



Introduction
Age-related cognitive impairment has become a major public concern as its associated loss of independence confers a substantial global health and economic burden. Vascular cognitive impairment and dementia (VCID) is the second most common cause of dementia only next to Alzheimer’s disease (AD) and accounts for almost 20–30% of cases. One of the primary clinical manifestations of vascular pathologies is impaired cerebral blood flow (CBF) or chronic cerebral hypoperfusion which has been reported to be positively associated with cognitive decline both in preclinical and clinical studies [1–5]. Microvascular endothelial cells, which form the inner lining of all cerebral vessels, play multifaceted roles in regulating cerebral blood flow and cognition. First, endothelial nitric oxide contributes to neurovascular coupling responses (NVC), a critical vasodilatory mechanism that maintains neuronal homeostasis and function by promptly matching local neuronal activity with the required increase in cerebral blood flow (CBF) [6, 7]. Secondly, endothelial glucose uptake through Glut1 (endothelial isoform 55kDa) controls the first point of glucose entry into the brain and critically contributes to the maintenance of whole-brain energy homeostasis in addition to supporting its metabolic needs [8]. Thirdly, endothelial cells maintain the structural integrity of the blood-brain barrier (BBB), which prevents the entry of serum constituents into the brain parenchyma, and subsequent glial activation and neuroinflammation [9, 10]. Lastly, endothelial angiogenesis is key to maintaining the optimal cerebral microvascular density needed to achieve adequate perfusion to the entire brain [11]. Accumulating evidence points to age-related impairment in endothelial function, metabolism, and structure resulting in impaired NVC [12, 13], attenuated glucose uptake leading to hypometabolism [14, 15], BBB leakage leading to neuroinflammation [16, 17] and microvascular rarefaction [18, 19]. All these mechanisms synergistically act to reduce cerebral blood flow or perfusion and contribute to cognitive decline in aging.
The majority of the previous research in the microvascular aging field has primarily focused on targeting cell-intrinsic mechanisms including senescence, oxidative stress, DNA damage, etc. However, a paradigm shift in the mechanistic view of endothelial dysfunction has occurred since the emergence of results from heterochronic parabiosis studies in recent years. These studies where young and old mice share the circulation for an extended period highlight the pro-aging role of circulating factors in accelerating endothelial dysfunction [20–24]. Especially, altered systemic and metabolic milieu in aging including chronic inflammation, hyperglycemia, hyperinsulinemia, and dyslipidemia has been implicated in endothelial dysfunction [25–30]. In addition, age-related reduction in the circulating vasoprotective factors such as IGF1 and adiponectin could also potentially contribute to microvascular aging [31–33]. Conforming to this overall idea, several meta-analysis studies have reported that patients diagnosed with age-related diabetes, dyslipidemia, and metabolic syndrome have an elevated risk of developing cognitive impairment later in their lifetime [34–37]. Overall, these studies signify the importance of interventions that improve metabolic dysfunction in treating and/or preventing VCI in aging.
Adipose tissue plays a central role in whole-body energy homeostasis through its direct involvement in glucose and lipid metabolism and also indirectly via its crosstalk with other systemic tissues through secreted factors. Age-related pathological changes in adipose tissue contribute to metabolic dysfunction through ectopic lipid deposition, insulin resistance, and low-grade chronic inflammation [33, 38–41], all of which have been implicated in accelerating endothelial aging. On the other hand, improvements in adipose tissue metabolism, at least in part, contribute to the delayed aging phenotype observed in response to several well-known anti-aging interventions [33, 41–43]. More importantly, adipose-related metabolic dysfunction in middle age precedes the onset of cognitive decline later in life [44–47], suggesting that interventions that restore adipose and systemic metabolism could be targeted to delay or prevent brain endothelial dysfunction and VCI in aging.
We have recently reported that the pharmacological activation of adipose thermogenesis, a catabolic phenomenon marked by increased fuel oxidation and energy expenditure, improved the overall systemic metabolism in aged mice [48]. Specifically, we used a β3-adrenergic receptor agonist (β3AR, CL, 216243) to stimulate thermogenesis in aged mice. β3ARs are predominantly expressed in the white and brown adipose tissue and play a critical role in the maintenance and activation of lipolytic and thermogenic machinery. β3AR agonists (CL, 316243 in rodents and FDA-approved mirabegron in humans) have been extensively validated as a pharmacological means to stimulate thermogenesis and improve systemic glucose and lipid metabolism [49–51], however, its relevance in aged subjects has remained questionable. Addressing this, we have recently shown that the metabolic benefits of β3AR agonist treatment are preserved in aged mice [48]. Chronic β3AR stimulation increased whole-body energy expenditure, reduced fat mass, improved glucose tolerance and insulin sensitivity, increased circulating adiponectin levels, and reduced ectopic lipid deposition in aged mice [48]. In the present study, we wanted to examine whether β3AR stimulation-mediated improvements in systemic metabolism and circulating milieu can mitigate microvascular endothelial dysfunction and cognitive decline in aged mice.
Materials and Methods
Animals and treatment
All animal protocols were approved by the Institutional Animal Care and Use Committee at the University of Oklahoma Health Sciences Center (OUHSC). Aged C57BL/6J male and female mice (18 months old) were obtained from the aging colony maintained by the National Institute on Aging and were fed a standard chow diet (PicoLab Rodent Diet 5053) with continuous access to water and enrichment. The animals were housed in the conventional animal housing facility with a 12:12-hour light-dark cycle at OUHSC. Aged mice were implanted subcutaneously with osmotic minipumps filled with saline or β3-AR agonist (CL 316,243 (CL) R&D Systems-Cat. No. 1499/50, 0.75 nmol/h) to enable continuous infusion for 6 weeks as previously described [51]. At the end of 4 weeks of treatment, the mice were subjected to behavioral assays including radial arm water maze (RAWM) and Y-maze to assess spatial learning and memory-related cognitive outcomes. A sub-cohort of animals underwent PET/CT imaging to assess glucose uptake at the end of 4 weeks. All the animals were sacrificed at the end of 6 weeks and brain tissues were collected and either stored at -80C for protein analysis or fixed in 10% formalin for paraffin embedding. A separate cohort of young (3–4 mos) and aged (20–22 mos) C57BL/6J animals were also used for validation of NVC and PET/CT imaging techniques. Power calculation: To achieve >80% power with 0.05 type I error rate, we require a minimum of 7 animals per group for protein and functional assessments and a minimum of 10 animals per group for behavioral analyses. These calculations assume an effect size comparable to that observed in our previously published data on the variables under study.
Radial arm water maze test
Spatial memory and long-term memory in mice were assessed by performance in the radial arm water maze (RAWM) test as described previously [52, 53]. The RAWM consisted of eight 9 cm wide arms that radiated out from the open central area, with a submerged escape platform located at the end of one of the arms. Food-grade white paint was added to make the water opaque and mask the escape platform. Visual cues were marked inside the maze at the end of each arm. The movement of mice was monitored by a video tracking system directly above the maze and the parameters including distance, time, and latency to escape were recorded using Ethovision software (Noldus Information Technology Inc., Leesburg, VA, USA). The experiment consisted of three consecutive days of learning trials (Days 1–3), followed by a 7-day break (Days 4–10), a probe trial on Day 11, and a reversal trial on Day 12. During the learning phase, mice underwent eight trials/day. The data from 8 trials were consolidated as a single block per day totaling 3 blocks for 3 learning days (Learning day 1, 2 and 3). In each trial, mice were started in an arm not containing the submerged escape platform and were allowed up to 1 minute to locate it. To ensure task familiarization, mice were allowed to spend 30 seconds on the platform after the first trial on Day 1. Following the learning phase, mice were housed in their home cages for 7 days before being subjected to the probe trial (Day 11), which assessed memory recall of the platform’s original location through four 1-minute trials (grouped into 1 block designated as Probe). On the reversal trial (Day 12), the platform was moved to a new arm (neither adjacent nor diametrically opposite to the original location), and mice were tested for their ability to relearn its location. This relearning phase involved eight trials, grouped into one block termed as Reversal. The mice were charged an error whenever they entered an incorrect arm (all four paws within the distal half of the arm) or spent 15 seconds at the center without entering any arm.
Neurovascular coupling assessments
After completion of behavioral tests, neurovascular coupling responses were assessed in a sub-group of animals using laser speckle contrast imaging as described previously [53, 54]. Briefly, mice were anesthetized with 2% isoflurane, endotracheally intubated and ventilated, but quickly switched to 0.5%-1% maintenance dose during the measurement of NVC responses. Cardio respiratory parameters such as blood pressure were monitored and maintained within the physiological range throughout the experiments. The right femoral artery was cannulated with a pressure transducer to continuously monitor and maintain the arterial blood pressure between 90–110 mmHg (Living Systems Instrumentations, Burlington, VT, USA). Further a thermostatic heating pad (Kent Scientific Co., Torrington, CT, USA) was used to maintain rectal temperature at 37°C. End-tidal CO2 (including dead space) was maintained between 3.2% and 3.7% to keep blood gas values within the physiological range (PaCO2 levels were consistently maintained between 35–45 mmHg, and PaO2 levels remained above 90 mmHg) during all procedures. Following immobilization in the stereotaxic frame, the scalp and periosteum were opened and the skull was thinned using a dental drill. To avoid overheating during drilling, dripping buffer was infused at the drilling site. After placement of the laser speckle contrast imager (Perimed, Jarfalla, Sweden) above the thinned site, CBF responses on the left somatosensory cortex were captured by stimulating the right whiskers for 30s at 5Hz from side to side. A total of six trials were performed with 5–10-minute intervals between them. The average of the CBF changes during the 6 trials was expressed as a % increase from the baseline values.
18F-FDG PET/CT imaging to assess brain glucose uptake
Briefly, overnight-fasted animals were injected with 18F-FDG (100 μCi) via the tail vein. After 2 hours of FDG uptake, a 15-minute PET image was acquired immediately followed by a 2-minute CT image. Both images were acquired using an MI Labs Vector6 machine (Utrecht, Netherlands). Images were reconstructed and registered using MI Labs software. ROI for the brain was then manually selected and the 18F activity in this ROI was quantified in the corresponding region of the registered PET image using AMIRA software (Thermo Fisher Scientific). Percent injected dose (% ID) was calculated as the activity (µCi) in the brain decay corrected to the time of the injection (i.t.) divided by the injected activity (µCi). The standard uptake value (SUV) was calculated by normalizing the % ID to the body weight of the animals. The imaging and analysis were performed in the Research imaging facility (RIF) at the College of Pharmacy in OUHSC.
BBB permeability assays
BBB permeability was assessed by quantifying the levels of extravasated fluorescent tracers in brain lysates as described previously by Devraj et al. [55]. Briefly, the mice were injected with 100 µl of 2 mM of 3Kda FITC dextran tracer (#D3305, Thermo Fisher Scientific, Waltham, MA, USA) by intraperitoneal injection. After 15 minutes, the mice were anesthetized and cardiac perfusion with ice-cold PBS was performed to remove the tracers from the vascular compartment. Cortex and hippocampus regions of the brain were dissected from one sagittal section of hemibrain and the other half of the brain was stored in formalin for immunofluorescence analysis. The permeability index was assessed by measuring the fluorescence intensity in the serum and brain homogenates (cortex and hippocampus) at an excitation/emission (nm) value of 490/520 using a plate reader. All raw fluorescence values (RFU) were corrected for background using tissue homogenates or serum from sham animals that did not receive tracer injection. The permeability index was calculated using the following formula: Permeability Index (mL/g) = (Tissue RFUs/g tissue weight)/(Serum RFUs/mL serum).
Immunohistochemistry for microvascular density
Microvascular density was assessed by immunostaining for endothelial cells. Frozen OCT-embedded brains were cut sagittally (35 μm) and stored in cryoprotectant solution (25% glycerol, 25% ethylene glycol, 25% of 0.1 m phosphate buffer, and 25% water) at −20°C. Sections were rinsed with Tris-buffered saline (TBS) and permeabilized with TBS with 0.05% Tween-20. After blocking with 5% BSA and 1% fish gelatin in TBS at room temperature for 2 h, sections were immunostained with a cocktail of anti-endomucin (1:75, Millipore) and anti-CD31 (1:50, BD biosciences) antibodies for 48 hrs at 4°C. Sections were washed for 5 min (3×) with TBST followed by incubation with goat anti-rat Alexa Fluor 488 secondary antibody (1:500, Thermo Fisher Scientific) for 2 hours at room temperature. The sections were then washed with TBST for 5 min (3×) and mounted onto slides using a Prolong antifade mounting medium (Thermo Fisher Scientific). Confocal images were obtained using Leica SP8 MP confocal laser scanning microscope using tiling mode and 10X objective. High resolution Z-stacks (tiles) were stitched to depict the whole brain followed by imaging with 20X objective. At least 2 representative z stacked images were captured in the cortex and hippocampus and the vessel density and vessel lengths were calculated using the AngioTool software.
Capillary-based immunoassay for Glut 1 protein expression
Frozen cortex and hippocampus samples were lysed using 1X cell lysis buffer (Cell Signaling Technology, Danvers, MA, USA) containing Halt protease and phosphatase inhibitor cocktail (Thermo Fisher Scientific, #PI78440). The lysates were obtained by mincing the tissue using a Dounce homogenizer followed by centrifugation at 16,000 g for 10 min at 4°C. The clear supernatant was collected and the protein concentrations were determined using the Pierce BCA Protein Assay Kit (Thermo Fisher Scientific, #23227). Automated western blots were performed using Jess capillary-based immunoassay using 12-230 kDa separation with protein normalization (PN) module using the Compass for SW Software 6.2.0 (Protein Simple). Protein samples were diluted with 0.1X sample buffer and loaded at 0.5 mg/mL optimized concentration. Anti-GLUT1 antibody (Abcam #ab115730) was loaded at 1:50 dilution. The peak area for 45kDa and 55kDa isoforms of GLUT1 were calculated using the dropped line peak integration and normalized to the total protein content (PN module) using the Compass for SW Software 6.2.0 (Protein Simple).
Milliplex assays for cytokine analysis
Protein lysates from cortex samples were analyzed for inflammatory markers (Millipore Sigma #MCYTOMAG-70K-PMX) using Milliplex kits. The values from protein lysates were normalized to the total protein content in each sample assessed by BCA assay and expressed as pg/ug of protein.
Statistical analysis
Statistical analyses were performed using Graph pad prism 9.3.1 (GraphPad Software, San Diego, CA, USA) and the data are expressed as mean ± SEM. Data were analyzed by two-tailed, unpaired student’s t-test and p < 0.05 were considered statistically significant.
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Results
Chronic β3AR stimulation improved neurovascular coupling responses and brain glucose uptake in aged mice
We assessed neurovascular coupling in the somatosensory cortex via laser speckle contrast imaging following 6 weeks of CL treatment in aged mice. Cerebral blood flow responses in the somatosensory cortex in response to contralateral whisker stimulation were significantly increased in aged mice treated with CL (~10% increase in CBF, comparable to levels observed in young mice as reported in previous publications [53, 56]) when compared with age-matched controls (representative pseudocolor flowmetry maps are shown in Figure 1A and the summary data are shown in Figure 1B). Next, we utilized 18F-FDG PET/CT imaging to measure in vivo brain glucose uptake, another critical endothelial function mediated by glucose transporters expressed on the luminal surface that regulates whole-brain energy metabolism. First, we validated 18F-FDG PET/CT imaging technique to detect age-related decreases in brain glucose uptake. In agreement with previous studies [57], we were able to demonstrate a significant age-dependent decline in brain glucose uptake (Figure 1C). Following validation of the imaging technique, we investigated whether CL treatment improved brain glucose uptake in aged animals. Consistent with improved NVC, brain glucose uptake was also significantly improved in aged animals following CL treatment (representative PET images are shown in Figure 1D and the summary data are shown in Figure 1E). Glucose uptake at the BBB is mediated by GLUT1 transporter, which has 2 isoforms: 55kDa isoform expressed in the luminal side of the BBB endothelial cells and the 45kDa isoform expressed in the astrocytic end-feet. Correlating with increased brain glucose uptake, CL treatment significantly increased GLUT1 levels in the hippocampus of the aged mice. However, interestingly only the 55kDa endothelial GLUT1 isoform was upregulated while no changes were observed with the 45kDa astrocytic isoform (Figure 1F–1K). CL-induced increase in endothelial GLUT1 was also region-specific as we did not observe similar changes in the cortex of aged mice (Figure 1F–1K). These results demonstrate that chronic β3AR treatment improved NVC and brain glucose uptake potentially mediated through endothelial GLUT1 in aged mice.
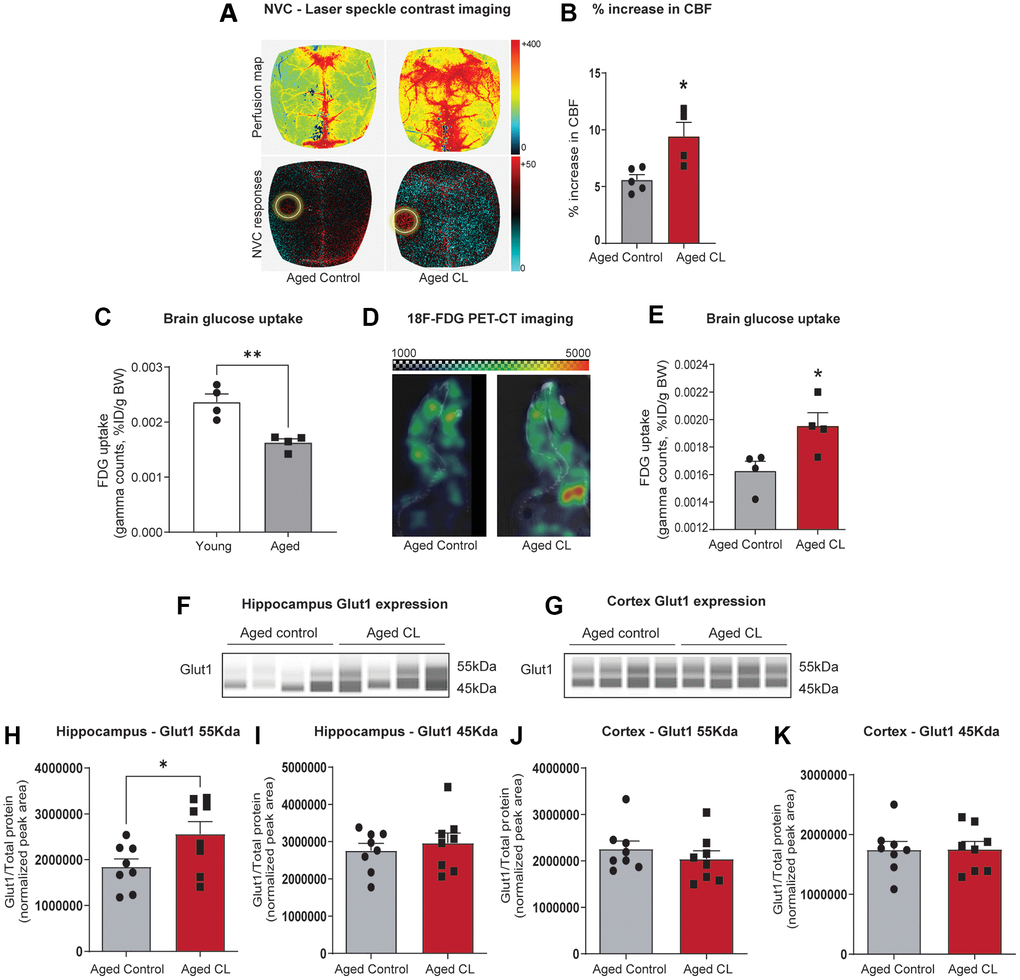
Figure 1. Effects of chronic β3-AR treatment on neurovascular coupling, brain uptake, and GLUT1 expression in aged mice. (A) Representative pseudocolor laser speckle flowmetry maps of baseline cerebral blood flow (CBF) (upper row; shown for orientation purposes) and CBF changes in the somatosensory cortex relative to baseline during contralateral whisker stimulation (bottom row, left circle, 30 s, 5 Hz) in aged mice treated with saline (aged controls) or CL 316,243 (aged CL). The color bar represents CBF as a percent change from the baseline. (B) Summary data as a % increase in CBF (n = 4–5 in each group, males). (C) 18F-FDG uptake in the young and aged brain represented as SUV (%ID/g body weight) (n = 4 in each group, males). (D) Representative 18F-FDG-PET images of aged control and CL-treated mice. Warmer colors represent higher activity in PET images. (E) Quantification of FDG uptake in the brain represented as SUV (%ID/g body weight) (n = 4 in each group, males). (F, G) Representative images of GLUT1 chemiluminescent signals for hippocampus and cortex lysates in capillaries created by the compass SW software for Jess analysis. (H–K) Peak areas for 55 and 45kDa GLUT1 isoforms normalized for total protein in the samples. Data are mean ± S.E.M. (n = 8–9 in each group, males). *P < 0.05 vs. aged controls.
Chronic β3AR stimulation attenuated BBB leakage and neuroinflammation, and increased microvascular density in aged mice
Next, we evaluated the effects of β3AR stimulation on microvascular endothelial structure which critically contributes to the maintenance of BBB integrity in aged mice. To determine BBB permeability, 3kDa FITC labeled dextran was injected intraperitoneally, and the extravasation of the injected tracer was quantified in the hippocampus and cortex tissue lysates after perfusion. CL treatment attenuated BBB leakage both in the hippocampus and cortex tissue of aged mice evident from the decreased permeability of the injected tracer in the brain parenchyma (Figure 2A, 2B).
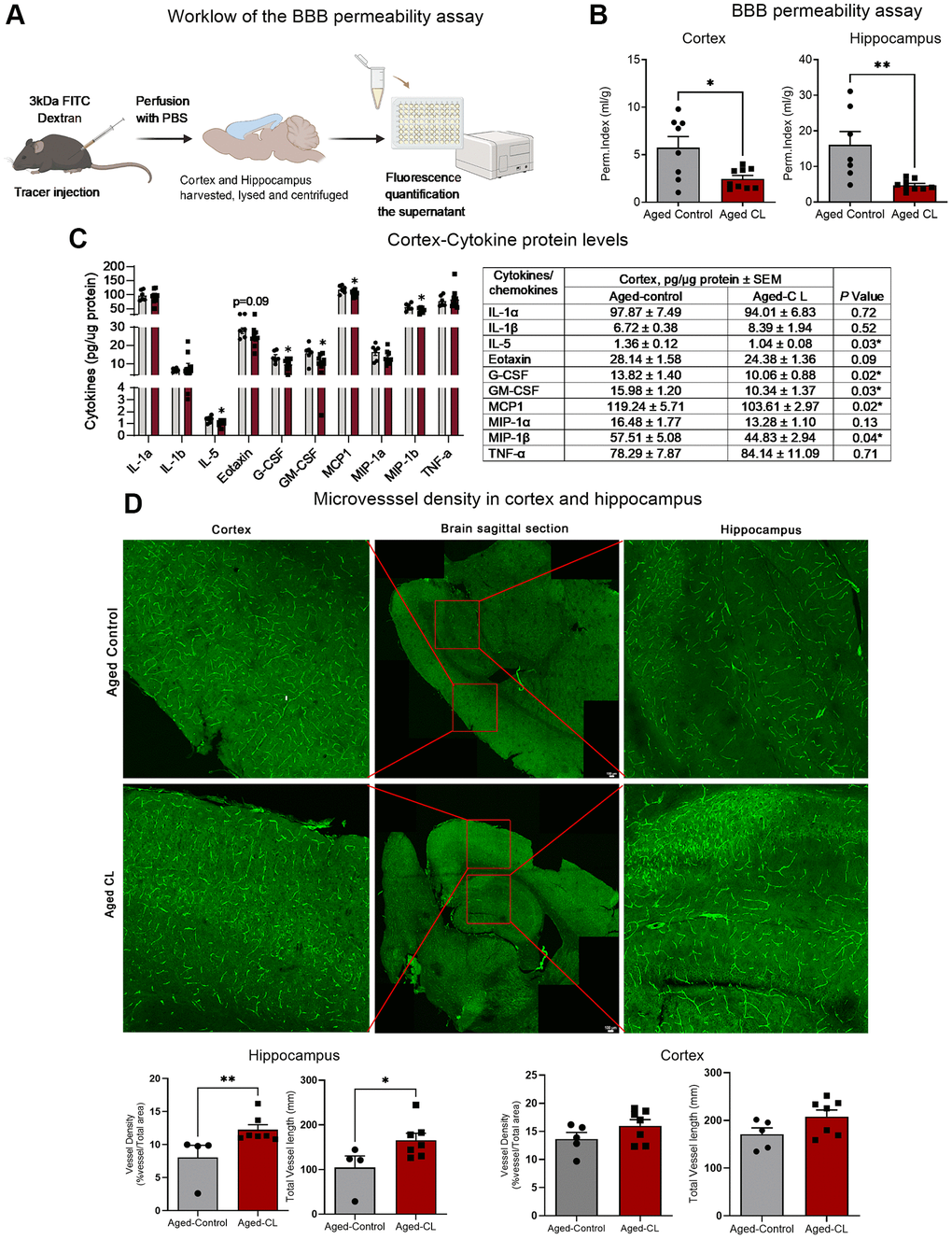
Figure 2. Effects of chronic β3-AR treatment on BBB permeability, inflammatory markers and microvascular density in aged mice. (A) Workflow representing the steps in the BBB permeability assay. (B) BBB permeability index calculated to assess the permeability of 3kDa FITC tracer in the cortex and hippocampus of aged controls and CL-treated mice (n = 7–11 in each group, males) (C) Protein levels of pro-inflammatory cytokines and chemokines assessed by multiplex magnetic assay (n = 6–9 in each group, both sexes). A table presenting the exact levels of cytokines and chemokines, normalized to total protein levels, in both the saline- and CL-treated aged mouse groups is provided. The table also includes p-values to indicate statistical differences between the two groups. (D) Representative images of brain sections stained with endomucin and CD31 in the cortex and hippocampus. Middle panel represents the whole brain picture stitched from images obtained by tiling mode. Red box indicates the region imaged at 20x magnification for quantification of the vessel density and vessel length in the cortex and the hippocampus. Bar graphs indicate the quantification data for vessel density and total vessel length analysis in the cortex and the hippocampus (n = 4–8 in each group, males). Data are mean ± S.E.M. *P < 0.05 vs. aged controls.
Intact BBB is crucial to prevent the trafficking of immune cells and other plasma proteins into the brain parenchyma. However, with aging, increased BBB permeability to blood constituents results in aberrant glial activation which ultimately leads to neuroinflammation. Hence, we wanted to assess if CL treatment mediated mitigation of BBB leakage improved inflammation in the aging brain. We assessed inflammation by measuring the protein levels of pro-inflammatory cytokines and chemokines in the cortex protein lysates. Conforming to improved BBB function, CL treatment significantly reduced the protein levels of various pro-inflammatory mediators such as IL5, Eotaxin, GCSF, GM-CSF, MIP-1b and MCP1 respectively (Figure 2C). Further, CL treatment improved the microvascular density and the total vessel length in the hippocampus, although such changes were not observed in the cortex region of aged mice (Figure 2D). These findings indicate that CL treatment significantly improved microvascular structure in aged mice.
Chronic β3AR stimulation improved spatial learning and memory in aged mice
To examine the effects of thermogenic stimulation on cognitive performance, especially spatial learning, and memory, we performed radial arm water maze test in aged mice after 6 weeks of CL treatment. First, we quantified the combined number of errors calculated across all the trials between the control and CL-treated aged mice. CL treatment significantly reduced the number of errors before reaching the target when compared to controls (Figure 3A, 3B). During the learning trials, we also observed that the mice from both groups progressively took less time to find the target suggestive of task learning (Figure 3C). On the last learning and probe trial, CL-treated aged mice took significantly less time to reach the target indicative of improved learning plasticity and memory when compared to controls. Further, a similar trend for improved relearning was also observed during the reversal trial in aged mice with CL treatment, however, they did not attain statistical significance. CL treatment did not affect swim speed during the test indicating that modulation of motor function did not contribute to the improved cognitive performance in CL-treated aged mice (Figure 3D).
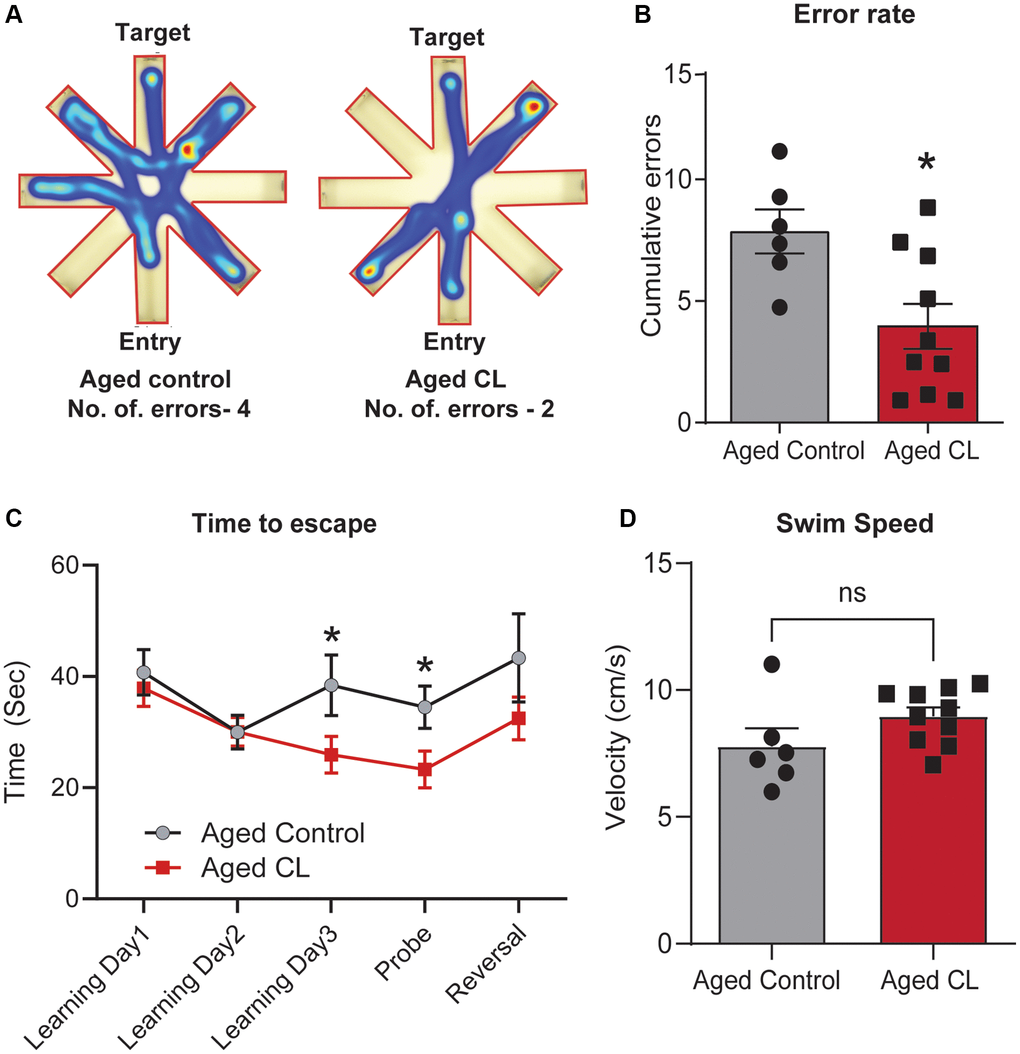
Figure 3. Effects of chronic β3-AR treatment on cognitive performance in aged mice. (A) Radial arm water maze- Heatmap showing an animal from each group that was chosen at random and the amount of time they spent in different arms and also the traces indicating the path the mice took to reach the target. Please take note that the aged controls took longer paths and made more errors in finding the target platform when compared to the CL-treated mice. (B) Cumulative errors calculated during the learning, probe, and reversal trials. (C) Time to escape calculated during each of the learning, probe and reversal day trials and (D). Swim speed (cm/s) in radial arm water maze test (n = 6–10/group, both sexes). Data are mean ± S.E.M. *P < 0.05 vs. aged controls, ns-not significant.
Discussion
Age-related metabolic diseases share strong pathogenic links with cerebromicrovascular dysfunction and cognitive impairment [58]. Supporting this idea, several epidemiological studies have demonstrated a causal association between metabolic syndrome in mid-life with decreased cerebral blood flow and cognitive decline later in life [44–47]. Adipose tissue dysfunction significantly contributes to the pathogenesis of metabolic disorders with aging through impaired glucose and lipid metabolism, altered adipokine secretion, increased secretion of pro-inflammatory mediators, and ectopic lipid deposition. Hence, interventions that improve adipose function and in turn peripheral metabolism might also confer protective effects on cerebral microvasculature and cognitive functions in aging. To test this, we chose thermogenic stimulation using β3AR agonists, a method previously well-established to improve adipose and systemic metabolism in both rodents and humans [59–62]. Although the effects of β3AR agonists on systemic metabolism have only been well-characterized in young animals, our studies showed that it also effectively improved multiple metabolic parameters including adiposity, glucose metabolism, insulin sensitivity, circulating adiponectin, and ectopic lipid deposition [48]. More importantly, these systemic improvements were associated with improved microvascular function, reduced neuroinflammation and enhanced cognition in aged mice, indicating that the beneficial effects of thermogenesis extend beyond metabolic tissues. Our results are in line with previous studies which also show that CL treatment improved brown adipose tissue thermogenesis and cognition in a triple transgenic mouse model of AD (3xTg-AD) [63] and chicks [64], albeit the mechanisms remain uncharacterized.
We posit that the mechanisms underlying the cognitive benefits of CL treatment in aging are multifactorial. First, it is highly likely that restoration of glucose uptake due to increased endothelial GLUT1 expression had a positive impact on brain energy metabolism and cognition in aging. GLUT1 is highly expressed in the brain microvascular endothelial cells [65], where it regulates BBB integrity and cerebral blood flow responses [66] in addition to supporting metabolic needs. Endothelial GLUT1 deficiency has been linked to impaired cerebral blood flow, BBB breakdown, and cognitive impairment in AD mice models [66]. Further, reduced GLUT1 expression anticipates the onset of microvascular dysfunction and clinical manifestations in mild cognitive impairment (MCI) and AD patients [67, 68], suggesting a pathogenic role for impaired endothelial glucose uptake in age-related vascular cognitive impairment. Based on these findings, it is highly likely that increased endothelial glucose uptake via GLUT1 (55kDa isoform) contributed to the restoration of BBB integrity leading to reduced neuroinflammation and improved learning and memory in aged mice. In addition, studies have also pointed to a role for GLUT1 in eNOS-mediated endothelial relaxation [69] and hence GLUT1 potentially also contributes to improved neurovascular coupling observed in CL-treated aged mice. However, the mechanistic contribution of GLUT1 deficiency to neurovascular uncoupling in aging is yet to be investigated.
Secondly, the potential role of adipose-secreted factors on cerebral microvasculature should also be considered as adipose tissue is the primary tissue target for CL. Specifically, we have observed that CL treatment significantly increased the circulating levels of adiponectin, a well-known vasoprotective adipokine with insulin-sensitizing [70] and anti-inflammatory properties [70–74]. Adiponectin has been shown to protect endothelial cells against high glucose and oxidized LDL-induced oxidative stress [75, 76], increase the production of NO by activating AMPK-eNOS signaling [11, 77, 78], and maintain capillarity and microvascular blood flow [79]. Adiponectin was also reported to inhibit atherogenesis [73] and to modulate inflammatory processes in cerebromicrovascular endothelial cells [77]. Further, studies have also established a critical role for adiponectin in the anti-aging vascular effects of caloric restriction [80, 81]. Given that adiponectin receptors (primarily AdipoR1) are expressed in brain microvascular endothelial cells [82], it raises the possibility that adiponectin could directly influence endothelial outcomes in CL-treated aged mice. It should also be noted that adipose tissue secretome is not just limited to adipokines but includes a wide repertoire of molecules such as bioactive lipids, peptides, and extracellular vesicles. Future studies should address whether CL treatment impacted these other adipose-secreted factors to modulate microvascular function and cognition in aging.
Thirdly, CL could also directly act on the brain to confer cognitive benefits in aging. The presence of β3AR mRNA has been documented in multiple brain regions [83], albeit at much lower levels than in adipose tissue. Amibegron, another β3AR agonist, has been shown to possess anxiolytic properties in rodents through modulation of neurotrophic and apoptotic pathways in the hippocampal neurons [84]. However, unlike amibegron which is BBB permeant, CL does not cross the BBB [85] and it is unlikely that CL directly influenced neuronal function. Alternatively, microvascular endothelial cells are indeed exposed to CL in circulation and are a potential target for its central actions. β3AR expression in the brain microvascular endothelial cells has not been defined yet, however, it is present and physiologically active in the coronary and retinal endothelial cells [86, 87]. In both the heart and retinal microvessels, stimulation of β3ARs induces eNOS activation and vasodilatory responses [86, 87]. Whether β3AR stimulation exerts similar actions in brain microvessels is yet to be characterized.
Although these findings are promising, several important caveats must be acknowledged. One of the major limitations in this study is that we could not consistently include both sexes in all the experiments and hence the sexual dimorphic effects of CL treatment could not be addressed. Additionally, the absence of a young control group limits the interpretation of the extent to which CL treatment improved cerebrovascular function in aged mice. Future studies incorporating both sexes and young controls will provide a more comprehensive understanding on the cerebrovascular rejuvenating effects of β3AR agonists in aging and to lay the groundwork for clinical investigations. Furthermore, this study did not address the possibility that changes in other cell types, such as pericytes, astrocytes, or vascular smooth muscle cells, may have contributed to the beneficial effects of β3AR agonists in the aging brain. Follow-up studies will focus on investigating the cell-type-specific effects of β3AR agonists in mediating improvements in NVC, BBB integrity and neuroinflammation in aging. Taken together, our findings show that chronic β3AR agonist treatment exerts robust microvascular protective effects in aged mice, which likely conferred cognitive benefits in aged mice. While β3AR agonists are being tested in clinical studies for metabolic disorders [49, 50], it could be a valuable therapeutic strategy to repurpose them for the treatment of age-related vascular cognitive impairment.
Author Contributions
Conceptualization, DN and PB; methodology and investigation, DN, SE, ST, AH, RN and VA; writing—original draft preparation, PB; review and editing, DN, MS, ST, AC, AY, VA and PB; funding acquisition, PB.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Ethical Statement
All animal experiments were approved by the Institutional Animal Care and Use Committee (Protocol#22-006) of the University of Oklahoma Health Sciences Center.
Funding
This work was supported by grants from the NIH (K01AG073613), American Heart Association (CDA1048544), The Irene Diamond Fund/AFAR Postdoctoral Transition Awards in Aging Supplement award, Presbyterian Health Foundation, and OUHSC College of Medicine Alumni Association (COMAA) to PB. This research was conducted while Priya Balasubramanian was an AFAR Grant for Junior Faculty awardee, supported by the WoodNext Foundation, a component fund administered by Greater Houston Community Foundation.
References
- 1. Wolters FJ, Zonneveld HI, Hofman A, van der Lugt A, Koudstaal PJ, Vernooij MW, Ikram MA, and Heart-Brain Connection Collaborative Research Group. Cerebral Perfusion and the Risk of Dementia: A Population-Based Study. Circulation. 2017; 136:719–28. https://doi.org/10.1161/CIRCULATIONAHA.117.027448 [PubMed]
- 2. Sorond FA, Kiely DK, Galica A, Moscufo N, Serrador JM, Iloputaife I, Egorova S, Dell'Oglio E, Meier DS, Newton E, Milberg WP, Guttmann CR, Lipsitz LA. Neurovascular coupling is impaired in slow walkers: the MOBILIZE Boston Study. Ann Neurol. 2011; 70:213–20. https://doi.org/10.1002/ana.22433 [PubMed]
- 3. Sorond FA, Schnyer DM, Serrador JM, Milberg WP, Lipsitz LA. Cerebral blood flow regulation during cognitive tasks: effects of healthy aging. Cortex. 2008; 44:179–84. https://doi.org/10.1016/j.cortex.2006.01.003 [PubMed]
- 4. Dichgans M, Leys D. Vascular Cognitive Impairment. Circ Res. 2017; 120:573–91. https://doi.org/10.1161/CIRCRESAHA.116.308426 [PubMed]
- 5. O'Brien JT, Erkinjuntti T, Reisberg B, Roman G, Sawada T, Pantoni L, Bowler JV, Ballard C, DeCarli C, Gorelick PB, Rockwood K, Burns A, Gauthier S, DeKosky ST. Vascular cognitive impairment. Lancet Neurol. 2003; 2:89–98. https://doi.org/10.1016/s1474-4422(03)00305-3 [PubMed]
- 6. Chen BR, Kozberg MG, Bouchard MB, Shaik MA, Hillman EM. A critical role for the vascular endothelium in functional neurovascular coupling in the brain. J Am Heart Assoc. 2014; 3:e000787. https://doi.org/10.1161/JAHA.114.000787 [PubMed]
- 7. Zhu WM, Neuhaus A, Beard DJ, Sutherland BA, DeLuca GC. Neurovascular coupling mechanisms in health and neurovascular uncoupling in Alzheimer's disease. Brain. 2022; 145:2276–92. https://doi.org/10.1093/brain/awac174 [PubMed]
- 8. Veys K, Fan Z, Ghobrial M, Bouché A, García-Caballero M, Vriens K, Conchinha NV, Seuwen A, Schlegel F, Gorski T, Crabbé M, Gilardoni P, Ardicoglu R, et al. Role of the GLUT1 Glucose Transporter in Postnatal CNS Angiogenesis and Blood-Brain Barrier Integrity. Circ Res. 2020; 127:466–82. https://doi.org/10.1161/CIRCRESAHA.119.316463 [PubMed]
- 9. Zhao Z, Nelson AR, Betsholtz C, Zlokovic BV. Establishment and Dysfunction of the Blood-Brain Barrier. Cell. 2015; 163:1064–78. https://doi.org/10.1016/j.cell.2015.10.067 [PubMed]
- 10. Daneman R, Prat A. The blood-brain barrier. Cold Spring Harb Perspect Biol. 2015; 7:a020412. https://doi.org/10.1101/cshperspect.a020412 [PubMed]
- 11. Ungvari Z, Tarantini S, Kiss T, Wren JD, Giles CB, Griffin CT, Murfee WL, Pacher P, Csiszar A. Endothelial dysfunction and angiogenesis impairment in the ageing vasculature. Nat Rev Cardiol. 2018; 15:555–65. https://doi.org/10.1038/s41569-018-0030-z [PubMed]
- 12. Tarantini S, Valcarcel-Ares MN, Toth P, Yabluchanskiy A, Tucsek Z, Kiss T, Hertelendy P, Kinter M, Ballabh P, Süle Z, Farkas E, Baur JA, Sinclair DA, et al. Nicotinamide mononucleotide (NMN) supplementation rescues cerebromicrovascular endothelial function and neurovascular coupling responses and improves cognitive function in aged mice. Redox Biol. 2019; 24:101192. https://doi.org/10.1016/j.redox.2019.101192 [PubMed]
- 13. Tarantini S, Tran CHT, Gordon GR, Ungvari Z, Csiszar A. Impaired neurovascular coupling in aging and Alzheimer's disease: Contribution of astrocyte dysfunction and endothelial impairment to cognitive decline. Exp Gerontol. 2017; 94:52–8. https://doi.org/10.1016/j.exger.2016.11.004 [PubMed]
- 14. Sakamuri SS, Sure VN, Kolli L, Evans WR, Sperling JA, Bix GJ, Wang X, Atochin DN, Murfee WL, Mostany R, Katakam PV. Aging related impairment of brain microvascular bioenergetics involves oxidative phosphorylation and glycolytic pathways. J Cereb Blood Flow Metab. 2022; 42:1410–24. https://doi.org/10.1177/0271678X211069266 [PubMed]
- 15. Zuendorf G, Kerrouche N, Herholz K, Baron JC. Efficient principal component analysis for multivariate 3D voxel-based mapping of brain functional imaging data sets as applied to FDG-PET and normal aging. Hum Brain Mapp. 2003; 18:13–21. https://doi.org/10.1002/hbm.10069 [PubMed]
- 16. Nyúl-Tóth Á, Tarantini S, DelFavero J, Yan F, Balasubramanian P, Yabluchanskiy A, Ahire C, Kiss T, Csipo T, Lipecz A, Farkas AE, Wilhelm I, Krizbai IA, et al. Demonstration of age-related blood-brain barrier disruption and cerebromicrovascular rarefaction in mice by longitudinal intravital two-photon microscopy and optical coherence tomography. Am J Physiol Heart Circ Physiol. 2021; 320:H1370–92. https://doi.org/10.1152/ajpheart.00709.2020 [PubMed]
- 17. Tucsek Z, Toth P, Sosnowska D, Gautam T, Mitschelen M, Koller A, Szalai G, Sonntag WE, Ungvari Z, Csiszar A. Obesity in aging exacerbates blood-brain barrier disruption, neuroinflammation, and oxidative stress in the mouse hippocampus: effects on expression of genes involved in beta-amyloid generation and Alzheimer's disease. J Gerontol A Biol Sci Med Sci. 2014; 69:1212–26. https://doi.org/10.1093/gerona/glt177 [PubMed]
- 18. Tucsek Z, Toth P, Tarantini S, Sosnowska D, Gautam T, Warrington JP, Giles CB, Wren JD, Koller A, Ballabh P, Sonntag WE, Ungvari Z, Csiszar A. Aging exacerbates obesity-induced cerebromicrovascular rarefaction, neurovascular uncoupling, and cognitive decline in mice. J Gerontol A Biol Sci Med Sci. 2014; 69:1339–52. https://doi.org/10.1093/gerona/glu080 [PubMed]
- 19. Li Y, Choi WJ, Wei W, Song S, Zhang Q, Liu J, Wang RK. Aging-associated changes in cerebral vasculature and blood flow as determined by quantitative optical coherence tomography angiography. Neurobiol Aging. 2018; 70:148–59. https://doi.org/10.1016/j.neurobiolaging.2018.06.017 [PubMed]
- 20. Kiss T, Nyúl-Tóth Á, Gulej R, Tarantini S, Csipo T, Mukli P, Ungvari A, Balasubramanian P, Yabluchanskiy A, Benyo Z, Conley SM, Wren JD, Garman L, et al. Old blood from heterochronic parabionts accelerates vascular aging in young mice: transcriptomic signature of pathologic smooth muscle remodeling. Geroscience. 2022; 44:953–81. https://doi.org/10.1007/s11357-022-00519-1 [PubMed]
- 21. Katsimpardi L, Litterman NK, Schein PA, Miller CM, Loffredo FS, Wojtkiewicz GR, Chen JW, Lee RT, Wagers AJ, Rubin LL. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science. 2014; 344:630–4. https://doi.org/10.1126/science.1251141 [PubMed]
- 22. Ximerakis M, Holton KM, Giadone RM, Ozek C, Saxena M, Santiago S, Adiconis X, Dionne D, Nguyen L, Shah KM, Goldstein JM, Gasperini C, Gampierakis IA, et al. Heterochronic parabiosis reprograms the mouse brain transcriptome by shifting aging signatures in multiple cell types. Nat Aging. 2023; 3:327–45. https://doi.org/10.1038/s43587-023-00373-6 [PubMed]
- 23. Chen MB, Yang AC, Yousef H, Lee D, Chen W, Schaum N, Lehallier B, Quake SR, Wyss-Coray T. Brain Endothelial Cells Are Exquisite Sensors of Age-Related Circulatory Cues. Cell Rep. 2020; 30:4418–32.e4. https://doi.org/10.1016/j.celrep.2020.03.012 [PubMed]
- 24. Gulej R, Nyúl-Tóth Á, Csik B, Patai R, Petersen B, Negri S, Chandragiri SS, Shanmugarama S, Mukli P, Yabluchanskiy A, Conley S, Huffman D, Tarantini S, et al. Young blood-mediated cerebromicrovascular rejuvenation through heterochronic parabiosis: enhancing blood-brain barrier integrity and capillarization in the aged mouse brain. Geroscience. 2024; 46:4415–42. https://doi.org/10.1007/s11357-024-01154-8 [PubMed]
- 25. Avogaro A, de Kreutzenberg SV, Federici M, Fadini GP. The endothelium abridges insulin resistance to premature aging. J Am Heart Assoc. 2013; 2:e000262. https://doi.org/10.1161/JAHA.113.000262 [PubMed]
- 26. Kim JA, Montagnani M, Chandrasekran S, Quon MJ. Role of lipotoxicity in endothelial dysfunction. Heart Fail Clin. 2012; 8:589–607. https://doi.org/10.1016/j.hfc.2012.06.012 [PubMed]
- 27. Stehouwer CDA. Microvascular Dysfunction and Hyperglycemia: A Vicious Cycle With Widespread Consequences. Diabetes. 2018; 67:1729–41. https://doi.org/10.2337/dbi17-0044 [PubMed]
- 28. Tangestani Fard M, Stough C. A Review and Hypothesized Model of the Mechanisms That Underpin the Relationship Between Inflammation and Cognition in the Elderly. Front Aging Neurosci. 2019; 11:56. https://doi.org/10.3389/fnagi.2019.00056 [PubMed]
- 29. Takechi R, Lam V, Brook E, Giles C, Fimognari N, Mooranian A, Al-Salami H, Coulson SH, Nesbit M, Mamo JCL. Blood-Brain Barrier Dysfunction Precedes Cognitive Decline and Neurodegeneration in Diabetic Insulin Resistant Mouse Model: An Implication for Causal Link. Front Aging Neurosci. 2017; 9:399. https://doi.org/10.3389/fnagi.2017.00399 [PubMed]
- 30. Balasubramanian P, Kiss T, Tarantini S, Nyúl-Tóth Á, Ahire C, Yabluchanskiy A, Csipo T, Lipecz A, Tabak A, Institoris A, Csiszar A, Ungvari Z. Obesity-induced cognitive impairment in older adults: a microvascular perspective. Am J Physiol Heart Circ Physiol. 2021; 320:H740–61. https://doi.org/10.1152/ajpheart.00736.2020 [PubMed]
- 31. Pasqualini L, Schillaci G, Innocente S, Pucci G, Coscia F, Siepi D, Lupattelli G, Ciuffetti G, Mannarino E. Lifestyle intervention improves microvascular reactivity and increases serum adiponectin in overweight hypertensive patients. Nutr Metab Cardiovasc Dis. 2010; 20:87–92. https://doi.org/10.1016/j.numecd.2009.03.002 [PubMed]
- 32. Csipo T, Fulop GA, Lipecz A, Tarantini S, Kiss T, Balasubramanian P, Csiszar A, Ungvari Z, Yabluchanskiy A. Short-term weight loss reverses obesity-induced microvascular endothelial dysfunction. Geroscience. 2018; 40:337–46. https://doi.org/10.1007/s11357-018-0028-9 [PubMed]
- 33. Miller KN, Burhans MS, Clark JP, Howell PR, Polewski MA, DeMuth TM, Eliceiri KW, Lindstrom MJ, Ntambi JM, Anderson RM. Aging and caloric restriction impact adipose tissue, adiponectin, and circulating lipids. Aging Cell. 2017; 16:497–507. https://doi.org/10.1111/acel.12575 [PubMed]
- 34. Xue M, Xu W, Ou YN, Cao XP, Tan MS, Tan L, Yu JT. Diabetes mellitus and risks of cognitive impairment and dementia: A systematic review and meta-analysis of 144 prospective studies. Ageing Res Rev. 2019; 55:100944. https://doi.org/10.1016/j.arr.2019.100944 [PubMed]
- 35. Atti AR, Valente S, Iodice A, Caramella I, Ferrari B, Albert U, Mandelli L, De Ronchi D. Metabolic Syndrome, Mild Cognitive Impairment, and Dementia: A Meta-Analysis of Longitudinal Studies. Am J Geriatr Psychiatry. 2019; 27:625–37. https://doi.org/10.1016/j.jagp.2019.01.214 [PubMed]
- 36. Pal K, Mukadam N, Petersen I, Cooper C. Mild cognitive impairment and progression to dementia in people with diabetes, prediabetes and metabolic syndrome: a systematic review and meta-analysis. Soc Psychiatry Psychiatr Epidemiol. 2018; 53:1149–60. https://doi.org/10.1007/s00127-018-1581-3 [PubMed]
- 37. Wee J, Sukudom S, Bhat S, Marklund M, Peiris NJ, Hoyos CM, Patel S, Naismith SL, Dwivedi G, Misra A. The relationship between midlife dyslipidemia and lifetime incidence of dementia: A systematic review and meta-analysis of cohort studies. Alzheimers Dement (Amst). 2023; 15:e12395. https://doi.org/10.1002/dad2.12395 [PubMed]
- 38. Stout MB, Justice JN, Nicklas BJ, Kirkland JL. Physiological Aging: Links Among Adipose Tissue Dysfunction, Diabetes, and Frailty. Physiology (Bethesda). 2017; 32:9–19. https://doi.org/10.1152/physiol.00012.2016 [PubMed]
- 39. Palmer AK, Kirkland JL. Aging and adipose tissue: potential interventions for diabetes and regenerative medicine. Exp Gerontol. 2016; 86:97–105. https://doi.org/10.1016/j.exger.2016.02.013 [PubMed]
- 40. Muzumdar R, Allison DB, Huffman DM, Ma X, Atzmon G, Einstein FH, Fishman S, Poduval AD, McVei T, Keith SW, Barzilai N. Visceral adipose tissue modulates mammalian longevity. Aging Cell. 2008; 7:438–40. https://doi.org/10.1111/j.1474-9726.2008.00391.x [PubMed]
- 41. Mau T, O'Brien M, Ghosh AK, Miller RA, Yung R. Life-span Extension Drug Interventions Affect Adipose Tissue Inflammation in Aging. J Gerontol A Biol Sci Med Sci. 2020; 75:89–98. https://doi.org/10.1093/gerona/glz177 [PubMed]
- 42. Stout MB, Steyn FJ, Jurczak MJ, Camporez JG, Zhu Y, Hawse JR, Jurk D, Palmer AK, Xu M, Pirtskhalava T, Evans GL, de Souza Santos R, Frank AP, et al. 17α-Estradiol Alleviates Age-related Metabolic and Inflammatory Dysfunction in Male Mice Without Inducing Feminization. J Gerontol A Biol Sci Med Sci. 2017; 72:3–15. https://doi.org/10.1093/gerona/glv309 [PubMed]
- 43. Kulkarni AS, Brutsaert EF, Anghel V, Zhang K, Bloomgarden N, Pollak M, Mar JC, Hawkins M, Crandall JP, Barzilai N. Metformin regulates metabolic and nonmetabolic pathways in skeletal muscle and subcutaneous adipose tissues of older adults. Aging Cell. 2018; 17:e12723. https://doi.org/10.1111/acel.12723 [PubMed]
- 44. Kloppenborg RP, van den Berg E, Kappelle LJ, Biessels GJ. Diabetes and other vascular risk factors for dementia: which factor matters most? A systematic review. Eur J Pharmacol. 2008; 585:97–108. https://doi.org/10.1016/j.ejphar.2008.02.049 [PubMed]
- 45. Rawlings AM, Sharrett AR, Schneider AL, Coresh J, Albert M, Couper D, Griswold M, Gottesman RF, Wagenknecht LE, Windham BG, Selvin E. Diabetes in midlife and cognitive change over 20 years: a cohort study. Ann Intern Med. 2014; 161:785–93. https://doi.org/10.7326/M14-0737 [PubMed]
- 46. Barbiellini Amidei C, Fayosse A, Dumurgier J, Machado-Fragua MD, Tabak AG, van Sloten T, Kivimäki M, Dugravot A, Sabia S, Singh-Manoux A. Association Between Age at Diabetes Onset and Subsequent Risk of Dementia. JAMA. 2021; 325:1640–9. https://doi.org/10.1001/jama.2021.4001 [PubMed]
- 47. Zhang X, Tong T, Chang A, Ang TFA, Tao Q, Auerbach S, Devine S, Qiu WQ, Mez J, Massaro J, Lunetta KL, Au R, Farrer LA. Midlife lipid and glucose levels are associated with Alzheimer's disease. Alzheimers Dement. 2023; 19:181–93. https://doi.org/10.1002/alz.12641 [PubMed]
- 48. Natarajan D, Plakkot B, Tiwari K, Ekambaram S, Wang W, Rudolph M, Mohammad MA, Chacko SK, Subramanian M, Tarantini S, Yabluchanskiy A, Ungvari Z, Csiszar A, Balasubramanian P. Chronic β3-AR stimulation activates distinct thermogenic mechanisms in brown and white adipose tissue and improves systemic metabolism in aged mice. Aging Cell. 2024; 23:e14321. https://doi.org/10.1111/acel.14321 [PubMed]
- 49. Finlin BS, Memetimin H, Zhu B, Confides AL, Vekaria HJ, El Khouli RH, Johnson ZR, Westgate PM, Chen J, Morris AJ, Sullivan PG, Dupont-Versteegden EE, Kern PA. The β3-adrenergic receptor agonist mirabegron improves glucose homeostasis in obese humans. J Clin Invest. 2020; 130:2319–31. https://doi.org/10.1172/JCI134892 [PubMed]
- 50. Cypess AM, Weiner LS, Roberts-Toler C, Franquet Elía E, Kessler SH, Kahn PA, English J, Chatman K, Trauger SA, Doria A, Kolodny GM. Activation of human brown adipose tissue by a β3-adrenergic receptor agonist. Cell Metab. 2015; 21:33–8. https://doi.org/10.1016/j.cmet.2014.12.009 [PubMed]
- 51. Mottillo EP, Balasubramanian P, Lee YH, Weng C, Kershaw EE, Granneman JG. Coupling of lipolysis and de novo lipogenesis in brown, beige, and white adipose tissues during chronic β3-adrenergic receptor activation. J Lipid Res. 2014; 55:2276–86. https://doi.org/10.1194/jlr.M050005 [PubMed]
- 52. Yabluchanskiy A, Tarantini S, Balasubramanian P, Kiss T, Csipo T, Fülöp GA, Lipecz A, Ahire C, DelFavero J, Nyul-Toth A, Sonntag WE, Schwartzman ML, Campisi J, et al. Pharmacological or genetic depletion of senescent astrocytes prevents whole brain irradiation-induced impairment of neurovascular coupling responses protecting cognitive function in mice. Geroscience. 2020; 42:409–28. https://doi.org/10.1007/s11357-020-00154-8 [PubMed]
- 53. Tarantini S, Balasubramanian P, Delfavero J, Csipo T, Yabluchanskiy A, Kiss T, Nyúl-Tóth Á, Mukli P, Toth P, Ahire C, Ungvari A, Benyo Z, Csiszar A, Ungvari Z. Treatment with the BCL-2/BCL-xL inhibitor senolytic drug ABT263/Navitoclax improves functional hyperemia in aged mice. Geroscience. 2021; 43:2427–40. https://doi.org/10.1007/s11357-021-00440-z [PubMed]
- 54. Tarantini S, Hertelendy P, Tucsek Z, Valcarcel-Ares MN, Smith N, Menyhart A, Farkas E, Hodges EL, Towner R, Deak F, Sonntag WE, Csiszar A, Ungvari Z, Toth P. Pharmacologically-induced neurovascular uncoupling is associated with cognitive impairment in mice. J Cereb Blood Flow Metab. 2015; 35:1871–81. https://doi.org/10.1038/jcbfm.2015.162 [PubMed]
- 55. Devraj K, Guérit S, Macas J, Reiss Y. An In Vivo Blood-brain Barrier Permeability Assay in Mice Using Fluorescently Labeled Tracers. J Vis Exp. 2018; 57038. https://doi.org/10.3791/57038 [PubMed]
- 56. Toth P, Tarantini S, Tucsek Z, Ashpole NM, Sosnowska D, Gautam T, Ballabh P, Koller A, Sonntag WE, Csiszar A, Ungvari Z. Resveratrol treatment rescues neurovascular coupling in aged mice: role of improved cerebromicrovascular endothelial function and downregulation of NADPH oxidase. Am J Physiol Heart Circ Physiol. 2014; 306:H299–308. https://doi.org/10.1152/ajpheart.00744.2013 [PubMed]
- 57. Subtirelu RC, Teichner EM, Su Y, Al-Daoud O, Patel M, Patil S, Writer M, Werner T, Revheim ME, Høilund-Carlsen PF, Alavi A. Aging and Cerebral Glucose Metabolism: 18F-FDG-PET/CT Reveals Distinct Global and Regional Metabolic Changes in Healthy Patients. Life (Basel). 2023; 13:2044. https://doi.org/10.3390/life13102044 [PubMed]
- 58. Takeda S, Sato N, Uchio-Yamada K, Sawada K, Kunieda T, Takeuchi D, Kurinami H, Shinohara M, Rakugi H, Morishita R. Diabetes-accelerated memory dysfunction via cerebrovascular inflammation and Abeta deposition in an Alzheimer mouse model with diabetes. Proc Natl Acad Sci U S A. 2010; 107:7036–41. https://doi.org/10.1073/pnas.1000645107 [PubMed]
- 59. Ramseyer VD, Granneman JG. Adrenergic regulation of cellular plasticity in brown, beige/brite and white adipose tissues. Adipocyte. 2016; 5:119–29. https://doi.org/10.1080/21623945.2016.1145846 [PubMed]
- 60. O'Mara AE, Johnson JW, Linderman JD, Brychta RJ, McGehee S, Fletcher LA, Fink YA, Kapuria D, Cassimatis TM, Kelsey N, Cero C, Sater ZA, Piccinini F, et al. Chronic mirabegron treatment increases human brown fat, HDL cholesterol, and insulin sensitivity. J Clin Invest. 2020; 130:2209–19. https://doi.org/10.1172/JCI131126 [PubMed]
- 61. Finlin BS, Memetimin H, Confides AL, Kasza I, Zhu B, Vekaria HJ, Harfmann B, Jones KA, Johnson ZR, Westgate PM, Alexander CM, Sullivan PG, Dupont-Versteegden EE, Kern PA. Human adipose beiging in response to cold and mirabegron. JCI Insight. 2018; 3:e121510. https://doi.org/10.1172/jci.insight.121510 [PubMed]
- 62. Bel JS, Tai TC, Khaper N, Lees SJ. Mirabegron: The most promising adipose tissue beiging agent. Physiol Rep. 2021; 9:e14779. https://doi.org/10.14814/phy2.14779 [PubMed]
- 63. Tournissac M, Vu TM, Vrabic N, Hozer C, Tremblay C, Mélançon K, Planel E, Pifferi F, Calon F. Repurposing beta-3 adrenergic receptor agonists for Alzheimer's disease: beneficial effects in a mouse model. Alzheimers Res Ther. 2021; 13:103. https://doi.org/10.1186/s13195-021-00842-3 [PubMed]
- 64. Gibbs ME, Maksel D, Gibbs Z, Hou X, Summers RJ, Small DH. Memory loss caused by beta-amyloid protein is rescued by a beta(3)-adrenoceptor agonist. Neurobiol Aging. 2010; 31:614–24. https://doi.org/10.1016/j.neurobiolaging.2008.05.018 [PubMed]
- 65. Zeller K, Rahner-Welsch S, Kuschinsky W. Distribution of Glut1 glucose transporters in different brain structures compared to glucose utilization and capillary density of adult rat brains. J Cereb Blood Flow Metab. 1997; 17:204–9. https://doi.org/10.1097/00004647-199702000-00010 [PubMed]
- 66. Winkler EA, Nishida Y, Sagare AP, Rege SV, Bell RD, Perlmutter D, Sengillo JD, Hillman S, Kong P, Nelson AR, Sullivan JS, Zhao Z, Meiselman HJ, et al. GLUT1 reductions exacerbate Alzheimer's disease vasculo-neuronal dysfunction and degeneration. Nat Neurosci. 2015; 18:521–30. https://doi.org/10.1038/nn.3966 [PubMed]
- 67. Simpson IA, Chundu KR, Davies-Hill T, Honer WG, Davies P. Decreased concentrations of GLUT1 and GLUT3 glucose transporters in the brains of patients with Alzheimer's disease. Ann Neurol. 1994; 35:546–51. https://doi.org/10.1002/ana.410350507 [PubMed]
- 68. Kalaria RN, Harik SI. Reduced glucose transporter at the blood-brain barrier and in cerebral cortex in Alzheimer disease. J Neurochem. 1989; 53:1083–8. https://doi.org/10.1111/j.1471-4159.1989.tb07399.x [PubMed]
- 69. Park JL, Heilig CW, Brosius FC
3rd . GLUT1-deficient mice exhibit impaired endothelium-dependent vascular relaxation. Eur J Pharmacol. 2004; 496:213–4. https://doi.org/10.1016/j.ejphar.2004.06.022 [PubMed] - 70. Hotta K, Funahashi T, Bodkin NL, Ortmeyer HK, Arita Y, Hansen BC, Matsuzawa Y. Circulating concentrations of the adipocyte protein adiponectin are decreased in parallel with reduced insulin sensitivity during the progression to type 2 diabetes in rhesus monkeys. Diabetes. 2001; 50:1126–33. https://doi.org/10.2337/diabetes.50.5.1126 [PubMed]
- 71. Kubota N, Terauchi Y, Yamauchi T, Kubota T, Moroi M, Matsui J, Eto K, Yamashita T, Kamon J, Satoh H, Yano W, Froguel P, Nagai R, et al. Disruption of adiponectin causes insulin resistance and neointimal formation. J Biol Chem. 2002; 277:25863–6. https://doi.org/10.1074/jbc.C200251200 [PubMed]
- 72. Yamauchi T, Kamon J, Minokoshi Y, Ito Y, Waki H, Uchida S, Yamashita S, Noda M, Kita S, Ueki K, Eto K, Akanuma Y, Froguel P, et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat Med. 2002; 8:1288–95. https://doi.org/10.1038/nm788 [PubMed]
- 73. Yamauchi T, Kamon J, Waki H, Imai Y, Shimozawa N, Hioki K, Uchida S, Ito Y, Takakuwa K, Matsui J, Takata M, Eto K, Terauchi Y, et al. Globular adiponectin protected ob/ob mice from diabetes and ApoE-deficient mice from atherosclerosis. J Biol Chem. 2003; 278:2461–8. https://doi.org/10.1074/jbc.M209033200 [PubMed]
- 74. Yamauchi T, Kamon J, Waki H, Terauchi Y, Kubota N, Hara K, Mori Y, Ide T, Murakami K, Tsuboyama-Kasaoka N, Ezaki O, Akanuma Y, Gavrilova O, et al. The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med. 2001; 7:941–6. https://doi.org/10.1038/90984 [PubMed]
- 75. Motoshima H, Wu X, Mahadev K, Goldstein BJ. Adiponectin suppresses proliferation and superoxide generation and enhances eNOS activity in endothelial cells treated with oxidized LDL. Biochem Biophys Res Commun. 2004; 315:264–71. https://doi.org/10.1016/j.bbrc.2004.01.049 [PubMed]
- 76. Ouedraogo R, Wu X, Xu SQ, Fuchsel L, Motoshima H, Mahadev K, Hough K, Scalia R, Goldstein BJ. Adiponectin suppression of high-glucose-induced reactive oxygen species in vascular endothelial cells: evidence for involvement of a cAMP signaling pathway. Diabetes. 2006; 55:1840–6. https://doi.org/10.2337/db05-1174 [PubMed]
- 77. Spranger J, Verma S, Göhring I, Bobbert T, Seifert J, Sindler AL, Pfeiffer A, Hileman SM, Tschöp M, Banks WA. Adiponectin does not cross the blood-brain barrier but modifies cytokine expression of brain endothelial cells. Diabetes. 2006; 55:141–7. [PubMed]
- 78. Chen H, Montagnani M, Funahashi T, Shimomura I, Quon MJ. Adiponectin stimulates production of nitric oxide in vascular endothelial cells. J Biol Chem. 2003; 278:45021–6. https://doi.org/10.1074/jbc.M307878200 [PubMed]
- 79. Tanigawa T, Shibata R, Ouchi N, Kondo K, Ishii M, Katahira N, Kambara T, Inoue Y, Takahashi R, Ikeda N, Kihara S, Ueda H, Murohara T. Adiponectin deficiency exacerbates age-related hearing impairment. Cell Death Dis. 2014; 5:e1189. https://doi.org/10.1038/cddis.2014.140 [PubMed]
- 80. Kondo M, Shibata R, Miura R, Shimano M, Kondo K, Li P, Ohashi T, Kihara S, Maeda N, Walsh K, Ouchi N, Murohara T. Caloric restriction stimulates revascularization in response to ischemia via adiponectin-mediated activation of endothelial nitric-oxide synthase. J Biol Chem. 2009; 284:1718–24. https://doi.org/10.1074/jbc.M805301200 [PubMed]
- 81. Shinmura K, Tamaki K, Saito K, Nakano Y, Tobe T, Bolli R. Cardioprotective effects of short-term caloric restriction are mediated by adiponectin via activation of AMP-activated protein kinase. Circulation. 2007; 116:2809–17. https://doi.org/10.1161/CIRCULATIONAHA.107.725697 [PubMed]
- 82. Pratap AA, Holsinger RMD. Altered Brain Adiponectin Receptor Expression in the 5XFAD Mouse Model of Alzheimer's Disease. Pharmaceuticals (Basel). 2020; 13:150. https://doi.org/10.3390/ph13070150 [PubMed]
- 83. Summers RJ, Papaioannou M, Harris S, Evans BA. Expression of beta 3-adrenoceptor mRNA in rat brain. Br J Pharmacol. 1995; 116:2547–8. https://doi.org/10.1111/j.1476-5381.1995.tb17205.x [PubMed]
- 84. Tamburella A, Micale V, Leggio GM, Drago F. The beta3 adrenoceptor agonist, amibegron (SR58611A) counteracts stress-induced behavioral and neurochemical changes. Eur Neuropsychopharmacol. 2010; 20:704–13. https://doi.org/10.1016/j.euroneuro.2010.04.006 [PubMed]
- 85. Mirbolooki MR, Schade KN, Constantinescu CC, Pan ML, Mukherjee J. Enhancement of 18F-fluorodeoxyglucose metabolism in rat brain frontal cortex using a β3 adrenoceptor agonist. Synapse. 2015; 69:96–8. https://doi.org/10.1002/syn.21789 [PubMed]
- 86. Dessy C, Moniotte S, Ghisdal P, Havaux X, Noirhomme P, Balligand JL. Endothelial beta3-adrenoceptors mediate vasorelaxation of human coronary microarteries through nitric oxide and endothelium-dependent hyperpolarization. Circulation. 2004; 110:948–54. https://doi.org/10.1161/01.CIR.0000139331.85766.AF [PubMed]
- 87. Dal Monte M, Filippi L, Bagnoli P. Beta3-adrenergic receptors modulate vascular endothelial growth factor release in response to hypoxia through the nitric oxide pathway in mouse retinal explants. Naunyn Schmiedebergs Arch Pharmacol. 2013; 386:269–78. https://doi.org/10.1007/s00210-012-0828-x [PubMed]