



Introduction
Reproductive aging
In middle-aged female (MA) rats, both the reproductive capacity and frequency of regular estrous cycles are decreased compared to their young adult counterparts [1]. The age-associated changes in estrous cyclicity observed in MA rats are associated to a delay as well as an attenuation of the preovulatory Luteinizing hormone (LH) surge [2], increased LH and follicle-stimulating hormone (FSH) serum levels, and altered circulating estradiol profiles with absence of the surge in the noon of the proestrus with respect to the young rats, [3]. There is clear evidence that in these animals, ovarian failure is preceded by the appearance of a diminished response of the neuroendocrine axis to positive feedback from estradiol [4, 5]. Our group has implemented strategies capable of prolonging reproductive function in female rats. Thus, in female MA rats we have previously demonstrated that intrahypothalamic gene therapy for insulin-like growth factor-I (IGF-I) started at 8 months of age extends the regular cyclicity of the animals beyond 10 months (the age at which MA rats stop ovulating) and preserves the integrity of ovarian structure. At 11 months of age, the treated rats showed, in general, a cyclicity preserved in its regularity as well as a normal ovarian histology, while the controls were, at the same age, mostly acyclic and exhibited a high percentage of polycystic ovaries and few corpora lutea [6].
Aging and epigenetics
The discovery of animal cloning [7, 8] and the subsequent development of cell reprogramming technology by means of the four reprogramming factors, Oct4, Sox2, Klf4, c-Myc (OSKM), also known as the Yamanaka genes, [9] ushered in a technological and conceptual revolution that led to the achievement of cell rejuvenation by full reprogramming and to the emerging view of aging as a reversible epigenetic process where cumulative DNA damage does not appear to play a central role as was thought for decades [10].
Regenerative effects of Yamanaka gene therapy
It is of significant interest that Yamanaka gene therapy in the retina of an experimental-glaucoma mouse model and in middle-aged mice ameliorated their visual acuity [11]. In progeric mice, transgenic for OSKM factors, cyclic partial reprogramming attenuated several signs of aging in visceral tissues and extended by 50% the survival of experimental versus control counterparts [12]. Furthermore, in a recent report [13] it was shown that intravenous gene therapy with a regulatable AAV9 vector system that expresses the OSK genes, implemented in senile mice (29.2 months), prolonged the survival of the animals by two months compared to control counterparts. This is the first report showing that Yamanaka gene therapy in WT mice can extend their survival. We have recently reported that 39-day OSKM gene therapy in the dorsal hippocampus of old rats significantly reverses the typical learning deficits displayed in aged rats [14].
This line of evidence prompted us to implement OSKM gene therapy in the hypothalamus of young female rats using an adenovector constructed by us that expresses a polycistronic cassette (the STEMCCA system) harboring the OSKM genes as well as the GFP gene, all under the control of a bidirectional regulatable promoter [15]. We hypothesized that long-term expression of the OSKM genes in the hypothalamus of young female rats could slow down the rate of decline of fertility in the animals as they near the age of ovulatory cessation (10 months in our rat colony). We report here that 9 months old female rats submitted to hypothalamic OSKM-GFP gene therapy at 4 months of age, display a higher fertility than control vector-treated counterparts and remain cycling regularly at the age of estropause (10 months).
Materials and Methods
Adenoviral vectors
Construction of a regulatable HD-recombinant adenoviral vector (RAd) Tet-Off adenovector harboring the GFP and Yamanaka genes
The adenovector was constructed using a commercial kit (Microbix Inc., Ontario, Canada) that provides the shuttle plasmid pC4HSU, the helper adenovirus H14 and the HEK293 Cre4 cell line. A full description of the adenovector constructed has been already documented [15]. Briefly, we cloned a construct harboring the bicistronic tandem Oct4-f2A-Klf4-ires-Sox2-p2A-cMyc (known as hSTEMCCA, generously provided by Dr. G. Mostoslavsky, Boston University) under the control of the bidirectional promoter PminCMV-TRE-PminCMV a regulatable Tet-Off promoter which on its second end is flanked by the gene for humanized green fluorescent protein (hGFP).
The hSTEMCCA cassette harbors the 4 Yamanaka genes grouped in pairs placed downstream and upstream of an internal ribosome entry site (IRES). In turn, each pair of genes is separated by a type 2A CHYSEL (cis-acting hydrolase element) self-processing short sequence which causes the ribosome to skip the Gly-Pro bond at the C-terminal end of the 2A sequence thus releasing the peptide upstream the 2A element but continuing with the translation of the downstream mRNA sequence. This allows near stoichiometric co-expression of the two cistrons flanking a 2A-type sequence [16]. The whole expression cassette (STEMCCA cassette 10,065 bp) was cloned into the pC4HSU HD shuttle at the AscI and SwaI sites giving rise to pC4HSU-STEMCCA-tTA (Figure 1). The pC4HSU HD shuttle consists of the inverted terminal repeats (ITRs) for Ad 5 virus, the packaging signal and part of the E4 adenoviral region plus a stuffer non-coding DNA of human origin which keeps a suitable size (28–31 Kbp) of the viral DNA.
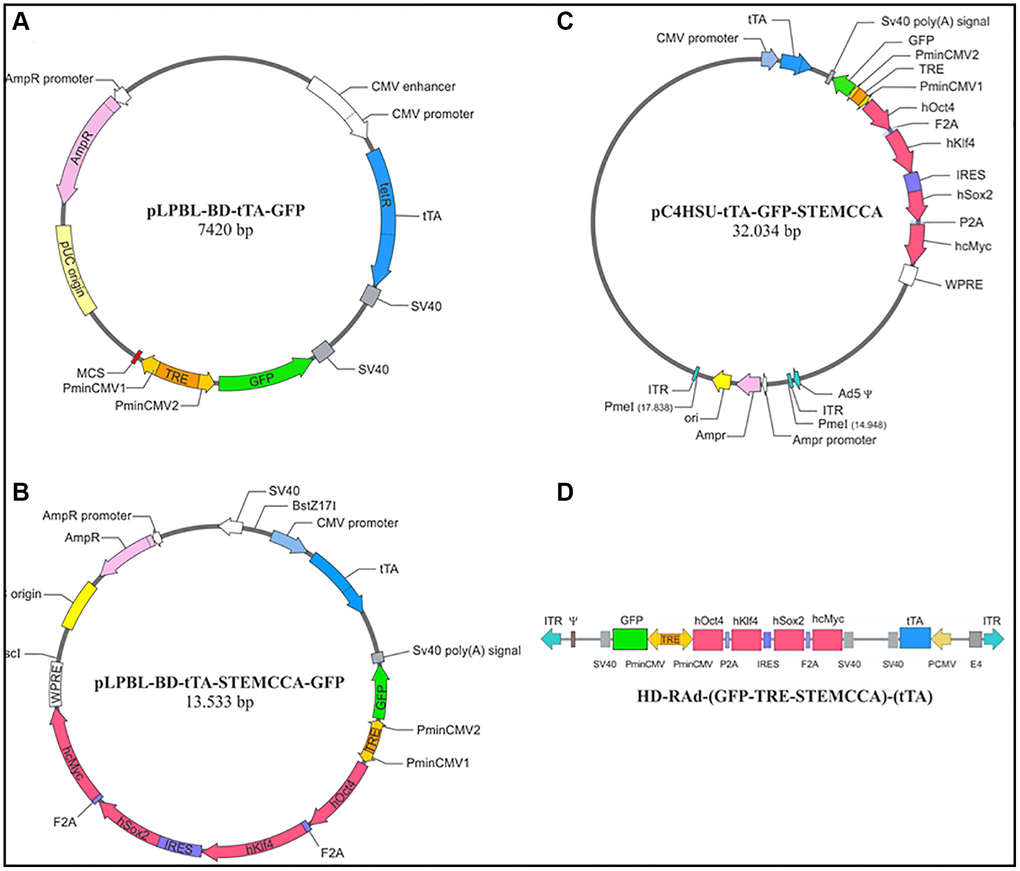
Figure 1. Diagrammatic representation of main intermediate constructs generated during the construction of HD-RAd-STEMCCA-GFP-Tet-Off. (A) Shows the base plasmid, pLPBL-BD-tTA-GFP, from which we departed in the generation of the above adenovector. It harbors the CMV1-TRE-CMV2 regulatable promoter flanked on one side by the gene for GFP and by a multiple cloning site (MCS) on the other side. Separately, the plasmid also harbors the gene for the chimeric tetracycline transactivator (tTA) protein, created by fusing one protein, TetR (tetracycline repressor), with the activation domain VP16, from the Herpes Simplex Virus. This construct is under the control of the human cytomegalovirus (CMV) promoter. (B) Displays the above plasmid now harboring the STEMCCA tandem, which was cloned into the MSC site. (C) Shows the full STEMCCA construct (including all associated regulatory components) cloned into the HD genomic plasmid which includes a bacterial sequence between its ITRs, flanked by PmeI sites. (D) Illustrates the basic components of HD-RAd-STEMCCA-GFP-Tet-Off genome after removing the bacterial sequence from pC4HSU-tTA-STEMCCA-GFP. For further details see Figure 2A legend and Ref. 15.4.
The linearized DNA backbone of the new HD-RAd (Figure 2A) was transfected in Cre 293 cells. For expansion, the helper H14 adenovirus was added to the cell cultures at a multiplicity of infection (MOI) of 5. In H14, the packaging signal is flanked by lox P sites recognized by the Cre recombinase expressed by the 293 Cre4 cells. Therefore, the helper virus provides in trans all of the viral products necessary for generation of the desired HD-RAd. The infected 293 Cre4 cells were left for 2–3 days until cytopathic effect (CPE) was evident. Cells and medium were collected and submitted to 3 freeze-thaw cycles to lyse them. Clear lysates were obtained, mixed with H14 helper virus and added to a fresh culture of Cre4 293 cells at MOI 1. When CPE appeared, passage 2 (P2) cell lysates were prepared. This iterative co-infection process was carried on five more times in order to generate enough HD-RAd particles as to proceed to the purification step. The newly generated HD-RAd was rescued from P5. The generated HD-RAd was purified by ultracentrifugation in CsCl gradients. Final virus stock was titrated by lysing the viral particles, extracting their DNA and determining its concentration in a Nanodrop spectrophotometer. For the first preparation, titer was 12 × 1011 physical viral particles (pvp)/ml.
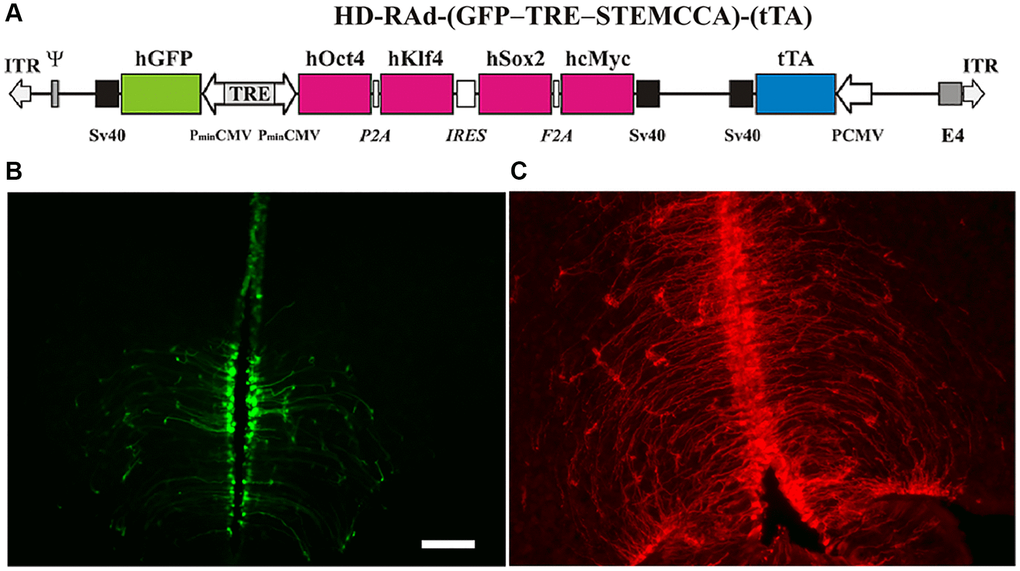
Figure 2. (A) The figure illustrates the basic components of HD-RAd-STEMCCA-GFP-Tet-Off genome. Abbreviations: GFP: humanized Green Fluorescent Protein; TRE: Tetracycline responsive element; tTA: chimeric regulatory protein; PminCMV: cytomegalovirus minimal promoter; SV40pA: polyadenylation signal; ITR: inverted terminal repeats; ψ: packaging signal. (B) Expression of GFP from the adenovector HD-RAd-STEMCCA-GFP in the third ventricle of the rat hypothalamus 6 days after vector injection in the lateral ventricles. Notice the processes of the ependymal tanycytes forming an extensive network in the hypothalamic parenchyma. Scale bar, 50 µm (for both, B, C). (C) Expression of DsRed2 from the control adenovector HD-RAd-DsRed2 in the rat hypothalamus 30 days after vector injection. Notice the processes of the ependymal tanycytes forming an extensive network in the hypothalamic parenchyma.
We also constructed two control regulatable HD-RAd Tet-Off harboring either the hGFP gene or the gene for the DsRed2, red fluorescent protein isolated from Discosoma sp.
Animals and in vivo procedures
Female Sprague-Dawley rats aged 3–4, 10 and 26 months were used. The animals were raised in our institution (INIBIOLP) and housed in a temperature-controlled room (22 ± 2ºC) on a 12:12 h light/dark cycle (lights on from 7 to 19 o’clock). Food and water were available ad libitum. All experiments with animals were performed according to the Animal Welfare Guidelines of NIH (INIBIOLP’s Animal Welfare Assurance No A5647-01).
Stereotaxic injections
Rats were anesthetized with ketamine hydrochloride (40 mg/kg; ip) plus xylazine (8 mg/kg; im) and placed on a stereotaxic apparatus. In order to access the mediobasal hypothalamus (MBH) the tip of a 26G needle fitted to a 10 µl syringe was brought to the following coordinates relative to the bregma: 3.0 mm posterior, 8.0 mm ventral and 0.6 mm right and left [17] and a suspension of 1.5 µl OSKM or control vector was bilaterally injected.
Vaginal smears
Vaginal secretion was collected daily, between 11 and 13 o’clock, with a plastic pipette filled with 20 µl normal saline (NaCl 0.9%) by inserting the tip into the rat vagina, but not deeply. A drop of vaginal fluid was smeared on a glass slide and the unstained material was observed under a light microscope, with a 40X phase-contrast objective. Three types of cells can be recognized: round and nucleated ones are epithelial cells; irregular ones without nucleus are cornified cells; and little round ones are leukocytes. The proportion among them was used for determination of the estrous cycle phases [18, 19], which are indicated as follows, P, proestrus; E, estrus, D1, diestrus 1; D2, diestrus 2.
In MA rats spending several days in a row in constant estrus (CE), a CE cycle was defined, for quantitation purposes, as a period of 5 consecutive days of vaginal smears showing only cornified cells. For instance, if an animal spent 13 days in a row in CE they were counted as 13/5 = 2.6 CE cycles.
Assessment of the effect of age on cycle regularity
Females aged 3, 10 and 26 months were submitted to daily vaginal smears sampling for 30 days.
Experimental design for long-term OSKM-GFP gene therapy in young females
Four-month-old cycling females were allotted to a control or experimental group of 12 animals each, thus forming 2 groups: Control, HD-RAd-GFP-injected control (GFP group) and HD-RAd-OSKM-GFP-injected experimental (OSKM group).
When the GFP and OSKM rats reached the age of 9 months and 10 days (40 weeks), a group of 12 intact female rats (3–4 months) was added (Y group). In each experimental group the 12 animals were divided into 4-rat subgroups, each of which was mated for one week with a young male rat (3–4 months). At the end of the pregnancy period pregnant rats were placed in single cages the number of pups per litter was counted and the newborn rats were weighed. At weaning, pups were weighed again in order to compare their BW gain.
At age 10 months, a sample of 6 nonpregnant OSKM-treated rats was submitted to vaginal smear assessment in order to determine their estrous cycle regularity.
Hypothalamic section assessment by fluorescence microscopy
Some of the rats were used for immunofluorescence studies in the hypothalamus. Animals were placed under deep anesthesia and perfused with phosphate buffered formaldehyde 4%, (pH 7.4) fixative. Each brain was removed and trimmed down to a block containing the whole hypothalamus. The block was then serially cut into coronal sections 40 µm thick on a vibratome (Leica, Nussloch, Germany).
Hypothalamic brain sections were mounted with Fluoromount water-soluble mounting medium. Images of hypothalamic and other brain sections were captured using a confocal Olympus FV1000 microscope (Tokyo, Japan). Digital images were analyzed using the ImagePro Plus (IPP™) v6.3.
Statistical analysis
The t-test was used to evaluate group differences.
Results
HD-RAd OSKM-GFP expression in the hypothalamus of young rats
Both, the HD-RAd OSKM-GFP and the control HD-RAd vectors were stereotaxically injected in the MBH of young female rats. Six days after injection, hGFP or DsRed expression was strong. The processes of the ependymal tanycytes form an extensive network in the hypothalamic parenchyma (Figure 2B). When our control vector HD-RAd-DsRed2 was injected into the hypothalamic parenchyma of young rats, DsRed 2 expression remained high for at least 30 days (Figure 2C).
Characterization of estrous cycle patterns throughout the lifespan
Using intact animals from our female rat colony, we performed an initial characterization of estrous cycle patterns from youth through very old age. As expected, the results showed that transition from regular to irregular cyclicity takes place at around 9 months of age and is followed by a prevalence of CE status from age 10 to 18–20 months (Figure 3). Animals older than 20 months progressively go into a constant diestrus phase. Thus, 3-month-old females typically show 4–5-day estrous cycles characterized by one day or less of P (in some cycles P was so short that when vaginal smear was performed the cell proportion was consistent with proestrus entering estrus or sometimes just estrus), one-two days of E and 2–3 days of D1 and/or D2. Ten-month old females typically show a prevalence of lengthy CE periods with interspersed irregular cycles (Figure 3, middle panel). Their ovarian histology showed numerous follicular cysts and scarce CL (data not shown, see ref 6). In senescent females (26 months) CE gave way to an increasing incidence of long periods of D with occasional 2–3 day-periods of P and E vaginal cytology as well as some CE periods. Senescent ovaries showed follicular cysts and some CL (data not shown).
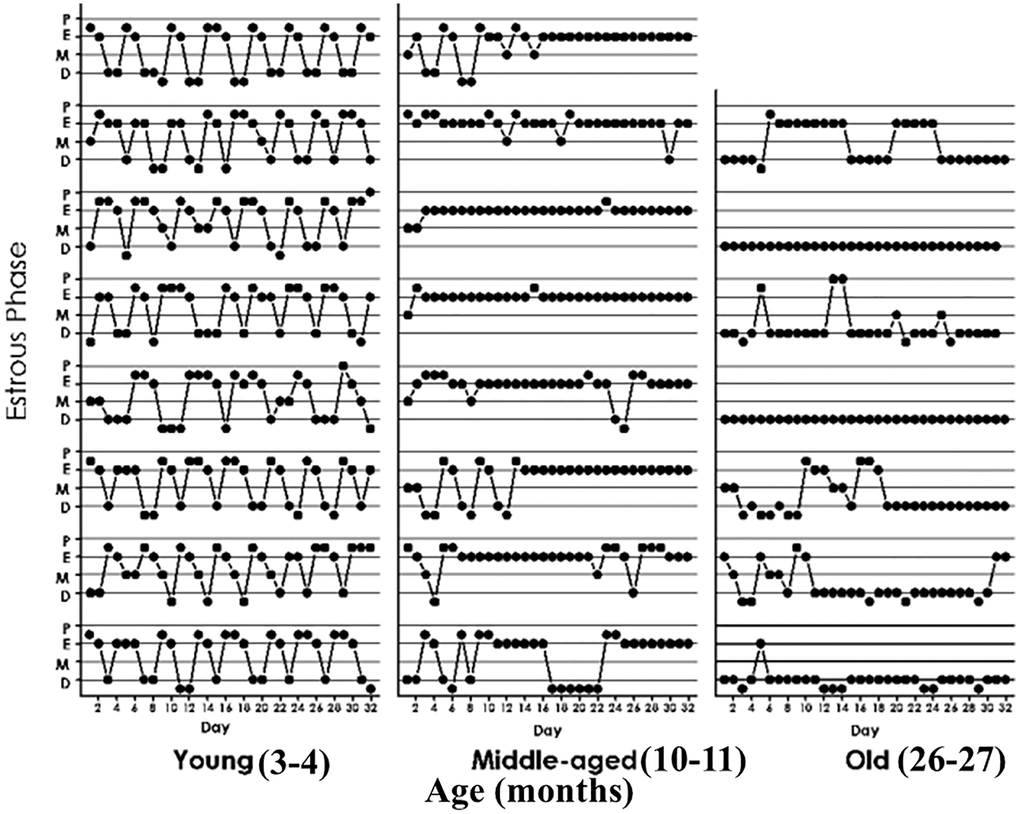
Figure 3. Estrous cycle patterns in young (3 momths, A), middle-aged (10 months, B) and old (26 months, C) female rats. The Y axis represents the estrous cycle phases of the animal on the indicated day of a 31-day window (X-axis) as determined by vaginal cytology. Four estrous phases were defined according to the proportion of epithelial and cornified cells or leukocytes; Proestrus (P) was characterized by abundance of epithelial cells and the presence of leukocytes. Estrus (E), showed abundance of cornified cells and to a lesser extent epithelial cells. Diestrus 1 (D1), was defined by the presence of an increasing number of leukocytes accompanied by substantial numbers of cornified cells. Diestrus 2 (D2) was characterized by high abundance of leukocyes which constituted the vast majority of cells present in the vaginal smears at D2.
Effect of hypothalamic OSKM gene therapy on fertility in MA female rats
As expected, the young group showed a much higher pregnancy rate (83%) than the MA controls (8.3%). Interestingly, MA OSKM treated rats showed a 25% rate of fertility, which although far lower than in young rats, was 3 times higher than in MA controls. (Figure 4B). When a z-test for proportions was applied to 8.3 vs. 25%, the two-sided p-value was 0.13104, while the one-sided p-value was 0.06552. Since the null hypothesis is one-sided, the one-sided value was used in our analysis.
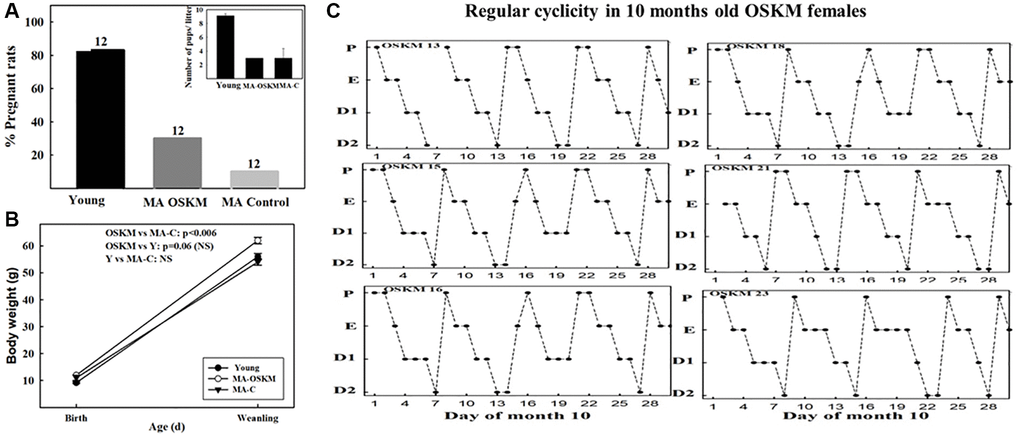
Figure 4. Effect of long-term OSKM-GFP gene therapy on the pregnancy rate, litter size, pup BW and estrous cyclicity in MA female rats. (A) There were three experimental groups; 4 months old intact females (Young, N = 12), 9.3 months old control rats (MA control N = 12) and 9.3 months old OSKM-treated rats (MA OSKM N = 12). These N values are indicated on each bar. (B) Pups were weighed when they were 1 or 2 days old and again at weaning (21 days). (C) During the 10th month of life daily vaginal smears were taken in the OSKM-treated rats in order to assess the effect of the OSKM gene therapy on regularity of estrous cyclicity. The estrous cycle phases are indicated as follows. Abbreviations: P: Proestrus; E: Estrus; D1: Early diestrus; D2: Late diestrus.
Also as expected, the litter size was higher in the young (mean litter size 9.1 pups) than in the MA controls (Figure 4B, inset) and OSKM-treated (mean litter size, 3 pups in each group). The body weight of the pups at birth was lower in the young rats (mean BW in pups from young rats, 9.2 g) than in the MA controls and treated (mean BW, 11.2 and 11.8 g, respectively). (Figure 4A). However, at weaning the pups of the OSKM animals had gained more weight than the pups of the MA-C females (Figure 4A). The BW of the pups from the young females remained lower than the OSKM counterparts, a fact that may be accounted for by the higher number of pups the young mothers had to feed as compared with the OSKM MA rats (8 vs. 3, respectively). In the case of the pups from the MA-control females, the lower BW at weaning suggests that milk production and/or quality declines with age, a change that was attenuated by hypothalamic OSKM gene therapy.
The total number of pups born from Y, MA-C and MA-OSKM was 83, 3 and 9, respectively. All pups survived and showed similarly normal behavior in the three groups.
Interestingly, most OSKM-treated MA rats were still cycling regularly at age 10 months (Figure 4C), which is in contrast with what happened in 10-month-old MA intact controls which showed irregular cycles with the prevalence of CE status (Figure 3, middle panel).
Discussion
Although the sequence of changes that take place during reproductive aging in female rats is qualitatively similar in most strains, the timing is likely to differ among strains and in different laboratory environments. This is why we considered it necessary to characterize the chronology of reproductive changes in our female rat colony before attempting to implement long-term protective gene therapy in young animals. In qualitative terms, the age changes in vaginal cytology observed in our Sprague-Dawley females are in agreement with early reports in Long-Evans rats. Thus, in Long-Evans females the vaginal smears show regular 4–5 day cycles from 2 to 10 months of age, transitioning to irregular cycles and persistent vaginal cornification (CE) during the following two months [20, 21]. In our MA females this transition takes place earlier. After 18–20 months, most of the animals progressively become pseudopregnant (PSP), a condition characterized by persistent diestrus vaginal cytology interrupted at irregular intervals, by 2–3 days of nucleated and cornified vaginal smears. In Long-Evans females the PSP stage has been reported to occur between 27–30 months of age [21]. While the CE stage is associated with an anovulatory condition, PSP females show intermittent ovulatory activity [22].
The MA females were mated 5.8 months (174 days) post adenovector injection, which, despite the long-term expression of HD-RAds, makes it likely that OSKM gene expression in the MBH at the time of mating was significantly weaker than at the time of injection. We have previously demonstrated that in our female rat colony, ovulation remains regular until 9 months of age, becoming irregular at age 10 months (6). Clearly, regular ovulation is a necessary but not sufficient condition for keeping the rats at optimal fertility levels, progressive decline in other components of the reproductive system has a marked overall impact on fertility, as shown in the MA controls. Considering that at the time of mating, MA rats were nearing reproductive cessation, the 25% fertility rate observed in the OSKM-treated rats reveals a significant beneficial effect of the OSKM treatment on the functional stability of the reproductive system. While litter size and mean pup BW at birth were not affected by the treatment, OSKM gene therapy seems to have exerted beneficial effects milk production or quality, as suggested the higher BW gain of the OSKM pups at weaning (Figure 4A). It is well-established that the OSKM genes have the ability to slow down the ticking rate of the epigenetic clock [23].
It should be pointed out that only a small proportion of hypothalamic cells are expected to have been transduced by our OSKM-GFP vector. However, since an extensive network of tanycytic processes perfuses the hypothalamic parenchyma (Figure 2B, 2C) it is likely that the influence of the relatively small number of transduced cells was amplified by this network. Those few cells expressing the OSKM genes may have generated a neuroprotective environment in the hypothalamus, effectively slowing down the effect of age on reproductive function. The observation that, unlike intact females, at 10 months of age the OSKM treated-animals remained cycling regularly, strongly supports the evidence that the OSKM genes effectively slowed down the age-related decline of the reproductive system. Neuroprotective gene therapy for IGF-1 was reported to keep MA females cycling regularly until 11 months of age (6), thus, our observation that 10 months old OSKM-treated females keep cycling regularly (Figure 4C) is in line with the results reported for IGF-1 gene therapy in the hypothalamus. Nevertheless, the more physiologically significant finding of the present report is the one related with an increased fertility in OSKM MA females as it reveals a prolongevity effect of OSKM genes on the hypothalamus.
It was not possible to assess the impact of the OSKM treatment on hypothalamic DNA methylation as pregnant rats and nonpregnant animals would have shown pregnancy-related differences in their hypothalamic DNA methylation profile. In spite of the lack of hypothalamic DNA methylation data, it seems reasonable to expect that whatever the effects of aging on hypothalamic DNA methylation, the OSKM treatment reversed, at least in part, such epigenetic changes. In rats, aging is known to induce hypermethylation of hippocampal DNA which is correlated with learning impairment [24]. Interestingly, OSKM gene therapy in the hippocampus of old rats significantly reverses this age-induced hippocampal DNA hypermethylation and significantly restores learning performance in old rats [14].
Our results are in line with the evidence that viral vector-mediated delivery of the Yamanaka genes in the brain has strong regenerative effects without adverse side effects [11]. The particular significance or the present results is that, for the first time, it is shown that long-term OSKM gene therapy in the hypothalamus is able to extend fertility, and regular cyclicity, which depend not only on the functionality of the hypothalamo-pituitary-ovarian axis but also on other reproduction related components.
Conclusions
In women, cessation of ovulation is caused by ovarian exhaustion, not by hypothalamic dysfunction, although it is likely that a functional decline in the human reproductive hypothalamus also occurs with age. In this context, our results in female rats demonstrating that hypothalamic gene therapy with prolongevity genes like OSKM prolongs ovulation and fertility suggest that minimally invasive gene transfer of OSKM/OSK into the hypothalamus of premenopausal women could prolong reproductive function, allowing their fertility to extend into older ages. The benefits of eventual prolongation of women fertility are difficult to overstate. Indeed, the potential of emerging technologies to prolong female fertility in modern society allows us to imagine a future where women can postpone conception long enough to consolidate their professional careers so that when they decide to form a family, their offspring can count on the full presence of the mother during the first years of childhood.
Author Contributions
Deflina Gallardo: Experimental design, animal handling, stereotaxic surgery, vaginal smears, cytological analysis and miscellaneous procedures. Conceptualization and reviewing. Mauricio Girard: Animal experiments including stereotaxic surgery. Enrique L. Portiansky: Hypothalamic image capturing, analysis and configuration. Rodolfo G. Goya: Supervision, conceptualization, writing - reviewing and editing.
Acknowledgments
The authors thank Ms. Yolanda E. Sosa for technical and editorial assistance.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Ethical Statement
All experiments with animals were performed according to the Animal Welfare Guidelines of NIH (INIBIOLP’s Animal Welfare Assurance No A5647-01).
Funding
Our studies are supported in part by research grant number SEGR-9-23-21 from the Society for Experimental Gerontological Research (SEGR), grant BRLS# 2024-02 from the Biomedical Research and Longevity Society, grant PICT 2018-00907 to RGG and Grant V309 to ELP.
References
- 1. Zheng W, Jimenez-Linan M, Rubin BS, Halvorson LM. Anterior pituitary gene expression with reproductive aging in the female rat. Biol Reprod. 2007; 76:1091–102. https://doi.org/10.1095/biolreprod.106.057877 [PubMed]
- 2. Neal-Perry GS, Zeevalk GD, Santoro NF, Etgen AM. Attenuation of preoptic area glutamate release correlates with reduced luteinizing hormone secretion in middle-aged female rats. Endocrinology. 2005; 146:4331–9. https://doi.org/10.1210/en.2005-0575 [PubMed]
- 3. Huang HH, Marshall S, Meites J. Capacity of old versus young female rats to secrete LH, FSH and prolactin. Biol Reprod. 1976; 14:538–43. https://doi.org/10.1095/biolreprod14.5.538 [PubMed]
- 4. Neal-Perry G, Nejat E, Dicken C. The neuroendocrine physiology of female reproductive aging: An update. Maturitas. 2010; 67:34–8. https://doi.org/10.1016/j.maturitas.2010.04.016 [PubMed]
- 5. Rubin BS, Lee CE, King JC. A reduced proportion of luteinizing hormone (LH)-releasing hormone neurons express Fos protein during the preovulatory or steroid-induced LH surge in middle-aged rats. Biol Reprod. 1994; 51:1264–72. https://doi.org/10.1095/biolreprod51.6.1264 [PubMed]
- 6. Rodríguez SS, Schwerdt JI, Barbeito CG, Flamini MA, Han Y, Bohn MC, Goya RG. Hypothalamic IGF-I gene therapy prolongs estrous cyclicity and protects ovarian structure in middle-aged female rats. Endocrinology. 2013; 154:2166–73. https://doi.org/10.1210/en.2013-1069 [PubMed]
- 7. Gurdon JB. The developmental capacity of nuclei taken from intestinal epithelium cells of feeding tadpoles. J Embryol Exp Morphol. 1962; 10:622–40. [PubMed]
- 8. Wilmut I, Schnieke AE, McWhir J, Kind AJ, Campbell KH. Viable offspring derived from fetal and adult mammalian cells. Nature. 1997; 385:810–3. https://doi.org/10.1038/385810a0 [PubMed]
- 9. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006; 126:663–76. https://doi.org/10.1016/j.cell.2006.07.024 [PubMed]
- 10. Goya RG, Lehmann M, Chiavellini P, Canatelli-Mallat M, Hereñú CB, Brown OA. Rejuvenation by cell reprogramming: a new horizon in gerontology. Stem Cell Res Ther. 2018; 9:349. https://doi.org/10.1186/s13287-018-1075-y [PubMed]
- 11. Lu Y, Brommer B, Tian X, Krishnan A, Meer M, Wang C, Vera DL, Zeng Q, Yu D, Bonkowski MS, Yang JH, Zhou S, Hoffmann EM, et al. Reprogramming to recover youthful epigenetic information and restore vision. Nature. 2020; 588:124–9. https://doi.org/10.1038/s41586-020-2975-4 [PubMed]
- 12. Ocampo A, Reddy P, Martinez-Redondo P, Platero-Luengo A, Hatanaka F, Hishida T, Li M, Lam D, Kurita M, Beyret E, Araoka T, Vazquez-Ferrer E, Donoso D, et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell. 2016; 167:1719–33.e12. https://doi.org/10.1016/j.cell.2016.11.052 [PubMed]
- 13. Macip CC, Hasan R, Hoznek V, Kim J, Lu YR, Metzger LE
4th , Sethna S, Davidsohn N. Gene Therapy-Mediated Partial Reprogramming Extends Lifespan and Reverses Age-Related Changes in Aged Mice. Cell Reprogram. 2024; 26:24–32. https://doi.org/10.1089/cell.2023.0072 [PubMed] - 14. Horvath S, Lacunza E, Mallat MC, Portiansky EL, Gallardo MD, Brooke RT, Chiavellini P, Pasquini DC, Girard M, Lehmann M, Yan Q, Lu AT, Haghani A, et al. Cognitive rejuvenation in old rats by hippocampal OSKM gene therapy. Geroscience. 2024. [Epub ahead of print]. https://doi.org/10.1007/s11357-024-01269-y [PubMed]
- 15. Lehmann M, Canatelli-Mallat M, Chiavellini P, Morel GR, Reggiani PC, Hereñú CB, Goya RG. Regulatable adenovector harboring the GFP and Yamanaka genes for implementing regenerative medicine in the brain. Gene Ther. 2019; 26:432–40. https://doi.org/10.1038/s41434-019-0063-x [PubMed]
- 16. Sommer CA, Stadtfeld M, Murphy GJ, Hochedlinger K, Kotton DN, Mostoslavsky G. Induced pluripotent stem cell generation using a single lentiviral stem cell cassette. Stem Cells. 2009; 27:543–9. https://doi.org/10.1634/stemcells.2008-1075 [PubMed]
- 17. Paxinos G, Watson C. The rat brain in stereotaxic coordinates. San Diego: Academic Press. 1998.
- 18. Marcondes FK, Bianchi FJ, Tanno AP. Determination of the estrous cycle phases of rats: some helpful considerations. Braz J Biol. 2002; 62:609–14. https://doi.org/10.1590/s1519-69842002000400008 [PubMed]
- 19. Astwood EB. Changes in the weight and water content of the uterus of the normal adult rat. Am J Physiol. 1939; 126:162–70. https://doi.org/10.1152/ajplegacy.1939.126.1.162
- 20. Huang HH, Meites J. Reproductive capacity of aging female rats. Neuroendocrinology. 1975; 17:289–95. https://doi.org/10.1159/000122367 [PubMed]
- 21. Lu KH, Hopper BR, Vargo TM, Yen SS. Chronological changes in sex steroid, gonadotropin and prolactin secretions in aging female rats displaying different reproductive states. Biol Reprod. 1979; 21:193–203. https://doi.org/10.1095/biolreprod21.1.193 [PubMed]
- 22. Aschheim P. [Repeated pseudogestation in senile rats]. C R Hebd Seances Acad Sci. 1961; 253:1988–90. [PubMed]
- 23. Horvath S, Raj K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat Rev Genet. 2018; 19:371–84. https://doi.org/10.1038/s41576-018-0004-3 [PubMed]
- 24. Chiavellini P, Lehmann M, Canatelli Mallat M, Zoller JA, Herenu CB, Morel GR, Horvath S, Goya RG. Hippocampal DNA Methylation, Epigenetic Age, and Spatial Memory Performance in Young and Old Rats. J Gerontol A Biol Sci Med Sci. 2022; 77:2387–94. https://doi.org/10.1093/gerona/glac153 [PubMed]