



Introduction
Malignant tumors of the hematopoietic system occur in the blood-forming tissues. In 2020, more than 1.2 million people worldwide were diagnosed with malignant tumors of the hematopoietic system, and over 700,000 people died from these malignancies [1]. Hematological malignancies are broadly categorized into lymphoma, which originates from the lymphatic system; multiple myeloma (MM), which affects plasma cells in the bone marrow; and leukemia, which impacts cells in the bone marrow or blood [2]. Currently, many observational studies have investigated the etiology of hematological malignancies, including factors such as air pollution, chlamydia contamination, and dietary habits, among others [3, 4]. However, due to the multitude of confounding factors in retrospective studies, many causative factors still cannot be conclusively determined.
Telomeres are natural ends of chromosomes characterized by variable numbers of TTAGGG repeat sequences and associated proteins [5]. The role of telomeres in human health and disease is still not fully understood. But more and more studies have demonstrated that telomeres play a crucial role in the development and progression of cancer [6]. Several studies have found that longer telomeres may be associated with an increased risk of various cancers, including melanoma, acute myeloid leukemia, and chronic lymphocytic leukemia in cancer-prone families [7–9]. Bao et al. found that longer telomeres in leukocytes were identified as important risk factors for the development of myeloproliferative neoplasms [10]. However, several studies found that starting life with shorter telomeres may increases the risk of cancer [11]. The possible reason for generating these opposing views is the insufficient research methods.
Mendelian randomization (MR) uses genetic variation as an instrumental variable (IV), which has advantages compared with other research methods [12]. This approach utilizes genetic variants associated with the exposure of interest to estimate the causal effect on the outcome, thereby providing valuable insights into disease etiology [13–15]. In recent years, there has been growing interest in exploring the potential association between telomere length and other type of carcinomas using Mendelian randomization analysis [16]. However, the precise causal relationship between telomere length and the hematopoietic malignancies remains unclear.
This Mendelian randomization study aims to investigate the causal relationship between telomere length and hematopoietic malignancies. By utilizing large-scale genome-wide association studies (GWAS) data and applying rigorous statistical methods, we seek to provide robust evidence regarding the role of telomeres in hematopoietic malignancies and may have implications for risk prediction, prevention, and potentially targeted therapies.
Materials and Methods
Study design
We performed a two-sample Mendelian randomization (MR) study to investigate the causal association between telomere length and hematological malignancies. As shown in Figure 1, in order for genetic variation to serve as a valid instrumental variable, it must adhere to three fundamental principles: (1) Genetic variants exhibit a robust correlation with the exposure of interest. (2) Genetic variants are not associated with potential confounders. (3) Genetic variants do not exert a direct influence on the outcome of interest. [17]. Figure 2 provides an overview of the study design. We reported this study according to the Strengthening the Reporting of Observational Studies in Epidemiology using Mendelian Randomization (STROBE-MR) [18].
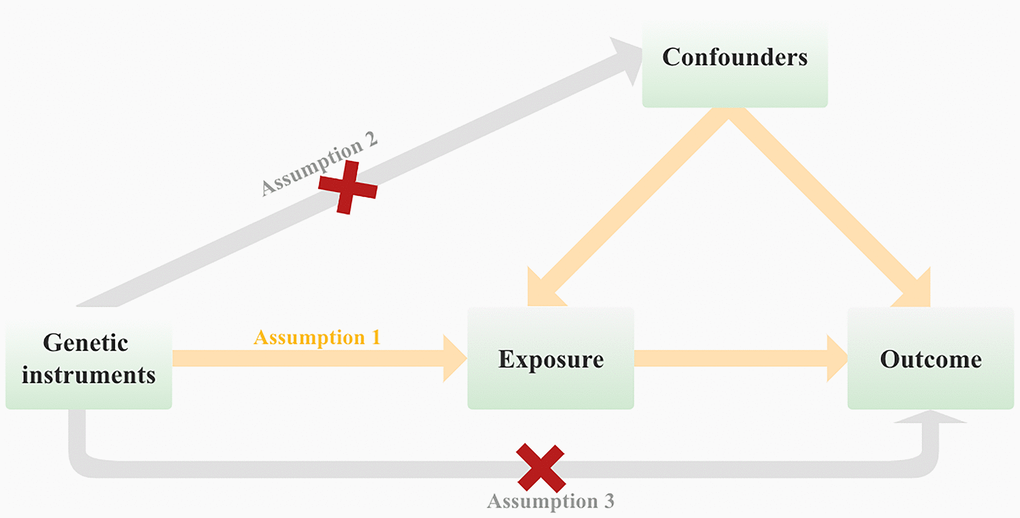
Figure 1. Mendelian randomization (MR) analysis is based on three fundamental assumptions at its core.
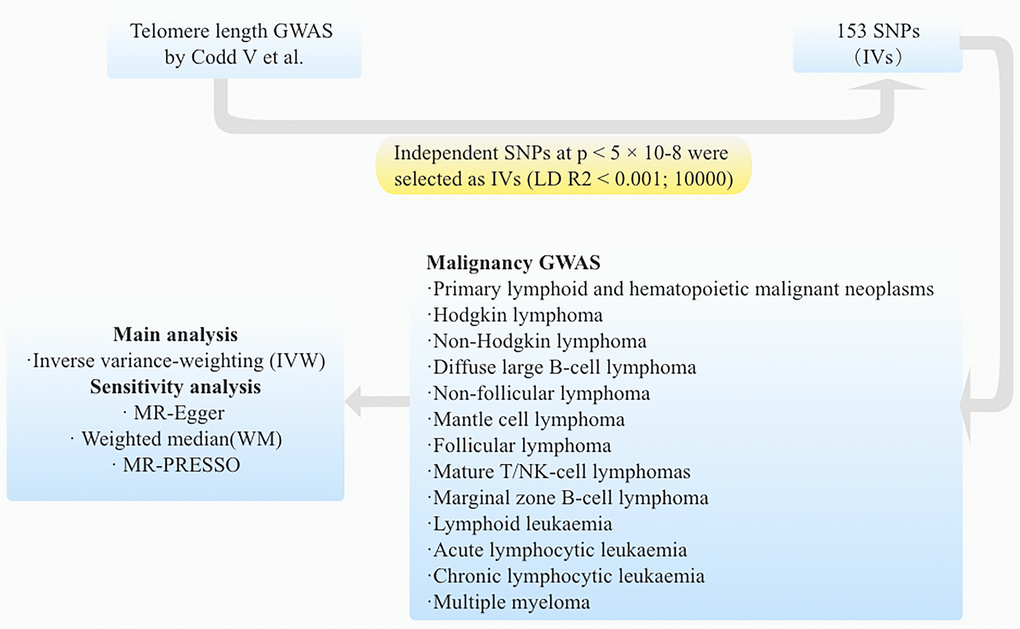
Figure 2. The flow diagram illustrates the sequential process of the MR study.
Data sources of exposure
Data on the association between genetics and telomere length were extracted from a GWAS conducted on a European cohort comprising 472,174 individuals (study ID “ieu-b-4879” and can be downloaded from the IEU GWAS database (https://gwas.mrcieu.ac.uk/datasets/) [19]. All participants fell within the age range of 40-69 years, with approximately equal representation of males (45.8%) and females (54.2%). The quantification of telomere length was performed using a well-established quantitative PCR method, and multiple quality control measures were implemented to account for potential influences of ethnicity, gender, age, and technical variables, as delineated in a previous investigation.
We included SNPs reaching GWAS (GWAS p < 5 × 10−8). Then, these SNPs were clumped based on the linkage disequilibrium (r 2 <0.001; kb = 10000) in the given genome region. Additionally, potential weak IVs (F-statistics <10) were excluded from the final analysis, as determined by calculating the F-statistics. Moreover, any palindromic SNPs with ambiguous minor allele frequencies (A/T or C/G) were discarded. Subsequently, we removed SNPs directly associated with hematological malignancies and confounding factors such as BMI [20] and tobacco smoking [21] through PhenoScanner datasets (http://www.phenoscanner.medschl.cam.ac.uk/).
Data sources of outcome
To investigate genetic associations with lymphoid and hematopoietic malignant neoplasms, we employed summary-level data obtained from the most recent publicly accessible R9 data release by Kurki et al. [22]. The FinnGen study is a comprehensive nationwide genetic investigation conducted in conjunction with electronic health records, aiming to collect genetic data. This study was adjusted for potential confounding factors including sex, age, genetic components, and genotyping batch.
Genetic associations with 13 lymphoid and hematopoietic malignant neoplasms GWAS databases were available from the FinnGen website (https://www.finngen.fi/en). Included outcomes were classified into five major categories according to the pathological pattern: (1) Primary lymphoid and hematopoietic malignant neoplasms (7519 cases and 299,952 controls); (2) HL (2602 cases and 299,952 controls); (3) NHL (1088 cases and 299,952 controls): FL (1081 cases and 299,952 controls), Non-follicular lymphoma (NFL) (2602 cases and 299,952 controls), DLBCL (1010 cases and 287,137 controls), Mature T/NK-cell lymphomas (335 cases and 299,952 controls), Mantle cell lymphoma (MCL) (119 cases and 287,173 controls), Marginal zone B-cell lymphoma (MZBL) (192 cases and 287,137 controls); and (4) Lymphoid leukaemia (1493 cases and 299,952 controls) (Acute lymphocytic leukaemia (ALL) (184 cases and 287,136 controls), Chronic lymphocytic leukaemia (CLL) (624 cases and 287,133 controls)); (5) Multiple myeloma (MM) (674 cases and 376,603 controls).
Primary MR analysis
The Wald ratio was used to assess the effect of telomere length on lymphoid and hematopoietic malignant neoplasms for each SNP. All SNP effects were meta-analyzed by the inverse-variance weighted (IVW) method [14]. This study used the multiplicative random-effects IVW method as the main MR analysis. In order to examine the potential causal relationship between telomere length and lymphoid and hematopoietic malignant neoplasms, we conducted MR analyses employing four distinct methods: IVW, MR-Egger, MR-PRESSO, and weighted median. The IVW method assumes the absence of pleiotropy, wherein instrumental variables (IVs) solely affect telomere length and not through alternative pathways. The MR-Egger approach provides a valid estimate of causal effect [23]. For the weighted median approach to be applicable, it necessitates that at least half of the IVs are valid [24]. The MR-PRESSO method effectively identifies potential IV abnormalities and automatically eliminates them to ensure an unbiased causal effect estimation. To assess heterogeneity, Cochran’s Q test was performed. In cases where no heterogeneity was observed in the IVW analysis, the fixed-effect model was utilized; otherwise, the random-effect model was employed.
Sensitivity analysis
In sensitivity analyses, MR-Egger [23] and weighted median (WM) methods [24] were applied to account for horizontal pleiotropic effects. The MR-Egger method was based on the Instrument Strength Independent of Direct Effect (InSIDE) assumption, which often provides imprecise and low statistical power MR results, especially when meeting small sizes of SNPs (e.g., <10) [23]. In our MR study, MR-Egger was mainly used to detect pleiotropy; the value of the intercept term is far from zero, indicating horizontal pleiotropy (P < 0.05) [25]. The WM method was more reliable if more than 50% of SNPs were invalid instruments (e.g., due to pleiotropy) [26]. In addition, MR-PRESSO analysis was used to detect outliers, which can reduce heterogeneity by removing those outliers that may lead to heterogeneity (Figure 3) [27]. We performed leave-one-out method analysis to determine potentially influential SNPs by removing each SNP. We adjusted the multiple testing by false discovery rate (FDR).
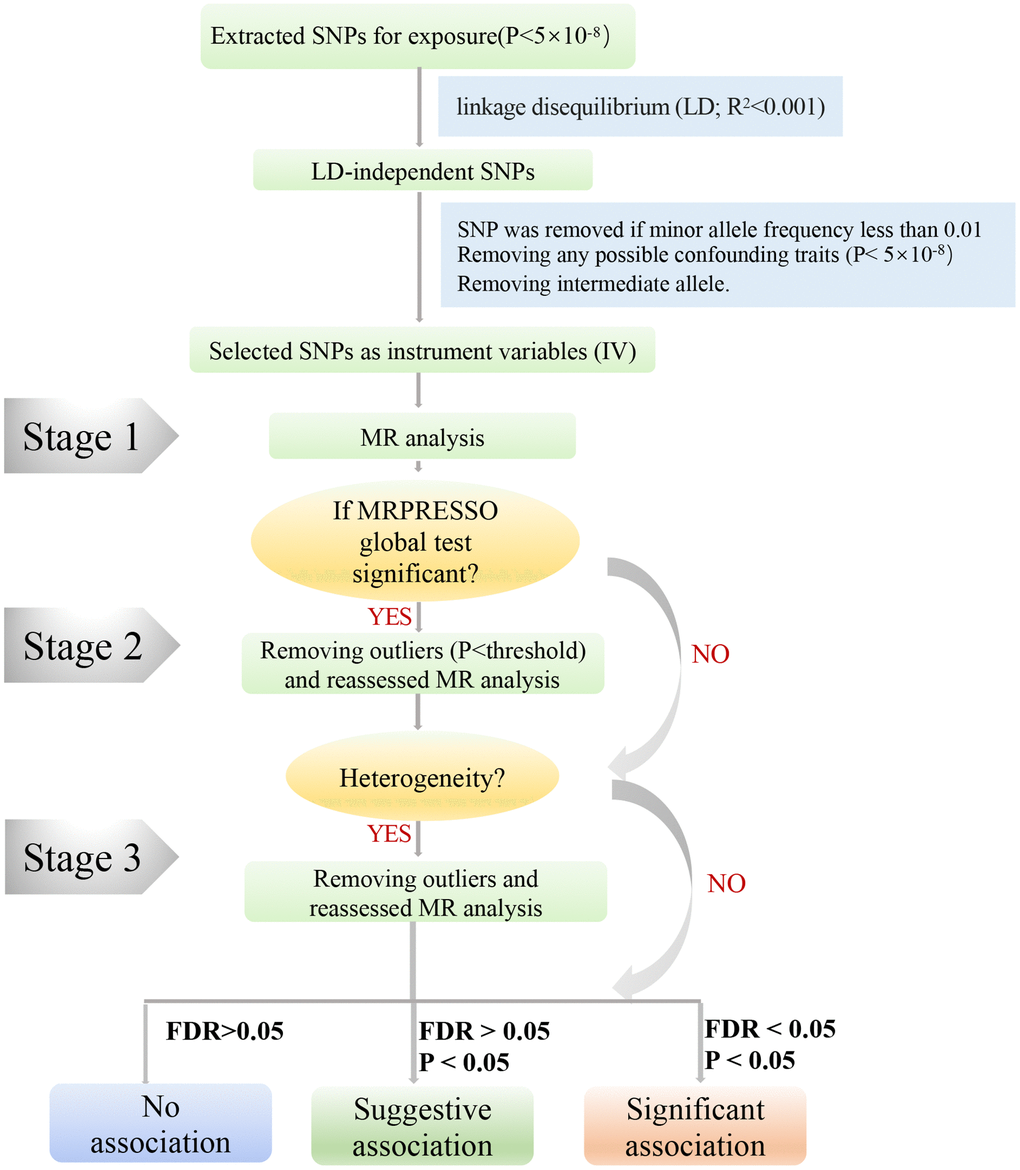
Figure 3. A step-by-step flow chart demonstrates the analytical methods employed and outlines the sequential execution of MR analysis. Step 1 involved conducting the MR analysis using the selected SNPs, followed by performing the MRPRESSO outlier test. If significance was detected (P < 0.05), we proceeded to step 2. In step 2, the MR analysis was reevaluated after removing all identified outliers (P < 0.05). Should heterogeneity persist, step 3 entailed excluding SNPs with a MR-PRESSO test P-value below 1 and reevaluating the MR analysis.
MR procedures
To ensure unbiased results by addressing potential heterogeneity, we followed a three-step approach in our study (Figure 3). In Step 1, we initially conducted MR analysis using the selected SNPs mentioned above, subsequently employing the MRPRESSO outlier test. If any outliers were detected (P < 0.05), we proceeded to Step 2. In Step 2, we reevaluated the MR analysis after excluding all outliers (P < 0.05). If heterogeneity persisted, we entered Step 3, wherein SNPs with a P-value less than 1 in the MR-PRESSO test were excluded, and the MR analysis was reevaluated. Furthermore, we exercised caution in interpreting the results if any potentially influential SNPs were identified through the leave-one-out test.
For our MR study, we utilized several R packages including “TwoSampleMR” [28], “MendelianRandomization” [24], and “MRPRESSO” [27] packages. The forestploter R packages were employed for data visualization. All statistical analyses were conducted using R software version 4.3.1 (R Foundation, Vienna, Austria, https://www.R-project.org/).
Results
We identified 34857 SNPs that showed a significant association with telomere length in the discovery cohort consisting of 472,174 European participants, as reported by Codd V et al. [19]. These SNPs reached the genome-wide significance level (p-value < 5 × 10−8). To ensure the independence of instrumental variables for telomere length (TL), SNPs in linkage disequilibrium (with r2 >0.001 and clump distance <10,000 kb) were excluded. Notably, rs7705526 was excluded due to its significant association with some hematological malignancies directly by screening PhenoScanner datasets (P < 5 × 10−8). Ultimately, 153 independent SNPs remained as instrumental variables. Supplementary Table 1 provides detailed information on the selected SNPs. The F statistics of these SNPs ranged from 29 to 1628, suggesting no weak instrumental variables exited [29]. The instrumental variables accounted for 3.36% of the variance in explaining the exposure.
MR main analysis
Genetically predicted longer telomere length could increase the risk of all types of primary lymphoid, Hodgkin lymphoma (HL), non-Hodgkin lymphoma (NHL), diffuse large B-cell lymphoma (DLBCL), non-follicular lymphoma (NFL), mantle cell lymphoma (MCL), acute lymphoid leukemia (ALL), chronic lymphoid leukemia (CLL) after FDR control (FDR <0.05; Figure 4 and Table 1). Specifically, a 1-SD increase of telomere length could increase the risk of all types of primary lymphoid (OR = 1.52, P = 2.11×10−6) by 52%, HL (OR = 1.64, P = 0.014) by 64%, NHL (OR = 1.70, P = 0.002) by 70%, DLBCL (OR = 1.57, P = 0.015) by 57%, NFL (OR = 1.58, P = 1.73 × 10−4) by 58%, MCL (OR = 3.13, P = 0.003) by 213%, ALL (OR = 2.65, P = 0.021) by 165%, CLL (OR = 2.80, P = 8.21 × 10−6) by 180%, and Multiple myeloma 1.85 (OR = 1.85, P = 0.016) by 85%. However, telomere length was not associated with increase in odds of follicular lymphomas (FL), mature T/NK cell lymphoma, and marginal zone B-cell lymphoma (MZBL).
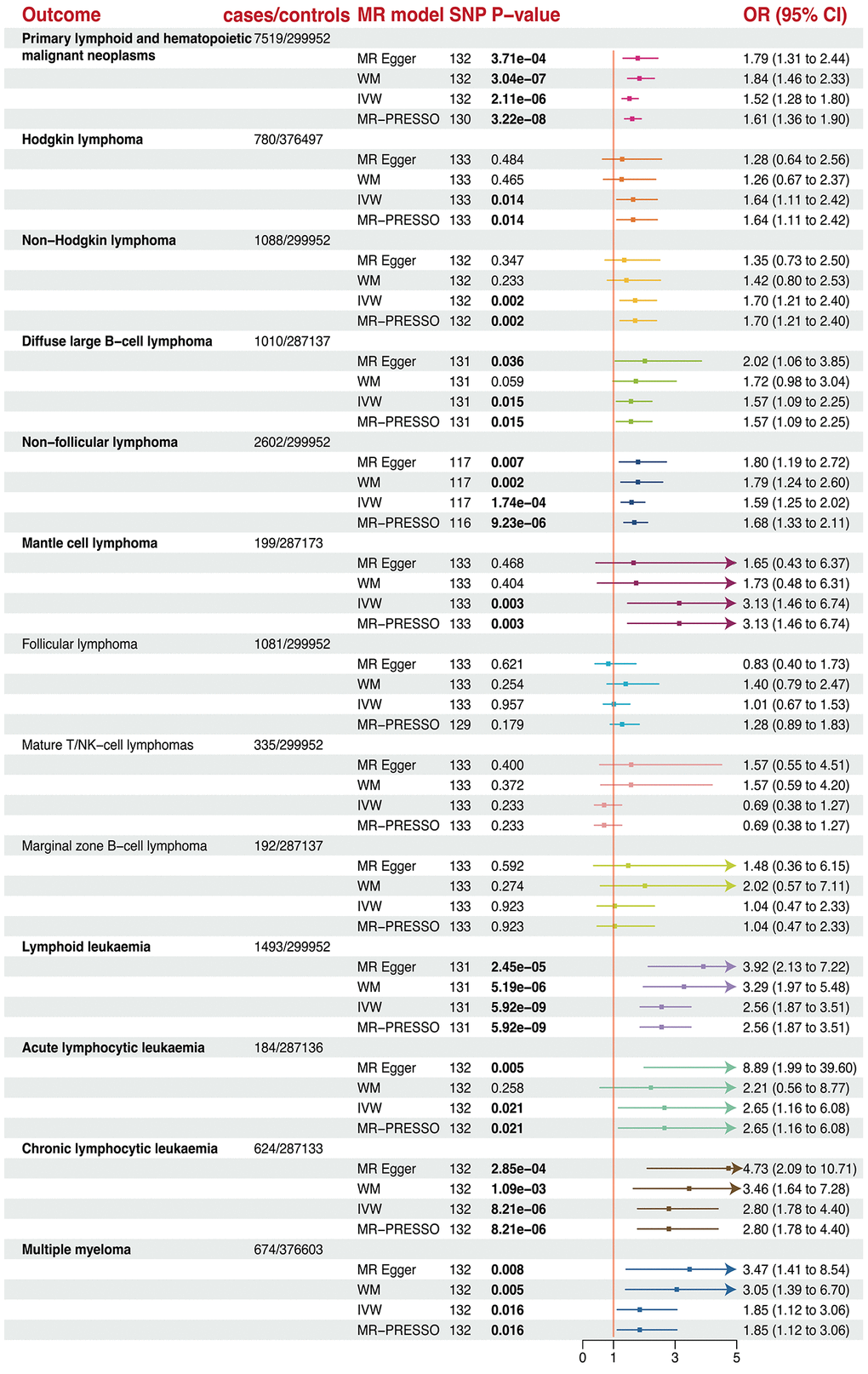
Figure 4. The forest plots revealed the causal association of telomere length with different lymphoma and hematopoietic malignancies.
Table 1. False discovery rate adjusted p-values for the tested associations of telomere length and outcomes.
| Outcome | SNPs | Original P-value | Benjamini-Hochberg adjusted P-value | Significant using an FDR of 0.05? |
| Primary lymphoid and hematopoietic malignant neoplasms | 132 | 2.11E-06 | 1.37E-05 | Yes |
| Hodgkin lymphoma | 117 | 0.014 | 0.026 | Yes |
| Non-Hodgkin lymphoma | 132 | 0.002 | 0.006 | Yes |
| Diffuse large B-cell lymphoma | 133 | 0.015 | 0.019 | Yes |
| Non-follicular lymphoma | 131 | 0.000 | 0.001 | Yes |
| Mantle cell lymphoma | 133 | 0.003 | 0.007 | Yes |
| Follicular lymphoma | 133 | 0.957 | 0.957 | No |
| Mature T/NK-cell lymphomas | 133 | 0.233 | 0.276 | No |
| Marginal zone B-cell lymphoma | 133 | 0.923 | 1.000 | No |
| Lymphoid leukaemia | 131 | 5.92E-09 | 7.69E-08 | Yes |
| Acute lymphocytic leukaemia | 132 | 2.09E-02 | 3.02E-02 | Yes |
| Chronic lymphocytic leukaemia | 132 | 0.000 | 0.000 | Yes |
| Multiple myeloma | 132 | 0.016 | 0.026 | Yes |
Sensitivity analysis
All WM and MR-Egger sensitivity analyses were directionally consistent in the IVW results except for FL and mature T/NK-cell lymphomas. The MR-PRESSO results suggested that the causal relationship still holds after removing outliers. No heterogeneity was detected except for primary lymphoid and FL. Furthermore, no horizontal pleiotropy was observed in this MR analysis across all subsets (Supplementary Table 2). Moreover, Leave-one-out analysis revealed that no SNP drove the results (Supplementary Figure 1).
To further investigate the causal relationship, we examined the effect of these malignancies as a risk factor for telomere length, thus ruling out the possibility of a bidirectional causal effect between telomere length and all types of lymphoma and lymphoid leukemia. Different SNPs associated with various lymphomas were considered, but no significant effect was found under any of the MR models. These results demonstrate the credibility of our conclusions.
Discussion
The objective of this study is to investigate the association between telomere length and various hematopoietic malignancies by MR analysis. Through our research, we found that telomere length may increase the risk of primary lymphoid, Hodgkin lymphoma, non-Hodgkin lymphoma, DLBCL, NFL, MCL, ALL, CLL and multiple myeloma. These findings were supported by rigorous statistical analysis, with a corrected FDR of less than 0.05. Furthermore, this study demonstrated no homogeneity and horizontal pleiotropy across all investigations.
Telomere length plays a critical role in maintaining genomic stability and preventing cellular senescence or apoptosis [30, 31]. Shortened telomeres are commonly observed in various cancer types and are generally associated with genomic instability and tumor progression [32–34]. Our findings add to the existing body of evidence by highlighting the importance of telomere length in hematopoietic malignancies. Previous studies have concluded that blood malignancies (NHL, MCL, ALL, CLL) have shorter telomere lengths than the control group [35–39]. Roos et al. found that shortened telomere length in chronic lymphocytic leukemia (CLL) patients is consistent with other classic biological factors of CLL, including unmutated immunoglobulin heavy chain variable region genes (UM-IGVH), positive CD38 and ZAP-70 (>30%), and short lymphocyte doubling time (<6 months) [40]. In a study conducted by Sellmann et al., a correlation was observed between the frequency of IGHV gene mutations and the length of telomeres [41]. The study conducted on CLL patients indicated that those with reduced telomere length demonstrated poorer clinical outcomes, including decreased progression-free survival (PFS) and overall survival (OS) [42]. However, their findings do not elucidate a causal relationship between telomere length and hematologic malignancies. Furthermore, it is worth noting that Furtado et al. [43] suggest that telomere shortening is an early event in the development of leukemia, as short telomeres are already present in small abnormal B-cell clones in monoclonal B-cell lymphocytosis. This disease precedes chronic lymphocytic leukemia, consistent with the causal relationship we deduced through Mendelian randomization. Several underlying mechanisms could explain the observed association between telomere length and hematopoietic cancers. One possibility is that telomere dysfunction is directly involved in the initiation and progression of these malignancies. Shortened telomeres may lead to chromosomal abnormalities, DNA damage, and genomic instability, ultimately contributing to the development of cancer cells. Additionally, alterations in telomerase activity or mutations in genes involved in telomere regulation could contribute to telomere length abnormalities. Telomerase, the enzyme responsible for adding telomeric repeats, is often upregulated in cancer cells, allowing them to maintain or even lengthen their telomeres [44]. Dysregulation of telomerase activity could lead to differences in telomere length among individuals affecting their susceptibility to hematopoietic malignancies [30, 31].
Currently, research on telomere therapy is still in the early stages, and there is no definitive treatment plan. However, some studies have begun to explore potential treatment methods. One possible treatment approach is to extend telomere length through stem cell transplantation. Researchers have found that during the process of differentiation, stem cells can restore telomere length, which may be helpful in treating certain telomere shortening-related diseases [45]. In addition, certain drugs and compounds are being studied for their potential in intervening with telomere length. For example, some anti-aging compounds are speculated to have potential telomere protection effects, such as telomerase activators, antioxidants, and certain vitamins [46–48]. However, it should be noted that the relationship between telomere length and various diseases is complex. Treating telomere length involves considering the cellular environment, genetic factors, and other relevant factors [16]. Telomere therapy is still in the research stage and requires further clinical trials and studies to validate its safety, efficacy, and applicability.
Importantly, our findings have clinical implications for the diagnosis, prognosis, and treatment of hematopoietic malignancies. Telomere length could serve as a potential biomarker for disease risk assessment, allowing for early detection and intervention. Moreover, telomere length could be used to predict treatment response and patient outcomes, enabling personalized therapeutic strategies [49].
While this study provides valuable insights, there are certain limitations that should be acknowledged. Genetic and environmental factors that influence telomere length were not extensively investigated in this study. Further research is needed to elucidate these factors and their interactions.
However, there are some concomitant limitations in our study. First, due to the unavailability of individual-level data, we can only perform causal association MR analysis and cannot further examine the sensitivity and specificity of the outcomes. Additionally, the FinnGen database does not disclose detailed disease diagnostic information, which may introduce errors in our phenotypic analysis. However, the FinnGen database links genotypes with specific data using unique national identification numbers, and the disease classification is primarily based on ICD. Therefore, the possibility of misclassification influencing the outcomes is likely to be small. Second, further investigation into the direct impact of telomerase activity on hematological tumor development is necessary as telomere length is primarily influenced by telomerase. This research may provide new insights into the mechanisms through which telomeres contribute to cancer development. However, due to the lack of comprehensive telomerase-related GWAS (genome-wide association studies) at present, we are currently unable to analyze the relationship between telomerase and hematological tumors. In the future, relevant studies will be necessary. Third, a significant portion of the participants included in this investigation were of European origin; hence, it is not possible to extrapolate our results to encompass all racial groups.
Conclusion
In conclusion, our study found that telomere length is a risk factor for a wide ride of hematopoietic malignancies. Understanding the role of telomere length in the pathogenesis of these cancers could pave the way for innovative diagnostic and therapeutic approaches. Further investigation into the underlying mechanisms and the identification of specific biomarkers associated with telomere length may contribute to improved clinical management and patient outcomes in hematopoietic malignancies.
Author Contributions
TF designed the study. TF, ZZ, and KR conducted data analysis. TF conceived the project and wrote the manuscript. LZ revised and approved the paper. All authors contributed to the article and approved the submitted version.
Acknowledgments
We would like to thank American Journal Experts (https://www.aje.com/) for English language editing.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Ethical Statement
No ethical statement is required because the study was based on GWAS publicly available data.
Funding
This work was supported by the Incubation Project, West China Hospital, Sichuan University (2019HXFH053).
References
- 1. Andrades A, Peinado P, Alvarez-Perez JC, Sanjuan-Hidalgo J, García DJ, Arenas AM, Matia-González AM, Medina PP. SWI/SNF complexes in hematological malignancies: biological implications and therapeutic opportunities. Mol Cancer. 2023; 22:39. https://doi.org/10.1186/s12943-023-01736-8 [PubMed]
- 2. Smith A, Howell D, Crouch S, Painter D, Blase J, Wang HI, Hewison A, Bagguley T, Appleton S, Kinsey S, Burton C, Patmore R, Roman E. Cohort Profile: The Haematological Malignancy Research Network (HMRN): a UK population-based patient cohort. Int J Epidemiol. 2018; 47:700. https://doi.org/10.1093/ije/dyy044 [PubMed]
- 3. Drexler HG, Dirks WG, MacLeod RA, Uphoff CC. False and mycoplasma-contaminated leukemia-lymphoma cell lines: time for a reappraisal. Int J Cancer. 2017; 140:1209–14. https://doi.org/10.1002/ijc.30530 [PubMed]
- 4. Taj T, Chen J, Rodopoulou S, Strak M, de Hoogh K, Poulsen AH, Andersen ZJ, Bellander T, Brandt J, Zitt E, Fecht D, Forastiere F, Gulliver J, et al. Long-term exposure to ambient air pollution and risk of leukemia and lymphoma in a pooled European cohort. Environ Pollut. 2024; 343:123097. https://doi.org/10.1016/j.envpol.2023.123097 [PubMed]
- 5. de Lange T. Shelterin-Mediated Telomere Protection. Annu Rev Genet. 2018; 52:223–47. https://doi.org/10.1146/annurev-genet-032918-021921 [PubMed]
- 6. Lansdorp PM. Telomeres, aging, and cancer: the big picture. Blood. 2022; 139:813–21. https://doi.org/10.1182/blood.2021014299 [PubMed]
- 7. McNally EJ, Luncsford PJ, Armanios M. Long telomeres and cancer risk: the price of cellular immortality. J Clin Invest. 2019; 129:3474–81. https://doi.org/10.1172/JCI120851 [PubMed]
- 8. Haycock PC, Burgess S, Nounu A, Zheng J, Okoli GN, Bowden J, Wade KH, Timpson NJ, Evans DM, Willeit P, Aviv A, Gaunt TR, Hemani G, et al, and Telomeres Mendelian Randomization Collaboration. Association Between Telomere Length and Risk of Cancer and Non-Neoplastic Diseases: A Mendelian Randomization Study. JAMA Oncol. 2017; 3:636–51. https://doi.org/10.1001/jamaoncol.2016.5945 [PubMed]
- 9. Schmutz I, Mensenkamp AR, Takai KK, Haadsma M, Spruijt L, de Voer RM, Choo SS, Lorbeer FK, van Grinsven EJ, Hockemeyer D, Jongmans MC, de Lange T. TINF2 is a haploinsufficient tumor suppressor that limits telomere length. Elife. 2020; 9:e61235. https://doi.org/10.7554/eLife.61235 [PubMed]
- 10. Bao EL, Nandakumar SK, Liao X, Bick AG, Karjalainen J, Tabaka M, Gan OI, Havulinna AS, Kiiskinen TTJ, Lareau CA, de Lapuente Portilla AL, Li B, Emdin C, et al, and FinnGen, and 23andMe Research Team. Inherited myeloproliferative neoplasm risk affects haematopoietic stem cells. Nature. 2020; 586:769–75. https://doi.org/10.1038/s41586-020-2786-7 [PubMed]
- 11. Nassour J, Schmidt TT, Karlseder J. Telomeres and Cancer: Resolving the Paradox. Annu Rev Cancer Biol. 2021; 5:59–77. https://doi.org/10.1146/annurev-cancerbio-050420-023410 [PubMed]
- 12. Boyko EJ. Observational research--opportunities and limitations. J Diabetes Complications. 2013; 27:642–8. https://doi.org/10.1016/j.jdiacomp.2013.07.007 [PubMed]
- 13. Martín-Masot R, Herrador-López M, Navas-López VM, Carmona FD, Nestares T, Bossini-Castillo L. Celiac Disease Is a Risk Factor for Mature T and NK Cell Lymphoma: A Mendelian Randomization Study. Int J Mol Sci. 2023; 24:7216. https://doi.org/10.3390/ijms24087216 [PubMed]
- 14. Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013; 37:658–65. https://doi.org/10.1002/gepi.21758 [PubMed]
- 15. Smith GD, Ebrahim S. 'Mendelian randomization': can genetic epidemiology contribute to understanding environmental determinants of disease? Int J Epidemiol. 2003; 32:1–22. https://doi.org/10.1093/ije/dyg070 [PubMed]
- 16. Ye Q, Apsley AT, Etzel L, Hastings WJ, Kozlosky JT, Walker C, Wolf SE, Shalev I. Telomere length and chronological age across the human lifespan: A systematic review and meta-analysis of 414 study samples including 743,019 individuals. Ageing Res Rev. 2023; 90:102031. https://doi.org/10.1016/j.arr.2023.102031 [PubMed]
- 17. Davey Smith G, Hemani G. Mendelian randomization: genetic anchors for causal inference in epidemiological studies. Hum Mol Genet. 2014; 23:R89–98. https://doi.org/10.1093/hmg/ddu328 [PubMed]
- 18. Skrivankova VW, Richmond RC, Woolf BAR, Davies NM, Swanson SA, VanderWeele TJ, Timpson NJ, Higgins JPT, Dimou N, Langenberg C, Loder EW, Golub RM, Egger M, et al. Strengthening the reporting of observational studies in epidemiology using mendelian randomisation (STROBE-MR): explanation and elaboration. BMJ. 2021; 375:n2233. https://doi.org/10.1136/bmj.n2233 [PubMed]
- 19. Codd V, Wang Q, Allara E, Musicha C, Kaptoge S, Stoma S, Jiang T, Hamby SE, Braund PS, Bountziouka V, Budgeon CA, Denniff M, Swinfield C, et al. Polygenic basis and biomedical consequences of telomere length variation. Nat Genet. 2021; 53:1425–33. https://doi.org/10.1038/s41588-021-00944-6 [PubMed]
- 20. Strongman H, Brown A, Smeeth L, Bhaskaran K. Body mass index and Hodgkin's lymphoma: UK population-based cohort study of 5.8 million individuals. Br J Cancer. 2019; 120:768–70. https://doi.org/10.1038/s41416-019-0401-1 [PubMed]
- 21. Sergentanis TN, Kanavidis P, Michelakos T, Petridou ET. Cigarette smoking and risk of lymphoma in adults: a comprehensive meta-analysis on Hodgkin and non-Hodgkin disease. Eur J Cancer Prev. 2013; 22:131–50. https://doi.org/10.1097/CEJ.0b013e328355ed08 [PubMed]
- 22. Kurki MI, Karjalainen J, Palta P, Sipilä TP, Kristiansson K, Donner KM, Reeve MP, Laivuori H, Aavikko M, Kaunisto MA, Loukola A, Lahtela E, Mattsson H, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023; 613:508–18. https://doi.org/10.1038/s41586-022-05473-8 [PubMed]
- 23. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017; 32:377–89. https://doi.org/10.1007/s10654-017-0255-x [PubMed]
- 24. Yavorska OO, Burgess S. MendelianRandomization: an R package for performing Mendelian randomization analyses using summarized data. Int J Epidemiol. 2017; 46:1734–9. https://doi.org/10.1093/ije/dyx034 [PubMed]
- 25. Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015; 44:512–25. https://doi.org/10.1093/ije/dyv080 [PubMed]
- 26. Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet Epidemiol. 2016; 40:304–14. https://doi.org/10.1002/gepi.21965 [PubMed]
- 27. Verbanck M, Chen CY, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. 2018; 50:693–8. https://doi.org/10.1038/s41588-018-0099-7 [PubMed]
- 28. Hemani G, Zheng J, Elsworth B, Wade KH, Haberland V, Baird D, Laurin C, Burgess S, Bowden J, Langdon R, Tan VY, Yarmolinsky J, Shihab HA, et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018; 7:e34408. https://doi.org/10.7554/eLife.34408 [PubMed]
- 29. Pierce BL, Ahsan H, Vanderweele TJ. Power and instrument strength requirements for Mendelian randomization studies using multiple genetic variants. Int J Epidemiol. 2011; 40:740–52. https://doi.org/10.1093/ije/dyq151 [PubMed]
- 30. Shay JW, Wright WE. Telomeres and telomerase: three decades of progress. Nat Rev Genet. 2019; 20:299–309. https://doi.org/10.1038/s41576-019-0099-1 [PubMed]
- 31. Vakonaki E, Tsiminikaki K, Plaitis S, Fragkiadaki P, Tsoukalas D, Katsikantami I, Vaki G, Tzatzarakis MN, Spandidos DA, Tsatsakis AM. Common mental disorders and association with telomere length. Biomed Rep. 2018; 8:111–6. https://doi.org/10.3892/br.2018.1040 [PubMed]
- 32. Tsatsakis A, Oikonomopoulou T, Nikolouzakis TK, Vakonaki E, Tzatzarakis M, Flamourakis M, Renieri E, Fragkiadaki P, Iliaki E, Bachlitzanaki M, Karzi V, Katsikantami I, Kakridonis F, et al. Role of telomere length in human carcinogenesis (Review). Int J Oncol. 2023; 63:78. https://doi.org/10.3892/ijo.2023.5526 [PubMed]
- 33. Bernal A, Tusell L. Telomeres: Implications for Cancer Development. Int J Mol Sci. 2018; 19:294. https://doi.org/10.3390/ijms19010294 [PubMed]
- 34. Schratz KE, Gaysinskaya V, Cosner ZL, DeBoy EA, Xiang Z, Kasch-Semenza L, Florea L, Shah PD, Armanios M. Somatic reversion impacts myelodysplastic syndromes and acute myeloid leukemia evolution in the short telomere disorders. J Clin Invest. 2021; 131:e147598. https://doi.org/10.1172/JCI147598 [PubMed]
- 35. Haydeé Cottliar AS, Noriega MF, Narbaitz M, Rodríguez A, Slavutsky IR. Association between telomere length and BCL2 gene rearrangements in low- and high-grade non-Hodgkin lymphomas. Cancer Genet Cytogenet. 2006; 171:1–8. https://doi.org/10.1016/j.cancergencyto.2006.05.016 [PubMed]
- 36. Widmann TA, Herrmann M, Taha N, König J, Pfreundschuh M. Short telomeres in aggressive non-Hodgkin's lymphoma as a risk factor in lymphomagenesis. Exp Hematol. 2007; 35:939–46. https://doi.org/10.1016/j.exphem.2007.03.009 [PubMed]
- 37. Adamson DJ, King DJ, Haites NE. Significant telomere shortening in childhood leukemia. Cancer Genet Cytogenet. 1992; 61:204–6. https://doi.org/10.1016/0165-4608(92)90088-p [PubMed]
- 38. Song DY, Kim JA, Jeong D, Yun J, Kim SM, Lim K, Park SN, Im K, Choi S, Yoon SS, Lee DS. Telomere length and its correlation with gene mutations in chronic lymphocytic leukemia in a Korean population. PLoS One. 2019; 14:e0220177. https://doi.org/10.1371/journal.pone.0220177 [PubMed]
- 39. Jebaraj BM, Kienle D, Lechel A, Mertens D, Heuberger M, Ott G, Rosenwald A, Barth TF, Möller P, Zenz T, Döhner H, Stilgenbauer S. Telomere length in mantle cell lymphoma. Blood. 2013; 121:1184–7. https://doi.org/10.1182/blood-2012-08-452649 [PubMed]
- 40. Roos G, Kröber A, Grabowski P, Kienle D, Bühler A, Döhner H, Rosenquist R, Stilgenbauer S. Short telomeres are associated with genetic complexity, high-risk genomic aberrations, and short survival in chronic lymphocytic leukemia. Blood. 2008; 111:2246–52. https://doi.org/10.1182/blood-2007-05-092759 [PubMed]
- 41. Sellmann L, de Beer D, Bartels M, Opalka B, Nückel H, Dührsen U, Dürig J, Seifert M, Siemer D, Küppers R, Baerlocher GM, Röth A. Telomeres and prognosis in patients with chronic lymphocytic leukaemia. Int J Hematol. 2011; 93:74–82. https://doi.org/10.1007/s12185-010-0750-2 [PubMed]
- 42. Counter CM, Gupta J, Harley CB, Leber B, Bacchetti S. Telomerase activity in normal leukocytes and in hematologic malignancies. Blood. 1995; 85:2315–20. https://doi.org/10.1182/blood.V85.9.2315.bloodjournal8592315 [PubMed]
- 43. Furtado FM, Scheucher PS, Santana BA, Scatena NF, Calado RT, Rego EM, Matos DM, Falcão RP. Telomere length analysis in monoclonal B-cell lymphocytosis and chronic lymphocytic leukemia Binet A. Braz J Med Biol Res. 2017; 50:e6019. https://doi.org/10.1590/1414-431X20176019 [PubMed]
- 44. Maciejowski J, de Lange T. Telomeres in cancer: tumour suppression and genome instability. Nat Rev Mol Cell Biol. 2017; 18:175–86. https://doi.org/10.1038/nrm.2016.171 [PubMed]
- 45. Wang L, Xiao H, Zhang X, Wang C, Huang H. The role of telomeres and telomerase in hematologic malignancies and hematopoietic stem cell transplantation. J Hematol Oncol. 2014; 7:61. https://doi.org/10.1186/s13045-014-0061-9 [PubMed]
- 46. Küçüksolak M, Yılmaz S, Ballar-Kırmızıbayrak P, Bedir E. Potent telomerase activators from a novel sapogenin via biotransformation utilizing Camarosporium laburnicola, an endophytic fungus. Microb Cell Fact. 2023; 22:66. https://doi.org/10.1186/s12934-023-02069-3 [PubMed]
- 47. Duman S, Ekiz G, Yılmaz S, Yusufoglu H, Ballar Kırmızıbayrak P, Bedir E. Telomerase activators from 20(27)-octanor-cycloastragenol via biotransformation by the fungal endophytes. Bioorg Chem. 2021; 109:104708. https://doi.org/10.1016/j.bioorg.2021.104708 [PubMed]
- 48. Fragkiadaki P, Renieri E, Kalliantasi K, Kouvidi E, Apalaki E, Vakonaki E, Mamoulakis C, Spandidos DA, Tsatsakis A. Τelomerase inhibitors and activators in aging and cancer: A systematic review. Mol Med Rep. 2022; 25:158. https://doi.org/10.3892/mmr.2022.12674 [PubMed]
- 49. Jones CH, Pepper C, Baird DM. Telomere dysfunction and its role in haematological cancer. Br J Haematol. 2012; 156:573–87. https://doi.org/10.1111/j.1365-2141.2011.09022.x [PubMed]