



Introduction
Circadian clocks generate daily endogenous rhythms in behavior, physiological functions, and cellular activities, which are coordinated with external day/night cycles [1,2]. Circadian rhythms become impaired with age as evidenced by the dampening of daily oscillations in melatonin and other hormones and the disruption of night-time sleep in aged rodents and humans [3,4,5]. Remarkably, age-associated sleep fragmentation was also reported in Drosophila melanogaster[6], suggesting that effects of aging on circadian systems may be evolutionarily conserved. While aging impairs the circadian systems, there is also evidence that loss of circadian rhythms may, in turn, contribute to aging. Genetic disruption of circadian rhythms by knockout of specific clock genes leads to various age related pathologies and visible signs of premature aging in mice [7,8]. In addition, chronic jet-lag which disrupts the circadian clock, increases mortality in aged mice [9]. As extension of healthspan is of critical importance in aging human population, there is a need to elucidate how strong circadian clocks may support healthy aging.
The mechanisms linking circadian rhythms to the rate of aging and healthspan are not well understood. To address these mechanisms, we investigated whether disruption of the circadian clock affects response to homeostatic challenge and aggravates selected aging biomarkers in the model organism Drosophila melanogaster. We used a null mutation in the circadian clock gene period (per01) [10]; this gene is one of the four core clock genes that act in a negative auto-regulatory feedback loop generating daily endogenous rhythms [11,12]. The loss of per function disrupts behavioral and molecular rhythms in flies [10,11,13].
To compare lifespan and healthspan in flies with normal or disrupted circadian clock, we measured their ability to maintain ROS homeostasis during aging. We probed the health status of aging flies by exposing them to mild oxidative stress of 24h hyperoxia at increasingchronological ages, followed by assessment of the resulting oxidative damage and mortality hazards. Hyperoxia was chosen as a homeostatic challenge, because it directly leads to ROS production irrespective of age-related changes in food consumption and other physiological parameters [14].
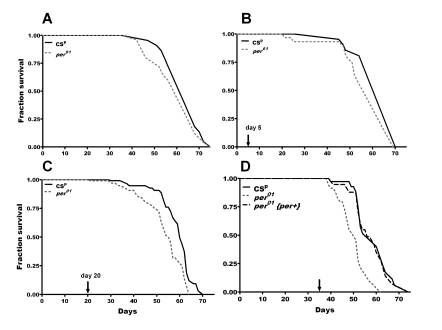
Figure 1. Lifespan of per01and CSpD. melanogaster in normoxia and following 24h
hyperoxia at different ages (marked by arrow in B-D). (A)
In normoxia, there was no significant difference in mean survival curves
(p=0.23) (B) Hyperoxia on day 5 did not significantly affect
longevity or survival curves (p=0.12) (C) Hyperoxia on day 20
resulted in a significant reduction (p<0.05) in average survival of per01flies compared to CSp with significant (p<0.0001)
difference in survival curves. (D) Hyperoxia on day 35 resulted in
more significant reduction (p<0.001) in average lifespan in per01flies compared to CSp and significant difference in survival
curves (p<0.0001). Males with rescued per function (per01{per+}) treated with hyperoxia on day 35 had average
lifespan similar to CSp but significantly different (p<0.001)
from per01mutants.
We report that per01 flies have shortened healthspan as evidenced by their increased mortality hazard in response to homeostatic challenge during aging. This conclusion is also supported by accelerated functional senescence, and increased signs of neurodegeneration in per mutants compared to age-matched controls with an intact circadian clock. In addition, we show that the expression of per gene declines with age leading to disruption of the circadian regulatory network in old wild type flies.
Results
Short-term oxidative stress shortens the lifespan in per01 mutants
To determine how loss of per affects lifespan and healthspan, per01 were backcrossed for 6 generations to Canton S strain, and this control stock was designated as CSp. Under normal laboratory conditions, the longevity of per01 males was similar to CSp controls (Figure 1A, Table 1). However, lifespan was significantly reduced in per01flies exposed to 24 h hyperoxia in middle age. Hyperoxia on day 20 shortened the average lifespan in per01 mutants by 12% while hyperoxia on day 35 decreased average lifespan of per01 flies by 20% compared to CSp males (Table 1); survival curves were significantly different in both ages (Figure 1C-D). We also calculated age specific mortality trajectories, and showed that mortality hazard significantly increased after exposure to 24 h hyperoxia on day 20 or 35 in per01 but remained unchanged in CSp males (see Supplementary Figure 1 and Supplementary Table 1). To verify that these effects are indeed linked to the lack of per gene function, we tested the lifespan of per01 flies transformed with a wild type copy of per, designated as per01{per+}. When flies with rescued per function were exposed to hyperoxia on day 35, their average survival (59 ± 2.0 days) and mortality trajectories were similar to CSp controls, but significantly different from per01 mutants (Figure 1D, Supplementary Figure 1D, and Supplementary Table 1). This verified that shortened lifespan and increased death-risk in per mutants are due to the loss of per gene. Importantly, exposure to hyperoxia on day 5 did not affect the average lifespan or mortality trajectories of per01 mutants (Figure 1B and Supplementary Figure 1B), demonstrating that hyperoxia sensitivity in these mutants is an age dependent phenotype.
Table 1. Average lifespan of CS p and per01 males exposed to 24h hyperoxia at indicated ages.
Values shown with SEM, n denotes the sample size. One-Way ANOVA with Tukey-Kramer multiple comparisons test. Statistical comparison across genotypes * = p<0.05, ** = p<0.001; within genotype, values with different superscripts are significantly different at p<0.05.
| Treatment | Genotypes | |
| CSp | per01 | |
| Normoxia | 61.5 ± 1.8a (n= 596) | 59.0 ± 1.02a (n= 640) |
| Hyperoxia day 5 | 60.4 ± 0.8a (n= 447) | 56.9 ± 0.93b (n= 480) |
| Hyperoxia day 20 | 58.4 ± 0.93a (n= 415) | 51.35 ± 1.07*c (n= 385) |
| Hyperoxia day 35 | 59.5 ± 1.03a (n = 328) | 47.8 ± 1.68**c (n= 350) |
per01 mutants accumulate more oxidative damage in response to stress and during normal aging
Given the increased mortality hazard in response to hyperoxia in per01 mutants, we next assessed the levels of oxidative damage incurred after 24 h hyperoxia exposure at the age of 5, 20, 35 and 50 days in both genotypes. Levels of protein carbonyls (PC) and the lipid peroxidation product 4-HNE were measured separately in heads and bodies. Exposure to hyperoxia induced significantly higher (p<0.001) PC levels in per01 than in CSp heads at all ages except day 5 (Figure 2A and Supplementary Table 2). Similar as in heads hyperoxia on day 35 or 50 led to moderate PC increase in CSp bodies and dramatic increase in the bodies of per01 flies (Figure 2B and Supplementary Table 2). Restoring per+ function in a per01 background resulted in PC content similar as in CSp and significantly lower than in per01males (Supplementary Table 2). Thus, the loss of per function leads to dramatically higher accumulation of PC in per01 flies faced with oxidative challenge. Similar as in the case of mortality hazard this deleterious phenotype is age dependent occurring in middle aged and old flies but not young per01mutants (Figure 1-2 and Supplementary Figure 1).
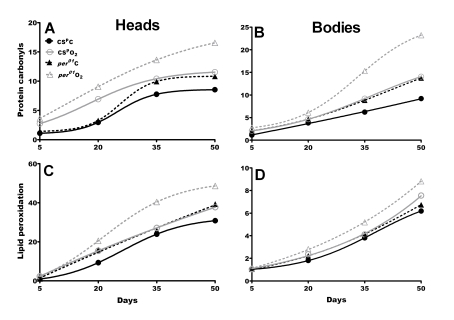
Figure 2. Oxidative damage accumulates to higher levels in aging per01flies.
Fold increase was calculated based on day 5 values in CSp males
under normoxia (numerical values are shown in Supplementary Table 2 and Supplementary Table 3). Top:
Protein carbonyls (PC) in heads (A) and bodies (B) of CSp
(solid line) and per01(broken line) in normoxia (black)
and after hyperoxia (gray). PC levels were significantly higher in per01than in CSp fly heads on day 35 and 50, and on day 50 in
bodies under normoxia. Hyperoxia on day 35 and 50 induced significantly
higher PC levels per01head and bodies compared to CSp
age-matched controls. Bottom: Lipid peroxidation product 4-HNE in
heads (C) and bodies (D). In normoxia, per01flies
accumulated significantly more 4-HNE in heads and bodies compared to CSp
in all ages except day 5. Under hyperoxia, significant increase in 4-HNE
accumulation was observed in per01heads and bodies on
day 20, 35 and 50 compared to CSp males. For statistical
analysis of PC and HNE data refer to Supplementary Table 2 and Supplementary Table 3.
The second indicator of oxidative damage, the lipid peroxidation product 4-HNE, was also measured in heads and bodies of CSp and per01 flies. Exposure to hyperoxia on day 35 and 50 significantly increased HNE in per01 heads compared to respective CSp controls (p<0.001) while exposure on day 5 or 20 had no significant effect (Figure 2C and Supplementary Table 3). Similar as in heads, hyperoxia administered on day 35 and 50 induced significantly more HNE in per01 than in CSp bodies, however, the increase was less pronounced than in fly heads (Figure 2C-D). These effects depend on the per gene as males with restored per function exhibited significantly lower HNE profiles than per01males, and similar as those observed in CSp flies (Supplementary Table 3).
Aging per01 mutants show greater mobility impairment and neurodegeneration
Our data show significantly higher accumulation of oxidative damage even in unchallenged per01 mutants under normoxia compared to age matched controls (Figure 2, Supplementary Table 2, Supplementary Table 3). As oxidative damage is one of the important biomarkers of aging, we asked whether other signs of aging are advanced in per01 mutants. First, we compared age-related locomotor performance between mutant and control flies. We used the RING assay, which utilizes negative geotaxis in Drosophila to assess climbing performance [15,16]. We measured climbing ability of per01 and CSp flies aged to day 5, 20, 35 or 50. Surprisingly, 5 day old per01 flies showed significantly higher climbing ability than control flies. In contrast, middle-aged and older per01 males showed significantly impaired climbing ability compared to age-matched controls (Figure 3). The difference was especially dramatic on day 50; at this age the average climbing ability of per01 males was approximately 4 fold lower than in CSp controls. This was partly caused by lack of vertical movement in many per01 flies at this age. The fact that young per01 mutant flies did not show impaired climbing demonstrate that the period gene does not affect fly geotaxis per se, but rather contributes to impaired climbing ability in an age-dependent fashion.
Another indicator of aging that we tested in per01 flies was the health of their nervous system. As aging is associated with degenerative morphological changes in the central nervous system, we examined brain sections from 50 day old per01, CSp, and per01{per+} males. We evaluated number of vacuoles, as they reflect the level of neurodegenerative damage in the brain [17]. Brains of per01 males showed significantly (p<0.05) greater number of vacuoles than control CSp and per01{per+} flies with restored per function (Figure 4). These vacuoles, which were found mainly in the neuropils of the optic lobes and the central brain, lead to disrupted neuronal connections. Increased vacuolization in 50 day old per01 flies is consistent with their severely impaired mobility (Figure 3).
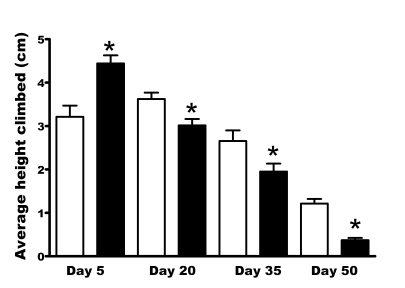
Figure 3. Vertical mobility deteriorates faster in per01flies,
as demonstrated by the RING assay. Bars represent mean height climbed
(with SEM) in CSp (open bars) and per01(black
bars) males at indicated age. The climbing performance of per01males on day 5 was significantly higher (p<0.001) compared to CSp.
With age, a rapid deterioration in climbing performance was noted in per01flies with mobility being significantly lower (* p<0.001) on day 20,
35, and 50 compared to age-matched CSp controls.
Expression of per gene declines significantly with age
Since age related functional decline is accelerated in per01 flies compared to flies with normal clock, it was of interest to investigate daily profiles of per expression during aging in control CSp flies. Therefore, we used qRT-PCR to measure the expression levels of per mRNA extracted from flies collected every 4h for 24h at age 5, 35 and 50 days. As expected [11], per mRNA levels showed daily cycling with lowest levels in the morning and a peak at early night in the heads of young flies (Figure 5A). The levels of per between peak and trough changed with a 12-fold amplitude. This amplitude dampened significantly in 35 day old flies; however, there was still pronounced cycling of per mRNA with 8-fold amplitude. A dramatic dampening of per oscillation was observed on day 50 with the amplitude reduced to 2-fold. Comparison of the relative per mRNA levels at the peak showed significant reduction by ca 70% in 50 day old flies relative to peak expression levels in young flies. Since per encodes an essential component of circadian clock, our data suggest that the circadian network is severely impaired in old flies.
Discussion
This study demonstrates healthspan extending role of the clock gene period and suggest that functional circadian clocks may prevent premature aging in flies. Research on Drosophila has demonstrated that different genetic manipulations and environmental interventions can extend fly lifespan [18]. Less attention has been paid to healthspan, despite that extension of healthspan is of critical importance in aging human population. Here, we show that healthspan can follow different trajectories in flies which have similar lifespan under stress-free laboratory conditions. Healthspan is an important but poorly defined concept, and there is an ongoing debate whether model organisms, such as Drosophila, can help to characterize parameters that could detect differences in healthspan [19]. We demonstrate that a relatively mild exogenous stress of 24 h hyperoxia, which revealed health impairment of per01 mutant, could be established as a convenient method to probe fly healthspan in a search for mechanisms supporting healthy aging.
Here, we show that healthspan, measured as the ability to respond to homeostatic challenge is reduced in per01 flies. Exposure to mild oxidative stress in middle age significantly shortened life expectancy in per01 flies but, importantly, not in control flies. The lower capacity of per01 mutants to buffer short-term oxidative challenge was linked to greatly increased accumulation of oxidative damage during hyperoxia exposure. Thus, it appears that increased mortality hazard in hyperoxia-exposed per01 mutants may be caused by their impaired ability to clear the oxidative damage which is suggested to be one of the major causes of aging [20].
The higher accrual of oxidative damage observed in per01 flies in normoxia and especially after hyperoxia could be influenced by a number of factors, with the primary suspect being higher production of endogenous ROS, which has been reported to increase in clock-disrupted flies [21] and mice [7]. Whether higher ROS is associated with decreased activity of ROS scavenging en- zymes remains to be determined. While microarray studies suggested that expression of superoxide dismutase and catalase may be controlled by the circadian clock in flies [22], qRT-PCR did not confirm such rhythm for catalase, but demonstrated that catalase activity is significantly lower in young clock-deficient flies [21]. It is currently unknown whether enzymes involved in protein repair are controlled by the circadian clock in animals, although such control was reported in plants [23]. Finally, excessive agglomeration of oxidatively damaged proteins in per01 flies could be related to impaired degradation as proteasome activity has been shown to decline with age in flies, and may be inhibited by PC and HNE [24,25]
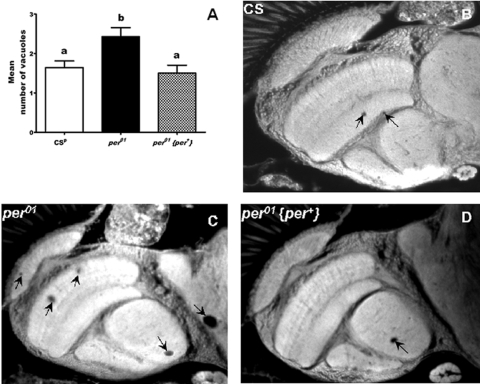
Figure 4. Neuronal
degeneration is accelerated in per01mutants compared to
CSp and flies with restored per function (per01{per+}) on day 50. (A) Mean number of
vacuoles (with SEM) representing neuronal degeneration was significantly
higher in per01mutants compared with wild type CSp
and flies with rescued per. Bars with different superscripts are
significantly different at p<0.05, data based on 10-15 heads for each
genotype. (B-D) Photomicrographs of representative brain sections of
CSp, per01, and per01{per+}
males. Arrows point to vacuolization.
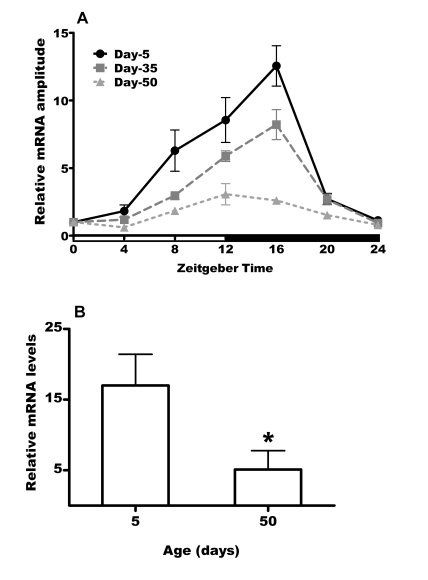
Figure 5. Expression of per mRNA declines with with age in heads of CSp
flies.
(A) Daily mRNA expression profiles of per in day 5, 35 and 50
male heads. White and black horizontal bars mark periods of light and
darkness respectively. Values were normalized to rp49 and calibrated
against ZT0 (taken as 1) for each age and represented as mean ± SEM of 3
bioreplicates. (B) The peak levels of per mRNA are
significantly reduced (* = p<0.05) in 50 day old males compared to young
control males. Values are mean ± SEM of 3 bioreplicates.
As in humans, age-related functional declines such as disrupted sleep and decreased mobility are observed in Drosophila[6,26]. The negative geotaxis assay revealed significant impairment in climbing ability in aging per01 flies relative to age-matched controls suggesting that lack of per impairs physical performance during aging. Importantly, exacerbated mobility decline in per01 flies was associated with increased neuronal degeneration in the brain. Neurodegenerative effects in the form of vacuoles in the neuropil region were observed with higher frequency in 50-day old per01 mutants than in CSp or per01{per+} flies with restored per function. The formation of vacuoles was previously linked to oxidative damage and accelerated aging in Drosophila with impaired carbonyl reductase gene [27], and in flies with Alzheimer-like phenotypes [28].
Our study suggests that functional circadian rhythms support healthy aging in flies. PER protein is the essential element of circadian clock and its absence disrupts molecular and cellular rhythms. We reported previously that young wild type flies have daily rhythms in ROS and PC levels, while in per01 flies levels of these deleterious compounds are significantly higher and arrhythmic [21]. We hypothesize that the circadian clock slows down the accumulation of oxidative damage in aging organisms by synchronizing the activities of enzymes involved in protein homeostasis. For example, microarray studies reported synchronous upregulation of several GST enzymes in flies [29], and it is known that glutathione participates in the conjugation of oxidized proteins [30]. In the absence of circadian clock, enzymes working in a specific pathway may become dysregulated leading to impaired removal of oxidative damage. However, we cannot exclude the possibility that per could affect efficiency of anti-oxidative defense systems independent of its role as a clock component, by acting in a pleiotropic non-circadian manner.
While loss of the circadian rhythms by disruption of the gene period accelerates aging, organisms with normal clocks also age. Our data demonstrate that at middle age per01 mutant shows aging phenotypes normally observed in chronologically older wild type flies, suggesting that clock gene activities may decline with age. Indeed, we demonstrate the amplitude of per mRNA oscillation is severely dampened in 50 day old flies and levels of per mRNA are significantly reduced at late night, when PER acts as essential element of clock negative feedback loop [11]. This suggests that circadian clocks and, consequently circadian rhythms are severely impaired in individuals of advanced age, which is consistent with declining strength of behavioral rhythms reported in aging flies [6]. While factors contributing to the decline of circadian rhythms in flies remain to be elucidated, oxidative stress is likely to be involved. We show here that oxidative damage accumulates to high levels even in wild type aging flies, and a previous report demonstrated that paraquat-induced oxidative stress, or decrease in FOXO expression, led to dampened per expression in Drosophila[31]. Decline in clock genes with age has been reported in zebrafish [32], rats [33] and most recently in rhesus monkey [34]. The intriguing similarities in the behavior of clock genes during aging between mammals, zebrafish, and flies warrants investigations of the mechanisms causing disruption of the circadian networks. Understanding these mechanisms will help to determine in future whether strong circadian clocks add water to the fountain of youth.
Materials and Methods
Fly rearing and l ife span analysis. Drosophila melanogaster were reared on yeast-cornmeal-molasses-agar diet (35g yeast/l) at 25°C in a 12-hour light/12-hour dark cycles; all experiments were performed 4-8 h after lights-on. The per01 mutant flies [10] were backcrossed 6 times to the Canton-S (CS) flies designated as CSP. To rescue per-function, we used transgenic flies carrying a wild-type copy of per (designated as perG) in a per01 background [35]. Males with two copies of perG (y wper01;{per+:32.1};+ were crossed with per01;+;+ females, and F1 males containing one copy of rescue construct designated per01{per+} were used. We confirmed their rhythmic locomotor activity indicating rescue of circadian clock function.
To determine lifespan, 3-4 cohorts of 100 flies of each genotype were housed in 16 oz transparent plastic bottles inverted over 60 mm Petri-dishes containing 15 ml of diet. Diet was replaced on alternate days without anesthesia, and mortality was recorded daily. For hyperoxia exposure, males were transferred from cages to narrow vials with diet in groups of 25, and placed in a Plexiglas chamber filled with oxygen (100% medical grade) flowing at a constant rate (300ml/min) for 24 h. Control flies were transferred to narrow vials as above and kept under normoxia. Hyperoxia-treated and control flies were then either frozen for oxidative damage analysis or returned to cages and monitored for mortality.
Oxidative damage assays. The amount of protein carbonyls was assayed separately in 25 heads and bodies. Carbonyls were quantified after reaction with 2,4-dinitrophenylhydrazine (DNPH) as described previously [21] at 370 nm in a BioTek Synergy 2 plate reader. Results were expressed as nmol.mg-1 protein using an extinction coefficient of 22,000 M-1cm-1. The lipid peroxidation product 4-hydroxy-2-nonenal (4-HNE) was assayed in heads and bodies by competitive enzyme-linked immunosorbent assay (ELISA) as described [36, 37]. Briefly, free HNE (Alpha Diagnostic, San Antonio, TX, USA) was conjugated to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) protein [38]. Wells in a 96-well plate were coated with 500 ng of HNE-GAPDH protein for 24h at 4°C, washed in PBS-Tween, and blocked with 1% BSA. A standard dose-response curve was developed from serial dilutions of HNE-GAPDH with polyclonal anti-HNE antibody (1:1000; Alpha Diagnostic). For samples, 10 μ;g of protein lysate was mixed with 1:1000 polyclonal rabbit anti-HNE antibody and added to wells in triplicate. Plates were incubated for 1 h, washed with buffer, incubated with 1:5000 secondary anti-rabbit antibody conjugated with horseradish peroxidase, washed, mixed with detection buffer TMB (Alpha Diagnostic), and read at OD 450nm in a BioTek plate reader.
Rapid iterative negative geotaxis (RING) assay. Vertical mobility was assayed using the RING method [15]. Briefly, 3 groups of 25 CSp or per01 flies were transferred into empty narrow vials, which were loaded into the RING apparatus. After 3 minutes rest, the apparatus was rapped sharply on the table three times in rapid succession to initiate a negative geotaxis response. The flies' movements in tubes were videotaped and digital images captured 4 s after initiating the behavior. Five consecutive trials were interspersed with a 30s rest. The climbing performance was calculated and expressed as average height climbed in the 4 s interval. The performance of flies in a single vial was calculated as the average of 5 consecutive trials to generate n = 1.
Neuronal degeneration. Paraffin-embedded sections of heads were used to examine neurodegenerative defects. Fly heads of all genotypes were processed and sectioned in parallel, and microscopic pictures taken at the same level of the brain and the number and volume of vacuoles counted in double-blind experiments using described methods [39, 40].
Quantitative Real-Time PCR. 25 male heads were collected for each time point in triplicate, homogenized in TriReagent (Sigma), and RNA was isolated following manufacturer protocol. Samples were purified using the RNeasy mini kit (Qiagen) with on-column DNAse digestion (Qiagen). Synthesis of cDNA was achieved with Sprint RT Complete kit (Clontech) or iScript cDNA synthesis kit (Biorad). Real-time PCR was performed on Step-One Plus real-time machine (Applied Biosystems) in triplicate under default thermal cycling conditions with a dissociation curve step. Each reaction contained iTaq SYBR Green Supermix with ROX (Biorad), 0.6-1ng cDNA, 80nM primers (IDT Technologies). Primers sequences are available upon request. Data were analyzed using the standard 2-∆∆CT method normalized to the gene rp49 and expressed relative to control samples at ZT0.
Statistical analyses. Life span and survival curves were plotted following Kaplan Meier survival analysis and statistical significance of curves assessed using the Log-Rank (Mantel-Cox) and Gehan-Breslow-Wilcoxon test (GraphPad Prism v 5.0).Age-specific mortality was calculated using the Gompertz's model of population aging. Ln values of instantaneous mortality (μx) were plotted against chronological time. Mortality calculations and Gompertz-Makeham maximum likelihood estimates were done using WinModest V1.0.2 [41] and plotted on GraphPad Prism. For statistical analysis of biochemical results three-way ANOVA with post-hoc tests were performed using OpenStat (William G. Miller © 2009). Statistical analysis of locomotor assays was done with one and two-way ANOVA for comparison between ages and genotypes.
Supplementary Materials
Acknowledgments
We thank Dr. M. Grotewiel for sharing RING protocols, Dr. P. Hardin for flies, and Drs. S. Pletcher and C. Davis for help with mortality hazard calculations. We thank Megan Mathes, Nick Meermeier, and Katie Sherman for excellent laboratory assistance, and Drs. L. Hooven, A. Sehgal, and P. Taghert for helpful comments on the manuscript. This work was supported in part by the NIH-NINDS NS047663 to DK, NIH-NIGMS GM073792, NRI, CSREES, USDA 2007-04617, and The Oregon Partnership for Alzheimer Research grants to JMG.
Conflicts of Interest
The authors have no conflict of interests to declare.
References
- 1. Hastings MH , Reddy AB and Maywood ES. A clockwork web: Circadian timing in brain and periphery in health and disease. Nature Rev Neurosci. 2003; 4: 649 -661. [PubMed] .
- 2. Green CB , Takahashi JS and Bass J. The meter of metabolism. Cell. 2008; 134: 728 -742. [PubMed] .
- 3. Huang YL , Liu RY , Wang QS , Van Someren EJ , Xu H and Zhou JN. Age-associated difference in circadian sleep-wake and rest-activity rhythms. Physiol Behav. 2002; 76: 597 -603. [PubMed] .
- 4. Turek FW , Penev P , Zhang Y , van Reeth O and Zee P. Effects of age on the circadian system. Neurosci Biobehav Rev. 1995; 19: 53 -58. [PubMed] .
- 5. Hofman MA and Swaab DF. Living by the clock: the circadian pacemaker in older people. Ageing Res Rev. 2006; 5: 33 -51. [PubMed] .
- 6. Koh K , Evans JM , Hendricks JC and Sehgal A. A Drosophila model for age-associated changes in sleep:wake cycles. Proc Natl Acad Sci USA. 2006; 103: 13843 -13847. [PubMed] .
- 7. Kondratov RV , Kondratova AA , Gorbacheva VY , Vykhovanets OV and Antoch MP. Early aging and age-related pathologies in mice deficient in BMAL1, the core component of the circadian clock. Genes Dev. 2006; 20: 1868 -1873. [PubMed] .
- 8. Lee CC Tumor suppression by the mammalian Period genes. Cancer Causes Control. 2006; 17: 525 -530. [PubMed] .
- 9. Davidson AJ , Sellix MT , Daniel J , Yamazaki S , Menaker M and Block GD. Chronic jet-lag increases mortality in aged mice. Curr Biol. 2006; 16: R914 -R916. [PubMed] .
- 10. Konopka RJ and Benzer S. Clock mutants of Drosophila melanogaster. Proc Natl Acad Sci U S A. 1971; 68: 2112 -2116. [PubMed] .
- 11. Hardin PE The circadian timekeeping system of Drosophila. Curr Biol. 2005; 15: R714 -R722. [PubMed] .
- 12. Zheng X and Sehgal A. Probing the relative importance of molecular oscillations in the circadian clock. Genetics. 2008; 178: 1147 -1155. [PubMed] .
- 13. Krishnan B , Dryer SE and Hardin PE. Circadian rhythms in olfactory responses of Drosophila melanogaster. Nature. 1999; 400: 375 -378. [PubMed] .
- 14. Kulkarni AC , Kuppusamy P and Parinandi N. Oxygen, the lead actor in the pathophysiologic drama enactment of the trinity of normoxia, hypoxia and hyperoxia in disease and therapy. Antioxid Redox Signal. 2007; 9: 1717 -1730. [PubMed] .
- 15. Gargano JW , Martin I , Bhandari P and Grotewiel MS. Rapid iterative negative geotaxis (RING): a new method for assessing age-related locomotor decline in Drosophila. Exp Gerontol. 2005; 40: 386 -395. [PubMed] .
- 16. Rhodenizer D , Martin I , Bhandari P , Pletcher SD and Grotewiel M. Genetic and environmental factors impact age-related impairment of negative geotaxis in Drosophila by altering age-dependent climbing speed. Exp Gerontol. 2008; 43: 739 -748. [PubMed] .
- 17. Kretzschmar D Neurodegenerative mutants in Drosophila: a means to identify genes and mechanisms involved in human diseases. Invert Neurosci. 2005; 5: 97 -109. [PubMed] .
- 18. Helfand SL and Rogina B. From genes to aging in Drosophila. Adv Genet. 2003; 49: 67 -109. [PubMed] .
- 19. Tatar M Can we develop genetically tractable models to assess healthspan (rather than life span) in animal models. J Gerontol A Biol Sci Med Sci. 2009; 64: 161 -163. [PubMed] .
- 20. Stadtman ER Protein oxidation and aging. Free Radic Res. 2006; 40: 1250 -1258. [PubMed] .
- 21. Krishnan N , Davis AJ and Giebultowicz JM. Circadian regulation of response to oxidative stress in Drosophila melanogaster. Biochem Biophys Res Commun. 2008; 374: 299 -303. [PubMed] .
- 22. Ceriani MF , Hogenesch JB , Yanovsky M , Panda S , Straume M and Kay SA. Genome-wide expression analysis in Drosophila reveals genes controlling circadian behavior. J Neurosci. 2002; 22: 9305 -9319. [PubMed] .
- 23. Bechtold U , Murphy DJ and Mullineaux PM. Arabidopsis peptide methionine sulfoxide reductase2 prevents cellular oxidative damage in long nights. Plant Cell. 2004; 16: 908 -919. [PubMed] .
- 24. Gaczynska M and Osmulski PA. Ward WF. Caretaker or undertaker? The role of the proteasome in aging. Mech Ageing Dev. 2001; 122: 235 -254. [PubMed] .
- 25. Vernace VA , Arnaud L , Schmidt-Glenewinkel T and Figueiredo-Pereira ME. Aging perturbs 26S proteasome assembly in Drosophila melanogaster. Faseb J. 2007; 21: 2672 -2682. [PubMed] .
- 26. Grotewiel MS , Martin I , Bhandari P and Cook-Wiens E. Functional senescence in Drosophila melanogaster. Aging Res Rev. 2005; 4: 372 -397. .
- 27. Botella JA , Ulschmid JK , Gruenewald C , Moehle C , Kretzschmar D , Becker K and Schneuwly S. The Drosophila carbonyl reductase sniffer prevents oxidative stress-induced neuro-degeneration. Curr Biol. 2004; 14: 782 -786. [PubMed] .
- 28. Carmine-Simmen K , Proctor T , Tschape J , Poeck B , Triphan T , Strauss R and Kretzschmar D. Neurotoxic effects induced by the Drosophila amyloid beta peptide suggest a conserved toxic function. Neurobiol Dis. 2009; 33: 274 -281. [PubMed] .
- 29. Wijnen H and Young MW. Interplay of circadian clocks and metabolic rhythms. Annu Rev Genet. 2006; 40: 409 -448. [PubMed] .
- 30. Tu CP and Akgul B. Drosophila glutathione S-transferases. Methods Enzymol. 2005; 401: 204 -226. [PubMed] .
- 31. Zheng X , Yang Z , Yue Z , Alvarez JD and Sehgal A. FOXO and insulin signaling regulate sensitivity of the circadian clock to oxidative stress. Proc Natl Acad Sci USA. 2007; 104: 15899 -15904. [PubMed] .
- 32. Zhdanova IV , Yu L , Lopez-Patino M , Shang E , Kishi S and Guelin E. Aging of the circadian system in zebrafish and the effects of melatonin on sleep and cognitive performance. Brain Res Bull. 2008; 75: 433 -441. [PubMed] .
- 33. Asai M , Yoshinobu Y , Kaneko S , Mori A , Nikaido T , Moriya T , Akiyama M and Shibata S. Circadian profile of Per gene mRNA expression in the suprachiasmatic nucleus, paraventricular nucleus, and pineal body of aged rats. J Neurosci Res. 2001; 66: 1133 -1139. [PubMed] .
- 34. Sitzmann BD , Lemos DR , Ottinger MA and Urbanski HF. Effects of age on clock gene expression in the rhesus macaque pituitary gland. Neurobiol Aging. 2008; in press; doi: In press .
- 35. Cheng Y , Gvakharia B and Hardin PE. Two alternatively spliced transcripts from the Drosophila period gene rescue rhythms having different molecular and behavioral characteristics. Mol Cell Biol. 1998; 18: 6505 -6514. [PubMed] .
- 36. Satoh K , Yamada S , Koike Y , Igarashi Y , Toyokuni S , Kumano T , Takahata T , Hayakari M , Tsuchida S and Uchida K. A 1-hour enzyme-linked immunosorbent assay for quantitation of acrolein and hydroxynonenal-modified proteins by epitope-bound casein matrix method. Anal Biochem. 1999; 270: 323 -328. [PubMed] .
- 37. Zheng J , Mutcherson R 2nd and Helfand SL. Calorie restriction delays lipid oxidative damage in Drosophila melanogaster. Aging Cell. 2005; 4: 209 -216. [PubMed] .
- 38. Uchida K and Stadtman ER. Covalent attachment of 4-hydroxynonenal to glyceraldehyde-3-phosphate dehydrogenase. A possible involvement of the intra- and intermolecular cross-linking reaction. J Biol Chem. 1993; 268: 6388 -6393. [PubMed] .
- 39. Tschape JA , Hammerschmied C , Muhlig-Versen M , Athenstaedt K , Daum G and Kretzschmar D. The neurodegeneration mutant lochrig interferes with cholesterol homeostasis and Appl processing. Embo J. 2002; 21: 6367 -6376. [PubMed] .
- 40. Bettencourt da Cruz A , Schwarzel M , Schulze S , Niyyati M , Heisenberg M and Kretzschmar D. Disruption of the MAP1B-related protein FUTSCH leads to changes in the neuronal cytoskeleton, axonal transport defects, and progressive neurodegeneration in Drosophila. Mol Biol Cell. 2005; 16: 2433 -2442. [PubMed] .
- 41. Pletcher SD Model fitting and hypothesis testing for age-specific mortality data. J Evol Biol. 1999; 12: 430 -439. .