



Introduction
Based on the Global Cancer Statistics Report, colorectal cancer (CRC) is currently the third most common cancer and the second leading cause of cancer death [1]. About 35% of all cancer patients are defined as having metastatic disease [2], of which less than 20% patients can survive more than 5 years from the date of diagnosis [3]. The therapeutic targets in clinical practice only include DNA microsatellite instability and the changes in B-Raf proto-oncogene (BRAF) gene [4]. Investigating the histological characteristics of CRC is conducive to the research of the biological characteristics of tumors. Because colorectal cancer’s migration and invasion are one of main factors to the severity of the disease, exploring the molecular mechanisms of proliferation, migration and invasion of CRC cells will contribute to the development of targeted therapy [5].
PTEN (phosphatase and tensin homolog deleted on chromosome 10) belongs to a cancer suppressor gene (one of the protein phosphatases). Its catalytic phosphatase active center is structurally similar to phosphatase, so it is named PTEN.
PTEN can inhibit cell cycle progression and induce cell death to stimulate angiogenesis [6]. Promoting PTEN expression in CRC cell lines will stop the activation of the phosphatidylinositol-3-kinase (PI3K)/sinkserine/threonine kinase (Akt) signaling pathway, which affects CRC cell growth, promotes apoptosis, and improves sensitivity to targeted therapy, immunotherapy, and conventional therapy. Therefore, targeting drugs molecules in the PTEN signaling pathway may be another strategy to suppress tumor cells by inhibiting tumor activity [7, 8]. The study of oncogenes or suppressor genes crossing with PTEN will facilitate drug discovery and interpretation of chemotherapy resistance mechanisms.
Recombinant Kinesin Family, Member 18A (KIF18A) belongs to Recombinant Kinesin Family [9], which is highly expressed in tumors and associated with poor prognosis [10, 11]. It is a necessity of cell division and plays a role by regulating microtubule dynamics and chromosome aggregation [12]. In lung adenocarcinoma and prostate cancer, overexpression of KIF18A can motivate cell proliferation and suppress apoptosis. Furthermore, it is associated with tumor development [13, 14], and involved in invasion and metastasis [15]. The study focused on the role of KIF18A in colorectal cancer cell progression, migration and invasion, along with the pathways that KIF18A may affect. What’s more, it offered a new target and therapeutic strategy for the treatment of CRC.
Methods
Cell culture
SW480 and HT29 (Human CRC cell lines) were bought from Procell (Wuhan, China) and they were cultured in DMEM with 10% fetal bovine serum and 1% penicillin/streptomycin. The cells were placed into a humid atmosphere of 37°C and with 5% carbon dioxide while the PTEN inhibitor SF1670 (MCE) was used to inhibit PTEN expression at a concentration of 10 μm.
Transcriptome sequencing analysis
SW480 cells (1 × 106) were put into each well of a 6-well plate. After 24 hours, the original medium was discarded, and the medium containing Rh3 (50 μg/mL) was added into the plate for another 24 hours. Then, the medium was removed, and the cells were rinsed with PBS, lysed and preserved with Trizol. GENEWIZ Co. (Beijing, China) carried on RNA purification, library preparation and RNA sequencing. Other steps were as follow: All RNA was extracted with Trizol; The libraries science was set by means of the NEBNext® Ultra™ RNA Library Prep Kit for Illumina® (NEB, Ipswich, MA, USA), and library preparations were sequenced on the Illumina Novaseq platform. The majority of the data of this research was analyzed and visualized by R packages. FastQC was used to check the quality of the reads. The adapter was cut out using Cutadapt (version 1.9.1) and mapped to the Ensembl human reference genome (EnsemblGRCh37 release 98) through Hisat2 (v2.0.2) aligner software. Differentially expressed genes (DEGs) were identified utilizing the R package, EdgeR. This package employs a negative binomial distribution model to account for both biological and technical variability inherent in RNA-seq data. Normalization of the raw read counts for each gene was achieved using the Trimmed Mean of M-values (TMM) method, which can adjust for the variances in library size and RNA composition. To define upregulation and downregulation, the log2 fold change (log2FC) for each gene was calculated. Genes exhibiting a log2FC greater than 1 were classified as upregulated, whereas those with a log2FC less than -1 were categorized as downregulated.
For statistical analysis, we employed the exact test provided by EdgeR, which is predicated on the negative binomial distribution. This test facilitated the comparison of gene expression differences between the experimental and control groups. To adjust for multiple testing, we utilized the Benjamini-Hochberg procedure, and genes with an adjusted p-value less than 0.05 were deemed significantly differentially expressed. The quantification of DEG expressions was determined using the counts per million (CPM) method, which can normalize gene counts based on the total number of reads in each sample, thereby enabling a fair comparison of gene expression levels across different samples. Visualization of these results was accomplished by generating a heatmap using the R tool. Lastly, GSEA version 4.1.0 was employed to execute and visualize Gene Ontology (GO) and KEGG GSEA queries.
Cell infection
The targeting sequences for KIF18A (5′-GACUCAGACUCCAACGAAUTT-3′) was mounted on a lentiviral vector, respectively. A scrambled shRNA lentiviral vector was used as a negative control. To construct the overexpression vectors, the coding sequences of KIF18A were linked into a lentiviral vector driven by the U6 promoter. HT29 and SW480 Cells were transfected with the shRNA and overexpression recombinant lentiviral vectors in accordance with the instructions [13]. Then SW480 and HT29 cells were divided into four groups including sh-control group, sh-RNA KIF18A group, control group and KIF18A-OE group respectively.
Experiments of subcutaneous tumorigenesis in nude mice
Five-week-old female BALB c nude mice were purchased from Nanjing Biomedical Research Institute of Nanjing University (Nanjing, China). SW480 cells were infected subcutaneously into each side of the back of every mouse for tumorigenesis experiments. The size of tumor was then detected by measuring the length (L) along with width (W) of the tumor with calipers every 3 days, and the tumor volume (V) was calculated by using the formula V = 1/2 × L × W [16].
HE staining
The tumor tissue sections were dewaxed. The sections were then added to distilled water, and stained with an aqueous solution of hematoxylin for several minutes. The Colors of each section were separated in acid water and ammonia for a few seconds. The sections were rinsed under running water for 1 hour and rinsed with the addition of distilled water for a while. Then, the sections were dehydrated in 70% and 90% alcohol for 10 min each and stained with alcohol eosin staining solution for 3 minutes. The stained sections were dehydrated by pure alcohol and cleared with xylene [17]. The transparent portion was dropped with neutral gum and covered with a coverslip.
Scratch wound assay
The cell migration capacity was measured by scratch wound assay. We scratched on the monolayers with sterile pipette tips (200 μL) and rinsed off the natant and separated cells with serum-free medium while the treated SW480 and HT29 cells were forming confluent monolayers in the wells of the 6-well plate [18]. Lastly, we took images with a reverse fluorescence microscope at 0 h and 48 h respectively [19]. For each image, we chose three locations to detect the width of the scratch at random and then averaged the results.
Transwell
Inserts pre-coated with matrigel was used to perform Transwell so that the migration and invasion ability of SW480 and HT29 cell could be tested. Next, the suspension of 4 × 105 cells was added to the supraventricular and the culture medium to the lower chamber in the migration experiment, while the lower chamber was added with a layer of culture medium and a layer of matrigel in the invasion experiment. After 48 hours, the non-invasive cells on the upper side of the membrane were cleaned up and the cells locating on the other side outside the membrane were treated with cold methanol (−20°C) and then dried in air [20]. Finally, the cells were stained with crystal violet and counted with an inverted microscope. The experiment was repeated three times.
Western blotting
The total protein was extracted from SW480 cells with modified RIPA buffer solution. The proteins with equal amounts were split on 10% polyacrylamide gels, and then transferred to polyvinylidene fluoride membranes, which were put into the primary antibody KIF18A (Abcam, ab72417, 1:2000), PTEN (Abcam, ab170941, 1:1000), p-PI3K (Abcam, ab182651, 1:500), p-Akt (Abcam, ab38449, 1:1000), MMP2 (Abcam, ab92536, 1:1000), MMP9 (Abcam, ab76003, 1:1000) and anti-GAPDH (Abeam, ab128915, 1:10000) at 4°C overnight after blocked with 5% BSA. After that, we incubated the membranes with horseradish peroxidase-conjugated secondary antibodies and observed them with an ECL chemiluminescence system [21, 22]. Densitometric analysis was performed with a Bio-Rad image detection system and Quantity One software (Bio-Rad, Hercules, CA, USA). Each experiment was repeated in triplicate for statistical analysis.
Statistical analysis
The statistical analyses were performed using GraphPad Prism 9.0 software. Comparisons among multiple sets of samples were performed with Studen’s t-test or one-way ANOVA with post-hoc testing (Dunnett multiple comparison). If P value is less than 0.05, the data is of statistical significance. Data is presented as mean ± standard deviations. Each experiment is performed in triplicate.
Results
Decreased expression of KIF18A has something to do with the malignancy of colorectal cancer
To evaluate the effect of KIF18A on CRC progression, the expression abundance of KIF18A in cancers was analysed according to the public data. The clinical data and the FPKM (fragments per kilobase of transcript per million mapped reads) matrices were downloaded from GEO (GEO 24572). Compared with normal tissue, the transcription abundance of KIF18A was promoted in most of the solid tumour tissues, including breast cancer, bladder cancer, colorectal cancer etc. (Figure 1A, 1B). In colorectal cancer, it was obvious that the expression of KIF18A was promoted in tumour tissues (275 vs. 349, P = 0.017) (Figure 1C). Differences in gene expression between the shKIF18A-SW480 and vector-SW480 cell lines (shown in Figure 1D) were compared by the transcriptome analysis.
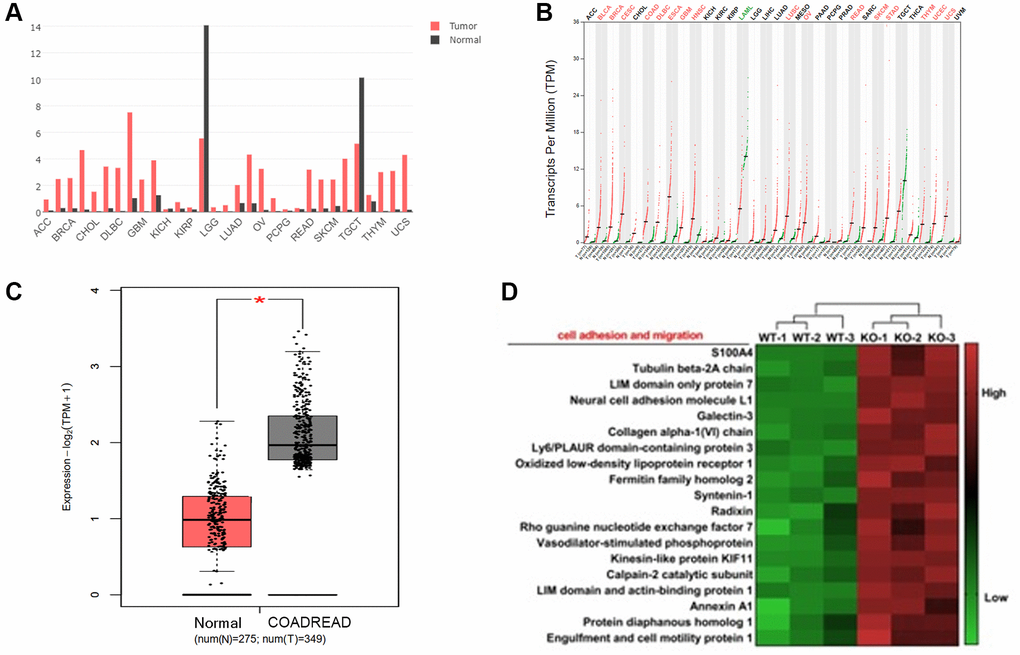
Figure 1. The expression of KIF18A in colorectal cancer tissues. (A, B) The transcriptional level (FPKM value) of KIF18A in pan-cancer tissues from the TCGA database. (Abbreviations: ACC: Adrenocortical Cancer; BRCA: Breast Invasive Carcinoma (Breast Cancer); CHOL: Cholangiocarcinoma (Bile Duct Cancer); DLBC: Diffuse Large B-Cell Lymphoma; GMB: Glioblastoma Multiforme (Brain Cancer); KICH: Kidney Chromophobe; KIRP: Kidney Renal Papillary Cell Carcinoma; LGG: Lower Grade Glioma; LUAD: Lung Adenocarcinoma; OV: Ovarian Serous Cystadenocarcinoma (Ovarian Cancer); PCPG: Pheochromocytoma and Paraganglioma; READ: Rectum Adenocarcinoma; SKCM: Skin Cutaneous Melanoma; TGCT: Testicular Germ Cell Tumors; THYM: Thymoma; UCS: Uterine Carcinosarcoma)). (C) The transcriptional level (FPKM value) of KIF18A in CRC and normal samples from the TCGA database. (COADREAD: Colon and Rectal Cancer). (D) The heatmap of genes involved in the cell adhesion and migration in shKIF18A-SW480 cells.
KIF18A can promote the progression of CRC
For further investigate the effect of KIF18A, we transfected SW480 cell lines with shKIF18A lentivirus and KIF18A overexpressing lentivirus. After transfecting, we injected them subcutaneously into nude mice to measure the size of tumors. It was shown that the size of tumors in the sh-RNA KIF18A group was distinctly smaller than those in the sh-control group. We also found that the size of tumors in the KIF18A-OE group was significantly larger than the control group. This result was then verified by HE staining (Figure 2). What mentioned above suggests that KIF18A can promote the progression of CRC.
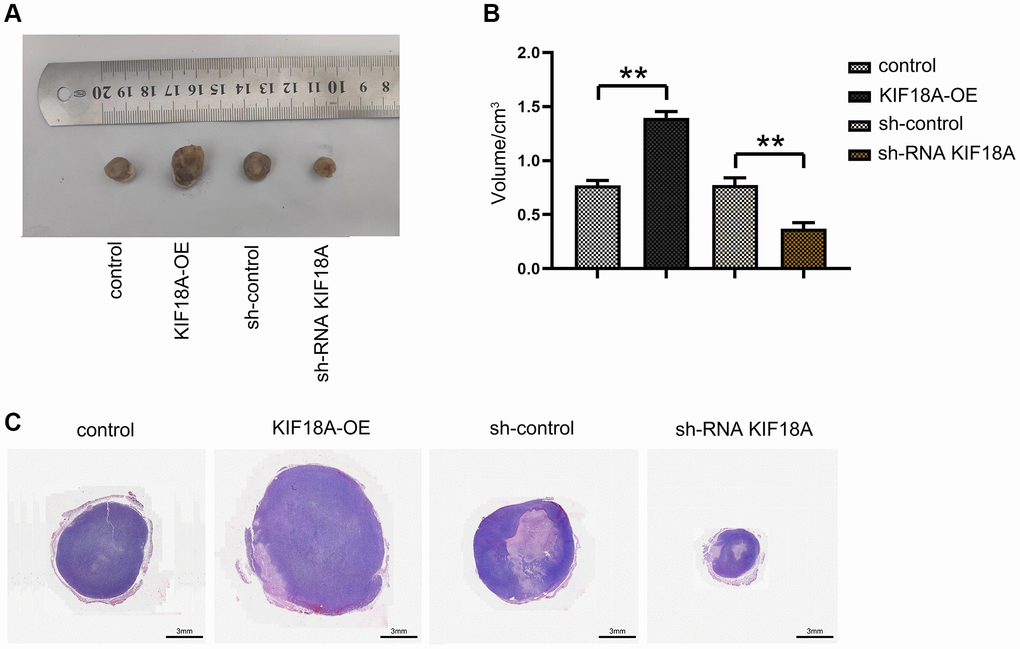
Figure 2. Effect of KIF18A on the development of CRC. (A) Diagrams of tumorigenesis in nude mice in sh-control group, sh-RNA KIF18Agroup, control group, KIF18A-OE group. (B) Statistics of tumor volume in nude mice in sh-control group, sh-RNA KIF18A group, control group, KIF18A-OE group. (C) Plots of HE staining results (**p < 0.01; N = 6).
KIF18A activates the PI3K/Akt signaling pathway by targeting PTEN to affect the expression of MMPs
In order to determine whether KIF18A can regulate the expression of PTEN signaling pathway, we transfected shKIF18A lentivirus and KIF18A overexpressing lentivirus into SW480 cells.
Western blotting results displayed that the relative protein expressions of KIF18A, p-PI3K, p-Akt, MMP2 and MMP9 were remarkably decreased and PTEN expression was signally risen in sh-RNA KIF18A group compared with the sh-control group, in the meantime, relative protein expressions of KIF18A, p-PI3K, p-Akt, MMP2 and MMP9 were going up and PTEN expression was dropping off in the KIF18A-OE group compared with the control group as plain as day (Figure 3).
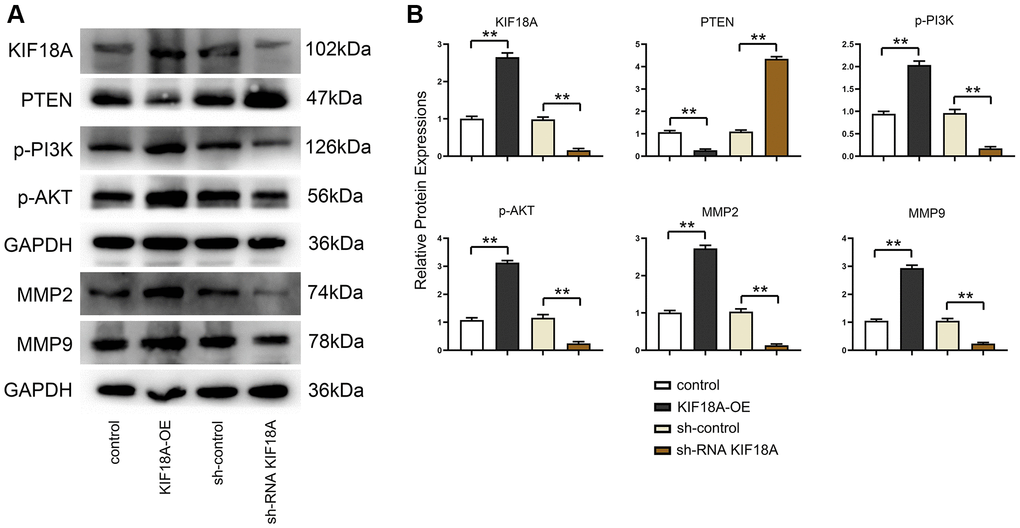
Figure 3. Influence of KIF18A on PTEN and PI3K/Akt signaling pathway. (A) Protein band diagrams of KIF18A, PTEN, p-PI3K, p-Akt, MMP2 and MMP9 in sh-control group, sh-RNA KIF18A group, control group, KIF18A-OE group. (B) Relative protein expressions of KIF18A, PTEN, p-PI3K, p-Akt, MMP2 and MMP9 in sh-control group, sh-RNA KIF18A group, control group, KIF18A-OE group (**p < 0.01; N = 3).
KIF18A can promote the migration and invasion of CRC cells
We tested the migration and invasion ability of SW480 AND HT29 cells by scratch wound assay, and Transwell assay was conducted for researching the biological functions of KIF18A and CRC. Based on scratch wound assay, sh-RNA KIF18A group significantly increased the cell scratch space in relative to sh-control group at 48 h. Meanwhile, in KIF18A-OE group, the cell scratch space was observably reduced in contrast to control group (Figure 4). In the migration experiment, the results suggested that the amount of migrated cells in sh-RNA KIF18A group was remarkably cut down compared with sh-control group, and the number of migrated cells in KIF18A-OE group was obviously risen compared with the control group. Invasion tests showed that the number of invasive cells in sh-RNA KIF18A group was reduced obviously in comparison with sh-control group. The number of invading cells was significantly risen in the KIF18A-OE group compared with the control group (Figure 5). The above-mentioned results indicate that KIF18A can improve the migration and invasion ability of CRC cells.
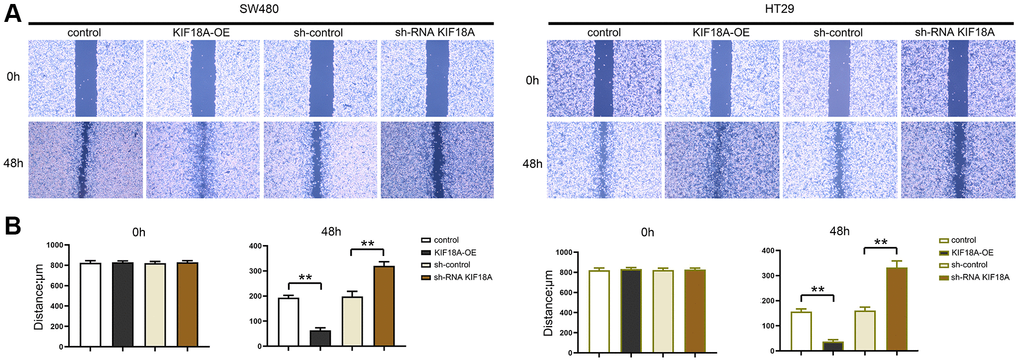
Figure 4. Effect of KIF18A on CRC cells migration ability. (A) Scratch wound diagrams at 0 h and 48 h in sh-control group, sh-RNA KIF18A group, control group, KIF18A-OE group. (B) Statistics of scratch-wound space at 0 h and 48 h in sh-control group, sh-RNA KIF18A group, control group, KIF18A-OE group (**p < 0.01; N = 3).
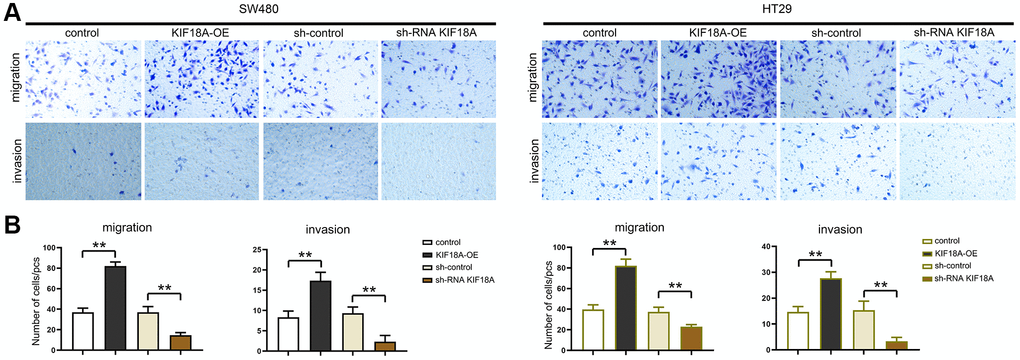
Figure 5. Influence of KIF18A on CRC cells migration and invasion ability. (A) Result diagrams of migration and invasion experiments in sh-control group, sh-RNA KIF18Agroup, control group, KIF18A-OE group. (B) Statistics of migration and invasion cells in sh-control group, sh-RNA KIF18A group, control group, KIF18A-OE group (**p < 0.01; N = 3).
KIF18A can promote migration and invasion of CRC cells through targeting PTEN to activate PI3K/Akt signaling pathway
Western blotting results displayed that the relative protein expressions of KIF18A, p-PI3K, p-Akt, MMP2 and MMP9 were remarkably decreased and PTEN expression was signally risen in sh-RNA KIF18A group compared with sh-control group (Figure 5).
After adding the PTEN inhibitor SF1670, we found that there were not significant differences of the relative protein expressions of PTEN, p-PI3K, p-Akt, MMP2 and MMP9 between the sh-RNA KIF18A+SF1670 group and the control+SF1670 group. Besides, another finding was that the number of migration and invasion cells in sh-RNA KIF18A group was significantly lower than control group through Transwell. However, there was no significant difference in the number of migration and invasion cells between the control+SF1670 group and the sh-RNA KIF18A+SF1670 group (Figure 6), which indicated that KIF18A can increase the migration and invasion of CRC cells by inhibiting PTEN to promote the activation of PI3K/Akt signaling pathway (Figure 7).
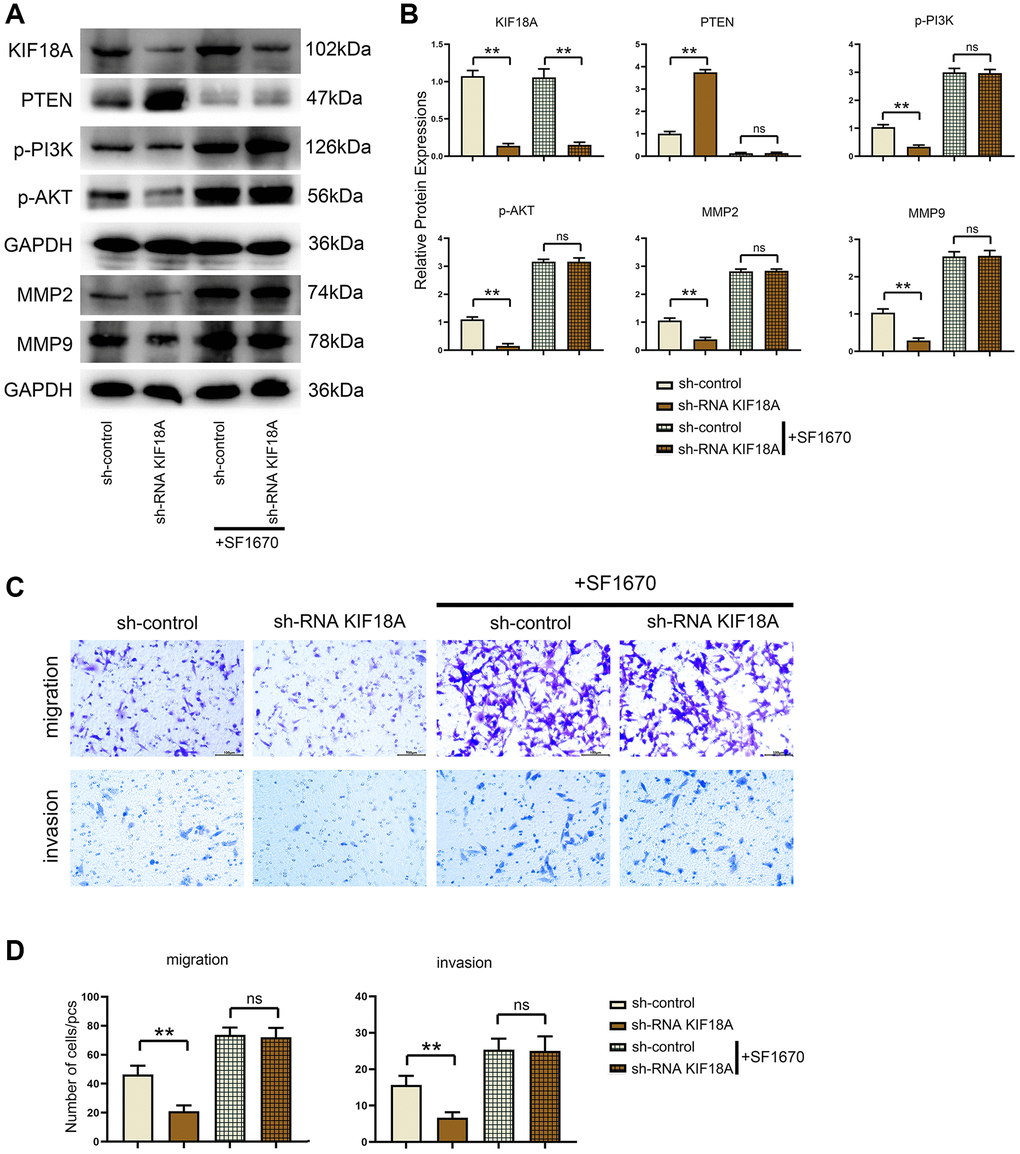
Figure 6. Influences of PTEN inhibitor on PTEN and PI3K/Akt signaling pathways and migration and invasion of CRC cells in control and KIF18A-OE groups. (A) Protein band diagrams of PTEN, p-PI3K, p-Akt, MMP2 and MMP9 in control group, KIF18A-OE group, control+SF1670 group and sh-RNA KIF18A+SF1670 group. (B) Relative protein expressions of PTEN, p-PI3K, p-Akt, MMP2 and MMP9 in control group, KIF18A-OE group, control+SF1670 group and sh-RNA KIF18A+SF1670 group. (C) Diagrams of migration and invasion experiments in control group, KIF18A-OE group, control+SF1670 group and sh-RNA KIF18A+SF1670 group. (D) Statistics of migration and invasion cells in control group, KIF18A-OE group, control+SF1670 group and sh-RNA KIF18A+SF1670 group (**p < 0.01; ns p > 0.05; N = 3).
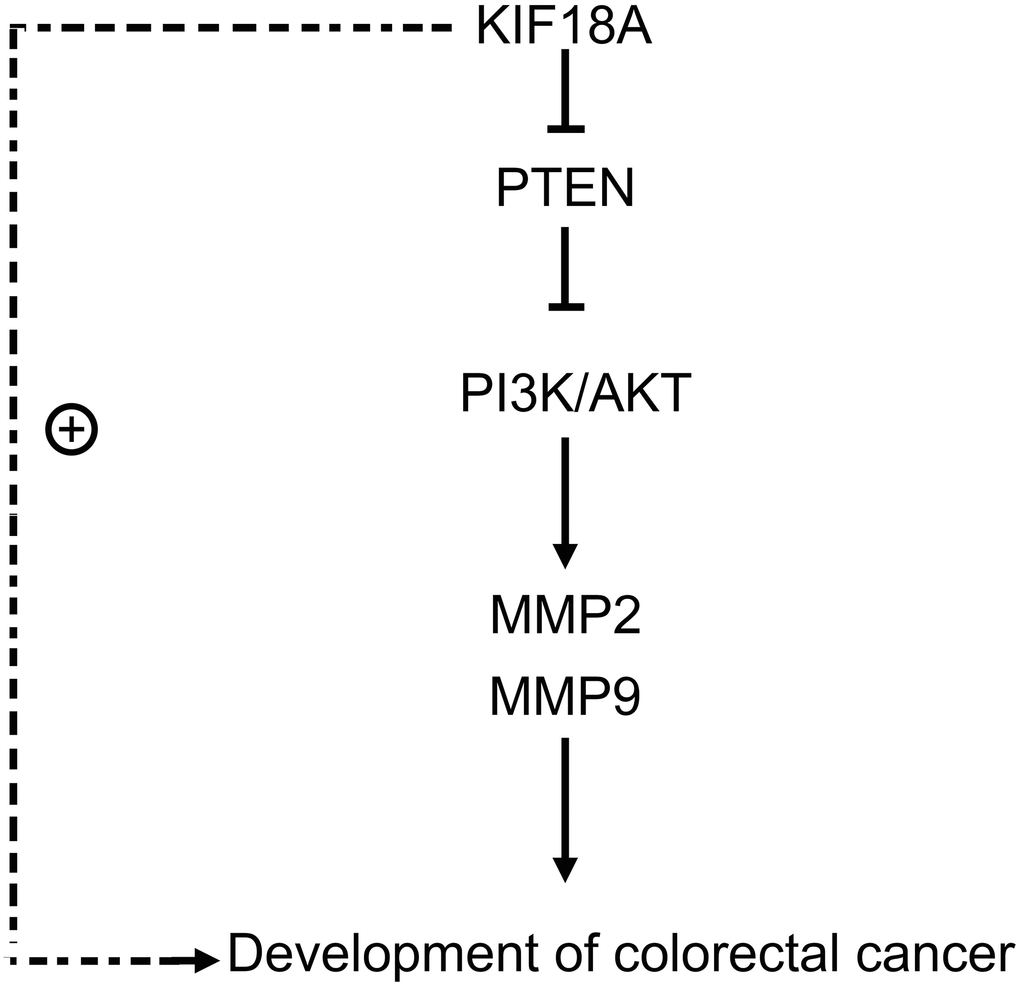
Figure 7. KIF18A can mediate PTEN, activate the PI3K/Akt signaling pathway, and promote the migration and invasion of CRC cells.
Discussion
CRC is one of the most common malignancies, and its occurrence and development are related to multiple gene abnormalities and signal pathway dysregulation. Despite increasing progress in targeted therapy and immunotherapy, the overall survival of metastatic colorectal cancer is poor [23–25]. Studying the mechanism of metastasis is help to the development of new therapeutic targets. Research steps of this study are as follows. First, by analyzing the published data of the expression of TCGA and GEO, we found that the expression of KIF18A gene is increased in tumor tissues. What’s more, the expression of genes containing cell proliferation, cell cycle, cell adhesion and migration as well as EMT signaling pathway is up-regulated in KIF18A knockout cells, so we concluded that KIF18A may also directly or indirectly regulate the expression of genes including these four signal transduction. In previous studies, it was reported that KIF18A was involved in cell proliferation, migration as well as invasion of esophageal cancer [26] and chromosomally unstable tumor cells [27, 28]. However, we have never discovered the regulatory molecules and signaling pathways. In the research, we sought to uncover the molecules that interact with KIF18A and explore the effect of KIF18A on CRC and its mechanism of action.
KIF18A, as kinesin-like protein 18A, is part of kinesin superfamily. It plays a key role in microtubule dynamics as it helps to maintain proper chromosome arrangement. KIF18A consists of a motor domain (responsible for ATP-dependent movement along microtubules) and a coil domain, which promotes double protein interactions. It has been shown to interact with several proteins involved in mitotic spindle assembly. Additionally, KIF18 interacts with tumors to inhibit p53, exhibiting a potential regulatory role in cancer progression. Current studies have uncovered that KIF18A has a high expression in many human tumors, including CRC [29–32]. However, its functional molecular mechanisms in CRC have not been fully explored. In the study, we found that PTEN expression was damaged in KIF18A knockout cells, but increased in KIF18A overexpression cells, which indicated that KIF18A can target PTEN to regulate CRC cells and also promote the activation of PI3K/Akt signaling pathway, leading to the upregulation of MMP2 and MMP9 expression.
PTEN is an impressive tumor suppressor, and its full name is Phosphatase and tensin homolog. It is considered an important factor in regulating embryonic development, cell survival and metabolic balance. PTEN can restrict Akt (protein kinase) signaling pathway and prevent cell proliferation and migration through catalyzing the conversion of phosphatidylinositol 3,5-triphosphate (PIP3) to phosphatidylinositol 4-bisphosphate (PIP2) [33]. Besides, PTEN is also involved in the pathogenesis and process of type I diabetes mellitus, dyslipidemia and nervous system diseases through transmembrane protein and enzyme mechanisms. Studies have shown that abnormal expression of PTEN gene is closely connected with the occurrence and development of many cancers like prostate cancer, breast cancer, ovarian cancer and so on. Therefore, studying PTEN will be conducive to a deeper understanding of the pathogenesis of cancer and provide new mechanisms for CRC treatment. In this study, it was found that the migration and invasion of CRC cells can be inhibited if we knockout KIF18A, and KIF18A overexpression can promote the migration and invasion of CRC cells. We discovered that KIF18A regulates CRC cells by targeting PTEN through PTEN inhibitor SF1670.
These findings show the important influence of KIF18A on the occurrence and development of colorectal cancer, providing new ideas for further exploring the pathogenesis of colorectal cancer.
Conclusion
We found that KIF18A whose expression could be up-regulated with the increase of tumor volume as a tumor-promoting gene in colorectal cancer. Down-regulation of KIF18A in tumors can control PTEN expression and promote activation of PI3K/Akt signaling pathway. Moreover, inhibition of KIF18A can control CRC cell migration and invasion. Based on the above-mentioned results, we can conclude that KIF18A can mediate PTEN and motivate the activation of PI3K/Akt signaling pathway, resulting to the progression of CRC.
Author Contributions
Wenyan Zhao: Conceptualization, Writing - Original Draft. Yuan Liu: Visualization, Investigation. Ming Sun: Data analysis, Writing - Original Draft. Bin Zhang and Wenyan Zhao: Methodology, Ming Sun and Wenyan Zhao: Conceptualization, Writing- Reviewing and Editing.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Ethical Statement
This study was approved by Ethics Committee of Shengjing Hospital of China Medical University (2022PS092K).
Funding
This work was supported by grants from the Natural Science Foundation of Liaoning Province (2019-MS-385).
References
- 1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021; 71:209–49. https://doi.org/10.3322/caac.21660 [PubMed]
- 2. Zacharakis M, Xynos ID, Lazaris A, Smaro T, Kosmas C, Dokou A, Felekouras E, Antoniou E, Polyzos A, Sarantonis J, Syrios J, Zografos G, Papalambros A, Tsavaris N. Predictors of survival in stage IV metastatic colorectal cancer. Anticancer Res. 2010; 30:653–60. [PubMed]
- 3. Biller LH, Schrag D. Diagnosis and Treatment of Metastatic Colorectal Cancer: A Review. JAMA. 2021; 325:669–85. https://doi.org/10.1001/jama.2021.0106 [PubMed]
- 4. Punt CJ, Koopman M, Vermeulen L. From tumour heterogeneity to advances in precision treatment of colorectal cancer. Nat Rev Clin Oncol. 2017; 14:235–46. https://doi.org/10.1038/nrclinonc.2016.171 [PubMed]
- 5. Piawah S, Venook AP. Targeted therapy for colorectal cancer metastases: A review of current methods of molecularly targeted therapy and the use of tumor biomarkers in the treatment of metastatic colorectal cancer. Cancer. 2019; 125:4139–47. https://doi.org/10.1002/cncr.32163 [PubMed]
- 6. Molinari F, Frattini M. Functions and Regulation of the PTEN Gene in Colorectal Cancer. Front Oncol. 2014; 3:326. https://doi.org/10.3389/fonc.2013.00326 [PubMed]
- 7. Lin PC, Lin JK, Lin HH, Lan YT, Lin CC, Yang SH, Chen WS, Liang WY, Jiang JK, Chang SC. A comprehensive analysis of phosphatase and tensin homolog deleted on chromosome 10 (PTEN) loss in colorectal cancer. World J Surg Oncol. 2015; 13:186. https://doi.org/10.1186/s12957-015-0601-y [PubMed]
- 8. Kotelevets L, Scott MGH, Chastre E. Targeting PTEN in Colorectal Cancers. Adv Exp Med Biol. 2018; 1110:55–73. https://doi.org/10.1007/978-3-030-02771-1_5 [PubMed]
- 9. Qian LX, Cao X, Du MY, Ma CX, Zhu HM, Peng Y, Hu XY, He X, Yin L. KIF18A knockdown reduces proliferation, migration, invasion and enhances radiosensitivity of esophageal cancer. Biochem Biophys Res Commun. 2021; 557:192–8. https://doi.org/10.1016/j.bbrc.2021.04.020 [PubMed]
- 10. Liu T, Yang K, Chen J, Qi L, Zhou X, Wang P. Comprehensive Pan-Cancer Analysis of KIF18A as a Marker for Prognosis and Immunity. Biomolecules. 2023; 13:326. https://doi.org/10.3390/biom13020326 [PubMed]
- 11. Marquis C, Fonseca CL, Queen KA, Wood L, Vandal SE, Malaby HLH, Clayton JE, Stumpff J. Chromosomally unstable tumor cells specifically require KIF18A for proliferation. Nat Commun. 2021; 12:1213. https://doi.org/10.1038/s41467-021-21447-2 [PubMed]
- 12. Tao BY, Liu YY, Liu HY, Zhang ZH, Guan YQ, Wang H, Shi Y, Zhang J. Prognostic Biomarker KIF18A and Its Correlations With Immune Infiltrates and Mitosis in Glioma. Front Genet. 2022; 13:852049. https://doi.org/10.3389/fgene.2022.852049 [PubMed]
- 13. Yang J, Zhang Q, Yang Z, Shu J, Zhang L, Yao Y, Wang X, Liu X. KIF18A interacts with PPP1CA to promote the malignant development of glioblastoma. Exp Ther Med. 2023; 25:154. https://doi.org/10.3892/etm.2023.11853 [PubMed]
- 14. Malaby HL, Lessard DV, Berger CL, Stumpff J. KIF18A's neck linker permits navigation of microtubule-bound obstacles within the mitotic spindle. Life Sci Alliance. 2019; 2:e201800169. https://doi.org/10.26508/lsa.201800169 [PubMed]
- 15. Liu G, Cai G, He X, Huang D, Zhu G, Chen C, Zhang X. KIF18A promotes head and neck squamous cell carcinoma invasion and migration via activation of Akt signaling pathway. Transl Cancer Res. 2019; 8:2252–63. https://doi.org/10.21037/tcr.2019.09.38 [PubMed]
- 16. Ma C, Wang X, Yang F, Zang Y, Liu J, Wang X, Xu X, Li W, Jia J, Liu Z. Circular RNA hsa_circ_0004872 inhibits gastric cancer progression via the miR-224/Smad4/ADAR1 successive regulatory circuit. Mol Cancer. 2020; 19:157. https://doi.org/10.1186/s12943-020-01268-5 [PubMed]
- 17. Liu X, Wang W, Liu X, Zhang Z, Yu L, Li R, Guo D, Cai W, Quan X, Wu H, Dai M, Liang Z. Multi-omics analysis of intra-tumoural and inter-tumoural heterogeneity in pancreatic ductal adenocarcinoma. Clin Transl Med. 2022; 12:e670. https://doi.org/10.1002/ctm2.670 [PubMed]
- 18. Jonkman JE, Cathcart JA, Xu F, Bartolini ME, Amon JE, Stevens KM, Colarusso P. An introduction to the wound healing assay using live-cell microscopy. Cell Adh Migr. 2014; 8:440–51. https://doi.org/10.4161/cam.36224 [PubMed]
- 19. Levoux J, Prola A, Lafuste P, Gervais M, Chevallier N, Koumaiha Z, Kefi K, Braud L, Schmitt A, Yacia A, Schirmann A, Hersant B, Sid-Ahmed M, et al. Platelets Facilitate the Wound-Healing Capability of Mesenchymal Stem Cells by Mitochondrial Transfer and Metabolic Reprogramming. Cell Metab. 2021; 33:283–99.e9. https://doi.org/10.1016/j.cmet.2020.12.006 [PubMed]
- 20. Chen S, Morine Y, Tokuda K, Yamada S, Saito Y, Nishi M, Ikemoto T, Shimada M. Cancer-associated fibroblast-induced M2-polarized macrophages promote hepatocellular carcinoma progression via the plasminogen activator inhibitor-1 pathway. Int J Oncol. 2021; 59:59. https://doi.org/10.3892/ijo.2021.5239 [PubMed]
- 21. Zhang Z, Wu Q, Fang M, Liu Y, Jiang J, Feng Q, Hu R, Xu J. HERC3 directly targets RPL23A for ubiquitination degradation and further regulates Colorectal Cancer proliferation and the cell cycle. Int J Biol Sci. 2022; 18:3282–97. https://doi.org/10.7150/ijbs.72014 [PubMed]
- 22. Liao H, Zhang L, Lu S, Li W, Dong W. KIFC3 Promotes Proliferation, Migration, and Invasion in Colorectal Cancer via PI3K/AKT/mTOR Signaling Pathway. Front Genet. 2022; 13:848926. https://doi.org/10.3389/fgene.2022.848926 [PubMed]
- 23. Xi Y, Xu P. Global colorectal cancer burden in 2020 and projections to 2040. Transl Oncol. 2021; 14:101174. https://doi.org/10.1016/j.tranon.2021.101174 [PubMed]
- 24. Rong Z, Luo Z, Fu Z, Zhang P, Li T, Zhang J, Zhu Z, Yu Z, Li Q, Qiu Z, Huang C. The novel circSLC6A6/miR-1265/C2CD4A axis promotes colorectal cancer growth by suppressing p53 signaling pathway. J Exp Clin Cancer Res. 2021; 40:324. https://doi.org/10.1186/s13046-021-02126-y [PubMed]
- 25. Yang L, Zheng L, Xie X, Luo J, Yu J, Zhang L, Meng W, Zhou Y, Chen L, Ouyang D, Zhou H, Tan Z. Targeting PLA2G16, a lipid metabolism gene, by Ginsenoside Compound K to suppress the malignant progression of colorectal cancer. J Adv Res. 2021; 36:265–76. https://doi.org/10.1016/j.jare.2021.06.009 [PubMed]
- 26. Nagahara M, Nishida N, Iwatsuki M, Ishimaru S, Mimori K, Tanaka F, Nakagawa T, Sato T, Sugihara K, Hoon DS, Mori M. Kinesin 18A expression: clinical relevance to colorectal cancer progression. Int J Cancer. 2011; 129:2543–52. https://doi.org/10.1002/ijc.25916 [PubMed]
- 27. Zhong Y, Jiang L, Lin H, Li X, Long X, Zhou Y, Li B, Li Z. Overexpression of KIF18A promotes cell proliferation, inhibits apoptosis, and independently predicts unfavorable prognosis in lung adenocarcinoma. IUBMB Life. 2019; 71:942–55. https://doi.org/10.1002/iub.2030 [PubMed]
- 28. Luo W, Liao M, Liao Y, Chen X, Huang C, Fan J, Liao W. The role of kinesin KIF18A in the invasion and metastasis of hepatocellular carcinoma. World J Surg Oncol. 2018; 16:36. https://doi.org/10.1186/s12957-018-1342-5 [PubMed]
- 29. Alfarsi LH, Elansari R, Toss MS, Diez-Rodriguez M, Nolan CC, Ellis IO, Rakha EA, Green AR. Kinesin family member-18A (KIF18A) is a predictive biomarker of poor benefit from endocrine therapy in early ER+ breast cancer. Breast Cancer Res Treat. 2019; 173:93–102. https://doi.org/10.1007/s10549-018-4978-5 [PubMed]
- 30. Chen QI, Cao B, Nan N, Wang YU, Zhai XU, Li Y, Chong T. Elevated expression of KIF18A enhances cell proliferation and predicts poor survival in human clear cell renal carcinoma. Exp Ther Med. 2016; 12:377–83. https://doi.org/10.3892/etm.2016.3335 [PubMed]
- 31. Chen FT, Zhong FK. Kinesin Family Member 18A (KIF18A) Contributes to the Proliferation, Migration, and Invasion of Lung Adenocarcinoma Cells In Vitro and In Vivo. Dis Markers. 2019; 2019:6383685. https://doi.org/10.1155/2019/6383685 [PubMed]
- 32. Wang LB, Zhang XB, Liu J, Liu QJ. The Proliferation of Glioblastoma Is Contributed to Kinesin Family Member 18A and Medical Data Analysis of GBM. Front Genet. 2022; 13:858882. https://doi.org/10.3389/fgene.2022.858882 [PubMed]
- 33. Cetintas VB, Batada NN. Is there a causal link between PTEN deficient tumors and immunosuppressive tumor microenvironment? J Transl Med. 2020; 18:45. https://doi.org/10.1186/s12967-020-02219-w [PubMed]