



Introduction
Immune checkpoint inhibitors (ICIs), particularly anti-programmed death receptor 1 (PD-1) therapies, have dramatically changed the face of cancer treatment in recent years [1–4]. Unfortunately, in colorectal cancer (CRC), one of the top three most malignant and prevalent cancers, the anti-tumor effect of immune checkpoint inhibitors (ICIs) is limited to those with high microsatellite instability (MSI-H) [5–8]. As for the microsatellite stable (mSS) CRC patients (approximately 90% of the cases), the response rate is only 5% to 10% [9–11]. A recent clinical report, the REGONIVO trial, indicated that combined treatment with Regorafenib with anti-PD-1 antibody achieved an objective response rate of 33% in CRC patients [12]. However, there is still an urgent need to explore the resistance mechanisms and improve the efficacy of immune checkpoint inhibitors in CRC due to the limited size of current clinical practice and insufficient evidence [13].
Traditional epigenetic regulation refers to the chemical modification of DNA or histones that can regulate gene expression independently of genome sequence changes [14, 15]. Studies have widely exposed the profound effect of dysregulated epigenetic modification enzymes on human diseases including cancers [16–19]. Similar with the epigenetic regulation of DNAs, RNAs also carry hundreds of different sites for different post-transcriptional modifications [17]. N6-methyl adenosine (m6A) is the most important mRNA modification in eukaryotic cells [20]. Mechanistically, m6A modification is a reversible chemical process controlled dynamically by the balanced activity of m6A methyltransferase and demethylase [21]. Since m6A modification has been shown to play a crucial role in RNA translation, stability, and variable splicing, perturbation of the m6A component has been indicated as regulatory factors of human disease, particularly cancer [22]. Dysregulation of the m6A “write” protein, such as the methyltransferase-like 3 (METTL3) and methyltransferase-like 16 (METTL16), has been reported in the disease development in recent years [23, 24]. Nevertheless, the role of METTL16 in immune regulation of CRC has not been clarified yet.
In this work, we explored the correlation between METTL16 with PD-L1 expression and determined the effects of METTL16 on CRC growth and immune evasion. Our findings may provide novel target for the treatment of CRC.
Materials and Methods
Evaluation of clinical samples
Patients diagnosed with CRC (n=20) and hospitalized in Tangshan Traditional Chinese Medicine Hospital were recruited in this study. Tumor and adjacent non-tumor tissues were collected during surgical operation and made into paraffin-embedded samples or frozen into liquid nitrogen for further experiments. All patients have signed the informed consents.
Xenograft model
Male BALB/c nude mice that aged 4 to 6-week-old were bought from Vital River Laboratory (China). SW480 cells transfected with control or METTL16 overexpression vectors (5 x 106 cells/site) were suspended in 50 μl saline and inoculated into the fact pat on back. Tumor size was measured and calculated every three days. After feeding for 30 days, the tumors were collected and made into paraffine-embedded samples.
IHC staining
Paraffine-embedded tissues were dewaxed and blocked with goat serum. After incubation with primary antibody against PD-L1 (1:200, Abcam, USA), METTL16 (1:200, Abcam, USA), and Ki-67 (1:200, Abcam, USA), the samples were probed with HRP-conjugated secondary antibodies and reacted with DAB for visualization. Images were taken using a light microscope (Leica, Germany).
Cell lines and transfection
CRC cell lines NCM460, RKO, HCT116, SW480, Colo320, DLD-1, HCE8693, SW620, and HT29, and the normal human colon epithelial cell line FHC were purchased from Wuhan Procell and ATCC. NCM460, DLD-1, HCE8693, and Colo320 were maintained in RPMI-1640, RKO in MEM, HCT116 and HT29 in McCoy’s 5A, SW620, SW480 in Leibovitz’s L-15 medium that supplied with 10% FBS (BI, Israel) and 1% penicillin and streptomycin (Sigma, USA) and maintained in 37° C incubator with 5% CO2.
METTL16 proteins siRNAs and overexpression vectors were obtained from GenePharma (Shanghai, China). Cells were transfected with indicated oligonucleotides using the Lipofectamine 2000 reagent (Thermo, USA) as per manufacturer’s protocol. Transfection efficacy was examined 2 days after transfection.
Cell viability and proliferation detection
Cell viability was measured by cell counting kit-8 (CCK-8, SolarBio, China) following the manufacturer’s protocol. CRC cells were seeded into 96-well plate with 5,000 cells per well and incubated at 37° C incubator for indicated time points. Then CCK-8 reagent was added and incubated for 2 hours. The absorbance values at 450 nm were measured by a microplate detector. For colony formation assay, cells were seeded into 6-well plate and incubated for 10 days. The visible colonies were then stained with crystal violet for 20 min. Images were taken by a digital camera.
Cell apoptosis
Cell apoptosis was measured using Annexin V/PI apoptosis detection kit (Beyotime, China) in line with manufacturer’s introduction. In brief, cells were collected and suspended in binding buffer that contains Annexin V and PI reagent. After incubation for 30 minutes in dark, the samples were loaded in a flow cytometer (BD Bioscience, USA) and the portion of apoptotic cells were measured.
Transwell assay
CRC cells were suspended in serum-free medium and seeded into the upper chambers of Transwell (Costar, USA). The lower chambers were filled with complete culture medium. After incubation for 24 hours, the upper chambers were collected, and the inner side of chamber was wiped with cotton sticks. The migrated cells were then stained with crystal violet for 20 min. To measure cell invasion, the Transwell chambers were pre-coated with Matrigel that mixed with Leibovitz’s L-15 at 1:1. Images of cells were taken under a microscope (Leica, Germany) [25].
Wound healing experiment
CRC cells were seeded into 6-well plate and cultured overnight to confluence. The monolayer of cells was then scratched with a sterile 200 μl pipette and washed with PBS to remove debris. Cells were then incubated for 24 hours, and images of the wounds were taken at 0. 24, and 48 h after scratching.
Western blotting assay
Cell pellets were homogenized with RIPA lysis buffer and quantified with BCA kit. Protein samples containing 40 mg of protein were separated by SDS-PAGE gel and transferred onto PVDF membranes, followed by blocking in 5% skim milk for 2 hours. The blots were then probed with primary antibodies that target E-cadherin (1:2000, ab231303, Abcam, USA), Vimentin (1:2000, ab92547, Abcam, USA), METTL16 (1:2000, ab252420, Abcam, USA), and β-actin (1:2000, ab8226, Abcam, USA) overnight at 4° C. The membranes were then hatched with HRP-conjugated secondary antibodies and ECL reagent for visualization.
Quantitative real time PCR (qPCR)
Total RNAs were extracted using the Trizol solution (Qiagen, USA). Then a total of 1 μg RNA was subjected to reverse transcription reactions using a Reverse Transcription kit (Qiagen, USA). PCR amplification was conducted using the SYBR GREEN qPCR mix (Thermo, USA).
PD-L1 forward primer: 5’- TGGCATTTGCTGAACGCATTT-3’,
PD-L1 reverse primer: 5’-TGCAGCCAGGTCTAATTGTTTT-3’;
METTL16 forward primer: 5’-CTCTGACGTGTACTCTCCTAAGG -3’,
METTL16 reverse primer: 5’- TACCAGCCATTCAAGGTTGCT-3’.
RNA stability detection
To detect the stability of METTL16 mRNA, CRC cells were treated with α-amanitin (50 μM; Sigma, USA) for indicated hours. Then total RNA was extracted and reverse transcribed. The relative level of METTL16 RNA to 0 hour was measured by qPCR assay.
Protein degradation
CRC cells were treated with the protein synthesis inhibitor cycloheximide (CHX) for 0, 3, 6, 12 hours, then homogenized to collect total protein. The expression of PD-L1 was detected by western blot assay.
RNA immunoprecipitation (RIP)
The enrichment of METTL16 on PD-L1 RNA was measured by using EZ-Magna RIP Kit (Millipore, USA) as per manufacturer’s instruction. In brief, magnetic beads were pre-coated with anti-METTL16 antibody overnight at 4° C in rotation. Next day, the antibody-beads conjugates were hatched with the isolated total RNAs for 6 hours at 4° C. The precipitated RNAs were then eluted and purified. The level of PD-L1 was measured by qPCR.
MeRIP-qPCR assay
The RNA was collected from cells and were hatched with anti-m6A antibody (Thermo, USA) for 6 hours at 4° C. The Dynabeads (Invitrogen, USA) were blocked by 2% BSA (Thermo, USA) for 3 hours at 4° C, followed by incubation with aforementioned RNA-antibody complex at 4° C overnight. After elution with SDS buffer, the RNAs were collected and level of PD-L1 was quantified by with qPCR assay [26].
T cell activation and co-culture system
Human peripheral blood samples were obtained from donors and the peripheral blood mononuclear cells (PBMCs) were purified by Ficoll methods. For T cell activation, PBMCs were incubated with Dynabeads Human T-Activator CD3 and CD28 (Gibco, USA) in line with the manufacturer’s protocol for 3 days. For co-culture, T cells and CRC cells were placed in the upper and lower chambers of Transwell, respectively. The ratio of T cells and cancer cells was 10:1 and cultured for 3 days. To determine the activation of T cells, PMSCs were incubated with anti-CD4, anti-CD-8, and anti-PD-1 antibody and analyzed by a flow cytometer.
Statistics
All statistical analyses were performed using the SPSS and GraphPad Prism 7.0 Software. The comparison between experimental groups was conducted using two-sided Student’s t-test and one-way ANOVA followed by Dunnett’s multiple comparison. P < 0.05 was set as significant.
Results
Expression of METTL16 and PD-L1 in clinical samples and cell lines of CRC
We first analyzed the expression and correlation of METTL16 and PD-L1 in CRC. As shown in Figure 1A, 1B, the RNA level of PDL-1 in collected CRC tumor tissues is notably elevated, whereas the level of METTL16 is significantly decreased, compared with the non-tumor tissues. The IHC analysis of protein expression also indicated elevated PD-L1 and suppressed METTL16 expression in CRC tumor tissues compared with the paired non-tumor tissues (Figure 1C). Results from qPCR indicated a negative correlation between expression of METTL16 and PD-L1 (R=0.7035, p=0.0081). Moreover, we observed significantly decreased level of METTL16 in CRC cell lines, including NCM460, RKO, HCT116, SW480, Colo320, DLD-1, HCE8693, SW620, and HT29, compared with the normal human colon epithelial cell line FHC (Figure 1E). These data indicated that METTL16 may be negatively correlated with PD-L1 expression in CRC.
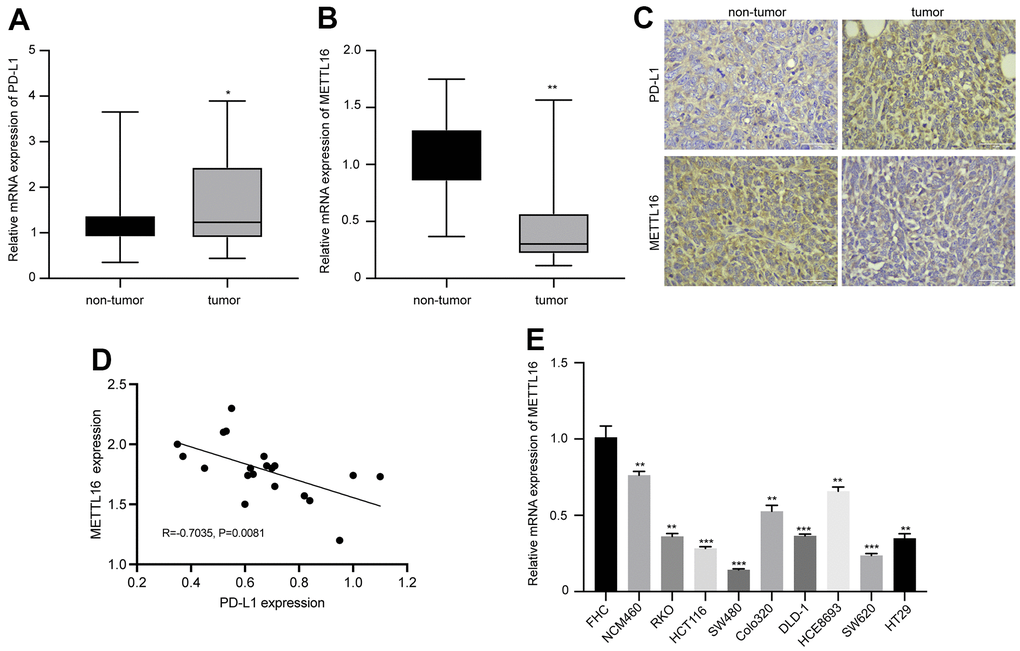
Figure 1. Expression of METTL16 and PD-L1 in clinical samples and cell lines of CRC. CRC tumor and non-tumor tissues were collected from patient with CRC. The RNA levels of (A) PD-L1 and (B) METTL16 were measured by qPCR assay. (C) The protein expression of PD-L1 and METTL16 was measured by IHC experiment. (D) The Pearson correlation analysis of PD-L1 and METTL16 in tumor tissues (n=20). (E) The RNA level of METTl16 in CRC cell lines (NCM460, RKO, HCT116, SW480, Colo320, DLD-1, HCE8693, SW620, and HT29) and normal human colon epithelial cell line FHC was measured by qPCR. *p<0.05, **p<0.01, ***p<0.001.
METTL16 regulates the proliferation, migration and invasion of CRC cells
Moreover, among the included CRC cell lines, the SW480 and SW620 exhibited the lowest level of METTL16 and hence were selected for further experiments. We subjected transfection of ectopic expression vectors and siRNAs of METTL16 in these two cell lines and assess cell proliferation, migration, and invasion. The results from western blotting assay demonstrated the successful overexpression and depletion of METTL16 in SW480 and SW620 cells (Figure 2A, 2B). As shown in Figure 2C, 2D, overexpression of METTL16 led to significantly suppressed growth (Figure 2C) and colonies (Figure 2D) of SW480 and SW620 cells. Besides, results from Transwell assay indicated that overexpression of METTL16 notably suppressed the number of migrated and invaded cells (Figure 3A). The suppressed healing of wounds in Figure 3B also suggested the impeded cell migration. Results from western blotting showed that overexpression of METTL16 caused increased level of E-cadherin and decreased level of Vimentin in SW480 and SW620 cells (Figure 3C), indicating the enhanced EMT. These data indicated that METTL16 suppressed CRC cell proliferation, migration and invasion.
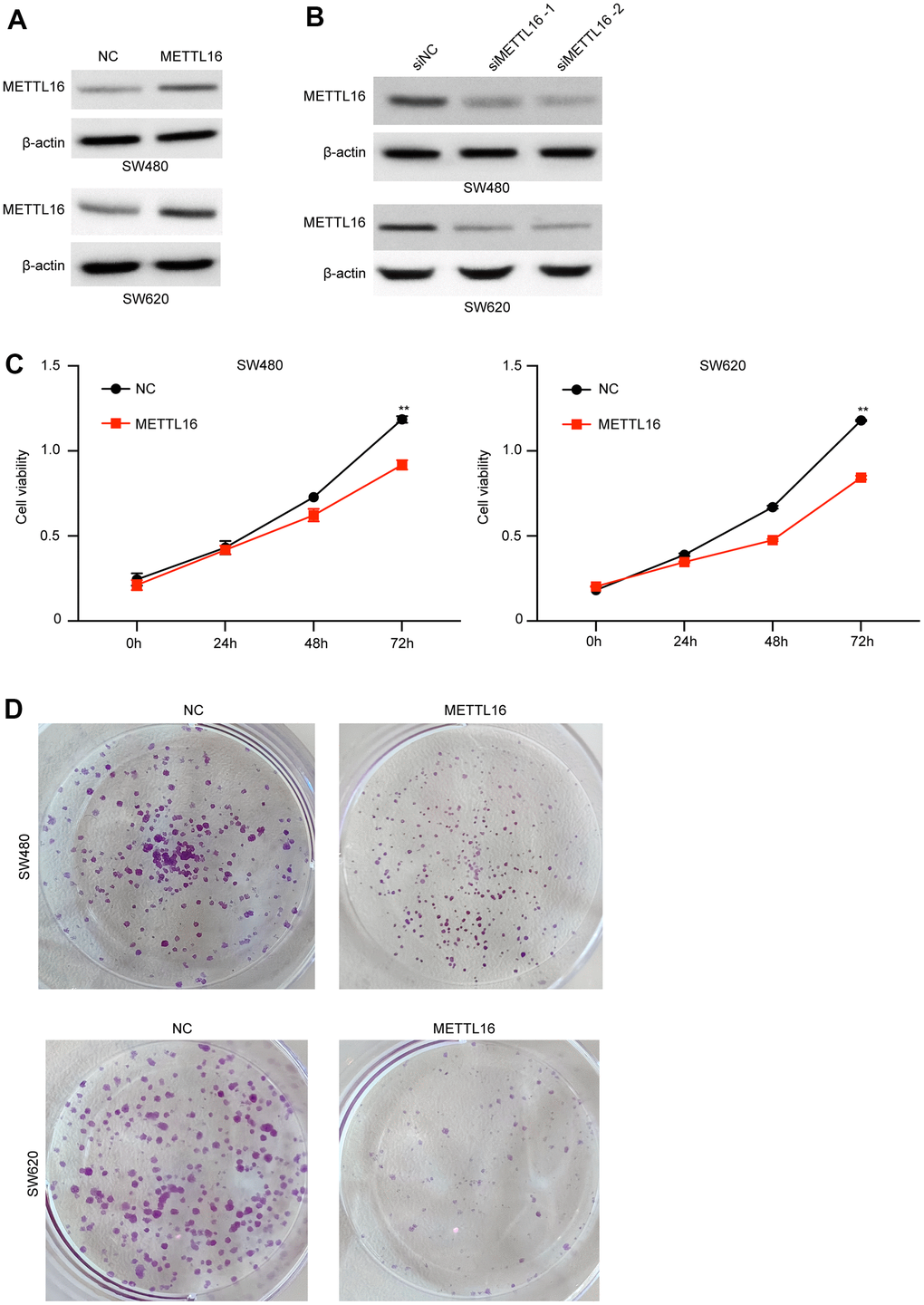
Figure 2. METTL16 regulates the proliferation of CRC cells. (A) SW480 and SW620 cells were transfected with METTL16 overexpression vectors, and protein level of METTL16 was measured by western blotting assay. (B) SW480 and SW620 cells were transfected with METTL16 siRNAs, then protein level of METTL16 was measured by western blotting assay. (C) Cell viability of SW480 and SW620 cells under overexpression of METTL16 was measured by CCK-8 assay. (D) Proliferation of SW480 and SW620 cells under overexpression of METTL16 was detected by using colony formation assay. **p<0.01.
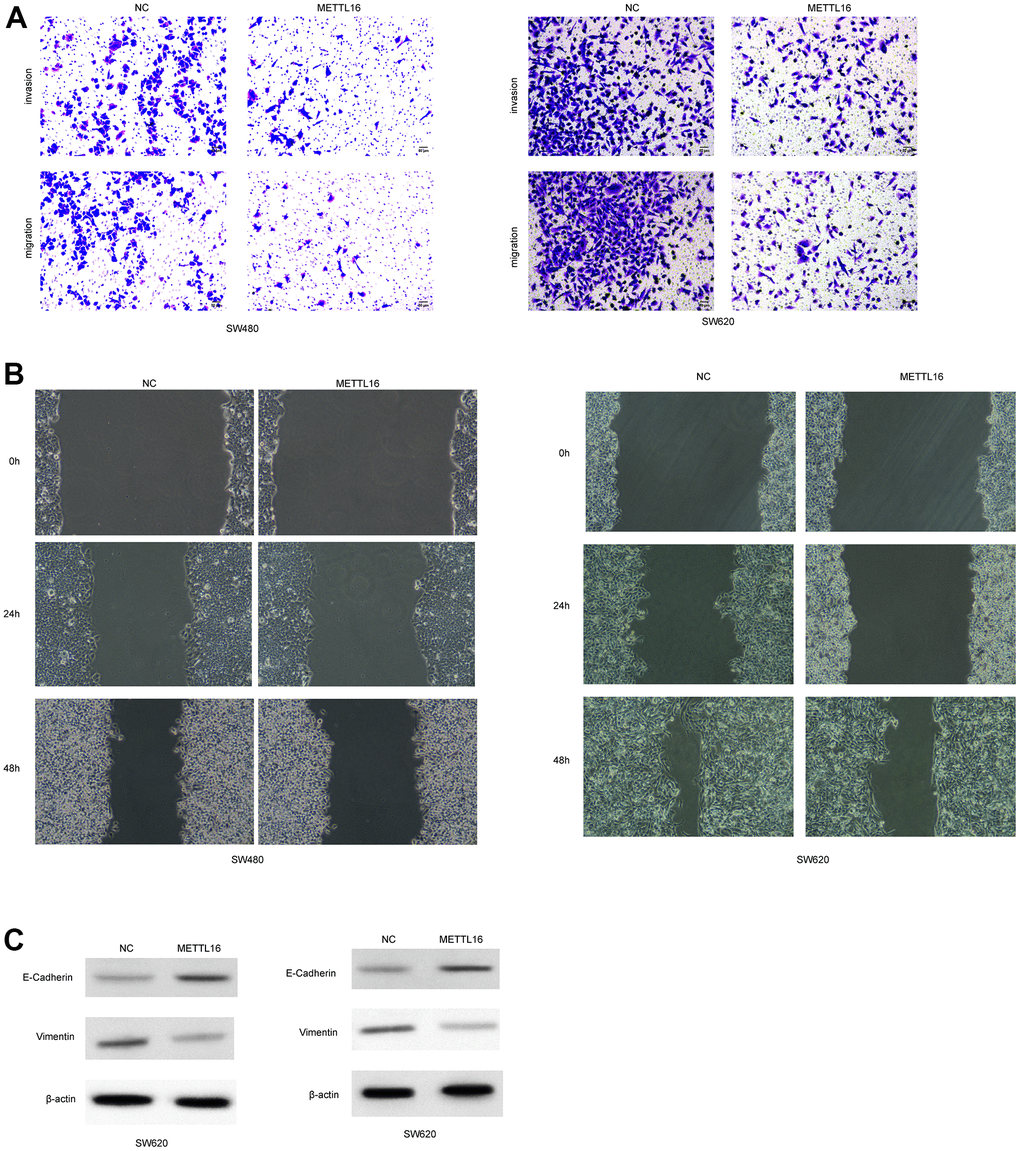
Figure 3. METTL16 regulates the migration and invasion of CRC cells. SW480 and SW620 cells were transfected with METTL16 overexpression vectors. (A) Cell migration and invasion were detected by using Transwell experiment. (B) Cell migration was measured by wound healing assay. (C) Protein levels of E-cadherin and Vimentin were detected by western blotting assay.
METTL16 modulates immune invasion in a CRC cell/T cell co-culture system
To explore the effects of METTL16 on immune invasion of CRC cells, we adopted a co-culture system of CRC cells with unstimulated or activated T cells and evaluated the activation, viability and apoptosis of T cells. As shown in Figure 1A, the portions of PD-1+ cell in activated T cells were notably decreased under co-culture with METTL16-overexpressed CRC cells, compared with those co-cultured with control CRC cells. Moreover, co-culture with CRC cells notably suppressed the viability and induced apoptosis of T cells (Figure 4B, 4C), which were significantly enhanced by depletion of METTL16 in CRC cells. Besides, overexpression of METTL16 reversed the suppressive effects of CRC cells on T cell viability (Figure 4B, 4C). These data suggested that METTL16 overexpression in CRC cells could promote immune response of T cells.
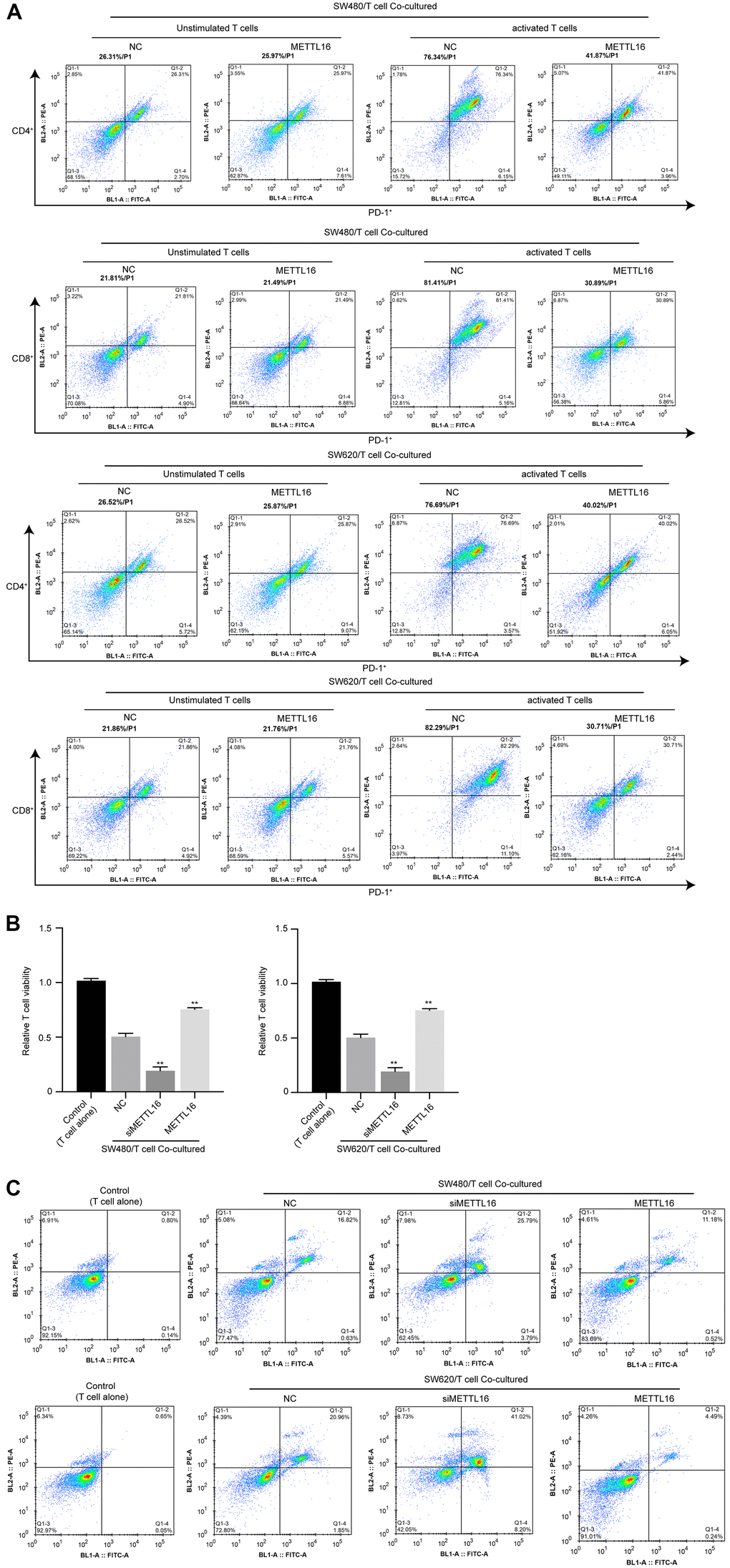
Figure 4. METTL16 modulates immune invasion in a CRC cell/T cell co-culture system. CRC cells were co-cultured with unstimulated or activated T cells. (A) The portion of PD-1+/CD4+ T cells and PD-1+/CD8+ T cells was measured by flow cytometry. (B) Cell viability was assessed by CCK-8 assay. (C) Cell apoptosis was measured by flow cytometry. **p<0.01.
METTL16-mediated m6A modification regulates PD-L1 expression in CRC cells
METTL16 is a well-recognized regulator of RNA methylation. Hence, we wondered if METTL16 directly modulates the expression of PD-L1. We observed that overexpression of METTL16 caused significant decrease in PD-L1 mRNA level (Figure 5A), and depletion of METTL16 markedly increased PD-L1 mRNA level (Figure 5B). We validated the elevated m6A modification level in several reported target of METTL16, including the RAB11B-AS1, MAT2A, and EEF1A1 (Supplementary Figure 1). Consistent with these findings, the results from RIP experiment indicated that the PD-L1 mRNA was modified by m6A (Figure 5C). Simultaneously, the m6A level of PD-L1 was upregulated by METTL16 overexpression and downregulated by METTL16 knockdown (Figure 5D, 5E), suggesting that METTL16 regulated the methylation of PD-L1 in CRC cells. Moreover, m6A modification has been reported to modulate RNA stability. Here, we examined the PD-L1 RNA stability after using α-amanitin to block new RNA synthesis. The degradation of PD-L1 mRNA was notably faster in METTL16-overexpressed cells, compared with the control cells (Figure 5F), and depletion of METTL16 maintained the stability of PD-L1 mRNA (Figure 5G). Moreover, treatment with Cycloheximide (CHX), the protein synthesis inhibitor, led to degradation of PD-L1 expression, and overexpression of METTL16 enhanced the PD-L1 degradation (Figure 5H). These data indicated that METTL16 could induce the m6A modification of PD-L1 mRNA and decreased its stability.
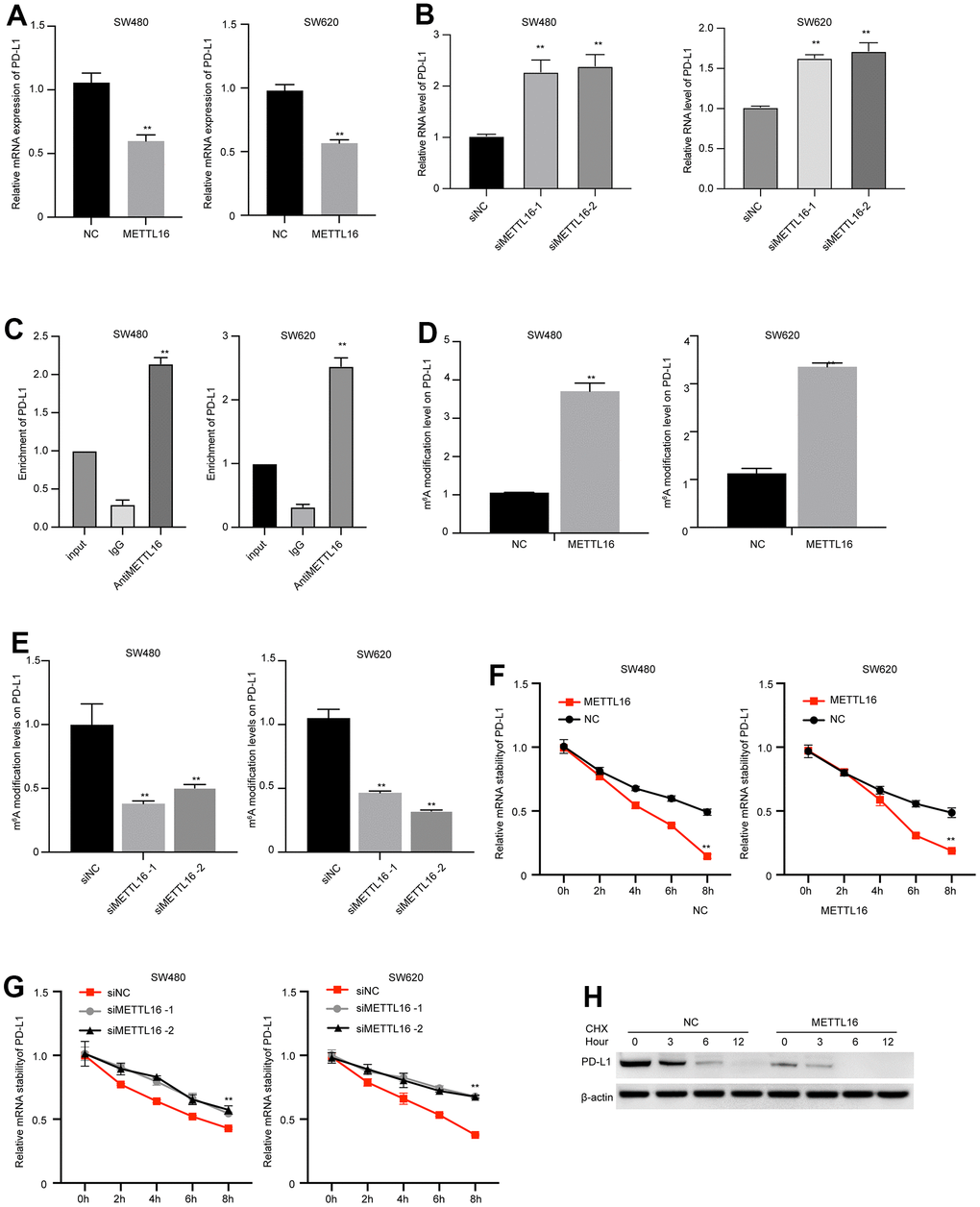
Figure 5. METTL16-mediated m6A modification regulates PD-L1 expression in CRC cells. (A, B) SW480 and SW620 cells were transfected with METTL16 overexpression vectors or siRNAs, then RNA level of PD-L1 was measured by qPCR assay. (C) The interaction between METTL16 with PD-L1 mRNA was measured by RIP assay. (D, E) SW480 and SW620 cells were transfected with METTL16 overexpression vectors or siRNAs, and MeRIP assay was conducted to assess m6A enrichment. (F, G) SW480 and SW620 cells were transfected with METTL16 overexpression vectors or siRNAs and treated with RNA synthesis inhibitor α-amanitin (50 μM). The stability of PD-L1 mRNA was checked by qPCR. (H) SW480 cells were treated with CHX for indicated time points and the expression of PD-L1 was measured by western blot. **p<0.01.
METTL16 modulates CRC cell growth in vivo
We next investigated the in vivo effects of METTL16 on CRC development. A xenograft tumor model was established using SW480 cells and treated with anti-PD-1 antibody. We observed that overexpression of METTL16 suppressed the tumor size and tumor growth curve and enhanced the tumor suppressive effect of anti-PD-1 antibody (Figure 6A, 6B). Analysis on tumor tissues demonstrated that overexpression of METTL16 and blocking of PD-1 could notably suppress the expression of proliferative biomarker Ki-67 (Figure 6C). Moreover, the levels of CD4 and CD8 positive cells in tumor tissues were notably elevated by overexpression of METTL16 and anti-PD-1 treatment compared with the control group and were further increased by their combination (Figure 6D).
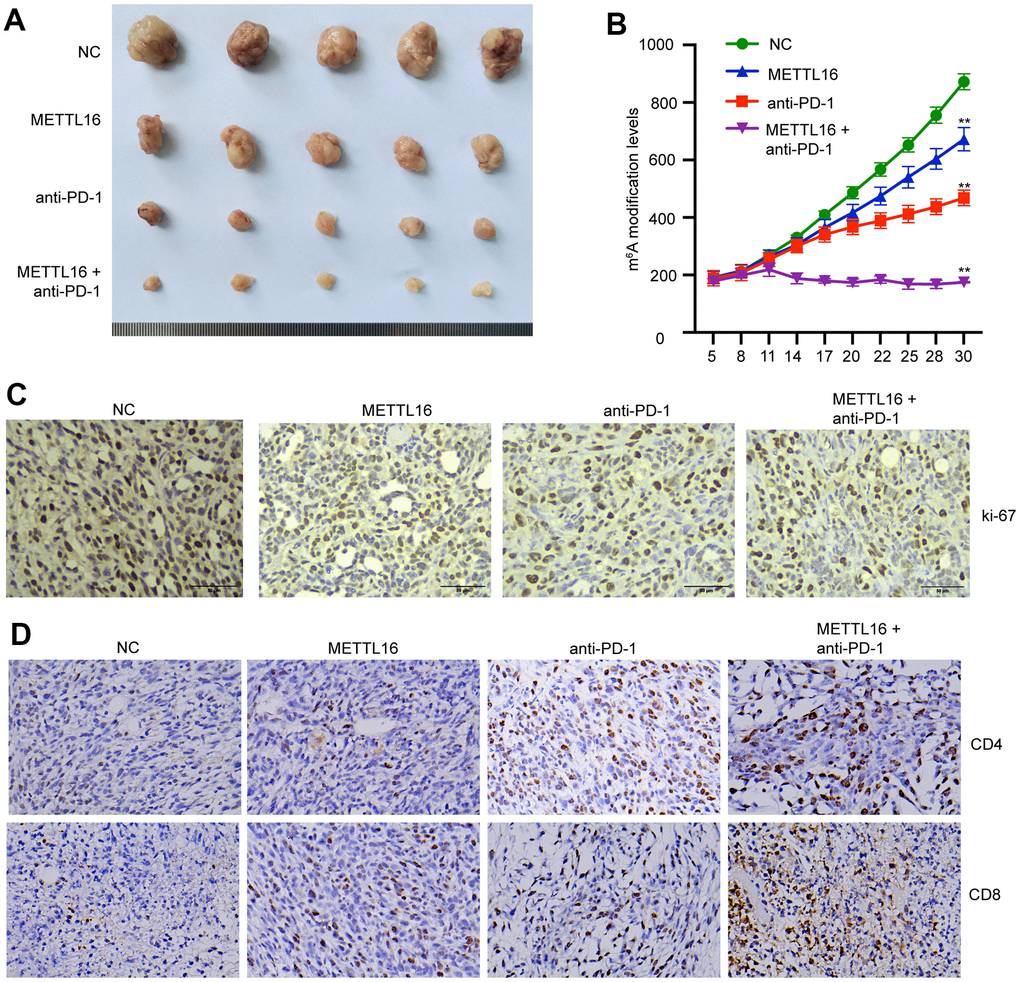
Figure 6. METTL16 modulates CRC cell growth in vivo. The xenograft mouse model was established using control and METTL16 overexpressed SW480 cells and treated with anti-PD-1 antibody. (A) Images of tumors were taken. (B) Tumor growth curve was recorded. (C) The expression of Ki-67 was measured by IHC assay. (D) The expression of CD8 and CD4 in tumor tissues was measured by IHC assay. **p<0.01.
Discussion
The involvement of m6A modification has been reported in various pathophysiological processes, and its aberrant regulation is correlated with the initiation and development of a variety of diseases [27]. The representative m6A methyltransferases and demethylases, such as ALKBH5, METTL3, and METTL14, have been widely studied in cancer development [27]. The correlation between METTL16 with cancers also draws great attention in recent years [28, 29]. The level of METTL16 is upregulated in glioma [30] and is correlated with poor prognosis of melanoma [31]. Wang et al. reported that METTL16 promotes the proliferation of gastric cancer cells via upregulating the expression of cyclin D1 [27]. In this study, we observed decreased level of METTL16 in clinical CRC tumor samples and CRC cell lines, compared with the non-tumor tissues and normal cell line. Further in vitro experiments revealed that overexpression of METTL16 led to suppressed CRC cell growth, migration, and invasion.
It has been demonstrated that METTL16 could function in m6A-dependent or -independent manner [32]. For example, METTL16 mediated the translation of CIDEA in a m6A-dependent manner and promoted non-alcoholic fatty liver disease [33]. Degradation of MAT2A pre-mRNA mediated by METTL16 was disrupted by oxidative stress, which consequently aggravated the apoptosis of nucleus pulposus cells and exacerbated the process of intervertebral disc degeneration [34]. Here, we identified that overexpression of METTL16 decreased the expression of PD-L1 in CRC cells through inducing the m6A modification and degradation of PD-L1 RNA. PD-L1 is the ligand protein produced by tumor cells, which can bind to PD-1 of T lymphocytes [35]. The interaction of PD-L1 with PD-1 consequently inhibits the immune recognition function and the cytotoxicity of T lymphocytes against tumor cells, and therefore facilitates the immune evasion and tumor growth [36, 37]. In this work, we established a CRC cell/T cell co-culture system and revealed that overexpression of METTL16 in CRC cells suppressed the portion of PD-1 positive cells in the activated T cells, implying the decreased immune evasion. The results from in vivo experiments presented synergetic tumor suppressive effects of METTL16 overexpression and PD-1 inhibition. Our findings provided novel regulatory mechanism underlying CRC progression and a potential target for its clinical therapy. Nevertheless, certain limitation exists in current study. For example, further investigation of the response of immune system upon METTL16 alteration is needed.
Conclusions
To conclude, we identified the suppressed METTL16 expression in CRC tumors and demonstrated that METTL16 overexpression suppressed CRC cell proliferation, migration and invasion. Mechanistically, METTL16 induced the m6A modification of PD-L1 to inhibit immune evasion and enhance the therapeutic effects of anti-PD-1 treatment on CRC.
Supplementary Materials
Author Contributions
Wang AL, Sun YJ analyzed clinical data and drafted the manuscript; Wang XC, Yan ZF, Wang DS advised and reviewed the manuscript; Zeng L, Lu QG performed the experiment.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement and Consent
The animal experiments were authorized by Ethics Committee of Tangshan Traditional Chinese Medicine Hospital. The approval for human experiments was received from the Ethics Committee of Tangshan Traditional Chinese Medicine Hospital. All patients have signed the informed consents.
Funding
The experiments of this research mentioned in the paper were funded by Tangshan Traditional Chinese Medicine Hospital.
References
- 1. Liu C, Liu R, Wang B, Lian J, Yao Y, Sun H, Zhang C, Fang L, Guan X, Shi J, Han S, Zhan F, Luo S, et al. Blocking IL-17A enhances tumor response to anti-PD-1 immunotherapy in microsatellite stable colorectal cancer. J Immunother Cancer. 2021; 9:e001895. https://doi.org/10.1136/jitc-2020-001895 [PubMed]
- 2. Bagchi S, Yuan R, Engleman EG. Immune Checkpoint Inhibitors for the Treatment of Cancer: Clinical Impact and Mechanisms of Response and Resistance. Annu Rev Pathol. 2021; 16:223–49. https://doi.org/10.1146/annurev-pathol-042020-042741 [PubMed]
- 3. Doroshow DB, Bhalla S, Beasley MB, Sholl LM, Kerr KM, Gnjatic S, Wistuba II, Rimm DL, Tsao MS, Hirsch FR. PD-L1 as a biomarker of response to immune-checkpoint inhibitors. Nat Rev Clin Oncol. 2021; 18:345–62. https://doi.org/10.1038/s41571-021-00473-5 [PubMed]
- 4. Afolabi BE. Cancer and Immunology-the Homeostasis Dance. J Mod Med Oncol. 2022; 2:5. https://doi.org/10.53964/jmmo.2022005
- 5. Vilar E, Gruber SB. Microsatellite instability in colorectal cancer-the stable evidence. Nat Rev Clin Oncol. 2010; 7:153–62. https://doi.org/10.1038/nrclinonc.2009.237 [PubMed]
- 6. Picard E, Verschoor CP, Ma GW, Pawelec G. Relationships Between Immune Landscapes, Genetic Subtypes and Responses to Immunotherapy in Colorectal Cancer. Front Immunol. 2020; 11:369. https://doi.org/10.3389/fimmu.2020.00369 [PubMed]
- 7. Lin A, Zhang J, Luo P. Crosstalk Between the MSI Status and Tumor Microenvironment in Colorectal Cancer. Front Immunol. 2020; 11:2039. https://doi.org/10.3389/fimmu.2020.02039 [PubMed]
- 8. Sharma BK. Innovative Translation Strategies in the Emergence of Next-Generation Drugs in Precision Cancer Medicine. J Mod Med Oncol. 2022; 2:4. https://doi.org/10.53964/jmmo.2022004
- 9. Müller MF, Ibrahim AE, Arends MJ. Molecular pathological classification of colorectal cancer. Virchows Arch. 2016; 469:125–34. https://doi.org/10.1007/s00428-016-1956-3 [PubMed]
- 10. Parikh AR, Szabolcs A, Allen JN, Clark JW, Wo JY, Raabe M, Thel H, Hoyos D, Mehta A, Arshad S, Lieb DJ, Drapek LC, Blaszkowsky LS, et al. Radiation therapy enhances immunotherapy response in microsatellite stable colorectal and pancreatic adenocarcinoma in a phase II trial. Nat Cancer. 2021; 2:1124–35. https://doi.org/10.1038/s43018-021-00269-7 [PubMed]
- 11. Sharma BK. Prospective Paradigms in Therapeutic Modalities in the Management of Human Cutaneous Melanoma: Plausible Role of Bioactive Compounds. J Mod Med Oncol. 2022; 2:1. https://doi.org/10.53964/jmmo.2022001
- 12. Fukuoka S, Hara H, Takahashi N, Kojima T, Kawazoe A, Asayama M, Yoshii T, Kotani D, Tamura H, Mikamoto Y, Hirano N, Wakabayashi M, Nomura S, et al. Regorafenib Plus Nivolumab in Patients With Advanced Gastric or Colorectal Cancer: An Open-Label, Dose-Escalation, and Dose-Expansion Phase Ib Trial (REGONIVO, EPOC1603). J Clin Oncol. 2020; 38:2053–61. https://doi.org/10.1200/JCO.19.03296 [PubMed]
- 13. Srikrishna D. Does Physical Activity Play a Role in the Efficacy of RegoNivo for MSS-mCRC? Clin Colorectal Cancer. 2022; 21:e76–7. https://doi.org/10.1016/j.clcc.2021.09.011 [PubMed]
- 14. Li Y, Chen X, Lu C. The interplay between DNA and histone methylation: molecular mechanisms and disease implications. EMBO Rep. 2021; 22:e51803. https://doi.org/10.15252/embr.202051803 [PubMed]
- 15. Tessarz P, Kouzarides T. Histone core modifications regulating nucleosome structure and dynamics. Nat Rev Mol Cell Biol. 2014; 15:703–8. https://doi.org/10.1038/nrm3890 [PubMed]
- 16. Hogg SJ, Beavis PA, Dawson MA, Johnstone RW. Targeting the epigenetic regulation of antitumour immunity. Nat Rev Drug Discov. 2020; 19:776–800. https://doi.org/10.1038/s41573-020-0077-5 [PubMed]
- 17. Ilango S, Paital B, Jayachandran P, Padma PR, Nirmaladevi R. Epigenetic alterations in cancer. Front Biosci (Landmark Ed). 2020; 25:1058–109. https://doi.org/10.2741/4847 [PubMed]
- 18. Sun L, Zhang H, Gao P. Metabolic reprogramming and epigenetic modifications on the path to cancer. Protein Cell. 2022; 13:877–919. https://doi.org/10.1007/s13238-021-00846-7 [PubMed]
- 19. Koshariya M, Agarwal B, Rathod R. Overview of Advancement and Management of Gastrointestinal Stromal Tumors. J Mod Med Oncol. 2021; 2:3. https://doi.org/10.53964/jmmo.2022003
- 20. Pan T. N6-methyl-adenosine modification in messenger and long non-coding RNA. Trends Biochem Sci. 2013; 38:204–9. https://doi.org/10.1016/j.tibs.2012.12.006 [PubMed]
- 21. Niu Y, Zhao X, Wu YS, Li MM, Wang XJ, Yang YG. N6-methyl-adenosine (m6A) in RNA: an old modification with a novel epigenetic function. Genomics Proteomics Bioinformatics. 2013; 11:8–17. https://doi.org/10.1016/j.gpb.2012.12.002 [PubMed]
- 22. Yang B, Wang JQ, Tan Y, Yuan R, Chen ZS, Zou C. RNA methylation and cancer treatment. Pharmacol Res. 2021; 174:105937. https://doi.org/10.1016/j.phrs.2021.105937 [PubMed]
- 23. Mendel M, Chen KM, Homolka D, Gos P, Pandey RR, McCarthy AA, Pillai RS. Methylation of Structured RNA by the m6A Writer METTL16 Is Essential for Mouse Embryonic Development. Mol Cell. 2018; 71:986–1000.e11. https://doi.org/10.1016/j.molcel.2018.08.004 [PubMed]
- 24. Perez-Pepe M, Alarcón CR. An RNA link for METTL16 and DNA repair in PDAC. Nat Cancer. 2022; 3:1018–20. https://doi.org/10.1038/s43018-022-00434-6 [PubMed]
- 25. Dou R, Liu K, Yang C, Zheng J, Shi D, Lin X, Wei C, Zhang C, Fang Y, Huang S, Song J, Wang S, Xiong B. EMT-cancer cells-derived exosomal miR-27b-3p promotes circulating tumour cells-mediated metastasis by modulating vascular permeability in colorectal cancer. Clin Transl Med. 2021; 11:e595. https://doi.org/10.1002/ctm2.595 [PubMed]
- 26. Liu X, He H, Zhang F, Hu X, Bi F, Li K, Yu H, Zhao Y, Teng X, Li J, Wang L, Zhang Y, Wu Q. m6A methylated EphA2 and VEGFA through IGF2BP2/3 regulation promotes vasculogenic mimicry in colorectal cancer via PI3K/AKT and ERK1/2 signaling. Cell Death Dis. 2022; 13:483. https://doi.org/10.1038/s41419-022-04950-2 [PubMed]
- 27. Wang XK, Zhang YW, Wang CM, Li B, Zhang TZ, Zhou WJ, Cheng LJ, Huo MY, Zhang CH, He YL. METTL16 promotes cell proliferation by up-regulating cyclin D1 expression in gastric cancer. J Cell Mol Med. 2021; 25:6602–17. https://doi.org/10.1111/jcmm.16664 [PubMed]
- 28. Zeng X, Zhao F, Cui G, Zhang Y, Deshpande RA, Chen Y, Deng M, Kloeber JA, Shi Y, Zhou Q, Zhang C, Hou J, Kim W, et al. METTL16 antagonizes MRE11-mediated DNA end resection and confers synthetic lethality to PARP inhibition in pancreatic ductal adenocarcinoma. Nat Cancer. 2022; 3:1088–104. https://doi.org/10.1038/s43018-022-00429-3 [PubMed]
- 29. Dai YZ, Liu YD, Li J, Chen MT, Huang M, Wang F, Yang QS, Yuan JH, Sun SH. METTL16 promotes hepatocellular carcinoma progression through downregulating RAB11B-AS1 in an m6A-dependent manner. Cell Mol Biol Lett. 2022; 27:41. https://doi.org/10.1186/s11658-022-00342-8 [PubMed]
- 30. Cong P, Wu T, Huang X, Liang H, Gao X, Tian L, Li W, Chen A, Wan H, He M, Dai D, Li Z, Xiong L. Identification of the Role and Clinical Prognostic Value of Target Genes of m6A RNA Methylation Regulators in Glioma. Front Cell Dev Biol. 2021; 9:709022. https://doi.org/10.3389/fcell.2021.709022 [PubMed]
- 31. Liu J, Zhou Z, Ma L, Li C, Lin Y, Yu T, Wei JF, Zhu L, Yao G. Effects of RNA methylation N6-methyladenosine regulators on malignant progression and prognosis of melanoma. Cancer Cell Int. 2021; 21:453. https://doi.org/10.1186/s12935-021-02163-9 [PubMed]
- 32. Pendleton KE, Chen B, Liu K, Hunter OV, Xie Y, Tu BP, Conrad NK. The U6 snRNA m6A Methyltransferase METTL16 Regulates SAM Synthetase Intron Retention. Cell. 2017; 169:824–35.e14. https://doi.org/10.1016/j.cell.2017.05.003 [PubMed]
- 33. Tang J, Zhao X, Wei W, Liu W, Fan H, Liu XP, Li Y, Wang L, Guo J. METTL16-mediated translation of CIDEA promotes non-alcoholic fatty liver disease progression via m6A-dependent manner. PeerJ. 2022; 10:e14379. https://doi.org/10.7717/peerj.14379 [PubMed]
- 34. Chen PB, Shi GX, Liu T, Li B, Jiang SD, Zheng XF, Jiang LS. Oxidative Stress Aggravates Apoptosis of Nucleus Pulposus Cells through m6A Modification of MAT2A Pre-mRNA by METTL16. Oxid Med Cell Longev. 2022; 2022:4036274. https://doi.org/10.1155/2022/4036274 [PubMed]
- 35. Kim W, Chu TH, Nienhüser H, Jiang Z, Del Portillo A, Remotti HE, White RA, Hayakawa Y, Tomita H, Fox JG, Drake CG, Wang TC. PD-1 Signaling Promotes Tumor-Infiltrating Myeloid-Derived Suppressor Cells and Gastric Tumorigenesis in Mice. Gastroenterology. 2021; 160:781–96. https://doi.org/10.1053/j.gastro.2020.10.036 [PubMed]
- 36. Shen X, Zhao B. Efficacy of PD-1 or PD-L1 inhibitors and PD-L1 expression status in cancer: meta-analysis. BMJ. 2018; 362:k3529. https://doi.org/10.1136/bmj.k3529 [PubMed]
- 37. Wu Q, Jiang L, Li SC, He QJ, Yang B, Cao J. Small molecule inhibitors targeting the PD-1/PD-L1 signaling pathway. Acta Pharmacol Sin. 2021; 42:1–9. https://doi.org/10.1038/s41401-020-0366-x [PubMed]