



Introduction
Aging is a progressive deterioration of physiological functions impairing the ability of an organism to cope with endogenous or exogenous stresses and maintain homeostasis. This in turn leads to increased susceptibility to diseases and death. The study of human progeroid syndromes have greatly advanced the understanding of the aging process [1]. One fascinating human aging disorder is Werner syndrome (WS). WS is a human autosomal recessive disorder characterized by genomic instability and the premature onset of a number of age-related diseases [2-5]. The defective enzyme responsible for WS possesses a 3′–5′ exonuclease activity in addition to a 3′–5′ helicase activity [6-8] and is involved in DNA repair, replication, transcription, and telomere maintenance [9-13]. We previously generated a mouse model with a deletion in the helicase domain of the murine WRN homologue (hereafter referred as WrnDhel/Dhel) [14] that recapitulates most of the WS phenotypes, including an abnormal hyaluronic acid excretion, higher reactive oxygen species (ROS) levels, dyslipidemia, increased genomic instability, and cancer incidence. Overall, such mutant mice have a 10-15% decreased of their mean life span [15, 16].
The WRN protein is a member of the RecQ family of DNA helicases [4]. It is highly conserved across species including in invertebrates such as the small worm Caenorhabditis elegans (C. elegans). Interestingly, the exonuclease and the DNA helicase domains homologous to the human WRN protein are encoded by two different genes in C. elegans [17]. The C. elegans wrn-1 gene codes for the ATP-dependent 3′–5′ DNA helicase capable of unwinding a variety of DNA structures [18]. Notably, it has been shown that the RNAi knockdown of the C. elegans wrn-1 gene shortens the life span, increases sensitivity to DNA damage, and accelerates aging phenotypes [17].
Recent discoveries in the fields of development, cancer, and aging have indicated that small non-coding RNAs play a major role in alterations associated with these biological processes. An important class of non-coding RNAs that has been studied in the context of C. elegans aging are the microRNAs (miRNAs) [19-23]. The miRNAs are short RNAs (~22nt) that regulate post-transcriptional gene expression via base pairing to partially complementary sites mainly found in the 3’ UTRs of messenger RNAs (mRNAs). miRNAs down regulate protein expression by inhibiting mRNA translation and/or mRNA stability [20]. Individual miRNAs can modulate multiple mRNA targets, and individual mRNAs can be regulated by multiple, distinct miRNAs [20]. Very few studies using rodent tissues have been performed to elucidate the role of miRNAs in aging [24-27], often with contradictory results [28, 29].
In this study, we report the differential expression of several miRNAs in the livers of young (three months old) WrnΔhel/Δhel mice compared to age-matched wild type animals. Among them, one conserved miRNA in animals (miR-124) was down regulated in both the liver of WrnDhel/Dhel mice and in whole wrn-1 C. elegans mutants. Deletion of mir-124 in C. elegans resulted in a decrease in life span, an increase in reactive oxygen species (ROS) production, a decrease in ATP levels, and an increase in the aging marker lipofuscin. All these phenotypes could be reversed in mir-124 mutation strains after vitamin C treatment. These results implicate a role for the conserved miR-124 in aging in C. elegans.
Results
The liver of WrnΔhel/Δhel mice show differential expression of miR-375 and miR-124
We have previously shown that in WrnΔhel/Δhel mice, the liver is the first tissue to show morphological changes compared to age-matched wild type animals [16, 30]. Interestingly, the liver undergoes substantial modifications in structure and function in old age including include alterations in liver mass, blood flow, and sinusoidal cell morphology [31]. These changes are associated with significant impairment of many hepatic metabolic and detoxification activities, with implications for systemic aging and age-related disease. We therefore focused our study on the hepatic tissue as the liver plays a pivotal role in whole body homeostasis through the maintenance of nutrient, drug, hormone, and metabolic processes. Total RNA from the liver of two WrnDhel/Dhel and two wild type mice at three months of age was extracted to analyze the expression of 755 different miRNAs using the TaqMan-based Array. Although no gross hepatic morphological difference could be observed between WrnDhel/Dhel mice and wild type mice at three months of age, the liver of WrnDhel/Dhel mice exhibited changes in the expression of a number of miRNAs compared to wild type mice. Supplementary Tables S1 and S2 provide the raw data on all miRNAs. Table 1 summarizes the list of differentially expressed miRNAs in the liver of WrnDhel/Dhel mice compared to wild type mice.
Table 1. List of miRNAs differentially expressed in the liver of three months old WrnΔhel/Δhel mutant compared to wild type mice with an adjusted P-value < 0.1
| miRNA | expression* | P-value | Adjusted P-value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| let-7i | down | 0.000058 | 0.0071 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-350 | down | 0.000250 | 0.0153 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-2183 | up | 0.000386 | 0.0157 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-375 | up | 0.000249 | 0.0362 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-15a | down | 0.002092 | 0.0638 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-124 | down | 0.000883 | 0.0641 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-509-3p | down | 0.001641 | 0.0641 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Expression of miRNAs in the liver of WrnΔhel/Δhel mutant compared to wild type mice. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
We next validated the differential expression of the seven miRNAs listed in Table 1 using the liver tissues of four different WrnDhel/Dhelmutant and four wild type mice (three months of age) Of the seven miRNAs tested, only miR-375 and miR-124 showed significant differential expressions in WrnDhel/Dhel mutant compared to wild type animals (Figure 1A and Supplementary Figure S1). miR-375 was up regulated more than three-fold and miR-124 was down regulated by ten-fold in the liver of WrnDhel/Dhel mutant mice compared to the liver of wild type animals (Figure 1A).
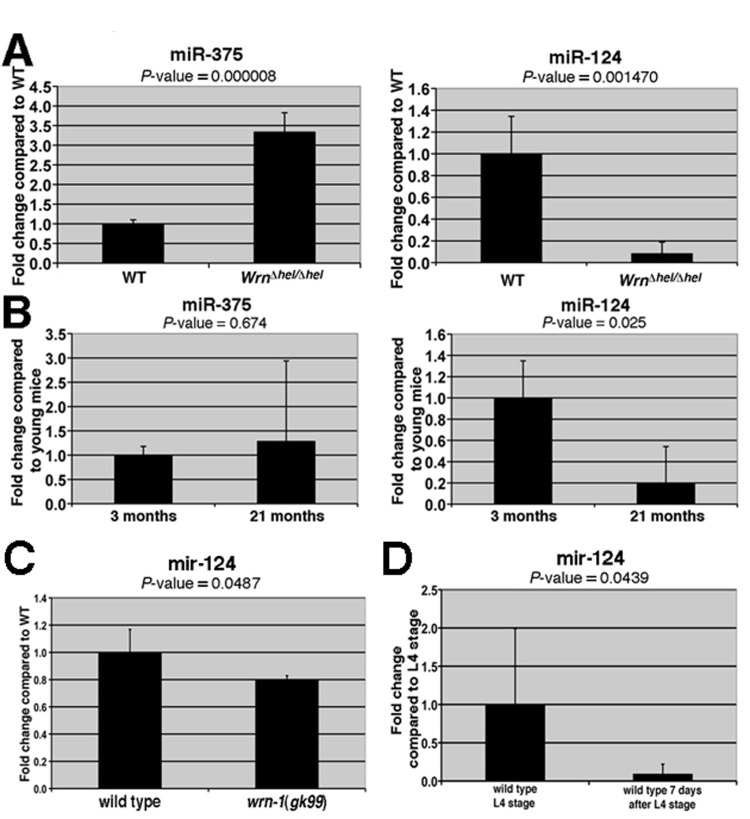
Figure 1. Expression levels of miRNAs in the liver of WrnDhel/Dhel mice compared to wild type mice and in the whole body of wild type and wrn-1(gk99) worms. (A) Total RNA from four mice (at three months of age) of the indicated genotype was used for the quantitative RT-PCR analyses. The levels of the indicated miRNAs in the WrnDhel/Dhel mice are relative to the wild type (WT) animals. (B) Expression levels of miR-375 and miR-124 in the liver of four young (three months old) and four old (21 months old) wild type animals. The levels of the indicated miRNAs in the old wild type mice are relative to the young wild type animals. (C) Expression level of mir-124 in wild type and wrn-1(gk99) strains. Twenty-five 7-day old adult worms (post-larval L4 stage) of each strain were sorted and collected for total RNA extraction. The quantification of mir-124 were measured by quantitative RT-PCR (TaqMan assay) and compared with the levels found in wild type animals. (D) mir-124 expression levels in young and older adults wild type worms. All data were normalized by the quantification of the small nucleolar RNA (sn2841). The error bars represent the 95% confidence interval of three independent experiments. The P-values (unpaired Student's t-test) are indicated above each graph.
To determine whether miR-375 and miR-124 were also differentially expressed during aging, quantitative RT-PCR was performed on the liver tissues of four young (three months) and four old (21 months) wild type mice. miR-124 was significantly decreased (by five-fold) in the livers of old wild type mice compared to young wild type mice (Figure 1B). In contrast, there was a non-significant increase in miR-375 level in the liver of old wild type animals. These results indicate that the expression of miR-124 correlates inversely with age in the liver of mice (Figure 1B).
The impact of the Wrn helicase on miR-124 expression is conserved in C. elegans
We next determine whether the observed alteration of the miRNAs in mice could be a global phenomenon during aging by studying these miRNAs in the nematode C. elegans. A search in the miRNA database miRBase (www.mirbase.org) revealed that miR-124 is conserved in the short-lived C. elegans but not the miR-375. We first determined if the modulation of miR-124 is also conserved in C. elegans animals carrying a loss-of-function deletion of the wrn-1 gene (wrn-1(gk99) allele) that encodes the human WRN helicase ortholog [32]. It has been reported that a depletion of the C. elegans wrn-1 gene product by RNAi reduces the life span of this animal [17]. Consistent with these findings, we found that the wrn-1(gk99) mutant animals had a reduced life span when compared to the wild type (N2) animals (Figure 2A). The median life span of the wrn-1(gk99) animals was 6.8 days compared to 9.0 days for the wild type strain (32% decrease; log-rank test: P-value = 1.4 × 10−11). Interestingly, we observed that the expression of the conserved miR-124 is significantly reduced by 20% in the wrn-1(gk99) animals (unpaired Student's t-test: P = 0.048) compared to the wild type strain (Figure 1C). Furthermore, we found that mir-124 expression is also reduced in older wild type worms (seven days after L4 stage) compared to young worms (at the L4-larvae developmental stage) (Figure 1D). These results indicate that miR-124 expression is decreased in both Mus musculus and C. elegans during aging and in animals with a mutation in the WRN helicase ortholog.
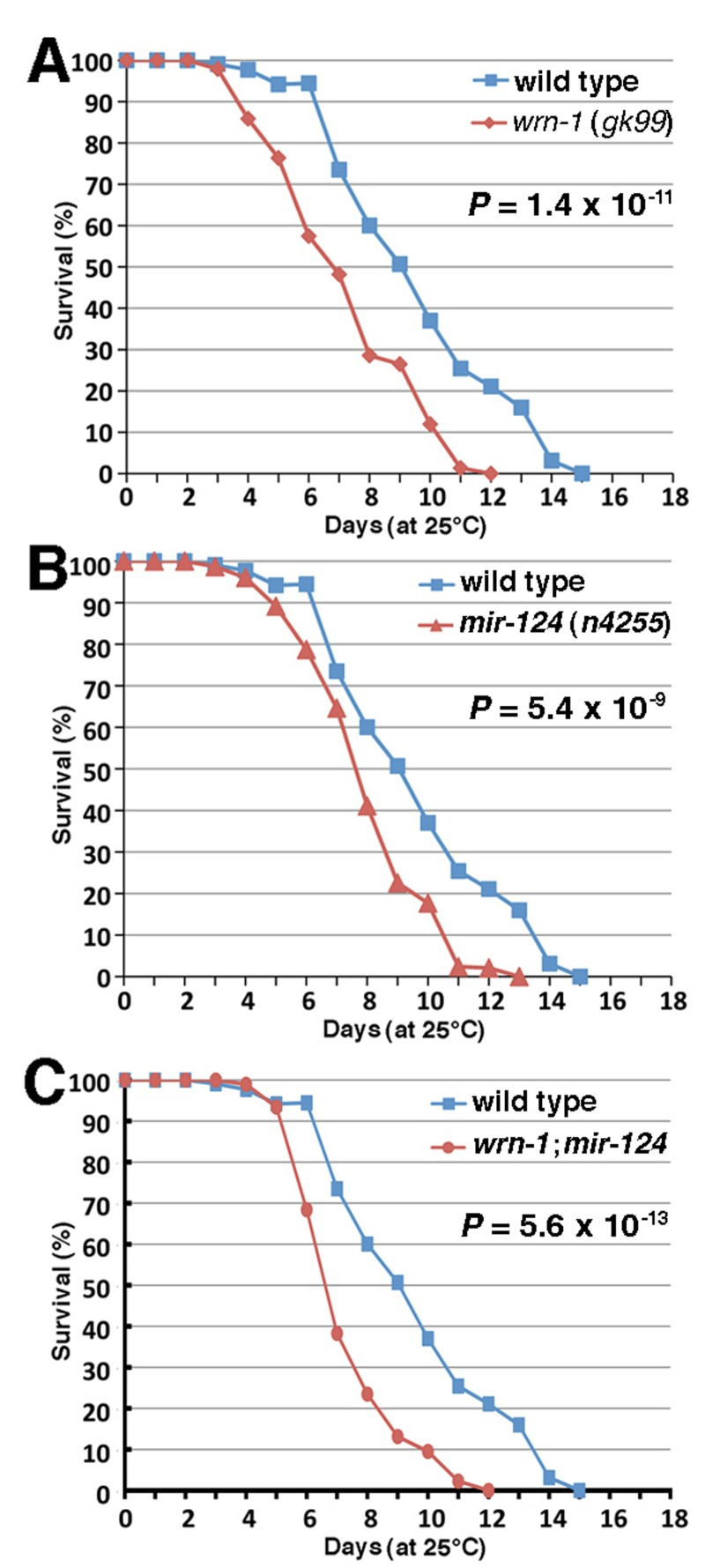
Figure 2. Life span of wild type and mutant C. elegans animals. (A) Survival curves of wild type (N2) and wrn-1(gk99) animals grown at 25°C. (B) Survival curves of wild type (N2) and mir-124 (n4255) strains grown at 25°C. (C) Survival curves of wild type (N2) and wrn-1;mir-124 double mutant animals grown at 25°C. All experiments were performed with five different pools of 20 to 30 animals of each genotype. The indicated P-values were obtained using the log-rank test method.
The loss of mir-124 causes a reduction of life span inC. elegans
To assess the impact of the loss of miR-124 on aging, we measured the life span of worms carrying a deletion of themir-124 gene (mir-124(n4255)) [33]. As shown in Figure 2B, the median life span of mir-124(n4255) worms was significantly decreased by 15% (7.7 days versus 9.0 days) compared to the wild type animals (log-rank test: P = 5.4 × 10−9). Notably, animals carrying both deletion of mir-124 and wrn-1 genes (wrn-1;mir-124 animals) displayed a more severe decrease in their life span (48% decrease compared to wild type; log-rank test: P = 5.6 × 10−13) than the single loss of either gene (Figure 2C). These results indicate that both genes are important in the life span of C. elegans.
The loss of wrn-1 and mir-124 leads to an increase in reactive oxygen species (ROS) generation and a reduction in ATP levels
We have reported that WrnDhel/Dhel mice exhibit increased ROS and decreased ATP levels in different tissues compared to age-matched wild type animals [16, 34]. To determine whether the loss of wrn-1 also affects ROS levels in C. elegans, we measured ROS levels in whole wrn-1(gk99) worms with dichlorofluorescein (DCFA) staining as described previously [34]. Although not significant, the wrn-1(gk99) mutant worms exhibited an 8% increase in overall ROS levels compared to the wild type strain (Figure 3A). The loss of mir-124, in return, led to a significant increase in overall ROS levels (16% increase; P = 0.0442). Interestingly, the loss of both wrn-1 and mir-124 resulted in a 40% increase in whole body ROS levels compared to wild type worms (P = 0.0008) (Figure 3A).
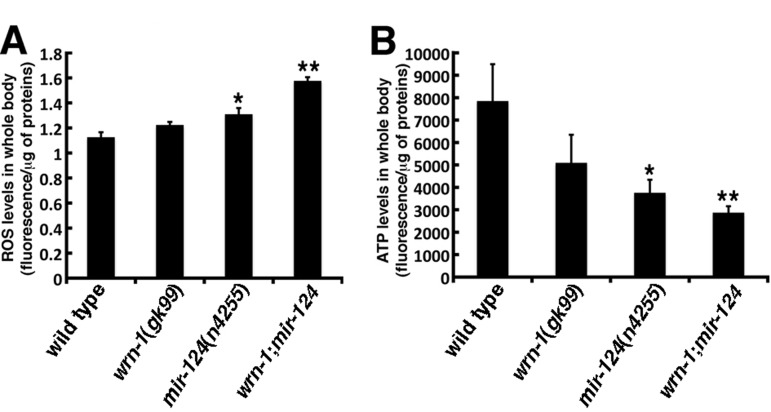
Figure 3. Reactive oxygen species (ROS) and ATP levels in wild type and mutant C. elegans strains. (A) ROS levels in whole body of 7-day adult old worms (post-larval L4 stage) detected with dichlorofluorescein. Data are expressed as units of fluorescence per micrograms of proteins. (Unpaired Student's t-test; *P = 0.0442 vs. wild type; **P = 0.0008 vs. wild type). (B) ATP levels in whole body of 7-day old adult worms (post-larval L4 stage). (Unpaired Student's t-test; *P = 0.0410 vs. wild type; **P = 0.0137 vs. wild type). Data are expressed as units of fluorescence per micrograms of proteins. Twenty-five worms of each genotype were collected for the ROS or ATP measurements. Experiments were performed with three independent pools of animals.
We next measured the impact of the loss of wrn-1 and/or mir-124 on ATP levels. The wrn-1(gk99) animals exhibited a 35% decrease in overall ATP levels compared to the wild type strain (Figure 3B), while the mir-124(n4255) animals exhibited a 52% decrease in the overall ATP levels compared to the wild type strain (P = 0.0409). Finally, the double mutant wrn-1;mir-124 worms showed a 63% decrease in whole body ATP levels compared to wild type animals (P = 0.0137). These results indicate that the loss of both wrn-1 and mir-124 functions significantly affect ATP levels in C. elegans.
The loss of wrn-1 and mir-124 lead to an increase of the aging marker lipofuscin
To determine whether the reduced life span observed inmir-124(n4255) worms was due to a progeroid phenotype, accumulation of the aging marker lipofuscin was examined. The intensity of the fluorescence observed in the wrn-1(gk99) animals was 21-fold stronger than wild type worms at the third day into adulthood (Figure 4). Similarly, the lipofuscin fluorescence observed in mir-124(n4255) mutant worms was also stronger than in the wild type animals. Finally, there was a synergic effect on the accumulation of lipofuscin in the double mutant wrn-1;mir-124 worms (Figure 4). These results indicate that the animals lacking either wrn-1 or mir-124 exhibit a progeroid phenotype that is exacerbated by the loss of both genes.
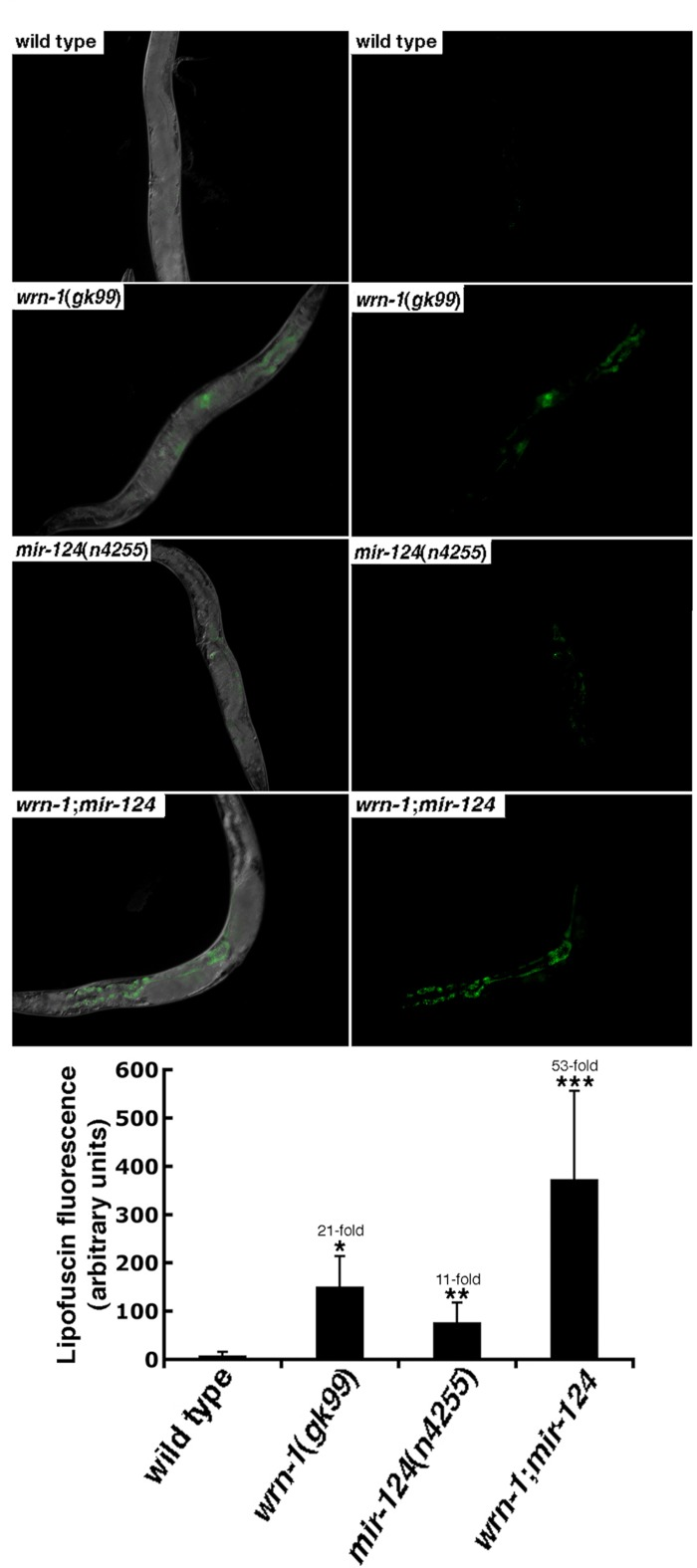
Figure 4. The aging marker lipofuscin is increased in mutant C. elegans strains. Representative photographs of wild type (N2), wrn-1(gk99), mir-124(n4255), and wrn-1;mir-124 double mutant worms at three days into adulthood. Panels on the right represent the lipofuscin autofluorescence alone. All pictures were taken at the same exposure time. Magnification is 10 X. The histogram at the bottom represents the average intensity of lipofuscin autofluorescence in the different C. elegans strains. Ten to fifteen three-days old (three days into adulthood) worms of each strain were photographed and the fluorescence intensity was quantified using Adobe Photoshop. The fold increase in fluorescence intensity compared to wild type animals is indicated. (Unpaired Student's t-test; *P = 0.00002 for wrn-1(gk99) vs. wild type; **P = 0.00222 for mir-124(n4255) vs. wild type; and ***P = 0.00078 for wrn-1;mir-124 vs. wild type).
Vitamin C restores the normal life span of wrn-1(gk99) and mir-124(n4255) mutant strains
Previously, we reported that vitamin C restored the normal life span of WrnDhel/Dhel mice [16]. We thus decided to test the impact of 10 mM ascorbate [35] on the life span of each C. elegans mutant strain. Vitamin C significantly increased the median life span of wrn-1(gk99) animals when they were grown with a diet containing vitamin C (log-rank test: P = 1.4 × 10−7; Figure 5A). Furthermore, this lifespan extension effect was comparable to wild type animals grown on normal media. The median life span of mir-124(n4255) mutant worms was also significantly increased to a level similar to wild type animals upon vitamin C supplementation (Figure 5B; log-rank test: P = 3.0 × 10−9). These results indicate that vitamin C significantly increased the life span of animals lacking the wrn-1 or mir-124 genes.
Finally, we determined the life span of double mutant wrn-1;mir-124 worms treated with vitamin C. While vitamin C did extend the lifespan of these double mutant worms from 6.6 days to 8.4 days (Figure 5C, P = 3.1 × 10−6), it was not as a dramatic effect as observed for the single mutant worms. Furthermore, vitamin C treatment did not increase the life span of the double mutants to that of the untreated wild type worms (Figure 5C; log-rank test: P = 0.0163).
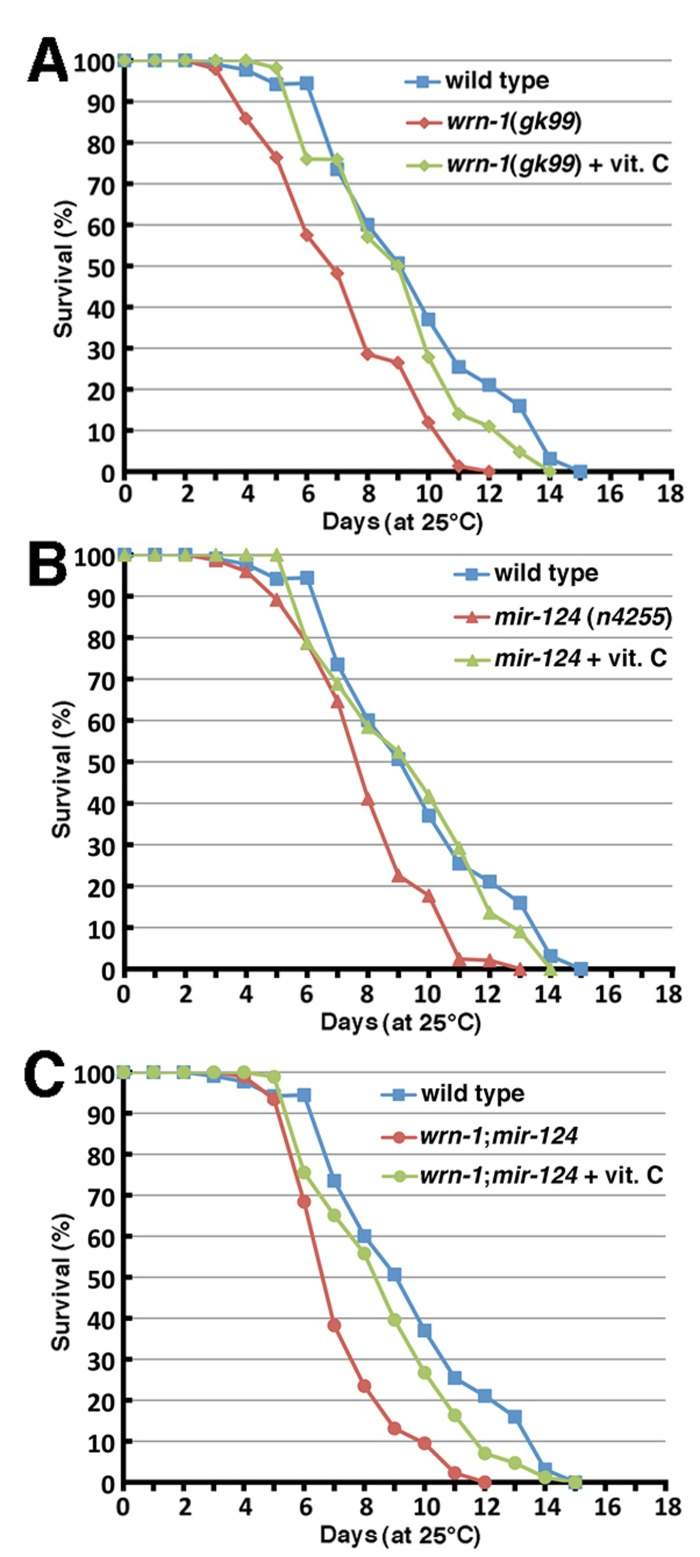
Figure 5. Impact of vitamin C on the life span of wild type and mutant C. elegans strains. (A) Survival curves of vitamin C treated wrn-1(gk99) and of untreated wild-type (N2) and wrn-1(gk99) C. elegans strains grown at 25°C (vitamin C treated wrn-1(gk99) vs. untreated wrn-1(gk99) worms: P = 1.4 × 10−7; vitamin C treated wrn-1(gk99) vs. untreated wild-type worms: P = 0.0778). (B) Survival curves of vitamin C treated mir-124(n4255) and of untreated wild-type (N2) and mir-124(n4255)C. elegans strains grown at 25°C (vitamin C treated mir-124(n4255) vs. untreated mir-124(n4255) worms: P = 3.0 × 10−9; vitamin C treated wrn-1(gk99) vs. untreated wild-type worms: P = 0.359). (C) Survival curves of vitamin C treated wrn-1;mir-124 and of untreated wild-type (N2) and wrn-1;mir-124 double mutant C. elegans strains grown at 25°C (vitamin C treated wrn-1;mir-124 vs. untreated wrn-1;mir-124 worms: P = 3.1 × 10−6; vitamin C treated wrn-1;mir-124 vs. untreated wild-type worms: P = 0.0163). All experiments were performed three to four times with 20 to 30 worms per genotype. P-values were obtained using the log-rank test method.
Vitamin C decreases ROS levels in all mutant strains
We next examined the effect of vitamin C on ROS levels in whole worms of each strain. Twenty-five 7-day old adult worms (timed from the post-larval L4 stage) of each genotype were treated with 10 mM vitamin C and then ROS levels were measured. There was no significant difference between untreated and vitamin C-treated wild type worms. In wrn-1(gk99) and mir-124(n4255) worms, vitamin C treatment significantly lowered ROS levels compared to untreated worms (Figure 6A; P < 0.00005). Finally, vitamin C also significantly decreased ROS levels in wrn-1;mir-124 worms compared to the untreated wrn-1;mir-124 animals (P = 0.00009) (Figures 6A). Overall, these results indicate that vitamin C significantly decreased ROS levels in all mutant strains tested.
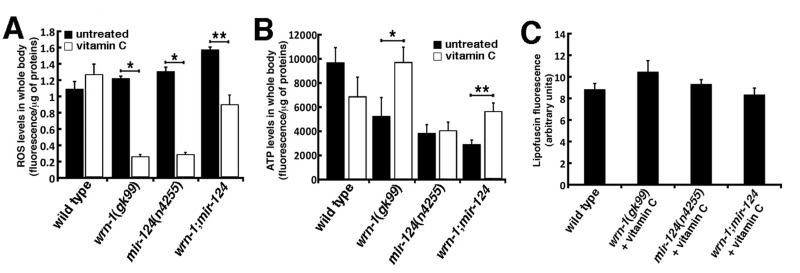
Figure 6. Impact of vitamin C on ROS, ATP, and lipofuscin levels in wild type and mutant C. elegans strains. (A) ROS levels in whole body of 7-day adult old worms (post-larval L4 stage) detected with dichlorofluorescein. Data are expressed as units of fluorescence per milligrams of proteins. (Unpaired Student's t-test; *P < 0.00005; ** P = 0.00009). (B) ATP levels in whole body of 7-day old adult worms (post-larval L4 stage). (Unpaired Student's t-test; *P = 0.00972; **P = 0.0002). Twenty-five worms of each genotype were collected for the ROS or ATP measurements. Experiments were performed in triplicate. (C) Histogram representing the average intensity of autofluorescence lipofuscin in the different C. elegans strains treated with vitamin C compared to untreated wild type worms. Ten to fifteen three-days old (three days into adulthood) worms of each strain were photographed and the fluorescence intensity was quantified using Adobe Photoshop.
Vitamin C increases ATP levels only in the wrn-1(gk99) mutant strain
We also measured ATP levels in vitamin C treated mutant worms. ATP levels were decreased in wild type treated worms compared to untreated wild type worms but this decrease was not statistically different. ATP levels in vitamin C-treated wrn-1(gk99) worms were similar to the ATP levels of untreated wild type worms (Figure 6B). In contrast, vitamin C significantly increased ATP levels in wrn-1(gk99) worms compared to the untreated wrn-1(gk99) animals by 1.9-fold (P = 0.00972; Figure 6B).
ATP level was not significantly increased in vitamin C treated mir-124(n4255) animals compared to the untreated mir-124(n4255) worms (Figure 6B) and was still at a lower level than untreated wild type animals (P = 0.0155). Thus, vitamin C did not normalized the amount of ATP in mir-124(n4255) worms to the wild type levels. There was a 1.9-fold increase in ATP levels in vitamin C treated wrn-1;mir-124 double mutant worms compared to untreated wrn-1;mir-124 animals (P = 0.0002; Figure 6B). However, the amount of ATP in vitamin C treated wrn-1;mir-124 double mutant worms was still lower than untreated wild type animals (P = 0.0440). Overall, these results indicate that vitamin C significantly increased ATP levels only in worms bearing the wrn-1(gk99) mutation.
Vitamin C decreases lipofuscin levels in all mutant strains to the level of untreated wild type animals
The intensity of autofluorescence from lipofuscin accumulation was examined in all the mutant strains treated with vitamin C. As indicated in Figure 6C, vitamin C decreased the intensity of autofluorescence in all the mutant strains to untreated wild type levels. These results indicate that vitamin C normalized lipofuscin accumulation in wrn-1(gk99), mir-124(n4255), and wrn-1;mir-124 worms.
Discussion
Important parallels between mouse and C. elegans with a mutation in the WRN helicase
In this study, we have demonstrated that a C. elegans animal carrying a deletion of the wrn-1 helicase have a reduced life span, and importantly this phenotype is similar to mice lacking the DNA helicase activity of the human WRN ortholog [16, 30, 34]. Thus, both models can be used to identify and assess the impact of specific genes that, with the WRN orthologs, affect health or life span. The short life span of the C. elegans allows a rapid evaluation of the impact of a gene on aging, which can then be translated to a more complex organism like the mouse. In this study, we identified miR-124 as a conserved miRNA in both mouse and worm animal models. miR-124 has a role in premature aging through the loss of a functional WRN ortholog helicase activity, although the mechanism by which the loss of WRN affects miR-124 expression remains somewhat unknown. Nevertheless, we demonstrate that a deletion of the mir-124 gene ortholog in C. elegans results in reduced life span, increased whole body ROS levels, and reduced ATP levels. Because total inactivation of both wrn-1 and mir-124 genes had a greater negative impact on ROS and ATP levels than inactivating wrn-1 alone, these results suggest that the decrease of the miR-124 miRNA can contribute to several key biological processes affected in WrnDhel/Dhel mice [15, 16, 34]. In addition, the deletion of mir-124 accelerated the accumulation of the aging marker lipofuscin in C. elegans and thus highlights the importance of this miRNA in the progeroid phenotype.
The expression of miR-124 was not only reduced in the livers of young WrnΔhel/Δhel mice compared to age-matched wild type mice, but it was also reduced in the livers of old wild type mice compared to young wild type mice. These results indicate that the miR-124 expression signature in the liver of young WrnΔhel/Δhel mice corresponds to the miR-124 signature in old wild type animals. To our knowledge, this is the first study showing a significant altered expression of miR-124 in the liver of aging mice. Previous studies have not shown an alteration of miR-124 during normal hepatic aging in mice or rats, or in the long-lived Ames dwarf mice [24, 27, 36]. This difference may be due to the different techniques used for the initial miRNA detection. Previous studies utilized hybridization of labeled molecules on nitrocellulose-based microarray [24, 27, 36] that may be less sensitive than direct quantitative RT-PCR of individual miRNA as was used in this study [37]. Interestingly, the level of miR-124 has also been reported to be down regulated in skeletal muscle of old mice compared to young mice [25]. These results, together with our data, indicate that a decrease of miR-124 can be considered as a common signature in the liver and muscle of aging mice. Our observation of a significant decrease in miR-124 levels in aging C. elegans further supports the role of this conserved miRNA in the molecular signature of aging in different animal species.
The miR-124 has been shown to be involved in neurogenesis not only in mouse but also in C. elegans [38, 39]. More precisely, the expression of miR-124 in the mouse brain is associated with the differentiation status of neuronal cells [38]. However, miR-124 is expressed in cell types other than neurons [40, 41]. Of relevance to our study, miR-124 is also expressed in the normal human liver [42]. As miR-124 is a regulator of several proteins involved in insulin exocytosis and intracellular signaling in pancreatic beta cell lines [40, 41], it is possible that miR-124 may alter insulin action in vivo directly impacting on organismal homeostasis and aging. Importantly, the insulin/insulin-like growth factor-1 signaling pathway is a strong regulator of longevity in C. elegans [23, 43, 44]. Noticeably, insulin-like peptides are primarily released from neurons in C. elegans [23]. Thus, the mutant C. elegans strains described in this study gives us relevant models to thoroughly decipher the molecular mechanisms involved in WS and aging in general. As miR-124 will affect protein expression by destabilizing RNA levels of target genes or by inhibiting translation of target mRNAs, the next step is to perform large scale proteomic analyses to identify proteins in our Mus musculus and C. elegans animal models involved in the insulin signaling pathway, redox balance, energy homeostasis, and healthy aging.
Vitamin C normalizes the life span of mutant wrn-1 and mir-124 strains We recently found that Vitamin C supplementation rescued the shorter mean life span of WrnDhel/Dhel mice and reversed several age-related abnormalities in adipose, cardiac, and liver tissues [16]. In this study, we show that vitamin C also rescued the shorter life span of both wrn-1(gk99) and the mir-124(n4255) mutant animals. Furthermore, vitamin C reversed the increased ROS levels, the decreased ATP levels, and the accelerated accumulation of the progeroid marker lipofuscin in both mutant strains. Lipofuscin is believed to be a mix of oxidized and cross-linked macromolecules, including proteins, lipids, and carbohydrates [45]. Such results point to metabolic abnormalities in worms lacking the helicase function of the human WRN ortholog like WrnΔhel/Δhel mice [16, 30, 34]. Importantly, we found that vitamin C reversed the metabolic abnormalities in both of these models.
To conclude, our data indicate that miR-124 is a conserved miRNA that is involved in the aging phenotype across mouse and worm species. Furthermore, the loss of miR-124 expression is associated with the lack of WRN helicase function in both species. Finally, the progeroid phenotypes associated with either WRN or miR-124 mutations can be reversed by vitamin C treatment. Finally, our results with both mouse [16] and worm models of WS suggest that vitamin C supplementation could have beneficial effects for patients with WS.
Methods
MicroRNA expression profiling
Care of mice was in accordance with the guidelines of the Centre de Recherche des Centres Hospitaliers Universitaires de Québec. The TaqMan® Array Rodent MicroRNA Card Set v3.0 is a two card set containing a total of 384 TaqMan® MicroRNA Assays per card. The set enables accurate quantification of 755 unique microRNAs for mouse. Included on each array is three TaqMan® MicroRNA Assay endogenous controls to aid in data normalization and one TaqMan® MicroRNA Assay not related to rodent as a negative control. Use of the Megaplex™ RT Primers, Rodent Pool Set v3.0 was required to run the array sets. An additional preamplification step was carried out with Megaplex™ PreAmp Primers. Reactions were performed on four animals, two for each genotype, and according to the manufacturer's protocol (Applied Biosystems, Carlsbad, CA). Raw CTs were then successively normalized using the endogenous U6 and quantile normalization. An empirical Bayesian method within the package limma in BioConductor (http://www.bioconductor.org) was used to identify the significantly modulated miRNAs. A miRNA was judged significantly modulated if the Benjamini-Hochberg adjusted P-value was lower than 0.1. All miRNA analyses were performed using R version 2.14.0.
Validation of miRNA expression
The quantitative measure of selected miRNA expressions was performed with TaqMan MicroRNA assays on extracted total RNA from four different WrnDhel/Dhelmutant and four wild type mice or on extracted total RNA from four young (three months) and four old (21 months) wild type mice following manufacturer's protocol (Life Techonology, USA).
Caenorhabditis elegans strains
All C. elegans strains were maintained as described [46]. Both wrn-1(gk99) and mir-124(n4255) strains obtained from the C. elegans Genetics Center (University of Minnesota, St Paul, MN) were out-crossed four times with the wild type N2 strain to remove possible unrelated mutations. The wrn-1(gk99) contains a 196 bps deletion that inhibits the expression of the protein [32]. The primers used to genotype this strain are 5’-CTGGCTGTAACT GCACCTGA-3’ and 5’-AAATGGGAGGGAAAGAGC AT-3’. The mir-124(n4255) strain contains a 212 bps deletion that spans the entire mir-124 sequence. The mir-124 sequence is localized in an intron of the trpa-1 gene. It has been shown that the n4255 deletion does not abrogate the expression of the trpa-1 gene in C. elegans [39]. The primers used to genotype the mir-124(n4255) strain are 5’-TTGCTTCTTCTTCGAGCA CA-3’ and 5’-AAATGGGAGGGAAAGAGCAT-3’.
Expression of mir-124 in C. elegans
Three hundred 7-days old adult worms (post-larval L4 stage) were sorted by size to exclude remaining larvae using a COPAS BIOSORT instrument (Union Biometrica, Inc., Somerville, MA, USA). Sorted worms were spun down in an eppendorf tube and lysed in TRIZOL (Invitrogen, Carlsbad, CA) to extract total RNA. To measure mir-124 expression, TaqMan Small RNA assays (Applied Biosystems) were performed as described before. Stem-loop qRT-PCR for mature miRNAs was performed on a real-time PCR system (AB 7900; Applied Biosystems). The short nuclear RNA sn2841 were measured and used as an endogenous control.
Measurement of life span and aging markers
Worms were transferred to fresh plates and were grown at 25°C. Death was scored by absence of any movement after several light pokes with a platinum wire. Lipofuscin was detected as autofluorescence in adult worms and images were captured using a Zeiss motorized Axioplan 2 microscope (with 525 nm filter) equipped with an AxioCam MRm camera and the AxioVision acquisition software (Carl Zeiss Microscopy GmbH, Jena, Germany).
ATP quantification in C. elegans
ATP levels were quantified with the ApoSensor ATP assay kit according to the manufacturer's instruction (BioVision, Mountain View, CA). Luminescence was measured with a Luminoskan Ascent luminometer (Thermo Electron Inc., Milford, MA). Twenty-five 7-days old adult worms (post-larval L4 stage) worms were collected spun in an eppendorf tube and resuspended in 250 mL of assay kit buffer. Worms were crushed in a Dounce homogenizer (25 strokes) and the homogenate was spun 5 min at 13,000 rpm on a bench top centrifuge at room temperature. The ATP level was measured from the homogenate. Protein concentrations were measured using the Bradford assay. Results were expressed as amount of ATP/mg of proteins. All experiments were performed in three independent pools of animals.
Reactive oxygen species (ROS) quantification in C. elegans
ROS quantification was performed on twenty-five 7-days old adult worms (post-larval L4 stage). Worms were collected spun in an eppendorf tube, resuspended in 250 mL of RIPA buffer (50 mM Tris HCl (pH 7.5), 150 mM NaCl, 1% NP-40, 0.1% SDS, 0.5% deoxycholate), crushed in a Dounce homogenizer (25 strokes). The homogenate (150 μl) was incubated with 10 μg/ml of the dye 2’-7’ dichlorofluorescein diacetate (Sigma-Aldrich) for one hour at 37°C. This dye is highly fluorescent upon oxidation. The oxidized dye was measured as described previously [34]. Protein concentrations were measured using the Bradford assay.
Statistical analysis
Data on graphs are presented as means + SD. The unpaired Student's t-test and the log-rank test were all performed using an alpha level of 0.05 and a two-sided hypothesis. Life span curves were build on differences between strains were considered significant at P-value lower than 0.05 in all statistical analyses. All statistical analyses were performed using R version 2.14.0 (www.r-project.org).
Supplementary Materials
Acknowledgments
We would like to thank Dr. Patrick Laprise (Centre de Recherche en Cancérologie de l'Université Laval, Canada) for allowing us to use his COPAS Biosort instrument. Nematode strains were provided by the Caenorhabditis Genetics Center, which is funded by the National Institute of Health National Center for Research Resources. This work was supported by the Canadian Institutes of Health Research to M.L. and M.J.S. R.d.C. is supported by the Intramural Research Program of the National Institute on Aging of the National Institutes of Health, USA. S.J.M. is supported by a National Health and Medical Research Council of Australia CJ Martin Early Career Fellowship (RIMS Project ID 2010-01671). M.J.S. is a Junior II scholar from the Fonds de la Recherche en Santé du Québec.
Conflicts of Interest
The authors of this manuscript have no conflict of interests to declare.
References
- 1. Kipling D, Davis T, Ostler EL, Faragher RG. What can progeroid syndromes tell us about human aging? Science. 2004; 305: 1426 -1431. [PubMed] .
- 2. Epstein CJ, Martin GM, Schultz AL, Motulsky AG. Werner's syndrome a review of its symptomatology, natural history, pathologic features, genetics and relationship to the natural aging process. Medicine (Baltimore). 1966; 45: 177 -221. [PubMed] .
- 3. Salk D. Werner's syndrome: a review of recent research with an analysis of connective tissue metabolism, growth control of cultured cells, and chromosomal aberrations. Hum Genet. 1982; 62: 1 -5. [PubMed] .
- 4. Yu CE, Oshima J, Fu YH, Wijsman EM, Hisama F, Alisch R, et al. Positional cloning of the Werner's syndrome gene. Science. 1996; 272: 258 -262. [PubMed] .
- 5. Ozgenc A and Loeb LA. Current advances in unraveling the function of the Werner syndrome protein. Mutat Res. 2005; 577: 237 -251. [PubMed] .
- 6. Kamath-Loeb AS, Shen JC, Loeb LA, Fry M. Werner syndrome protein. II. Characterization of the integral 3’ → 5’ DNA exonuclease. J Biol Chem. 1998; 273: 34145 -34150. [PubMed] .
- 7. Shen JC, Gray MD, Oshima J, Kamath-Loeb AS, Fry M, Loeb LA. Werner syndrome protein. I. DNA helicase and dna exonuclease reside on the same polypeptide. J Biol Chem. 1998; 273: 34139 -34144. [PubMed] .
- 8. Huang S, Beresten S, Li B, Oshima J, Ellis NA, Campisi J. Characterization of the human and mouse WRN 3’-->5’ exonuclease. Nucleic Acids Res. 2000; 28: 2396 -2405. [PubMed] .
- 9. Balajee AS, Machwe A, May A, Gray MD, Oshima J, Martin GM, et al. The Werner syndrome protein is involved in RNA polymerase II transcription. Mol Biol Cell. 1999; 10: 2655 -2668. [PubMed] .
- 10. Cooper MP, Machwe A, Orren DK, Brosh RM, Ramsden D, Bohr VA. Ku complex interacts with and stimulates the Werner protein. Genes Dev. 2000; 14: 907 -912. [PubMed] .
- 11. Shen JC and Loeb LA. The Werner syndrome gene: the molecular basis of RecQ helicase-deficiency diseases. Trends Genet. 2000; 16: 213 -220. [PubMed] .
- 12. Saintigny Y, Makienko K, Swanson C, Emond MJ, Monnat RJ Jr.. Homologous recombination resolution defect in werner syndrome. Mol Cell Biol. 2002; 22: 6971 -6978. [PubMed] .
- 13. Crabbe L, Verdun RE, Haggblom CI, Karlseder J. Defective telomere lagging strand synthesis in cells lacking WRN helicase activity. Science. 2004; 306: 1951 -1953. [PubMed] .
- 14. Lebel M and Leder P. A deletion within the murine Werner syndrome helicase induces sensitivity to inhibitors of topoisomerase and loss of cellular proliferative capacity. Proc Natl Acad Sci U S A. 1998; 95: 13097 -13102. [PubMed] .
- 15. Lebel M, Lavoie J, Gaudreault I, Bronsard M, Drouin R. Genetic cooperation between the Werner syndrome protein and poly(ADP-ribose) polymerase-1 in preventing chromatid breaks, complex chromosomal rearrangements, and cancer in mice. Am J Pathol. 2003; 162: 1559 -1569. [PubMed] .
- 16. Massip L, Garand C, Paquet ER, Cogger VC, O'Reilly JN, Tworek L, et al. Vitamin C restores healthy aging in a mouse model for Werner syndrome. FASEB J. 2010; 24: 158 -172. [PubMed] .
- 17. Lee SJ, Yook JS, Han SM, Koo HS. A Werner syndrome protein homolog affects C. elegans development, growth rate, life span and sensitivity to DNA damage by acting at a DNA damage checkpoint. Development. 2004; 131: 2565 -2575. [PubMed] .
- 18. Hyun M, Bohr VA, Ahn B. Biochemical characterization of the WRN-1 RecQ helicase of Caenorhabditis elegans. Biochemistry. 2008; 47: 7583 -9753. [PubMed] .
- 19. Ibanez-Ventoso C, Yang M, Guo S, Robins H, Padgett RW, Driscoll M. Modulated microRNA expression during adult lifespan in Caenorhabditis elegans. Aging Cell. 2006; 5: 235 -246. [PubMed] .
- 20. Ibanez-Ventoso C and Driscoll M. MicroRNAs in C. elegans Aging: Molecular Insurance for Robustness? Curr Genomics. 2009; 10: 144 -153. [PubMed] .
- 21. Bates DJ, Liang R, Li N, Wang E. The impact of noncoding RNA on the biochemical and molecular mechanisms of aging. Biochim Biophys Acta. 2009; 1790: 970 -979. [PubMed] .
- 22. Grillari J and Grillari-Voglauer R. Novel modulators of senescence, aging, and longevity: Small non-coding RNAs enter the stage. Exp Gerontol. 2010; 45: 302 -311. [PubMed] .
- 23. Smith-Vikos T and Slack FJ. MicroRNAs and their roles in aging. J Cell Sci. 2012; 125: 7 -17. [PubMed] .
- 24. Maes OC, An J, Sarojini H, Wang E. Murine microRNAs implicated in liver functions and aging process. Mech Ageing Dev. 2008; 129: 534 -541. [PubMed] .
- 25. Hamrick MW, Herberg S, Arounleut P, He HZ, Shiver A, Qi RQ, et al. The adipokine leptin increases skeletal muscle mass and significantly alters skeletal muscle miRNA expression profile in aged mice. Biochem Biophys Res Commun. 2010; 400: 379 -383. [PubMed] .
- 26. Li N, Bates DJ, An J, Terry DA, Wang E. Up-regulation of key microRNAs, and inverse down-regulation of their predicted oxidative phosphorylation target genes, during aging in mouse brain. Neurobiol Aging. 2011; 32: 944 -955. [PubMed] .
- 27. Li N, Muthusamy S, Liang R, Sarojini H, Wang E. Increased expression of miR-34a and miR-93 in rat liver during aging, and their impact on the expression of Mgst1 and Sirt1. Mech Ageing Dev. 2011; 132: 75 -85. [PubMed] .
- 28. Williams AE, Perry MM, Moschos SA, Lindsay MA. microRNA expression in the aging mouse lung. BMC Genomics. 2007; 8: 172 [PubMed] .
- 29. Izzotti A, Calin GA, Steele VE, Croce CM, De Flora S. Relationships of microRNA expression in mouse lung with age and exposure to cigarette smoke and light. FASEB J. 2009; 23: 3243 -3250. [PubMed] .
- 30. Labbe A, Garand C, Cogger VC, Paquet ER, Desbiens M, Le Couteur DG, et al. Resveratrol improves insulin resistance hyperglycemia and hepatosteatosis but not hypertriglyceridemia, inflammation, and life span in a mouse model for Werner syndrome. J Gerontol A Biol Sci Med Sci. 2011; 66: 264 -278. [PubMed] .
- 31. Le Couteur DG, Warren A, Cogger VC, Smedsrod B, Sorensen KK, De Cabo R, et al. Old age and the hepatic sinusoid. Anat Rec (Hoboken). 2008; 291: 672 -683. [PubMed] .
- 32. Lee SJ, Gartner A, Hyun M, Ahn B, Koo HS. The Caenorhabditis elegans Werner syndrome protein functions upstream of ATR and ATM in response to DNA replication inhibition and double-strand DNA breaks. PLoS Genet. 2010; 6: e1000801 [PubMed] .
- 33. Miska EA, Alvarez-Saavedra E, Abbott AL, Lau NC, Hellman AB, McGonagle SM, et al. Most Caenorhabditis elegans microRNAs are individually not essential for development or viability. PLoS Genet. 2007; 3: e215 [PubMed] .
- 34. Massip L, Garand C, Turaga RV, Deschenes F, Thorin E, Lebel M. Increased insulin, triglycerides, reactive oxygen species, and cardiac fibrosis in mice with a mutation in the helicase domain of the Werner syndrome gene homologue. Exp Gerontol. 2006; 41: 157 -168. [PubMed] .
- 35. Huang J and Lemire BD. Mutations in the C. elegans succinate dehydrogenase iron-sulfur subunit promote superoxide generation and premature aging. Journal of molecular biology. 2009; 387: 559 -569. [PubMed] .
- 36. Bates DJ, Li N, Liang R, Sarojini H, An J, Masternak MM, et al. MicroRNA regulation in Ames dwarf mouse liver may contribute to delayed aging. Aging Cell. 2010; 9: 1 -18. [PubMed] .
- 37. Khanna A, Muthusamy S, Liang R, Sarojini H, Wang E. Gain of survival signaling by down-regulation of three key miRNAs in brain of calorie-restricted mice. Aging (Albany NY). 2011; 3: 223 -236. [PubMed] .
- 38. Cheng LC, Pastrana E, Tavazoie M, Doetsch F. miR-124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nat Neurosci. 2009; 12: 399 -408. [PubMed] .
- 39. Clark AM, Goldstein LD, Tevlin M, Tavare S, Shaham S, Miska EA. The microRNA miR-124 controls gene expression in the sensory nervous system of Caenorhabditis elegans. Nucleic Acids Res. 2010; 38: 3780 -3793. [PubMed] .
- 40. Baroukh N, Ravier MA, Loder MK, Hill EV, Bounacer A, Scharfmann R, et al. MicroRNA-124a regulates Foxa2 expression and intracellular signaling in pancreatic beta-cell lines. J Biol Chem. 2007; 282: 19575 -19588. [PubMed] .
- 41. Lovis P, Gattesco S, Regazzi R. Regulation of the expression of components of the exocytotic machinery of insulin-secreting cells by microRNAs. Biol Chem. 2008; 389: 305 -312. [PubMed] .
- 42. Hatziapostolou M, Polytarchou C, Aggelidou E, Drakaki A, Poultsides GA, Jaeger SA, et al. An HNF4alpha-miRNA inflammatory feedback circuit regulates hepatocellular oncogenesis. Cell. 2011; 147: 1233 -1247. [PubMed] .
- 43. Friedman DB and Johnson TE. A mutation in the age-1 gene in Caenorhabditis elegans lengthens life and reduces hermaphrodite fertility. Genetics. 1988; 118: 75 -86. [PubMed] .
- 44. Kenyon C, Chang J, Gensch E, Rudner A, Tabtiang R. A C. elegans mutant that lives twice as long as wild type. Nature. 1993; 366: 461 -464. [PubMed] .
- 45. Yin D. Biochemical basis of lipofuscin, ceroid, and age pigment-like fluorophores. Free radical biology & medicine. 1996; 21: 871 -888. [PubMed] .
- 46. Brenner S. The genetics of Caenorhabditis elegans. Genetics. 1974; 77: 71 -94. [PubMed] .