https://doi.org/10.1098/rspb.2004.2897 [PubMed]
- 80. Anderson JL, Reynolds RM, Morran LT, Tolman-Thompson J, Phillips PC. Experimental evolution reveals antagonistic pleiotropy in reproductive timing but not life span in Caenorhabditis elegans.. J Gerontol A Biol Sci Med Sci. 2011; 66:1300–08. https://doi.org/10.1093/gerona/glr143 [PubMed]
- 81. Qian W, Ma D, Xiao C, Wang Z, Zhang J. The genomic landscape and evolutionary resolution of antagonistic pleiotropy in yeast. Cell Reports. 2012; 2:1399–410. https://doi.org/10.1016/j.celrep.2012.09.017 [PubMed]
- 82. Medawar PB. An Unsolved Problem of Biology. 1952, London, HK Lewis.
- 83. Charlesworth B. Fisher, Medawar, Hamilton and the evolution of aging. Genetics. 2000; 156:927–31. [PubMed]
- 84. Charlesworth B. Patterns of age-specific means and genetic variances of mortality rates predicted by the mutation-accumulation theory of ageing. J Theor Biol. 2001; 210:47–65. https://doi.org/10.1006/jtbi.2001.2296 [PubMed]
- 85. Williams GC. Pleiotropy, natural selection, and the evolution of senescence. Evolution. 1957; 11:398–411. https://doi.org/10.2307/2406060
- 86. Kirkwood TB. Evolution of ageing. Nature. 1977; 270:301–04. https://doi.org/10.1038/270301a0 [PubMed]
- 87. Kirkwood TB, Holliday R. The evolution of ageing and longevity. Proc R Soc Lond B Biol Sci. 1979; 205:531–46. https://doi.org/10.1098/rspb.1979.0083 [PubMed]
- 88. Kirkwood TB, Austad SN. Why do we age? Nature. 2000; 408:233–38. https://doi.org/10.1038/35041682 [PubMed]
- 89. Garay E, Campos SE, González de la Cruz J, Gaspar AP, Jinich A, Deluna A. High-resolution profiling of stationary-phase survival reveals yeast longevity factors and their genetic interactions. PLoS Genet. 2014; 10:e1004168. https://doi.org/10.1371/journal.pgen.1004168 [PubMed]
- 90. Madeo F, Herker E, Maldener C, Wissing S, Lächelt S, Herlan M, Fehr M, Lauber K, Sigrist SJ, Wesselborg S, Fröhlich KU. A caspase-related protease regulates apoptosis in yeast. Mol Cell. 2002; 9:911–17. https://doi.org/10.1016/S1097-2765(02)00501-4 [PubMed]
- 91. Fabrizio P, Battistella L, Vardavas R, Gattazzo C, Liou LL, Diaspro A, Dossen JW, Gralla EB, Longo VD. Superoxide is a mediator of an altruistic aging program in Saccharomyces cerevisiae. J Cell Biol. 2004; 166:1055–67. https://doi.org/10.1083/jcb.200404002 [PubMed]
- 92. Herker E, Jungwirth H, Lehmann KA, Maldener C, Fröhlich KU, Wissing S, Büttner S, Fehr M, Sigrist S, Madeo F. Chronological aging leads to apoptosis in yeast. J Cell Biol. 2004; 164:501–07. https://doi.org/10.1083/jcb.200310014 [PubMed]
- 93. Wissing S, Ludovico P, Herker E, Büttner S, Engelhardt SM, Decker T, Link A, Proksch A, Rodrigues F, Corte-Real M, Fröhlich KU, Manns J, Candé C, et al. An AIF orthologue regulates apoptosis in yeast. J Cell Biol. 2004; 166:969–74. https://doi.org/10.1083/jcb.200404138 [PubMed]
- 94. Mazzoni C, Herker E, Palermo V, Jungwirth H, Eisenberg T, Madeo F, Falcone C. Yeast caspase 1 links messenger RNA stability to apoptosis in yeast. EMBO Rep. 2005; 6:1076–81. https://doi.org/10.1038/sj.embor.7400514 [PubMed]
- 95. Allen C, Büttner S, Aragon AD, Thomas JA, Meirelles O, Jaetao JE, Benn D, Ruby SW, Veenhuis M, Madeo F, Werner-Washburne M. Isolation of quiescent and nonquiescent cells from yeast stationary-phase cultures. J Cell Biol. 2006; 174:89–100. https://doi.org/10.1083/jcb.200604072 [PubMed]
- 96. Li W, Sun L, Liang Q, Wang J, Mo W, Zhou B. Yeast AMID homologue Ndi1p displays respiration-restricted apoptotic activity and is involved in chronological aging. Mol Biol Cell. 2006; 17:1802–11. https://doi.org/10.1091/mbc.E05-04-0333 [PubMed]
- 97. Büttner S, Eisenberg T, Carmona-Gutierrez D, Ruli D, Knauer H, Ruckenstuhl C, Sigrist C, Wissing S, Kollroser M, Fröhlich KU, Sigrist S, Madeo F. Endonuclease G regulates budding yeast life and death. Mol Cell. 2007; 25:233–46. https://doi.org/10.1016/j.molcel.2006.12.021 [PubMed]
- 98. Fabrizio P, Longo VD. Chronological aging-induced apoptosis in yeast. Biochim Biophys Acta. 2008; 1783:1280–85. https://doi.org/10.1016/j.bbamcr.2008.03.017 [PubMed]
- 99. Goldberg AA, Bourque SD, Kyryakov P, Gregg C, Boukh-Viner T, Beach A, Burstein MT, Machkalyan G, Richard V, Rampersad S, Cyr D, Milijevic S, Titorenko VI. Effect of calorie restriction on the metabolic history of chronologically aging yeast. Exp Gerontol. 2009; 44:555–71. https://doi.org/10.1016/j.exger.2009.06.001 [PubMed]
- 100. Burstein MT, Kyryakov P, Beach A, Richard VR, Koupaki O, Gomez-Perez A, Leonov A, Levy S, Noohi F, Titorenko VI. Lithocholic acid extends longevity of chronologically aging yeast only if added at certain critical periods of their lifespan. Cell Cycle. 2012; 11:3443–62. https://doi.org/10.4161/cc.21754 [PubMed]
- 101. Sheibani S, Richard VR, Beach A, Leonov A, Feldman R, Mattie S, Khelghatybana L, Piano A, Greenwood M, Vali H, Titorenko VI. Macromitophagy, neutral lipids synthesis, and peroxisomal fatty acid oxidation protect yeast from “liponecrosis”, a previously unknown form of programmed cell death. Cell Cycle. 2014; 13:138–47. https://doi.org/10.4161/cc.26885 [PubMed]
- 102. Richard VR, Beach A, Piano A, Leonov A, Feldman R, Burstein MT, Kyryakov P, Gomez-Perez A, Arlia-Ciommo A, Baptista S, Campbell C, Goncharov D, Pannu S, et al. Mechanism of liponecrosis, a distinct mode of programmed cell death. Cell Cycle. 2014; 13:3707–26. https://doi.org/10.4161/15384101.2014.965003 [PubMed]
- 103. Arlia-Ciommo A, Svistkova V, Mohtashami S, Titorenko VI. A novel approach to the discovery of anti-tumor pharmaceuticals: searching for activators of liponecrosis. Oncotarget. 2016; 7:5204–25. [PubMed]
- 104. Goldberg AA, Kyryakov P, Bourque SD, Titorenko VI. Xenohormetic, hormetic and cytostatic selective forces driving longevity at the ecosystemic level. Aging (Albany NY). 2010; 2:461–70. https://doi.org/10.18632/aging.100186 [PubMed]
- 105. Calabrese EJ, Mattson MP. Hormesis provides a generalized quantitative estimate of biological plasticity. J Cell Commun Signal. 2011; 5:25–38. https://doi.org/10.1007/s12079-011-0119-1 [PubMed]
- 106. Burstein MT, Beach A, Richard VR, Koupaki O, Gomez-Perez A, Goldberg AA, Kyryakov P, Bourque SD, Glebov A, Titorenko VI. Interspecies chemical signals released into the environment may create xenohormetic, hormetic and cytostatic selective forces that drive the ecosystemic evolution of longevity regulation mechanisms. Dose Response. 2012; 10:75–82. https://doi.org/10.2203/dose-response.11-011.Titorenko [PubMed]
- 107. Calabrese V, Cornelius C, Dinkova-Kostova AT, Iavicoli I, Di Paola R, Koverech A, Cuzzocrea S, Rizzarelli E, Calabrese EJ. Cellular stress responses, hormetic phytochemicals and vitagenes in aging and longevity. Biochim Biophys Acta. 2012; 1822:753–83. https://doi.org/10.1016/j.bbadis.2011.11.002 [PubMed]
- 108. Arlia-Ciommo A, Piano A, Leonov A, Svistkova V, Titorenko VI. Quasi-programmed aging of budding yeast: a trade-off between programmed processes of cell proliferation, differentiation, stress response, survival and death defines yeast lifespan. Cell Cycle. 2014; 13:3336–49. https://doi.org/10.4161/15384101.2014.965063 [PubMed]
- 109. Lutchman V, Medkour Y, Samson E, Arlia-Ciommo A, Dakik P, Cortes B, Feldman R, Mohtashami S, McAuley M, Chancharoen M, Rukundo B, Simard É, Titorenko VI. Discovery of plant extracts that greatly delay yeast chronological aging and have different effects on longevity-defining cellular processes. Oncotarget. 2016; 7:16542–66. [PubMed]