



Introduction
Worldwide, hepatocellular carcinoma (HCC) is one of the major diseases threatening the health of humans [1–3]. Although the diagnosis and treatment of it are becoming more perfect, the prognosis of HCC patients has not been significantly improved [4]. Reliable biomarkers are not only helpful for early diagnosis and prognosis of cancer patients, but also can reduce medical costs [5–7]. Thus, elucidating the specific biomarkers in the process of HCC development has a critical research significance for early diagnosis and personalized medicine in patients.
Coiled-coil domain-containing protein 50 (CCDC50) was first identified as mapping to chromosome 3q28 [8]. As a negative regulator of IFN signalling, it is ubiquitously expressed in human tissues [9]. It has been reported that CCDC50 was enhanced by viral infection, and could inhibit the NF-κB-mediated apoptotic pathway, enhance the viral resistance, and regulate the p53 signalling pathways [8, 10, 11]. Further, CCDC50 could regulate Ras signalling pathway and promote mice HCC [12]. However, the potential role of CCDC50 in human HCC progression remains unclear. Our study aimed to examine the relationships between CCDC50 expression and diverse features in HCC. Furthermore, the CCK8 and transwell assays were employed to determine the biological role of CCDC50 in HCC progression.
Materials and Methods
Analysis of CCDC50 expression in pan-cancer
GEPIA (http://gepia.cancer-pku.cn/) is a web-based tool that quickly outputs customizable data results, and can be interactively used for analysing gene expression data of the TCGA clinical data and RNA-seq [13]. We analysed the CCDC50 expression across TCGA tumors, and the GTEx data and matched TCGA normal were contained as controls. UALCAN database is a powerful web-portal [14], herein, we detached CCDC50 protein expression via UALCAN web-portal.
The prognostic and clinical information of CCDC50 in HCC
We used the GEPIA [13] and PrognoScan [15] to examine the prognostic OS, and DSS of CCDC50 in HCC. The gene mutation features of CCDC50 in HCC were analysed via the cBioPortal [16]. We chose the three datasets: “MSK, Clin Cancer Res 2018 [17], MERiC/Basel, Nat Commun. 2022 [18], and TCGA. We explored the TIMER2.0 to estimate the immunological roles of CCDC50 in HCC [19].
Cell culture, siRNA and qRT-PCR
LIHC cell lines and LO2 cells lines were obtained from ATCC and cultured in RPMI-1640 medium supplemented with 10% FBS. CCDC50 siRNA kits (si-CCDC50#1: 5’- CCGUGCUUAUGCAGAUAGUTT-3’; si-CCDC50#2: 5’- GCAGCAAAUUCCAAGUCAATT-3’) and negative control siRNAs (si-NC: 5’-UUCUCCGAACGUGUC ACG UTT-3’) were purchased from GenePharma. Transfections were performed as previously described [20]. The qRT-PCR assay was conducted as previously documented [20]. The following primer sequences were used in this study: CCDC50: 5’-GACGACGCATTCAGGAGAAGA-3’, 5’-ACTATCTGCATAAGCACGGGTT-3’; β-actin: 5’-GTCTTCCCCTCCATCGTG-3’’, 5’-AGGGTGAGGATGCCTCTCTT-3’.
Western blot
Western blot assays were fulfilled as described previously [20]. The following primary antibodies were used in this finding: CCDC50 (A17836; Abclonal) and β-actin (sc-47778; Santa Cruz). All the assays were independently repeated thrice.
Cell proliferation assay
CCK8 assays were fulfilled as described previously [20]. For CCK8 assay, Hep3B and Huh7 cells were cultured in a 96-well plate supplemented with 200 μL of RPMI-1640 medium.
Correlation between CCDC50 and cancer drug sensitivity
GSCA [21] is an integrated platform combining clinical and small molecular drugs information, it can help us easily analyse CCDC50 expression with drug sensitivity GDSC and CTRP [22, 23].
Data availability statement
The data from this article can be obtained from the public database The Cancer Genome Atlas (https://portal.gdc.cancer.gov/).
Results
CCDC50 was differentially expressed in multiple cancer including HCC
Firstly, we combined the TCGA and GTEx databases, and confirmed that the expression of CCDC50 was significantly lower in 14 cancer tissues than normal tissues. In addition, CCDC50 was up-regulated in 12 types of cancer including HCC (Figure 1A). As the main undertaker of life activities, the change of protein expression level is directly related to cancer progression [24]. To determine the protein levels of CCDC50 in different types of cancer, we analysed the UALCAN database and found that the protein expression of CCDC50 was low in 4 types of cancer, whereas it was high in 5 types of cancer including HCC (Figure 1B).
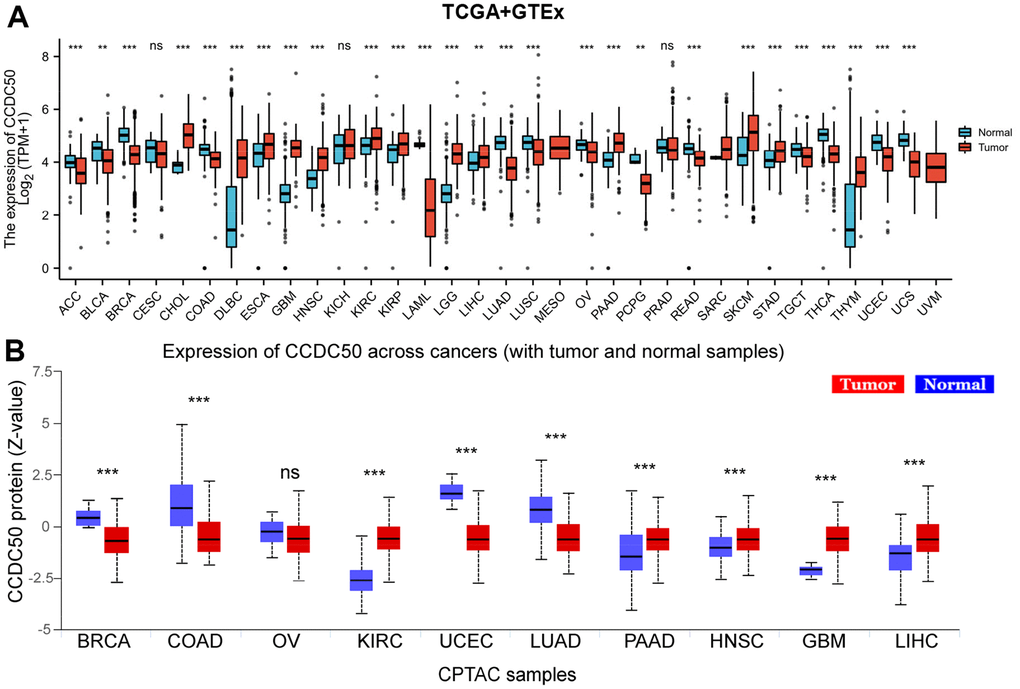
Figure 1. CCDC50 mRNA and protein expressions between tumour and normal tissues. (A) The CCDC50 expression in pan-cancer analysis via the TCGA/GTEx databases. (B) The protein of CCDC50 in pan-cancer analysis from CPTAC samples via the UALCAN web-portal. Z-values represent standard deviations from the median across samples for the given cancer type. ns, p > 0.05; *p < 0.05; **p < 0.01; ***p < 0.001.
To explore CCDC50 expression levels in HCC, we used HPA and GEO datasets to validate its expression in HCC. We showed that CCDC50 was upregulated in HCC (Figure 2A, 2B). Furthermore, we showed that CCDC50 was correlated with adverse clinical features, such as grade, pathological stage and OS event, not related to NM stage (Figure 2C–2G). GEO datasets also confirmed that high CCDC50 expression had worse OS in HCC cancer patients (Figure 2H, 2I).
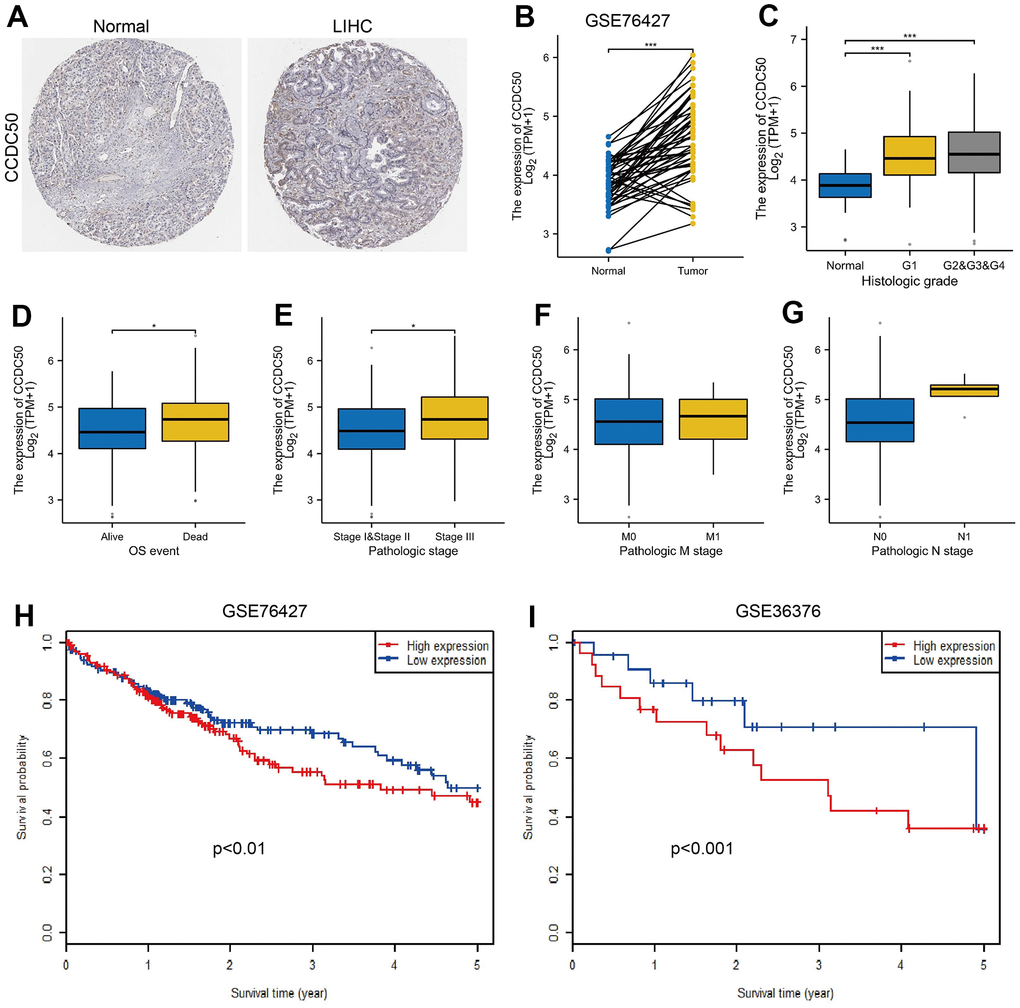
Figure 2. CCDC50 was highly expressed in HCC. (A) CCDC50 protein was highly expressed in HCC tissues than normal liver tissues examined by HPA database. (B) Validation of the expression of CCDC50 in HCC by GEO dataset. (C–G) Relationships between the expression of CCDC50 and diverse clinical features in HCC. (H, I) Validation of the prognosis of CCDC50 in HCC by GEO dataset.
Prognosis value of CCDC50 in HCC
As CCDC50 expression was up-regulated in HCC tissues, we explored the prognostic value of CCDC50 in HCC. The KM survival curve results showed that increased expression of CCDC50 related to poor OS and DSS in HCC (Figure 3A, 3B). We comprehensively analysed the relationship between CCDC50 expression and OS in HCC. High CCDC50 expression had worse OS in diverse subgroups of HCC, including residual tumour (Figure 3C), gender (Figure 3D), age (Figure 3E), race (Figure 3F), histologic grade (Figure 3G), weight (Figure 2H), TNM stage (Figure 3H–3K), and tumour status (Figure 3L).
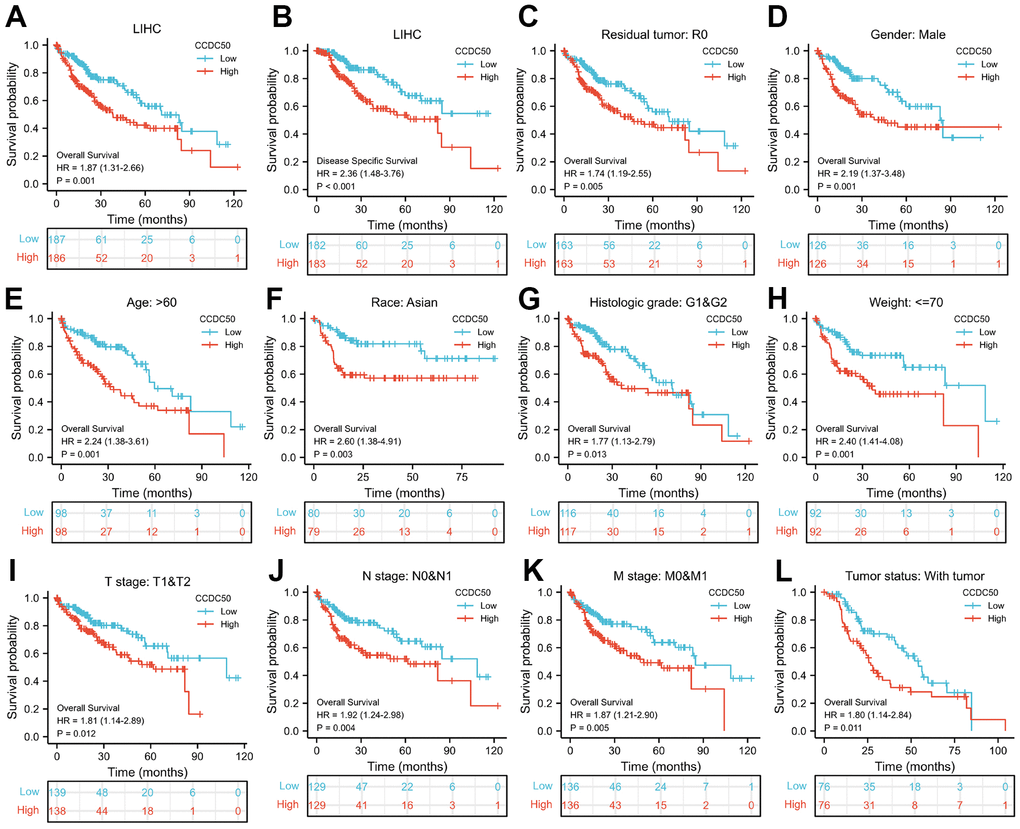
Figure 3. Prognostic analysis of CCDC50 in HCC. (A, B) The correlation between CCDC50 and OS, and DSS. (C–L) The correlation between CCDC50 and OS in different clinical subgroups of HCC, including residual tumour, gender, age, race, histologic grade, weight, TNM stage, AND tumour status.
To examine whether CCDC50 can be used in HCC prognosis, we constructed a nomogram to predict the OS, and DSS in HCC patients and found that pathological stage and CCDC50 expression act as prognostic factors (Figure 4A, 4B and Tables 1–3). The calibration curves indicated that the nomogram can reliably predict the 1-, 3-, and 5-year OS, and DSS in HCC (Figure 4C, 4D). In summary, the model constructed above can optimize the prediction of different survival rates for liver cancer patients, and has high performance in prediction.
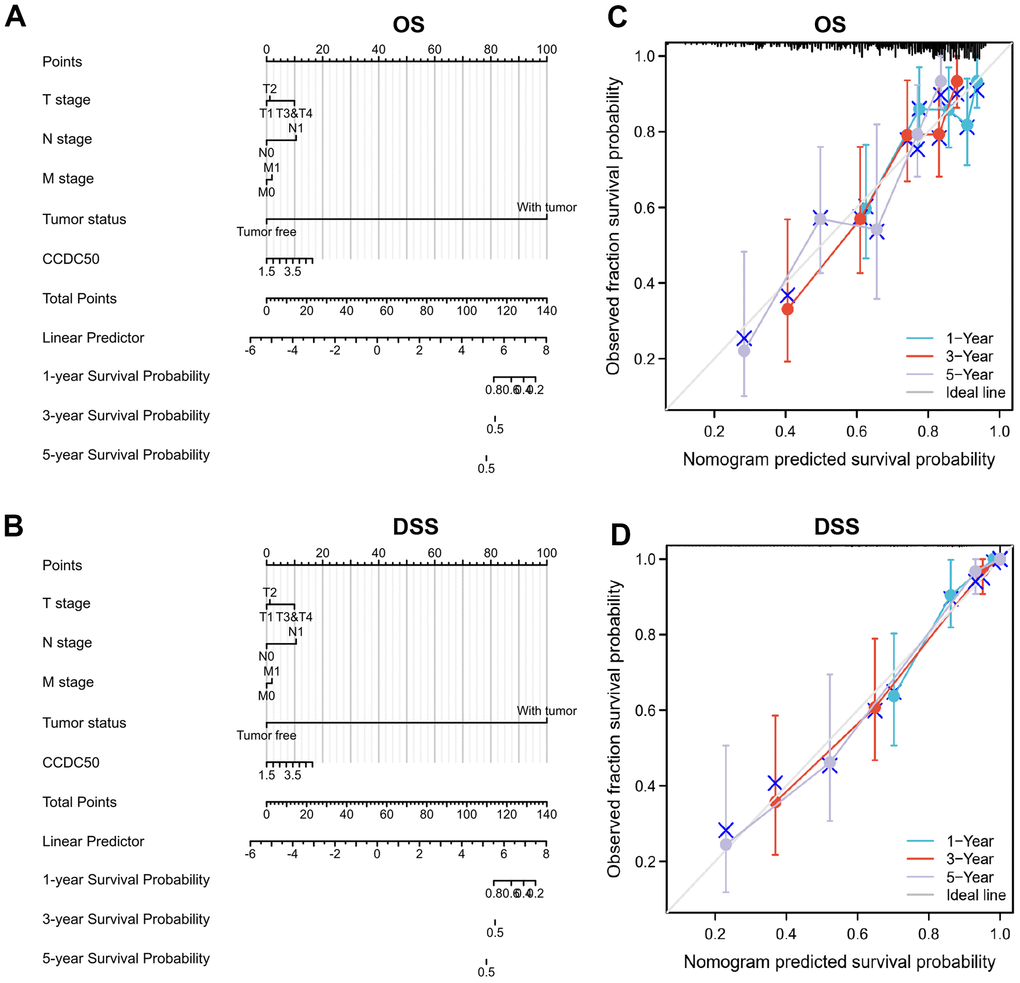
Figure 4. Nomogram and calibration curve for predicting the probability of 1-, 3-, and 5-year OS, and DSS in HCC patients. (A, B) A nomogram integrates CCDC50 and other prognostic factors in HCC from the TCGA data. (C, D) The calibration curve of the nomogram.
Table 1. Univariate and multivariate Cox regression analyses of different parameters of overall survival in liver hepatocellular carcinoma.
| Characteristics | Total (N) | Univariate analysis | Multivariate analysis | |||
| Hazard ratio (95% CI) | P-value | Hazard ratio (95% CI) | P-value | |||
| T stage | 370 | |||||
| T1 | 183 | |||||
| T2 | 94 | 1.428 (0.901–2.264) | 0.129 | 1.594 (0.874–2.907) | 0.128 | |
| T3 and T4 | 93 | 2.949 (1.982–4.386) | <0.001 | 1.694 (0.226–12.699) | 0.608 | |
| N stage | 258 | |||||
| N0 | 254 | |||||
| N1 | 4 | 2.029 (0.497–8.281) | 0.324 | |||
| M stage | 272 | |||||
| M0 | 268 | |||||
| M1 | 4 | 4.077 (1.281–12.973) | 0.017 | 1.304 (0.310–5.480) | 0.717 | |
| Pathologic stage | 349 | |||||
| Stage I and Stage II | 259 | |||||
| Stage III and Stage IV | 90 | 2.504 (1.727–3.631) | <0.001 | 1.727 (0.237–12.611) | 0.590 | |
| Tumour status | 354 | |||||
| Tumour free | 202 | |||||
| With tumour | 152 | 2.317 (1.590–3.376) | <0.001 | 1.953 (1.226–3.110) | 0.005 | |
| CCDC50 | 373 | 4.512 (0.270–75.472) | 0.295 | |||
Table 2. Univariate and multivariate Cox regression analyses of different parameters of disease-specific survival in liver hepatocellular carcinoma.
| Characteristics | Total (N) | Univariate analysis | Multivariate analysis | |||
| Hazard ratio (95% CI) | P-value | Hazard ratio (95% CI) | P-value | |||
| T stage | 362 | |||||
| T1 | 180 | |||||
| T2 | 92 | 1.619 (0.869–3.016) | 0.129 | 1.459 (0.608–3.498) | 0.397 | |
| T3 and T4 | 90 | 4.328 (2.583–7.251) | <0.001 | 14.188 (0.791–254.370) | 0.072 | |
| N stage | 253 | |||||
| N0 | 249 | |||||
| N1 | 4 | 3.612 (0.870–14.991) | 0.077 | 9.081 (1.141–72.298) | 0.037 | |
| M stage | 268 | |||||
| M0 | 265 | |||||
| M1 | 3 | 5.166 (1.246–21.430) | 0.024 | 1.997 (0.466–8.557) | 0.352 | |
| Pathologic stage | 341 | |||||
| Stage I and Stage II | 254 | |||||
| Stage III and Stage IV | 87 | 3.803 (2.342–6.176) | <0.001 | 0.332 (0.018–6.028) | 0.456 | |
| Tumour status | 354 | |||||
| Tumour free | 202 | |||||
| With tumour | 152 | 775790759.389 (0.000–Inf) | 0.994 | |||
| CCDC50 | 365 | 0.073 (0.000–31.328) | 0.397 | |||
Table 3. Univariate and multivariate Cox regression analyses of different parameters of progression-free interval in liver hepatocellular carcinoma.
| Characteristics | Total (N) | Univariate analysis | Multivariate analysis | |||
| Hazard ratio (95% CI) | P-value | Hazard ratio (95% CI) | P-value | |||
| T stage | 370 | |||||
| T1 | 183 | |||||
| T2 | 94 | 2.017 (1.409–2.888) | <0.001 | 1.100 (0.694–1.742) | 0.686 | |
| T3 and T4 | 93 | 2.798 (1.969–3.975) | <0.001 | 0.895 (0.212–3.785) | 0.880 | |
| N stage | 258 | |||||
| N0 | 254 | |||||
| N1 | 4 | 1.370 (0.338–5.552) | 0.659 | |||
| M stage | 272 | |||||
| M0 | 268 | |||||
| M1 | 4 | 3.476 (1.091–11.076) | 0.035 | 1.462 (0.447–4.782) | 0.530 | |
| Pathologic stage | 349 | |||||
| Stage I and Stage II | 259 | |||||
| Stage III and Stage IV | 90 | 2.201 (1.591–3.046) | <0.001 | 2.043 (0.488–8.555) | 0.328 | |
| Tumour status | 354 | |||||
| Tumour free | 202 | |||||
| With tumour | 152 | 11.342 (7.567–17.000) | <0.001 | 15.140 (9.082–25.237) | <0.001 | |
| CCDC50 | 373 | 2.303 (0.186–28.565) | 0.516 | |||
Gene mutation landscape of CCDC50 in HCC
The mutational data of CCDC50 from the cBioPortal that showed genetic alterations in the CCDC50 gene were observed in only 1.4% of the HCC patients (Figure 5A), and 1 missense site was found between amino acids ‘0’ and ‘306’ (Figure 5B). These results indicate that genetic alterations of CCDC50 may not be the main factor affecting its prognostic ability.
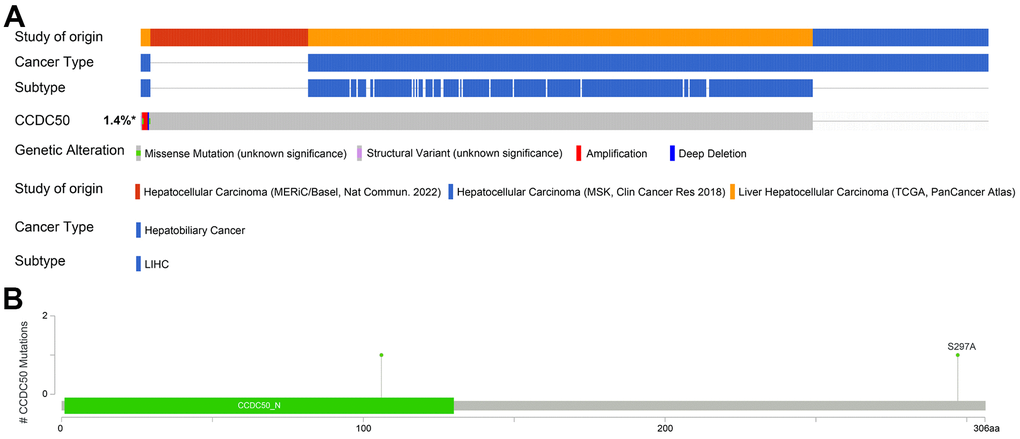
Figure 5. Mutational analysis of CCDC50 in HCC. (A) OncoPrint visual summary of the alterations in the CCDC50 gene. (B) The hot spots of mutations in CCDC50.
DNA methylation modulates CCDC50 expression in HCC
DNA methylation and m6A methylation play crucial roles in cancer progression. Therefore, it is crucial to reveal whether CCDC50 is regulated by DNA methylation and then abnormally expressed in hepatocellular carcinoma. First, we confirmed that CCDC50 was positively correlated with m6A regulatory factors expression in HCC (Figure 6A). Furthermore, we found that the level of DNA methylation of CCDC50 was downregulated in liver cancer tissue and was negatively correlated with liver cancer metastasis (Figure 6B, 6C). Finally, we found that the level of DNA methylation of CCDC50 was negatively related to the expression of CCDC50 in HCC (Figure 6D). However, the level of DNA methylation of CCDC50 did not affect the prognosis of HCC patients in TCGA LIHC datasets (Figure 6E, 6F). Interestingly, the level of DNA methylation of CCDC50 was positively correlated with the Tr1 and iTreg cell infiltration level (Figure 6G). These results partially suggest that DNA methylation may affect the infiltration levels of diverse immune cells by regulating the expression of CCDC50 in HCC.
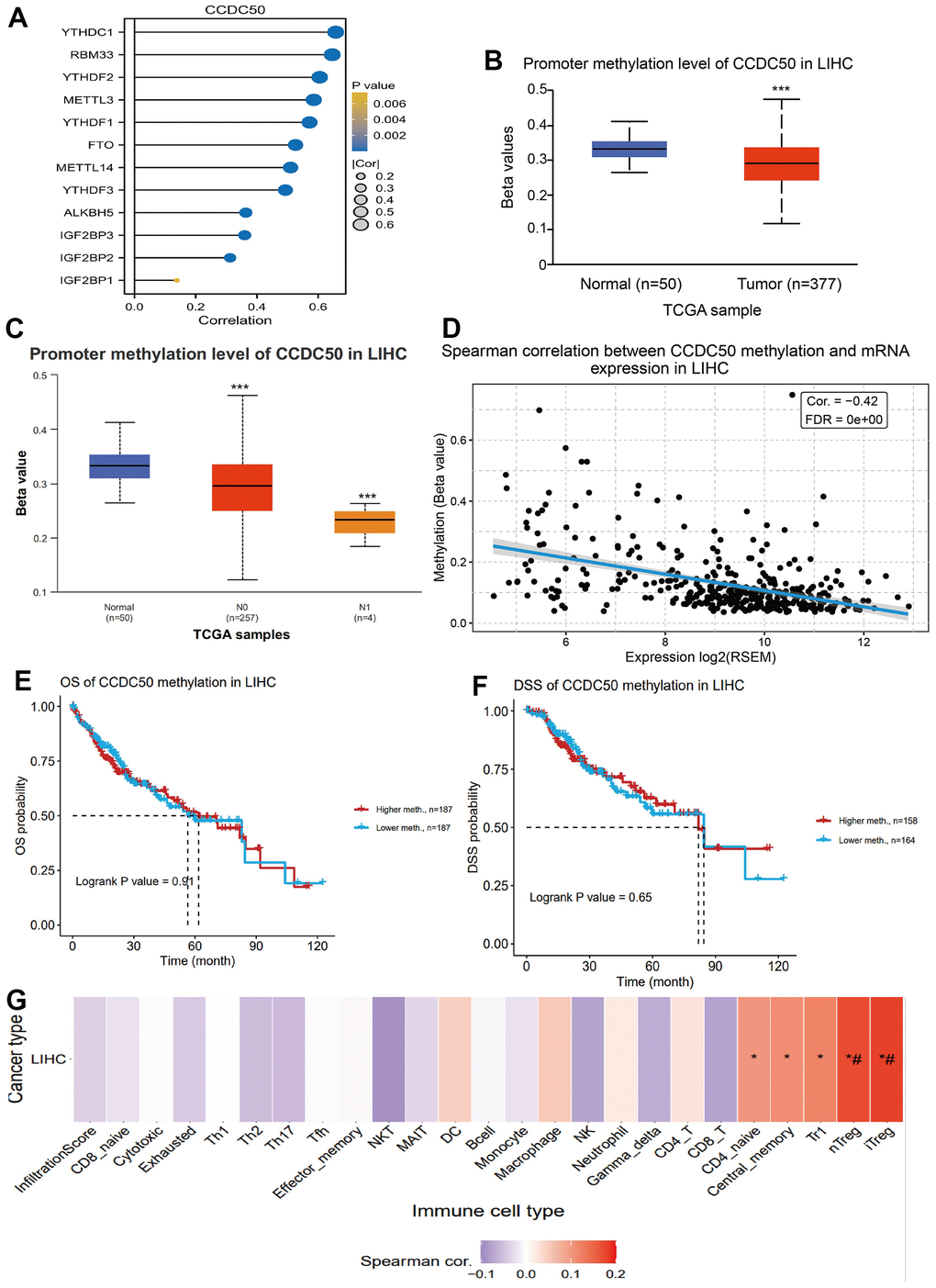
Figure 6. DNA and RNA methylation analysis of CCDC50 in HCC. (A) Correlation between m6A-related modulator genes and CCDC50 in LIHC. (B, C) The level of DNA methylation of CCDC50 in liver cancer tissue was significantly lower than that in normal liver tissue, and was negatively correlated with liver cancer metastasis. (D) The level of DNA methylation of CCDC50 was significantly negatively correlated with the expression of CCDC50 in HCC. (E, F) Correlation between DNA methylation of CCDC50 and prognosis in LIHC. (G) Correlation between DNA methylation of CCDC50 and the level of immune cell infiltration in LIHC. RSEM is a software for quantifying gene expression based on STAR sequence comparison. ns, p > 0.05; *p < 0.05; **p < 0.01; ***p < 0.001.
Biological functions of CCDC50 in LIHC
Using the “clusterProfiler” R package, we performed functional annotations of CCDC50-associated differentially expressed genes (DEGs) in HCC patients, and 423 DEGs (mRNA and lncRNA) were captured, including 331 upregulated and 92 downregulated genes (Figure 7A–7D). The GO enrichment results showed that the DEGs were mainly involved in the extracellular space, acute-phase response, cell differentiation, chemoattractant activity, and epidermal growth factor receptor interactions (Figure 7E). KEGG results showed that the DEGs were mainly involved in the gastric cancer, pancreatic secretion, gastric acid secretion, pertussis, glycosaminoglycan biosynthesis-keratan sulphate, axon guidance, phenylalanine metabolism, nitrogen metabolism and protein digestion and absorption (Figure 7F).
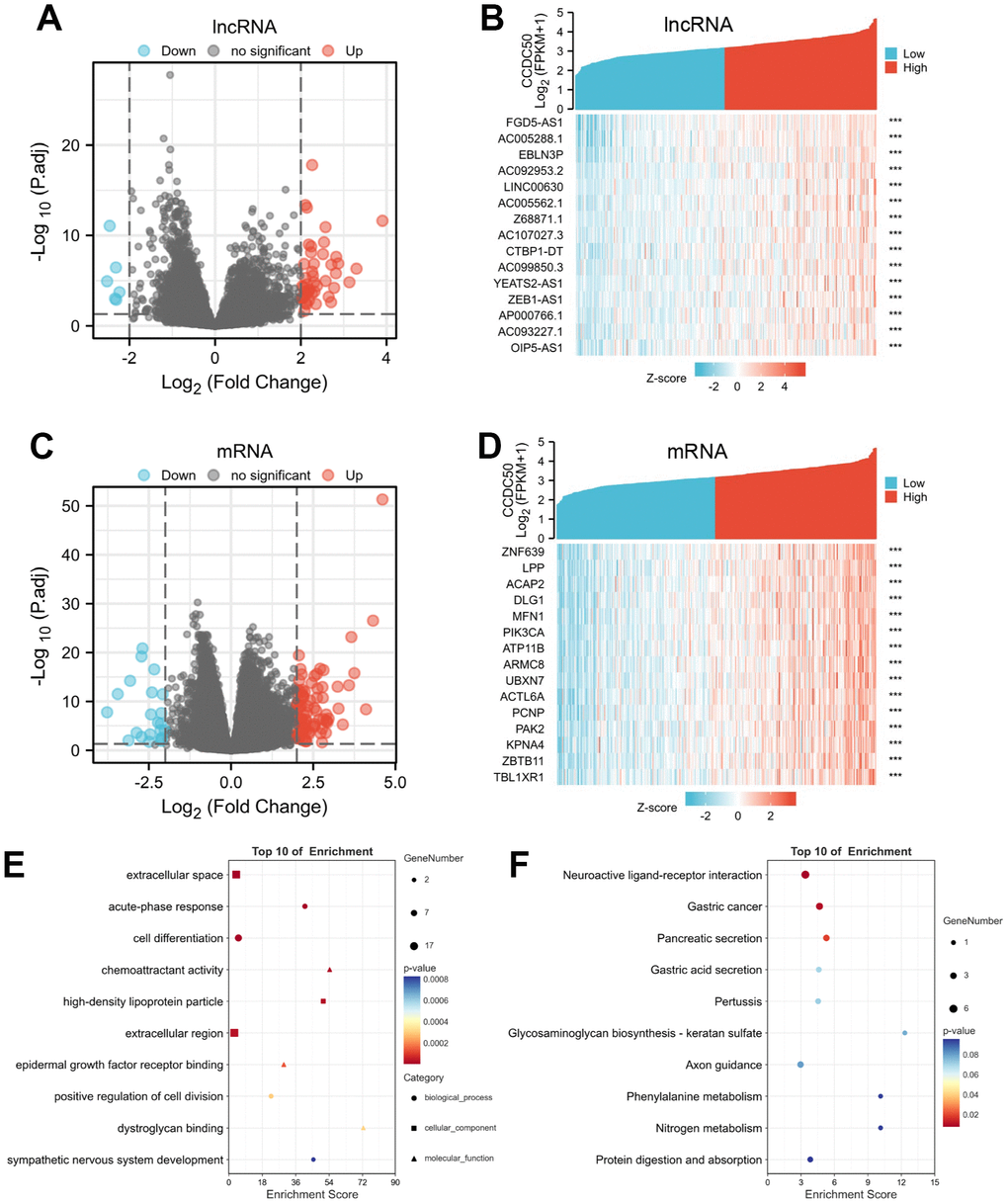
Figure 7. Identifying differentially expressed genes (DEGs) between high and low expression of CCDC50 groups. (A) The volcano plot of differential lncRNA profiles between CCDC50 high expression and CCDC50 low expression. (B) The heat map of the top 15 DEGs (lncRNA) between CCDC50 high expression and CCDC50 low expression. (C) The volcano plot of differential mRNA profiles between CCDC50 high expression and CCDC50 low expression. (D) The heat map of the top 15 DEGs (mRNA) between CCDC50 high expression and CCDC50 low expression. (E) The gene ontology term of CCDC50 analysis by using DEGs. (F) The KEGG term of CCDC50 analysis by using DEGs. ***p < 0.001.
GSEA results suggested that CCDC50 mainly participated in TOLL like receptor, T cell receptor, MAPK and wnt signaling pathway, chemokine signaling pathway (Figure 8A–8D). These results confirmed that CCDC50 regulated immune-related pathways and participate in the malignant progression of liver cancer.
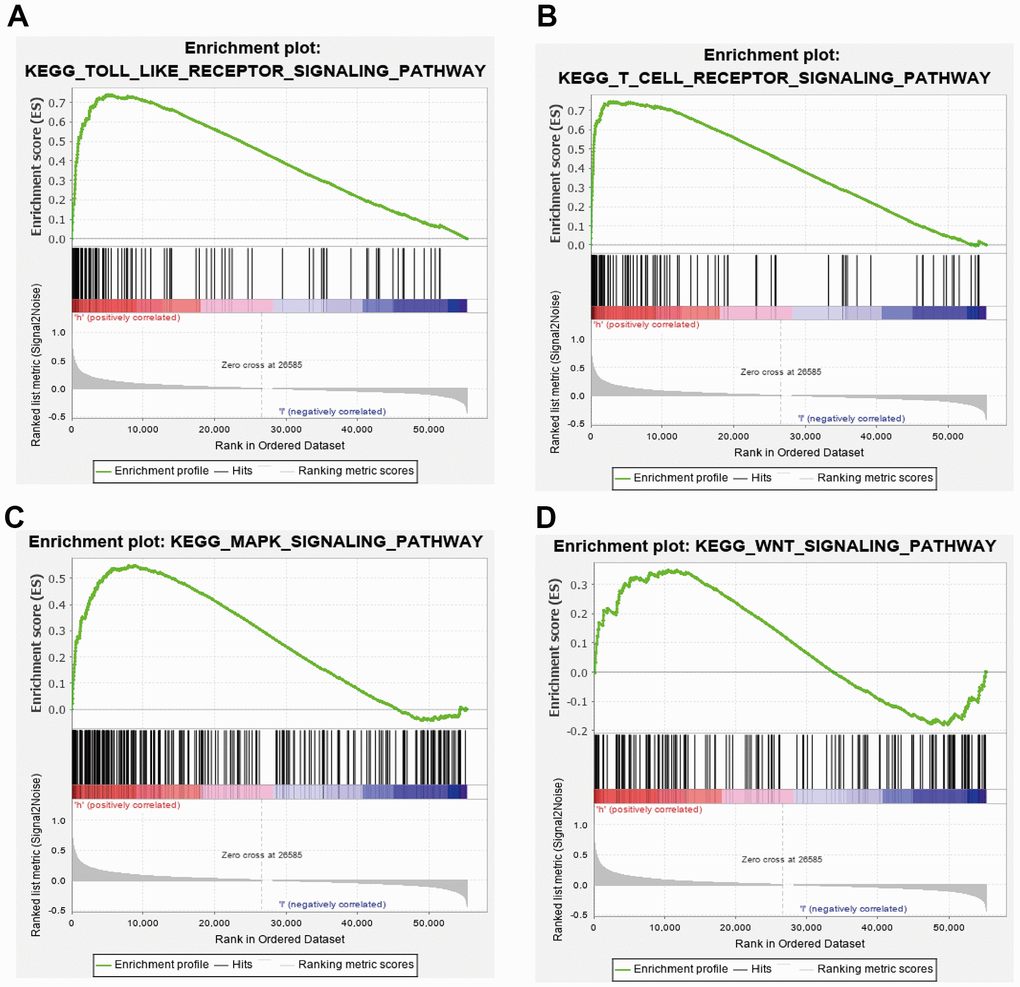
Figure 8. GSEA of CCDC50 in HCC. (A–D) The top GSEA results of CCDC50 in HCC.
Immune cell infiltration of CCDC50 in HCC
Immune cells have a critical role in cancer progression [25]. We used the TIMER database to reveal the relationship between CCDC50 expression and the infiltration levels of 24 immune cell types in HCC. The expression of CCDC50 significantly correlated with these 8 major immune cells in HCC (Figure 9A). CCDC50 expression levels showed a negative correlation with pDC (Figure 9B), Cytotoxic cells (Figure 9C), DC (Figure 9D), and Tgd (Figure 9E), and positive association with Eosinophils (Figure 9F), Tcm (Figure 9G), Th2 cells (Figure 7H), and T helper cells (Figure 9I). Due to the heterogeneity of tumor cells and immune cells, the expression of CCDC50 in hepatocellular carcinoma may be significantly different from the correlation between different immune cells. It is this difference that leads to tumor immune escape or the formation of tumor immune tolerance.
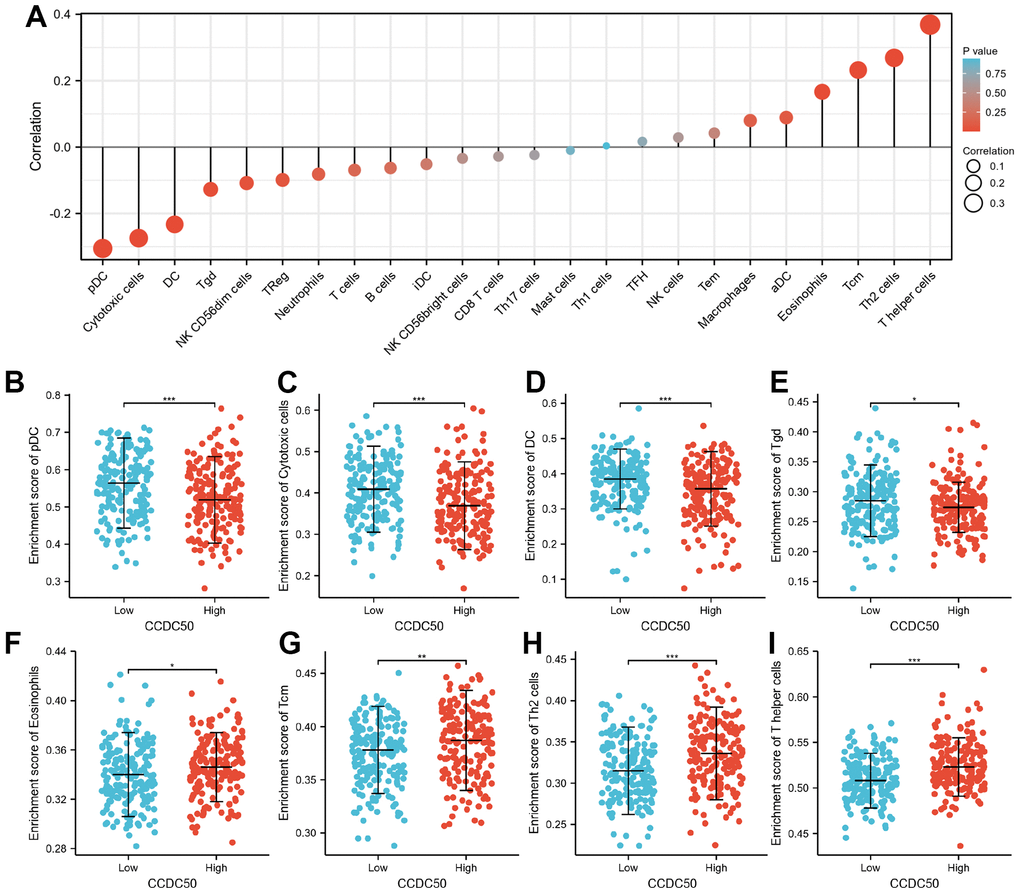
Figure 9. The correlation between CCDC50 expression and the level of immune cell infiltration. (A) The correlation between CCDC50 expression and the level of immune cell infiltration in HCC by using the TIMER database. (B–I) The correlations between CCDC50 expression and the level of pDC, Cytotoxic cells, DC, Tgd, Eosinophils, Tcm, Th2, and T helper cells. *p < 0.05, **p < 0.01, ***p < 0.001.
Knockdown of CCDC50 suppresses the malignant phenotype of LIHC
In vitro cell biology functional experiment validation of the functional role of CCDC50 in HCC was performed. Compared to LO2, a human normal hepatocyte, CCDC50 was highly expressed in LIHC cell lines (Figure 10A) and inhibited by siRNA in Hep3B and Huh7 cells. QRT-PCR and western blot were employed to examined the knockdown efficacy (Figure 10B, 10C). As expected, CCDC50 knockdown inhibited the proliferation and migration abilities of Hep3B and Huh7 cells (Figure 10D, 10E).
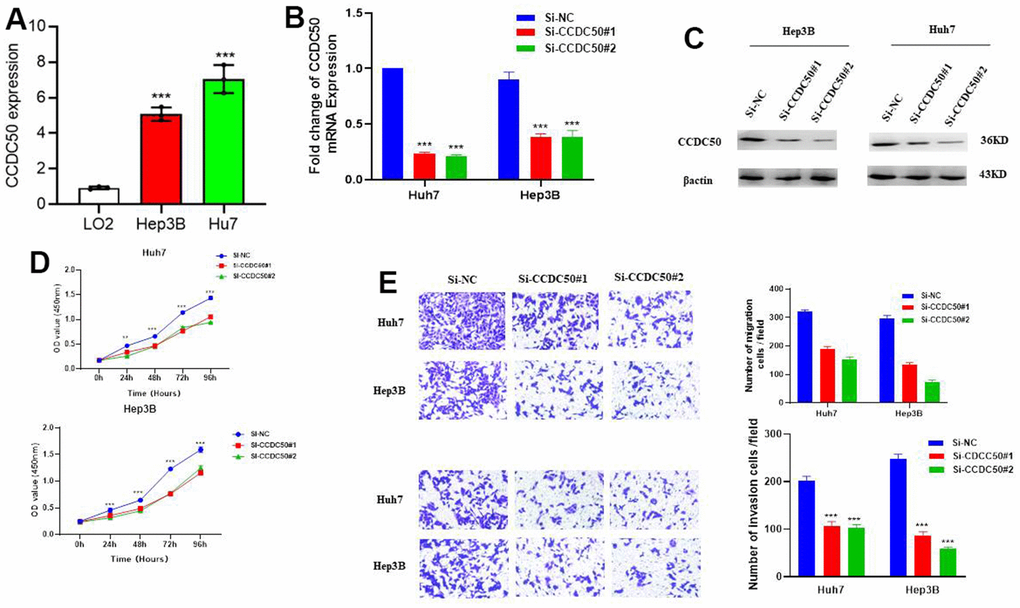
Figure 10. CCDC50 promotes the proliferation, migration, and invasion of LIHC cells. (A) The expression of CCDC50 in LIHC cell lines was examined via qRT-PCR assay. (B, C) The establishment of CCDC50 knockdown cell lines in Hep3B and Huh7 was verified via qRT-PCR assay and western blot. (D) The knockdown of CCDC50 dramatically inhibits the proliferation of Hep3B and Huh7 cells, examined via cell counting kit-8 assay. (E) The knockdown of CCDC50 dramatically inhibits the migration and invasion abilities of Hep3B and Huh7 cells. Data are presented as the mean ± SD of three independent experiments. **p < 0.01, ***p < 0.001.
Cancer drug sensitivity analysis of CCDC50
In the GDSC database, the expression of CCDC50 positively related to the following drugs: EKB-569, WZ3105, KIN001-102, AT-7519, GSK690693, BMS345541, AT-7519, GSK1070916, NPK76-II-72-1 and TAK-715. However, it was negatively correlated with the following drugs: PLX4720, dabrafenib, (5Z)-7-Oxozeaenol, bleomycin, SB590885, CHIR-99021, selumetinib, TGX221, PD-0325901, AG-014699 and piperlongumine (r < −0.13, p < 0.0001) (Figure 11A). In the CTRP database, the expression of CCDC50 positively correlated with the following drugs: SR8278, GSK-J4, austocystin D, afatinib, linifanib and BRD-K41597374 (r > 0.16, p < 0.0001). However, it was negatively associated with vemurafenib, MLN2480, fluvastatin and lovastatin (r < −0.13, p < 0.0001) (Figure 11B). Due to the heterogeneity of tumor cells, there are certain differences in the correlation between CCDC50 and different drugs in different tumors, which may have a certain impact on different tumor treatments. These results show that CCDC50 is significantly related to drug sensitivity in diverse cancer cell lines, and it has the potential to be a promising cancer therapeutic target.
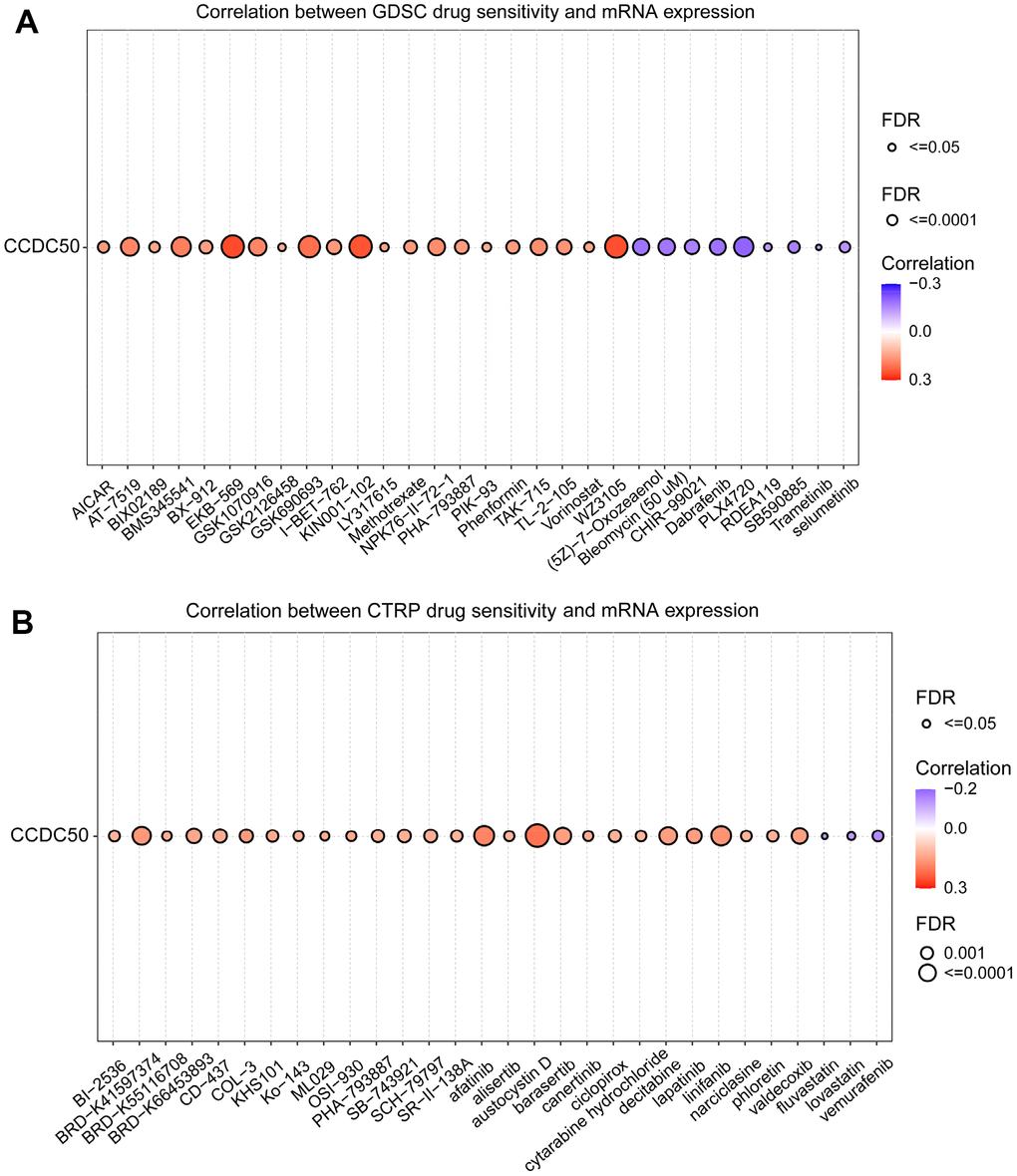
Figure 11. Analysis of the correlation between CCDC50 expression and drug sensitivity in various cancers. (A) The correlation between CCDC50 expression and drug sensitivity in various cancers was analysed via the GDSC database. (B) The correlation between CCDC50 expression and drug sensitivity in various cancers was analysed via the CTRP database.
Discussion
Prevention and treatment are important scientific problems for HCC patients. The bioinformatic analysis is critical to compare the heterogeneity among different tumours and identify novel cancer biomarkers and therapeutic targets [26, 27]. In the early stage, we also reported the relationship between CDH11 and malignant progression of gastric cancer through the public cancer database [28]. CCDC50, as a negative regulator of IFN signalling, is ubiquitously expressed in human tissues [9]. It has been reported CCDC50 is involved in the progression of renal clear cell carcinoma and mantle cell lymphoma [10, 29]. Further, CCDC50 could regulate Ras signalling pathway and promote mice hepatocellular carcinoma [12]. However, until now, no studies have assessed whether CCDC50 is associated with human HCC prognosis.
In this study, we firstly reported that CCDC50 expression was up-regulated in HCC. In addition, our results indicated that high CCDC50 expression correlated with poor prognosis in HCC, the calibration curves indicated that a nomogram is reliable to predict the 1-, 3-, and 5-year OS, and DSS in HCC. To better understand the role of CCDC50 in HCC, the KEGG enrichment analysis was performed, and it indicated that the DEGs in HCC were primarily involved in the neuroactive ligand-receptor interaction, gastric cancer, pancreatic secretion, gastric acid secretion, pertussis, glycosaminoglycan biosynthesis-keratan sulphate, axon guidance, phenylalanine metabolism, nitrogen metabolism and protein digestion and absorption.
A previous study found that 25% of HCC samples expressed inflammatory biomarkers [30]. Tumor immune cell infiltration might be associated with CDC50 expression, as shown in our study. We showed that CCDC50 correlated with these 8 major immune cells in HCC. CCDC50 expression levels showed a negative correlation with pDC, Cytotoxic cells, DC, and Tgd, and positive association with Eosinophils, Tcm, Th2, and T helper cells.
Based on the comprehensive bioinformatics analysis of liver cancer related datasets and the results of our cell biology functional experiments, we can preliminarily conclude that CCDC50 may play a role as an oncogene in the progression of liver cancer, promoting the malignant progression of liver cancer cells. In the future, it can be used as an important liver cancer biomarker and a potential therapeutic target.
Author Contributions
Qiang Wang, and Chunhong Li designed this work and performed related assay. Yingdong Jia, Na Li, Qiang Zhou, and Rui Liu analyzed data. Qiang Wang and Chunhong Li supervised and wrote the manuscript. All authors have read and approved the final version of the manuscript.
Acknowledgments
We thank Bullet Edits Limited for the linguistic editing and proofreading of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interests.
Ethical Statement
This study was approved by the Ethics Committee of Suining Central Hospital.
Funding
This study was supported by grants from the Scientific Research Project of Suining Central Hospital (No. 2022yz103), Sichuan Medical Research Project Plan (No. S22045).
References
- 1. Zhang X, Li C, Wen T, Yan L, Li B, Yang J, Wang W, Xu M, Lu W, Jiang L. Appropriate treatment strategies for intrahepatic recurrence after curative resection of hepatocellular carcinoma initially within the Milan criteria: according to the recurrence pattern. Eur J Gastroenterol Hepatol. 2015; 27:933–40. https://doi.org/10.1097/MEG.0000000000000383 [PubMed]
- 2. Chen W, Zheng R, Baade PD, Zhang S, Zeng H, Bray F, Jemal A, Yu XQ, He J. Cancer statistics in China, 2015. CA Cancer J Clin. 2016; 66:115–32. https://doi.org/10.3322/caac.21338 [PubMed]
- 3. Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer Statistics, 2021. CA Cancer J Clin. 2021; 71:7–33. https://doi.org/10.3322/caac.21654 [PubMed]
- 4. Anwanwan D, Singh SK, Singh S, Saikam V, Singh R. Challenges in liver cancer and possible treatment approaches. Biochim Biophys Acta Rev Cancer. 2020; 1873:188314. https://doi.org/10.1016/j.bbcan.2019.188314 [PubMed]
- 5. Sauerbrei W, Taube SE, McShane LM, Cavenagh MM, Altman DG. Reporting Recommendations for Tumor Marker Prognostic Studies (REMARK): An Abridged Explanation and Elaboration. J Natl Cancer Inst. 2018; 110:803–11. https://doi.org/10.1093/jnci/djy088 [PubMed]
- 6. Li X, Liu R, Su X, Pan Y, Han X, Shao C, Shi Y. Harnessing tumor-associated macrophages as aids for cancer immunotherapy. Mol Cancer. 2019; 18:177. https://doi.org/10.1186/s12943-019-1102-3 [PubMed]
- 7. Ebrahimi N, Faghihkhorasani F, Fakhr SS, Moghaddam PR, Yazdani E, Kheradmand Z, Rezaei-Tazangi F, Adelian S, Mobarak H, Hamblin MR, Aref AR. Tumor-derived exosomal non-coding RNAs as diagnostic biomarkers in cancer. Cell Mol Life Sci. 2022; 79:572. https://doi.org/10.1007/s00018-022-04552-3 [PubMed]
- 8. Bohgaki M, Tsukiyama T, Nakajima A, Maruyama S, Watanabe M, Koike T, Hatakeyama S. Involvement of Ymer in suppression of NF-kappaB activation by regulated interaction with lysine-63-linked polyubiquitin chain. Biochim Biophys Acta. 2008; 1783:826–37. https://doi.org/10.1016/j.bbamcr.2007.09.006 [PubMed]
- 9. Vazza G, Picelli S, Bozzato A, Mostacciuolo ML. Identification and characterization of C3orf6, a new conserved human gene mapping to chromosome 3q28. Gene. 2003; 314:113–20. https://doi.org/10.1016/s0378-1119(03)00710-8 [PubMed]
- 10. Farfsing A, Engel F, Seiffert M, Hartmann E, Ott G, Rosenwald A, Stilgenbauer S, Döhner H, Boutros M, Lichter P, Pscherer A. Gene knockdown studies revealed CCDC50 as a candidate gene in mantle cell lymphoma and chronic lymphocytic leukemia. Leukemia. 2009; 23:2018–26. https://doi.org/10.1038/leu.2009.144 [PubMed]
- 11. Hou P, Yang K, Jia P, Liu L, Lin Y, Li Z, Li J, Chen S, Guo S, Pan J, Wu J, Peng H, Zeng W, et al. A novel selective autophagy receptor, CCDC50, delivers K63 polyubiquitination-activated RIG-I/MDA5 for degradation during viral infection. Cell Res. 2021; 31:62–79. https://doi.org/10.1038/s41422-020-0362-1 [PubMed]
- 12. Wang H, Zhang CZ, Lu SX, Zhang MF, Liu LL, Luo RZ, Yang X, Wang CH, Chen SL, He YF, Xie D, Xu RH, Yun JP. A Coiled-Coil Domain Containing 50 Splice Variant Is Modulated by Serine/Arginine-Rich Splicing Factor 3 and Promotes Hepatocellular Carcinoma in Mice by the Ras Signaling Pathway. Hepatology. 2019; 69:179–95. https://doi.org/10.1002/hep.30147 [PubMed]
- 13. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017; 45:W98–102. https://doi.org/10.1093/nar/gkx247 [PubMed]
- 14. Chandrashekar DS, Bashel B, Balasubramanya SA, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BV, Varambally S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia. 2017; 19:649–58. https://doi.org/10.1016/j.neo.2017.05.002 [PubMed]
- 15. Mizuno H, Kitada K, Nakai K, Sarai A. PrognoScan: a new database for meta-analysis of the prognostic value of genes. BMC Med Genomics. 2009; 2:18. https://doi.org/10.1186/1755-8794-2-18 [PubMed]
- 16. Cerami E, Gao J, Dogrusoz U, Gross BE, Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, Antipin Y, Reva B, Goldberg AP, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012; 2:401–4. https://doi.org/10.1158/2159-8290.CD-12-0095 [PubMed]
- 17. Harding JJ, Nandakumar S, Armenia J, Khalil DN, Albano M, Ly M, Shia J, Hechtman JF, Kundra R, El Dika I, Do RK, Sun Y, Kingham TP, et al. Prospective Genotyping of Hepatocellular Carcinoma: Clinical Implications of Next-Generation Sequencing for Matching Patients to Targeted and Immune Therapies. Clin Cancer Res. 2019; 25:2116–26. https://doi.org/10.1158/1078-0432.CCR-18-2293 [PubMed]
- 18. Ng CK, Dazert E, Boldanova T, Coto-Llerena M, Nuciforo S, Ercan C, Suslov A, Meier MA, Bock T, Schmidt A, Ketterer S, Wang X, Wieland S, et al. Integrative proteogenomic characterization of hepatocellular carcinoma across etiologies and stages. Nat Commun. 2022; 13:2436. https://doi.org/10.1038/s41467-022-29960-8 [PubMed]
- 19. Li T, Fan J, Wang B, Traugh N, Chen Q, Liu JS, Li B, Liu XS. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017; 77:e108–10. https://doi.org/10.1158/0008-5472.CAN-17-0307 [PubMed]
- 20. Ning Y, Deng C, Li C, Peng W, Yan C, Ran J, Chen W, Liu Y, Xia J, Ye L, Wei Z, Xiang T. PCDH20 inhibits esophageal squamous cell carcinoma proliferation and migration by suppression of the mitogen-activated protein kinase 9/AKT/β-catenin pathway. Front Oncol. 2022; 12:937716. https://doi.org/10.3389/fonc.2022.937716 [PubMed]
- 21. Liu CJ, Hu FF, Xie GY, Miao YR, Li XW, Zeng Y, Guo AY. GSCA: an integrated platform for gene set cancer analysis at genomic, pharmacogenomic and immunogenomic levels. Brief Bioinform. 2023; 24:bbac558. https://doi.org/10.1093/bib/bbac558 [PubMed]
- 22. Basu A, Bodycombe NE, Cheah JH, Price EV, Liu K, Schaefer GI, Ebright RY, Stewart ML, Ito D, Wang S, Bracha AL, Liefeld T, Wawer M, et al. An interactive resource to identify cancer genetic and lineage dependencies targeted by small molecules. Cell. 2013; 154:1151–61. https://doi.org/10.1016/j.cell.2013.08.003 [PubMed]
- 23. Yang W, Soares J, Greninger P, Edelman EJ, Lightfoot H, Forbes S, Bindal N, Beare D, Smith JA, Thompson IR, Ramaswamy S, Futreal PA, Haber DA, et al. Genomics of Drug Sensitivity in Cancer (GDSC): a resource for therapeutic biomarker discovery in cancer cells. Nucleic Acids Res. 2013; 41:D955–61. https://doi.org/10.1093/nar/gks1111 [PubMed]
- 24. Hanahan D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022; 12:31–46. https://doi.org/10.1158/2159-8290.CD-21-1059 [PubMed]
- 25. Gajewski TF, Schreiber H, Fu YX. Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol. 2013; 14:1014–22. https://doi.org/10.1038/ni.2703 [PubMed]
- 26. Dohlman AB, Arguijo Mendoza D, Ding S, Gao M, Dressman H, Iliev ID, Lipkin SM, Shen X. The cancer microbiome atlas: a pan-cancer comparative analysis to distinguish tissue-resident microbiota from contaminants. Cell Host Microbe. 2021; 29:281–98.e5. https://doi.org/10.1016/j.chom.2020.12.001 [PubMed]
- 27. Yuan Y, Jiang X, Tang L, Wang J, Duan L. Comprehensive Analysis of the Prognostic and Immunological Role of PAFAH1B in Pan-Cancer. Front Mol Biosci. 2022; 8:799497. https://doi.org/10.3389/fmolb.2021.799497 [PubMed]
- 28. Wang Q, Jia Y, Peng X, Li C. Clinical and prognostic association of oncogene cadherin 11 in gastric cancer. Oncol Lett. 2020; 19:4011–23. https://doi.org/10.3892/ol.2020.11531 [PubMed]
- 29. Sun G, Zhou H, Chen K, Zeng J, Zhang Y, Yan L, Yao W, Hu J, Wang T, Xing J, Xiao K, Wu L, Ye Z, Xu H. HnRNP A1 - mediated alternative splicing of CCDC50 contributes to cancer progression of clear cell renal cell carcinoma via ZNF395. J Exp Clin Cancer Res. 2020; 39:116. https://doi.org/10.1186/s13046-020-01606-x [PubMed]
- 30. Sia D, Jiao Y, Martinez-Quetglas I, Kuchuk O, Villacorta-Martin C, Castro de Moura M, Putra J, Camprecios G, Bassaganyas L, Akers N, Losic B, Waxman S, Thung SN, et al. Identification of an Immune-specific Class of Hepatocellular Carcinoma, Based on Molecular Features. Gastroenterology. 2017; 153:812–26. https://doi.org/10.1053/j.gastro.2017.06.007 [PubMed]