


Introduction
Studies from different model organisms have shown that nutrient sensing factors or pathways [1, 2] such as AMP-activated kinase (AMPK) [3], insulin-IGF signaling (IIS) [4, 5], target of rapamycin (TOR) [6] and Sirtuins (Sir2-like) [7, 8] are key players in maintaining metabolic homeostasis. Further, the response of an organism to a metabolic stress, such as starvation, is largely dependent on its capacity to derive energy from stored reserves like triglyceride (lipid) [9, 10] and glycogen (glucose) [11, 12]. Genetic perturbations that impinge on lipid metabolism have been implicated as one of the causes of metabolic deregulation, and often associated with obesity and type II diabetes [13-16].
Sirtuins or Sir2-like proteins are NAD+-dependent enzymes that link cellular physiology with metabolic changes [7, 17]. Sir2 was identified as a key determinant of longevity in Saccharomyces cerevisiae and its dependence on NAD+ for its activity is crucial for its ability to link calorie/dietary restriction (CR/DR) with lifespan extension [18, 19]. Although, initial observations in Caenorhabditis elegans [20] and Drosophila melanogaster [21] indicated the evolutionary significance of Sir2 in extending lifespans, recent reports have questioned these findings in metazoans [22, 23, 24]. More importantly, in the absence of a clear understanding of the evolutionary conservation of molecular functions of Sir2 in metazoans, its roles in regulating organismal physiology and survival are still not well appreciated. In this regard, studies that dissect out molecular functions of Sir2 will provide insights into its role in response to calorie restriction and aging.
Although, it is intuitive to expect Sir2 to maintain metabolic homeostasis, across species, only mammalian SIRT1 has been clearly shown to do so [17, 25, 26]. Cell culture studies clearly indicated the role of SIRT1 in glucose and fat metabolism and recent papers have highlighted its importance in metabolically relevant tissues such as the liver and muscles [26-36]. Further, the contributions of SIRT1 functions in different tissues in maintaining organismal physiology are being appreciated only recently [36, 37]. However, it is still unclear if these functions of SIRT1 are evolutionarily conserved in lower organisms. Therefore, there is a need to study the roles of Sir2 orthologs in mediating systemic changes, and those emanating from metabolic tissues, to provide a comprehensive understanding of Sir2 biology. Although, studies in mammals provide clear insights into the ability of SIRT1 in metabolically relevant tissues to affect organismal physiology [36, 37], the relevance under altered nutrient conditions is not known. Importantly, it is unclear whether the ability of Sir2 in different tissues to maintain metabolic homeostasis has any bearing on organismal survival.
The fruit fly Drosophila melanogaster has been successfully used for studying many molecular factors that link dietary or metabolic changes to organismal physiology [38]. The molecular mechanisms downstream to dSir2 and its ability to regulate metabolism are poorly appreciated in flies. dSir2 was previously shown to genetically interact with rpd3 (histone deacetylase) and mediate lifespan extension [21]. However, this does not provide a functional link with the metabolic functions (if any) of dSir2 [39]. dSir2 has also been shown to interact with Dmp53 and is implicated in p53-dependent functions [40, 41]. Except for a very recent study in larvae, its role in regulating metabolism in adult flies is still unclear [42]. Importantly, the tissue specific role of dSir2 in modulating metabolism and therefore, organismal physiology has not been addressed in adult flies. Given the tissue complexity and conservation of metabolic pathways, Drosophila is a useful system to investigate dSir2-dependent alterations in molecular mechanisms, which might have a bearing on organismal survival.
Here, we report the central role of endogenous dSir2 in maintaining metabolic and energy homeostasis. In addition to using backcrossed dSir2 mutant flies, we have knocked down dSir2 using gene-switch lines that negate background genetic differences (between control and dSir2 knockdown flies). We show that dSir2 plays a crucial role in fat metabolism and systemic insulin signaling. By knocking down dSir2 in the muscles and fatbody, we have compared its functions in these tissues to maintain metabolic homeostasis and mediate organismal survival in response to starvation. These findings not only establish Drosophila as a useful system to investigate Sir2 functions, but also show that dSir2 is a critical factor in fat mobilization from the fatbody during starvation. Our results highlight the key role that dSir2 plays in the adult fatbody in bringing about systemic changes in physiology that determine the ability of an organism to cope with metabolic stress. Importantly, contrasting effects of dSir2 deficiency in the fatbody and muscles signify the tissue dependent roles of dSir2 in organismal survival.
Results
dSir2 affects starvation survival in flies
Starvation, unlike other dietary regimes, is an acute manipulation, which induces nutrient sensing pathways [46]. In addition, it reflects the ability of an organism to maintain metabolic and energy homeostasis [46]. Although, SIRT1 in mammals has been shown to respond to starvation and mediate transcription of genes involved in metabolic homeostasis [27, 34, 47], such a role for Sir2 in invertebrates is not known. Moreover, it is still unclear if absence of Sir2/SIRT1 affects starvation resistance. In order to study the importance of Drosophila Sir2 (dSir2) in maintaining metabolic homeostasis, we subjected the flies to nutrient deprivation (starvation). We found that backcrossed dSir2 mutant (Sir22A.7.11) flies (Supplementary Figure 1) were more sensitive to starvation when compared to the controls (Figure 1A and Supplementary Figure 2). In order to confirm this and also negate any potential background effects, we knocked down dSir2 ubiquitously using the inducible pSwitch-tub-gal4 driver (Supplementary Figure 1). Knocking down dSir2 (+ RU486) also led to decreased starvation survival similar to the mutant flies (Figure 1B and Supplementary Figure 2).
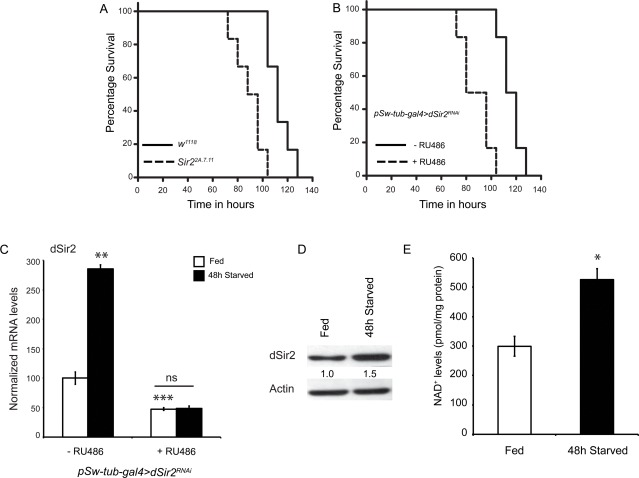
Figure 1. dSir2 activity increases in response to starvation and its absence decreases starvation resistance (A) Starvation survival of dSir2 mutants (Sir22A.7.11) (p < 0.001) and (B) whole body dSir2RNAi (+ RU486) (p < 0.001), with respective controls (n = 60). (C)dSir2 transcript levels increase in response to 48-hours starvation in control (- RU486) but not in whole body dSir2RNAi(+ RU486) flies (n = 8/24). (D) Increase in dSir2 protein levels in control flies in response to 48-hours starvation. (E) Increase in NAD+ levels in response to 48-hours starvation in control flies (n = 36). 200 μM RU486 was used to knockdown dSir2 expression inpSw-tub-gal4>dSir2RNAi flies. Log Rank was used to plot survival curves and Mantel-Cox test was used for statistical analysis. Student's t-test and ANOVA were used to analyze statistical significance of the data (*, p < 0.05; **, p < 0.01; ***, p < 0.001 or mentioned otherwise).
SIRT1 activity and expression have been shown to increase during starvation in mammals [48, 49]. To check if starvation resistance in control flies involved an upregulation of dSir2 expression, we assayed for its transcript and protein levels. dSir2 expression, both mRNA and protein, was induced in response to starvation in control flies (Figures 1C and 1D). It is important to note that dSir2RNAi (+ RU486) flies did not show any increase in dSir2 transcript levels (Figure 1C) upon starvation.
The activity of Sir2-like proteins has been demonstrated to be dependent upon intracellular NAD+ levels. Across organisms, increased NAD+ levels have been associated with increased activity and functions of Sir2 proteins [50-54]. To check if in addition to induction of dSir2 expression, there was an increase in NAD+ levels during starvation, we measured NAD+ in fed and starved, control and dSir2RNAi (+ RU486) flies (Figure 1E and Supplementary Figure 2). We found that NAD+ levels increased by 1.8 folds in response to starvation, similarly, in control and dSir2RNAi(+ RU486) flies (Figure 1E and Supplementary Figure 2). These results indicate that dSir2 expression and activity, in vivo, increases during starvation in flies.
dSir2 in the fatbody regulates metabolic homeostasis in flies
Abrogated starvation resistance indicated metabolic defects in flies that lacked dSir2. To appreciate the importance of dSir2 in maintaining metabolic homeostasis, total glucose and total triglyceride levels were measured in control and backcrossed dSir2 mutant flies (Sir22A.7.11). Interestingly, we found that both were significantly higher in dSir2 mutants (Sir22A.7.11) (Figure 2A and Supplementary Figure 3A). The deregulated fat metabolism and increased triacylglycerol (TAG) levels were corroborated by oil red staining of fatbodies (Figure 2B) in these flies. In order to check if dSir2 was also responsible for regulating metabolism under altered dietary conditions, we fed the control and mutant flies with diets containing variable yeast contents (0.25%, 2.5% and 5.0% with 5.0% sucrose and 8.6% cornmeal). It is important to note that irrespective of the dietary regime, flies that lackeddSir2 exhibited increased glucose and triglyceride levels (Supplementary Figures 3B and 3C).
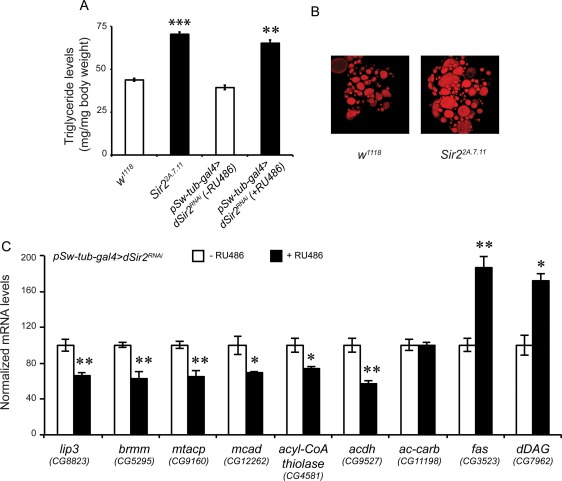
Figure 2. dSir2 regulates fat metabolism (A) Total body triglyceride (TAG) in dSir2 mutants (Sir22A.7.11) and whole body dSir2RNAi (+ RU486) flies, with respective controls (n = 36). (B) Oil-Red O staining of fatbodies from control and Sir22A.7.11. (C) Relative expression of fat metabolism genes lipase-3 (lip3), brummer (brmm), mitochondrial acyl carrier protein (mtACP), medium chain acyl CoA dehydrogenase (mcad), aceto acetyl CoA thiolase (ACoT), long chain acyl CoA dehydrogenase (lcad), acetyl CoA carboxylase (ACC), fatty acid synthase (fas) and diacyl glycerol synthase (dDAG) (n = 24). 200 μM RU486 was used to knockdown dSir2 expression in pSw-tub-gal4>dSir2RNAi flies. Student's t-test was used to analyze statistical significance of the data (*, p < 0.05; **, p < 0.01; ***, p < 0.001 or mentioned otherwise).
When glucose levels were assayed in whole body dSir2 knockdown flies, we observed decreased levels of glucose, contrary to what was found in the dSir2 mutant flies (Supplementary Figure 3A). This difference could be due to the complete absence of dSir2 expression in the mutants, as against a reduced expression in the RNAi flies (Supplementary Figure 1). Importantly, we saw an increase in the TAG levels when dSir2 was knocked down in the whole body and this effect was consistent with the mutants (Figures 2A and 2B). We did not observe a significant difference in the weights of these flies (Supplementary Figure 3D). To confirm the role of dSIr2 in regulating metabolic output in a bidirectional manner, we overexpressed dSir2 using pSw-tub-gal4>Sir2EP2300. The flies overexpressing dSir2 exhibited decreased TAG levels (Supplementary Figure 4A). These results clearly showed that an absence of dSir2 leads to metabolic defects, specifically of fat metabolism, under basal conditions and high nutrient conditions.
To investigate the underlying molecular mechanism that renders this phenotype, we quantified the expression of genes involved in fat metabolism by RT-qPCR analyses. From Figure 2C it is evident that in dSir2RNAiflies (+ RU486), the transcript levels of brummer (lipase ATGL), lipase-3, medium chain acyl-CoA dehydrogenase (MCAD), mitochondrial acyl carrier protein (mtACP), aceto-acetyl-CoA-thiolase (ACoT) and long chain acyl-CoA-dehydrogenase (LCAD) are decreased. We also observed that expression of fatty acid synthase (fas) and diacyl glycerol synthetase (dDAG) were upregulated in dSir2RNAi flies (+ RU486) (Figure 2D). Corroborating the importance of dSir2 in the expression of these genes, we observed the opposite effects in the dSir2 overexpression flies (Supplementary Figure 4).
In order to check if the accumulation of TAGs was due to a failure in mobilization or utilization, we knocked down dSir2 in a tissue specific manner. While muscles utilize fat, fatbodies are considered equivalent to mammalian liver and adipose tissue. We knocked down dSir2 in the fatbody using the specific pSw-S1106-gal4 driver, which has previously been reported to be expressed in abdominal and head fatbodies [55, 56]. To knockdown dSir2 in the muscles, we employed the muscles specific pSwitch-MHC-gal4 driver [57]. Figure 3A (and Supplementary Figure 5) clearly show tissue specific knockdown of dSir2 in these flies. It is very interesting to note that fatbody knockdown of dSir2 led to an increase in TAG levels and mimicked the whole body dSir2RNAi (+ RU486) flies (Figure 2A). Surprisingly, knocking down dSir2 in the muscles did not affect TAG levels (Figure 3C), suggesting the importance of dSir2 in fat mobilization.
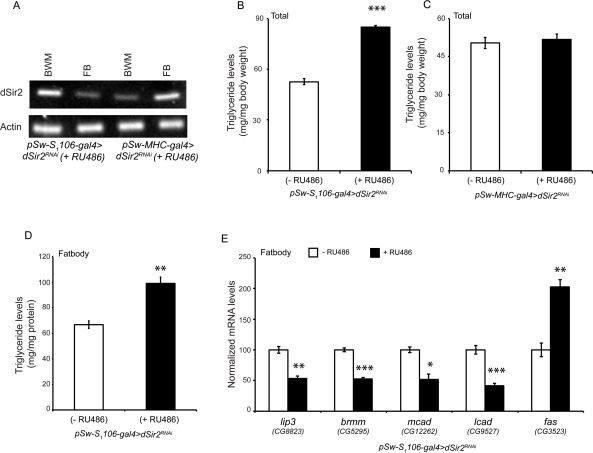
Figure 3. dSir2 in the fatbody, but not in the muscles regulates fat metabolism (A) RT-PCR to show knockdown of dSir2 in the fatbody (FB) of fatbody dSir2RNAiand body wall with muscles (BWM) of muscle dSir2RNAi, with respective controls (n = 24). (B-C) Total body triglyceride levels in (B) fatbody dSir2RNAi flies and (C) muscles dSir2RNAi flies (n = 36). (D) Triglyceride levels in the isolated fatbody of fatbody dSir2RNAi flies (n = 60). (E) Relative expression of fat metabolism genes lipase-3 (lip3), brummer (brmm), medium chain acyl CoA dehydrogenase (mcad), long chain acyl CoA dehydrogenase (lcad) and fatty acid synthase (fas) in fatbody isolated from fatbody dSir2RNAi flies (n = 24). 200 μM RU486 was used to knockdown dSir2 expression pSw-S1106-gal4>dSir2RNAi and pSw-MHC-gal4> dSir2RNAi flies. Student's t-test was used to analyze statistical significance of the data (*, p < 0.05; **, p < 0.01; ***, p < 0.001 or mentioned otherwise).
These results indicated that in the absence of dSir2 the defects in fat metabolism originate in the fatbodies. The importance of dSir2 in this tissue was confirmed when we observed increased TAG levels in fatbodies isolated from fatbody dSir2RNAi (+ RU486) flies (Figure 3B). This increase was also corroborated by reduced expression of fat breakdown genes and an increase in fatty acid synthase (Figure 3E) in the fatbody. These findings show that dSir2 is important for metabolic homeostasis, and highlight its role in the adult fatbody in regulating fat metabolism. To our knowledge, this is the first study linking dSir2 to metabolic homeostasis in adult flies, and with the recent findings in larvae [42], highlights its role in regulating fat metabolism in invertebrates. Importantly, from our results, and studies on mammalian SIRT1 [29, 33, 58] an evolutionarily conserved role for Sir2 (dSir2/SIRT1) in regulating fat metabolism becomes obvious.
Knockdown of dSir2 in the fatbody, but not in muscles, affects starvation survival
Based on the molecular results, we wanted to check if knocking down dSir2 in specific tissues, such as fatbody and muscles, affected starvation survival. Control and dSir2RNAi (+ RU486) flies, in both muscles and fatbodies, were assayed for starvation survival. Our results clearly demonstrate that knocking down dSir2 in the fatbody phenocopied (Figure 4A and Supplementary Figure 6) the ubiquitous dSir2 knockdown (Figure 1B) and these flies succumbed to starvation earlier than the controls. Interestingly, subjecting pSwitch-MHC-gal4 (+ RU486) flies to starvation phenocopied control flies (Figure 4B). These results clearly delineate the functions of dSir2 in the two different metabolic tissues in affecting starvation survival.
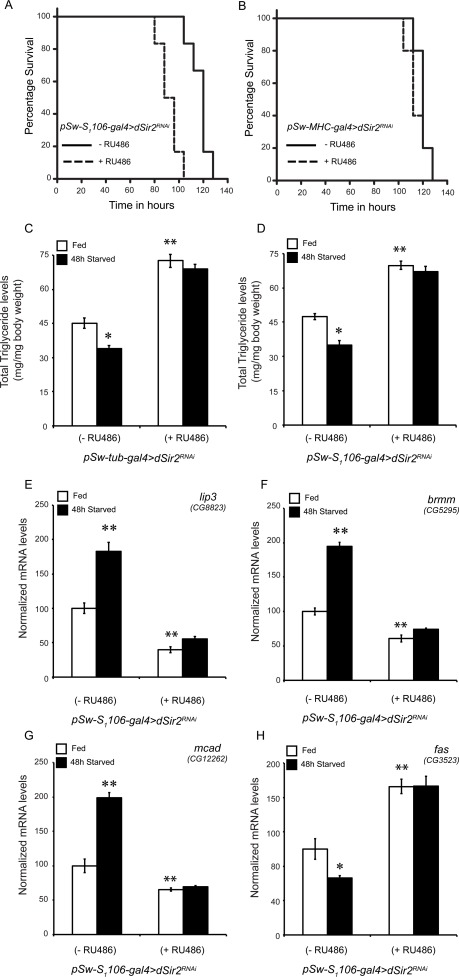
Figure 4. Fatbody dSir2, but not muscle specific dSir2 regulates starvation survival and regulates fat mobilization (A and B) Starvation survival in (A) fatbody dSir2 knockdown flies (p < 0.001) and (B) muscle dSir2 knockdown flies (non-significant, ns)(n = 60). 200 μM RU486 was used to knockdown dSir2 expression pSw-S1106-gal4>dSir2RNAi and pSw-MHC-gal4> dSir2RNAi flies. Log Rank was used to plot survival curves and Mantel-Cox test was used for statistical analysis. (C-D) Total body triglyceride levels in fed and starved conditions in (C) whole bodydSir2RNAiflies, with respective controls (n = 36) and (D) fatbody dSir2RNAi (n = 36). Relative expression of fat metabolism genes (E) lipase-3 (lip3), (F) brummer (brmm), (G) medium chain acyl CoA dehydrogenase (mcad), (H) fatty acid synthase (fas) in fatbody dSir2 knockdown flies under fed and starved conditions (n = 24). 200 μM RU486 was used to knockdown dSir2 expression pSw-S1106-gal4>dSir2RNAi and pSw-MHC-gal4> dSir2RNAi flies. ANOVA was used to analyze statistical significance of the data (*, p < 0.05; **, p < 0.01; ***, p < 0.001 or mentioned otherwise).
Failure to mobilize energy reserves affects starvation survival in fatbody dSir2 knockdown flies
We were intrigued to find that, although, dSir2-RNAi and -mutant flies had increased fat storage (Figures 2A and 2B), these flies survived less upon starvation. Tissue specific knockdown of dSir2 in the fatbody and the muscles indicated that these flies had defective fat mobilization and not utilization (Figures 3B to 3D). To check if this was true, we analyzed TAG levels and genes involved in fat metabolism in response to starvation. While control flies showed a significant decrease in TAG levels after 48-hour starvation, whole body dSir2RNAi (+ RU486) flies had elevated TAG levels and were comparable to their fed condition (Figure 4C). Importantly, the same effect was observed when dSir2 was knocked down specifically in the fatbody (Figure 4D). Corroborating these results, we found that the expression of genes involved in fat breakdown increased in control flies during starvation, but not in fatbody dSir2RNAi (+ RU486) flies (Figures 4E to 4G). Surprisingly, we also observed an increase in the expression of fatty acid synthase in fatbody dSir2 knockdown flies under fed and starved conditions (Figure 4H).
Fatbody specific knockdown of dSir2 affects insulin signaling
Insulin-IGF signaling (IIS) is another important nutrient sensing pathway that has been shown to mediate metabolic homeostasis and also affect organismal survival [59, 60]. Therefore, we wanted to check if dSir2RNAi (+ RU486) flies displayed abrogated IIS. Expression of dilp (insulin-like-peptide) genes has been documented to reflect changes in IIS under fed and starved conditions [61]. Using RNA isolated from fly heads, RT-qPCR analyses were performed to quantify the expression dilp-2, -3 and -5 from pSw-tub-gal4> dSir2RNAi (+ RU486) flies. We found that only dilp-2 and -5 levels were substantially altered in these flies (Figure 5A and Supplementary Figures 7A and 7B). However, it should be noted that the decrease in dilp5 expression that was observed during starvation in control flies [61] was absent in dSir2RNAi (+ RU486) flies (Figure 5A). We also did not see any changes in the expression of dilp6 levels in the fatbody of control and dSir2RNAi (+ RU486) flies (Supplementary figure 7C).
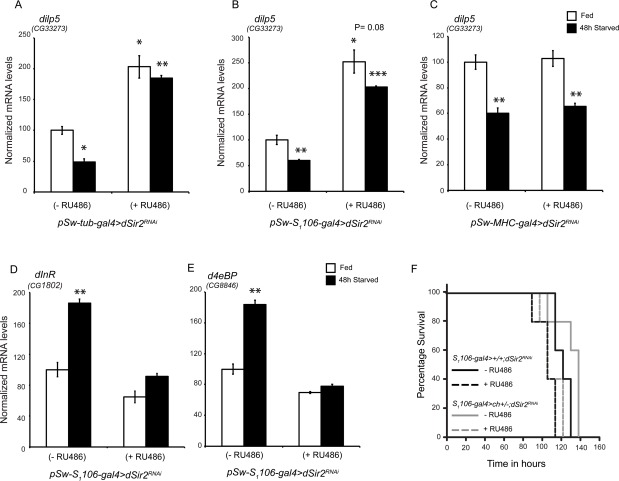
Figure 5. Fatbody dSir2 regulates dilp5 meditated Insulin Signaling (A-C) Relative dilp5 levels in the heads of (A) whole body dSir2RNAi (B) fatbody dSir2RNAi and (C) muscle dSir2RNAi flies, with respective controls, under fed and starved conditions (n = 24). Relative expression of (D) dInR and (E) d4eBP in fatbody dSir2RNAi flies under fed and starved conditions (n = 24). (F) Starvation survival of fatbody dSir2 knockdown (pSw-S1106-gal4>+/+;dSir2RNAi + RU486) and control (pSw-S1106-gal4>+/+;dSir2RNAi - RU486) flies, fatbody dSir2 knockdown in chico heterozygote flies (pSw-S1106-gal4>ch+/-; dSir2RNAi + RU486) and (pSw-S1106-gal4>ch+/-; dSir2RNAi- RU486) (statistical significance indicated in supplementary figure 9) (n = 60).200 μM RU486 was used to knockdown dSir2 expression. Log Rank was used to plot survival curves and Mantel-Cox test was used for statistical analysis. Student's t-test and ANOVA were used to analyze statistical significance of the data (*, p < 0.05; **, p < 0.01; ***, p < 0.001 or mentioned otherwise).
Next, we wanted to investigate the contributions of dSir2 activity in the fatbody and muscles in regulating IIS. Interestingly, we found that while knocking down dSir2 in the fatbody led to an increase in dilp5 expression, dSir2 knock down in muscles did not have any effect (Figures 5B and 5C). Further, we observed that dilp5 expression was reduced during starvation in muscle specificdSir2RNAi (+ RU486) flies similar to controls (Figure 5C), as expected. In contrast, the transcript levels of dilp5 remained high in fatbody specific dSir2RNAi (+ RU486) flies after 48-hour starvation (Figure 5B).
Since increased dilp5 expression has been associated with an increase in IIS, we assayed for the expression of insulin receptor and the downstream target gene d4eBP in control and fatbody dSir2RNAi (+ RU486) flies. It is important to note that decreased expression of d4eBP is well characterized as a marker of signaling through IIS. When expression of these genes was assayed in fatbody specific dSir2RNAi (+ RU486) flies, we found that their levels were reminiscent of increased insulin signaling (Figures 5D and 5E).
In order to confirm the role of dSir2 in affecting systemic insulin signaling, we assayed for the expression of dilp5, dInR and d4eBP in pSw-tub-gal4>Sir2EP2300 flies. As shown in Supplementary Figure 8, overexpression of dSir2 resulted in decreased dilp5 mediated insulin signaling. Further, these results made us wonder if starvation sensitivity of fatbody dSir2RNAi (+ RU486) flies was due to increased insulin signaling. To specifically analyze this, we employed chico heterozygote flies which exhibit reduced insulin signaling. Importantly, these flies have been shown to outlive control flies when subjected to starvation [62, 63]. To address the effect of fatbody dSir2 knockdown in the background of chico (ch) mutation, we generated pSw-S1106-gal4>ch+/-; dSir2RNAi flies and administered them with RU486. As reported earlier, chico heterozygotes (pSw-S1106-gal4>ch+/-; dSir2RNAi without RU486) [62] displayed starvation resistance when compared to controls. Comparing median and maximal survival showed that flies pSw-S1106-gal4>ch+/-; dSir2RNAi with RU486 (chico heterozygotes + dSir2knockdown) were similar to pSw-S1106-gal4>ch+/-; dSir2RNAi with RU486 flies (only dSir2 knockdown) (Figure 5F and Supplementary Figure 9). Interestingly, we did not observe any significant effect of insulin signaling on starvation survival when dSir2 was knocked down in the fatbody. These results suggest that reduced starvation resistance in the absence of dSir2 expression is caused by an inability to mobilize energy reserves.
Discussion
Here, we report that dSir2 is a critical factor that regulates metabolic homeostasis and mediates organismal physiology (Figure 6). Using genetic tools (inducible RNAi) that negate background effects, we provide concrete results that highlight the importance of endogenous dSir2 in the whole body, and in metabolically relevant tissues, such as fatbody and muscle. Our findings point out the importance of nutrient signaling in eliciting dSir2-dependent molecular changes, which play an important role in tissue specific metabolic functions that affect systemic outputs in flies (Figure 6). By describing a metabolic phenotype in flies that lack dSir2, we not only reiterate that Drosophila can be used to study sirtuin biology, but also highlight the evolutionary conservation of dSir2/SIRT1 functions in regulating organismal physiology.
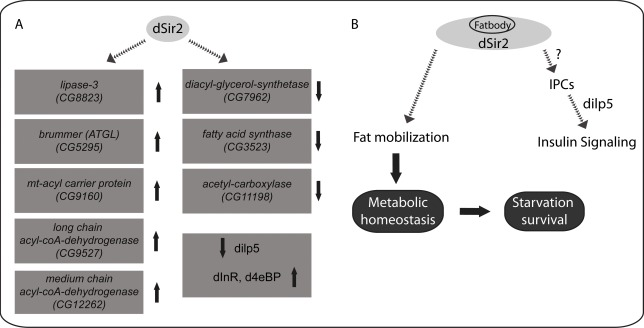
Figure 6. dSir2 regulates metabolic and energy homeostasis and its functions in the fatbody affect systemic insulin signaling
Until now, the conservation of molecular mechanisms underlying Sir2 biology was poorly addressed in invertebrates. It is only in mammals that a functional interplay between metabolic flux, SIRT1 and its downstream molecular factors has been addressed, thus far [17, 25, 26]. Results from backcrossed dSir2 mutant and whole body dSir2 knockdown flies indicated that absence or down-regulation of dSir2 expression results in gross metabolic defects (Figure 2). Interestingly, we observed that the effects on glucose levels were different in these two cases (Supplementary Figure 3A). The differences in glucose levels might reflect the systemic alterations in response to a complete absence of the protein in the case of mutants and down-regulation of expression in the case of knockdowns. It is interesting to note that studies in Sirt1+/-, liver specific Sirt1 knockout [36] and knockdown mice [27, 28] have also yielded seemingly conflicting. Specifically, with respect to glucose metabolism, these differences indicate that the manifestation of functions of Sir2/SIRT1 might be dependent upon the extent to which its expression is altered. Importantly, this underpins the need to further investigate the molecular interactions that bring about such varied phenotypes, in both mammals and flies.
It is important to note that we obtained consistent phenotypic, metabolic and molecular readouts with respect to fat metabolism in dSir2-mutant and -RNAi flies. We have found that a decrease (or absence) of dSir2 expression results in increased fat storage in the fatbodies, as determined by oil red staining and biochemical analyses (Figures 2 and 3). This fat accumulation is due to altered expression of genes involved in fat metabolism (Figures 2 and 3). Importantly, we show that genes involved in fat breakdown are downregulated in the dSir2 knockdown flies, in addition to an upregulation of genes involved in fat synthesis (Figures 2 and 3). These findings are not only in accordance with the results obtained from dSir2 mutant larvae [54] but also implicate dSir2 as a key player in fat metabolism in adult flies.
We have also uncovered a role for dSir2 in regulating systemic insulin signaling in flies. To investigate if the ability of dSir2 to mediate insulin signaling emanated from a specific tissue, we assayed for dilp5 expression in fatbody and muscle specific dSir2RNAi flies. Interestingly, we have found that knocking down dSir2 only in the fatbody, but not muscles led to increased dilp5 expression, and mimicked dSir2 mutants and whole body dSir2RNAi flies (Figure 5 and data not shown). Specifically, we have addressed the role of dSir2 in the fatbody to mediate systemic effects on insulin signaling (Figure 5 and Supplementary Figure 9). Further investigations should help us understand the dSir2-dependent molecular and physiological links between the fatbody and medial secretory neurons (MSNs). Very recently, hepatic SIRT1 was shown to mediate peripheral insulin signaling in mice [36]. Importantly, our findings underpin the importance of dSir2/SIRT1 in the homologous metabolic tissues, fatbody and liver, on systemic insulin signaling.
Our efforts to link the molecular functions of dSir2 and organismal physiology led us to implicate dSir2 in starvation survival. dSir2 mutants and whole body dSir2RNAi flies succumb to starvation earlier than the controls and interestingly, are phenocopied by fatbody dSir2RNAi flies (Figures 1A, 1B and 4A). Moreover we have shown that this is due to an inability to mobilize fat reserves from the fatbody, and a resultant of decreased expression of lipid breakdown genes, both under fed and starved conditions (Figures 2 to 4). The importance of dSir2 in the fatbody and fat mobilization is corroborated by an absence of deregulated fat metabolism in muscle specific dSir2RNAi flies (Figure 3). Further, a lack of starvation phenotype when dSir2 is knocked down from the muscles highlights the physiological relevance of fatbody (Figure 4B).
In summary, we have elucidated the significance of the functions of dSir2 in the fatbody in mediating central and peripheral effects on metabolic homeostasis and insulin signaling (Figure 6). Therefore, we conclude that dSir2 is a key component that links dietary inputs with organismal physiology and survival (Figure 6). Most importantly, our study highlights the functions of dSir2 in the fatbody as a deterministic factor in governing fly physiology. This study delineates the functions of dSir2 in two metabolic tissues in affecting organismal survival. Metabolic homeostasis and the ability to utilize stored energy reserves are also crucial for mediating the effects of calorie restriction. We believe that our results, which emphasize the importance of dSir2 in maintaining homeostasis reiterates its role in calorie restriction. Finally, our report highlights the need to further investigate the functions ofdSir2, and should motivate future studies to understanding Sir2's interactions with other pathways and importance during aging.
Methods
Fly Strains
w1118, Sir22A.7.11,Sir2EP2300, cn[1] P{ry[+t7.2]=ry11}chico[1]/CyO;ry[506], and P{Switch1}106-gal4 were obtained from Bloomington Stock Center (Indiana University, USA). P{Switch-tubulin}-gal4 was a kind gift from Dr. Stephen Helfand's Lab. The rp298; P{Switch-MHC}-gal4 was obtained from NCBS, Bangalore, India. The Sir22A.7.11stock was backcrossed with w1118 for 10 generations before they were used for the assays.dSir2RNAi (CG5216:23201/GD) was obtained from the Vienna Drosophila RNAi Center (VDRC). Flies were grown on normal food under non-crowding conditions at 25°C with 12/12h light/dark cycle. Age-matched virgin female flies were used for this study.
Fly Diets
Following different dietary regimes were used for metabolic alterations [44]. Normal Diet (ND) contained yeast extract (2.5%), sucrose (5%) and cornmeal (8.6%). For low yeast medium, yeast extract was reduced to 0.25%, and for high yeast medium yeast extract was increased to 5%, keeping all the other components the same.
Starvation Assay
Three-day-old flies were kept in vials (10 per vial) containing 2% agar and transferred into fresh vials every 8 hours. To determine starvation survival response, dead flies were scored after every transfer. To characterize metabolic and molecular effects of starvation flies were starved (as mentioned earlier) for 48 hours and snap frozen in liquid nitrogen.
Activation of inducible gal4
The inducible gal4 (PSwitch lines) was activated by rearing flies on medium containing 200 μM RU486 (Mifepristone; Sigma Cat No.M8046) (in ethanol). Flies reared on a diet containing only ethanol were used as controls.
RNA isolation and Reverse Transcription
Total RNA was isolated from 8 flies (pooled together) using Trizol (Invitrogen, Cat. no. 15596-026) according to the manufacturer's instructions. 1μg of RNA was used for cDNA synthesis using SSIII reverse transcriptase kit (Invitrogen, Cat. no. 18080-044) as per the manufacturer's instructions.
Real time PCR analyses
qPCR was performed using Quantifast SYBR green (Qiagen, Cat. no. 204054) and Eppendorf Realplex instrument. The cycling conditions were as prescribed by the manufacturer. The primer pairs used for the assay are tabulated in Supplementary Table 1. Levels of Actin (actin5c) were used for normalization.
Lipid and Glucose Measurement
Six (3-5 day old) flies reared on normal diet were snap frozen in liquid nitrogen and homogenized in 600 μL PBST (PBS with 0.05% Tween-20). The homogenate was heated to 70°C for 10 minutes. Following centrifugation at 12000 rpm for 15 minutes, the supernatant was transferred to fresh tubes and were assayed at a pathological laboratory (Shahbazker's Lab, Colaba Mumbai-400005).
Oil-Red staining of fat bodies
Fat bodies from adult flies were dissected out and fixed in 4% formaldehyde. Followed by washing with running tap water for 10 minutes, the fixed samples were stained with freshly prepared 60% Oil Red O (Sigma Aldrich Cat. # 0625-100G) solution for 20 minutes and then rinsed with 60% isopropanol and PBS. The samples were mounted onto a slide and imaged using LSM-510 Carl Zeiss microscope.
Western Blot
Lysates were prepared, by incubating on ice for 15-20 minutes, in a buffer containing 50mM Tris-HCl pH7.5, 150mM NaCl, 1mM EDTA, 6mM EGTA, 20mM NaF, 1% Triton X-100, and protease inhibitors (Roche, Cat no. 05-056-489001). The lysates were then centrifuged at 12,000 rpm for 15 minutes at 4C to pellet the debris. Supernatants were used for protein estimations using BCA kit (Sigma, Cat no. BCA9643) and resolved on a 10% SDS-PAGE. Following electrotransfer onto PVDF membranes they were probed with appropriate antibodies according to standard procedures. Anti-Sir2 (sc-300) and anti-Actin antibodies were purchased from Santa Cruz Biotechnology (Cat. no. Sc-98262) and Sigma Aldrich (Cat. no. A1978), respectively. Chemiluminescence detection (Roche Cat. no. 12-015-196001) was used to visualize the bands.
NAD Assay
NAD+ was extracted as explained by McElfresh and McDonald [45]. Briefly, 8 flies were homogenized in 1mL of 0.5M Perchloric Acid (PCA) and centrifuged at 10, 000g for 3 minutes. The supernatant was treated with equal volumes of 1M potassium phosphate and neutralized with 3N KOH followed by 2 hours incubation at 4°C. To 350 μl of neutralized extracts, 310 μl of reaction buffer (0.232M Bicine, 2mg/ml BSA, 1.16M ethanol, 9.67mM EDTA, 3.87mM phenazineethosulphate, 1mM MTT) was added and incubated at 37°C for 5 minutes. Then cycling assay was initiated by the addition of 60μl of 0.6mg/ml alcohol dehydrogenase. After 30 minutes adding 300μl of 12mM iodoacetate terminated the reaction and the absorbance was read at 570nm. Total NAD+ was normalized to protein content.
Statistical analyses
Graphpad Instat3 and SigmaPlot were used for all statistical analyses. Log Rank was used to plot survival curves and Mantel-Cox test was used for statistical analysis. Student's t-test and ANOVA were used to analyze statistical significance of the data (*, p < 0.05; **, p < 0.01; ***, p < 0.001 or mentioned otherwise).
Supplementary Materials
Acknowledgments
We are extremely grateful to (Late) Prof. Veronica Rodrigues for her support and scientific discussions. We acknowledge critical inputs from Prof. Heidi Tissenbaum, Prof. Vidita Vaidya and Prof. BJ Rao. We also thank Ms. Deepti Ramachandran and Ms. Upasana Roy for providing useful insights. We thank Ms. Sharon Mary and Mr. Vinesh Mandot for technical support and standardization of many assays. Financial support from Tata Institute of Fundamental Research (DAE) is sincerely acknowledged.
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Haigis MC and Yankner BA. The aging stress response. Molecular Cell. 2010; 40: 333 -344. [PubMed] .
- 2. Mair W and Dillin A. Aging and survival: the genetics of life span extension by dietary restriction. Annual review of biochemistry. 2008; 77: 727 -754. .
- 3. Xue B and Kahn BB. AMPK integrates nutrient and hormonal signals to regulate food intake and energy balance through effects in the hypothalamus and peripheral tissues. The Journal of physiology. 2006; 574: 73 -83. [PubMed] .
- 4. Broughton S and Partridge L. Insulin/IGF-like signalling, the central nervous system and aging. The Biochemical journal. 2009; 418: 1 -12. [PubMed] .
- 5. Brown-Borg HM. Hormonal regulation of aging and life span. Trends in endocrinology and metabolism: TEM. 2003; 14: 151 -153. [PubMed] .
- 6. Rohde JR and Cardenas ME. The tor pathway regulates gene expression by linking nutrient sensing to histone acetylation. Molecular and cellular biology. 2003; 23: 629 -35. [PubMed] .
- 7. Bishop NA and Guarente L. Genetic links between diet and lifespan: shared mechanisms from yeast to humans. Nature reviews. Genetics. 2007; 8: 835 -844. .
- 8. Longo VD. Linking sirtuins, IGF-I signaling, and starvation. Experimental gerontology. 2009; 44: 70 -74. [PubMed] .
- 9. Stout RW, Henry RW, Buchanan KD. Triglyceride metabolism in acute starvation: the role of secretin and glucagon. European journal of clinical investigation. 1976; 6: 179 -185. [PubMed] .
- 10. Neumann-Haefelin C, Beha A, Kuhlmann J, Belz U, Gerl M, Quint M, Biemer-Daub G, Broenstrup M, Stein M, Kleinschmidt E, Schaefer HL, Schmoll D, Kramer W, Juretschke HP, Herling AW. Muscle-type specific intramyocellular and hepatic lipid metabolism during starvation in wistar rats. Diabetes. 2004; 53: 528 -534. [PubMed] .
- 11. Hedeskov CJ and Capito K. The effect of starvation on insulin secretion and glucose metabolism in mouse pancreatic islets. The Biochemical journal. 1974; 140: 423 -433. [PubMed] .
- 12. Klein S, Young VR, Blackburn GL, Bistrian BR, Wolfe RR. Palmitate and glycerol kinetics during brief starvation in normal weight young adult and elderly subjects. The Journal of clinical investigation. 1986; 78: 928 -933. [PubMed] .
- 13. Schuller E, Patel N, Item C, Greber-Platzer S, Baran H, Patsch W, Strobl W. The genetic background modifies the effects of the obesity mutation, 'fatty', on apolipoprotein gene regulation in rat liver. International journal of obesity and related metabolic disorders: journal of the International Association for the Study of Obesity. 2000; 24: 460 -467. [PubMed] .
- 14. Gil-Campos M, Canete RR, Gil A. Adiponectin, the missing link in insulin resistance and obesity. Clinical nutrition. 2004; 23: 963 -974. [PubMed] .
- 15. Raz I, Eldor R, Cernea S, Shafrir E. Diabetes: insulin resistance and derangements in lipid metabolism. Cure through intervention in fat transport and storage. Diabetes/metabolism research and reviews. 2005; 21: 3 -14. [PubMed] .
- 16. Kobayashi K, Inoguchi T, Maeda Y, Nakashima N, Kuwano A, Eto E, Ueno N, Sasaki S, Sawada F, Fujii M, Matoba Y, Sumiyoshi S, Kawate H, Takayanagi R. The lack of the C-terminal domain of adipose triglyceride lipase causes neutral lipid storage disease through impaired interactions with lipid droplets. The Journal of clinical endocrinology and metabolism. 2008; 93: 2877 -2884. [PubMed] .
- 17. Longo VD and Kennedy BK. Sirtuins in aging and age-related disease. Cell. 2006; 126: 257 -268. [PubMed] .
- 18. Delaney JR, et al. Sir2 deletion prevents lifespan extension in 32 long-lived mutants. Aging cell. 2011; 10: 1089 -1091. [PubMed] .
- 19. Kaeberlein M, McVey M, Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Saccharomyces cerevisiae by two different mechanisms. Genes & development. 1999; 13: 2570 -2580. [PubMed] .
- 20. Tissenbaum HA and Guarente L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature. 2001; 410: 227 -230. [PubMed] .
- 21. Rogina B and Helfand SL. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proceedings of the National Academy of Sciences of the United States of America. 2004; 101: 15998 -16003. [PubMed] .
- 22. Burnett C, et al. Absence of effects of Sir2 overexpression on lifespan in C. elegans and Drosophila. Nature. 2011; 477: 482 -485. [PubMed] .
- 23. Viswanathan M and Guarente L. Regulation of Caenorhabditis elegans lifespan by sir-2.1 transgenes. Nature. 2011; 477: E1 -2. [PubMed] .
- 24. Piper MD, Mair W, Partridge L. Counting the calories: the role of specific nutrients in extension of life span by food restriction. The journals of gerontology. Series A, Biological sciences and medical sciences. 2005; 60: 549 -555. .
- 25. Canto C and Auwerx J. Caloric restriction, SIRT1 and longevity. Trends in endocrinology and metabolism: TEM. 2009; 20: 325 -331. [PubMed] .
- 26. Finkel T, Deng CT, Mostoslavsky R. Recent progress in the biology and physiology of sirtuins. Nature. 2009; 460: 587 -591. [PubMed] .
- 27. Rodgers JT and Puigserver P. Fasting-dependent glucose and lipid metabolic response through hepatic sirtuin 1. Proceedings of the National Academy of Sciences of the United States of America. 2007; 104: 12861 -12866. [PubMed] .
- 28. Erion DM, et al. SirT1 knockdown in liver decreases basal hepatic glucose production and increases hepatic insulin responsiveness in diabetic rats. Proceedings of the National Academy of Sciences of the United States of America. 2009; 106: 11288 -11293. [PubMed] .
- 29. Li X, Zhang S, Blander G, Tse JG, Krieger M, Guarente L. SIRT1 deacetylates and positively regulates the nuclear receptor LXR. Molecular cell. 2007; 28: 91 -106. [PubMed] .
- 30. Nemoto S, Fergusson MM, Finkel T. Nutrient availability regulates SIRT1 through a forkhead-dependent pathway. Science. 2004; 306: 2105 -2108. [PubMed] .
- 31. Picard F, Kurtev M, Chung N, Topark-Ngarm A, Senawong T, Machado De Oliveira R, Leid M, McBurney MW, Guarente L. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-gamma. Nature. 2004; 429: 771 -776. [PubMed] .
- 32. Ponugoti B, Kim DH, Xiao Z, Smith Z, Miao J, Zang M, Wu SY, Chiang CM, Veenstra TD, Kemper JK. SIRT1 deacetylates and inhibits SREBP-1C activity in regulation of hepatic lipid metabolism. The Journal of biological chemistry. 2010; 285: 33959 -33970. [PubMed] .
- 33. Purushotham A, Schug TT, Xu Q, Surapureddi S, Guo X, Li X. Hepatocyte-specific deletion of SIRT1 alters fatty acid metabolism and results in hepatic steatosis and inflammation. Cell metabolism. 2009; 9: 327 -338. [PubMed] .
- 34. Rodgers JT, Lerin C, Haas W, Gygi SP, Spiegelman BM, Puigserver P. Nutrient control of glucose homeostasis through a complex of PGC-1alpha and SIRT1. Nature. 2005; 434: 113 -118. [PubMed] .
- 35. Liu Y, Dentin R, Chen D, Hedrick S, Ravnskjaer K, Schenk S, Milne J, Meyers DJ, Cole P, Yates J 3rd, Olefsky J, Guarente L, Montminy M. A fasting inducible switch modulates gluconeogenesis via activator/coactivator exchange. Nature. 2008; 456: 269 -273. [PubMed] .
- 36. Wang RH, Kim HS, Xiao C, Xu X, Gavrilova O, Deng CX. Hepatic Sirt1 deficiency in mice impairs mTorc2/Akt signaling and results in hyperglycemia, oxidative damage, and insulin resistance. The Journal of clinical investigation. 2011; 121: 4477 -4490. [PubMed] .
- 37. Schenk S, McCurdy CE, Philp A, Chen MZ, Holliday MJ, Bandyopadhyay GK, Osborn O, Baar K, Olefsky JM. Sirt1 enhances skeletal muscle insulin sensitivity in mice during caloric restriction. The Journal of clinical investigation. 2011; 121: 4281 -4288. [PubMed] .
- 38. Helfand SL and Rogina B. Genetics of aging in the fruit fly, Drosophila melanogaster. Annual review of genetics. 2003; 37: 329 -348. .
- 39. Frankel S, Ziafazeli T, Rogina B. dSir2 and longevity in Drosophila. Experimental gerontology. 2011; 46: 391 -396. [PubMed] .
- 40. Antosh M, Whitaker R, Kroll A, Hosier S, Chang C, Bauer J, Cooper L, Neretti N, Helfand SL. Comparative transcriptional pathway bioinformatic analysis of dietary restriction, Sir2, p53 and resveratrol life span extension in Drosophila. Cell cycle. 2011; 10: 904 -911. [PubMed] .
- 41. Bauer JH, Morris SN, Chang C, Flatt T, Wood JG, Helfand SL. dSir2 and Dmp53 interact to mediate aspects of CR-dependent lifespan extension in D. melanogaster. Aging. 2009; 1: 38 -48. [PubMed] .
- 42. Reis T, Van Gilst MR, Hariharan IK. A buoyancy-based screen of Drosophila larvae for fat-storage mutants reveals a role for Sir2 in coupling fat storage to nutrient availability. PLoS genetics. 2010; 6: e1001206 [PubMed] .
- 43. Gerhart-Hines Z, Rodgers JT, Bare O, Lerin C, Kim SH, Mostoslavsky R, Alt FW, Wu Z, Puigserver P. Metabolic control of muscle mitochondrial function and fatty acid oxidation through SIRT1/PGC-1alpha. The EMBO journal. 2007; 26: 1913 -1923. [PubMed] .
- 44. Ja WW, Carvalho GB, Zid BM, Mak EM, Brummel T, Benzer S. Water- and nutrient-dependent effects of dietary restriction on Drosophila lifespan. Proceedings of the National Academy of Sciences of the United States of America. 2009; 106: 18633 -18637. [PubMed] .
- 45. McElfresh KC and McDonald JF. The effect of temperature on biochemical and molecular properties of Drosophila alcohol dehydrogenase. Biochemical genetics. 1986; 24: 873 -889. [PubMed] .
- 46. Cahill GF Jr. Fuel metabolism in starvation. Annual review of nutrition. 2006; 26: 1 -22. .
- 47. Bordone L, Motta MC, Picard F, Robinson A, Jhala US, Apfeld J, McDonagh T, Lemieux M, McBurney M, Szilvasi A, Easlon EJ, Lin SJ, Guarente L. Sirt1 regulates insulin secretion by repressing UCP2 in pancreatic beta cells. PLoS biology. 2006; 4: e31 [PubMed] .
- 48. Kanfi Y, Peshti V, Gozlan YM, Rathaus M, Gil R, Cohen HY. Regulation of SIRT1 protein levels by nutrient availability. FEBS letters. 2008; 582: 2417 -2423. [PubMed] .
- 49. Lee IH, Cao L, Mostoslavsky R, Lombard DB, Liu J, Bruns NE, Tsokos M, Alt FW, Finkel T. A role for the NAD-dependent deacetylase Sirt1 in the regulation of autophagy. Proceedings of the National Academy of Sciences of the United States of America. 2008; 105: 3374 -3379. [PubMed] .
- 50. Landry J, Sutton A, Tafrov ST, Heller RC, Stebbins J, Pillus L, Sternglanz R. The silencing protein SIR2 and its homologs are NAD-dependent protein deacetylases. Proceedings of the National Academy of Sciences of the United States of America. 2000; 97: 5807 -58011. [PubMed] .
- 51. Zhang T, Berrocal JG, Frizzell KM, Gamble MJ, DuMond ME, Krishnakumar R, Yang T, Sauve AA, Kraus W. Enzymes in the NAD+ salvage pathway regulate SIRT1 activity at target gene promoters. The Journal of biological chemistry. 2009; 284: 20408 -20417. [PubMed] .
- 52. van der Horst A, Schavemaker JM, Pellis-van Berkel W, Burgering BM. The Caenorhabditis elegans nicotinamidase PNC-1 enhances survival. Mechanisms of ageing and development. 2007; 128: 346 -349. [PubMed] .
- 53. Anderson RM, Bitterman KJ, Wood JG, Medvedik O, Sinclair DA. Nicotinamide and PNC1 govern lifespan extension by calorie restriction in Saccharomyces cerevisiae. Nature. 2003; 423: 181 -185. [PubMed] .
- 54. Nakahata Y, Kaluzova M, Grimaldi B, Sahar S, Hirayama J, Chen D, Guarente LP, Sassone-Corsi P. The NAD+-dependent deacetylase SIRT1 modulates CLOCK-mediated chromatin remodeling and circadian control. Cell. 2008; 134: 329 -340. [PubMed] .
- 55. Giannakou ME, Goss M, Jacobsen J, Vinti G, Leevers SJ, Partridge L. Dynamics of the action of dFOXO on adult mortality in Drosophila. Aging cell. 2007; 6: 429 -438. [PubMed] .
- 56. Roman G, Endo K, Zong L, Davis RL. P[Switch], a system for spatial and temporal control of gene expression in Drosophila melanogaster. Proceedings of the National Academy of Sciences of the United States of America. 2001; 98: 12602 -12607. [PubMed] .
- 57. Osterwalder T, Yoon KS, White BH, Keshishian H. A conditional tissue-specific transgene expression system using inducible GAL4. Proceedings of the National Academy of Sciences of the United States of America. 2001; 98: 12596 -12601. [PubMed] .
- 58. Hou X, Xu S, Maitland-Toolan KA, Sato K, Jiang B, Ido Y, Lan F, Walsh K, Wierzbicki M, Verbeuren TJ, Cohen RA, Zang M. SIRT1 regulates hepatocyte lipid metabolism through activating AMP-activated protein kinase. The Journal of biological chemistry. 2008; 283: 20015 -20026. [PubMed] .
- 59. Tissenbaum HA and Ruvkun G. An insulin-like signaling pathway affects both longevity and reproduction in Caenorhabditis elegans. Genetics. 1998; 148: 703 -717. [PubMed] .
- 60. Taguchi A and White M. Insulin-like signaling, nutrient homeostasis, and life span. Annual review of physiology. 2008; 70: 191 -212. .
- 61. Ikeya T, Galic M, Belawat P, Nairz K, Hafen E. Nutrient-dependent expression of insulin-like peptides from neuroendocrine cells in the CNS contributes to growth regulation in Drosophila. Current biology:CB. 2002; 12: 1293 -1300. [PubMed] .
- 62. Clancy DJ, Gems D, Harshman GL, Oldham S, Stocker H, Hafen E, Leevers SJ, Partridge L. Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein. Science. 2001; 292: 104 -106. [PubMed] .
- 63. Yamamoto R and Tatar M. Insulin receptor substrate chico acts with the transcription factor FOXO to extend Drosophila lifespan. Aging cell. 2011; 10: 729 -732. [PubMed] .