



Introduction
Clear Cell Renal Cell Carcinoma (ccRCC) is a common malignant tumor with rapid development and no obvious early clinical symptoms [1, 2]. Renal cell carcinoma is a middle-aged and elderly disease. About 90% of patients diagnosed with RCC are aged 45 or above, more than 50% of patients are between 60 and 70 years old [3]. About 28% of ccRCC patients have local invasion or distant metastasis at initial diagnosis [4]. After surgical treatment, about 1/4 of localized ccRCC patients have local recurrence or distant metastasis [5]. Distant metastases are most common in lungs, lymph nodes, liver, bones and brain. The 5-year survival rate of patients with ccRCC increased from 57% in 1987–1989 to 74% in 2007–2013. However, the effective systemic efficacy of recurrent and metastatic ccRCC is still lacking, and the prognosis has been poor [2]. CcRCC patients are not sensitive to radiotherapy and chemotherapy, and surgery is still the first choice for treating ccRCC.
Related studies have shown that the invasion and distant metastasis of cancer cells are closely related to the remodeling of their energy metabolism [6, 7]. Unlike normal differentiated cells, tumor cells mainly rely on glycolysis to provide the necessary energy for rapid proliferation, which increases the metabolism and utilization of glucose [7]. Glycolysis metabolic intermediates, such as glucose-6-phosphate and pyruvate, can also be directly used by tumor cells for the synthesis of nucleotide or amino acid building blocks [8–10]. Moreover, studies have found that glycolysis metabolites can acidify the tumor microenvironment, which is conducive to the infiltration and metastasis of tumor cells and inhibit the immune effect of the body [11, 12]. Fluorouracil (5-FU) is one of the most commonly used conventional anti-metabolites chemotherapy drugs. It inhibits the DNA synthesis and repair in cancer cells. However, enhanced glycolysis in tumor cells promotes chemoresistance in cancer. Hexokinase 2 (HK2) is the key rate-limiting enzyme in the glycolysis pathway, and the expression of HK2 is increased in various tumor cells [13, 14]. At present, metabolic inhibitors for glycolysis pathway have shown a particularapplication prospect. RBM3 is a member of highly conserved RNA-binding family proteins. It is thought to be a proto-oncogene promoting cell differentiation and preventing apoptosis in several cancers [15, 16]. Under general physiological conditions, the expression of RBM3 is related to the stage of the tumor, indicating that it may be related to tumorigenesis [17]. RBM3 can improve the translation of cyclooxygenase-2, IL-8 and vascular endothelial growth factor and is essential for cells to progress through mitosis [18].
In the present study, we compared the expression of RBM3 in ccRCC tissue and normal renal tissue through bioinformatics data mining of Oncomine database and GEPIA database, and measured the level of RBM3 protein in tumor specimens of ccRCC patients by immunohistochemistry. We explored the effect of RBM3 expression level on glycolysis, chemoresistance, as well as the regulation of HK2 activity in ccRCC. Our study may help to find new potential therapeutic targets of the energy metabolism remodeling pathway for the targeted therapy of ccRCC.
Results
High expression of RBM3 in ccRCC
We first analyzed the expression of RBM3 in renal clear cell carcinoma through the oncomine database (Figure 1A). The results shown that the RBM3 in tumor tissue was hyper-expressed than that in renal tissue (p < 0.05) (Figure 1A). Similarly, the results of GEPIA database showed that RBM3 was overexpressed in renal cell carcinoma (p < 0.05) (Figure 1B). To verify these data, we collected cancer and paracancerous tissues from 30 patients with ccRCC for detection. RT-PCR results showed that RBM3 level was higher in tumor tissues (p < 0.01) (Figure 1C). Moreover, Western blot (p < 0.05) (Figure 1D) and immunohistochemical experiments (Figure 1E) showed that the level of RBM3 protein was increased in tumors. Meanwhile, the expression of higher in pT3/pT4 stage patients than that in pT1/pT2 stage patients (Figure 1F). The high expression of RBM3 was positive correlated with the poor survival of the patients (Figure 1G). Therefore, the overexpression of RBM3 may be related to the ccRCC (Figure 1).
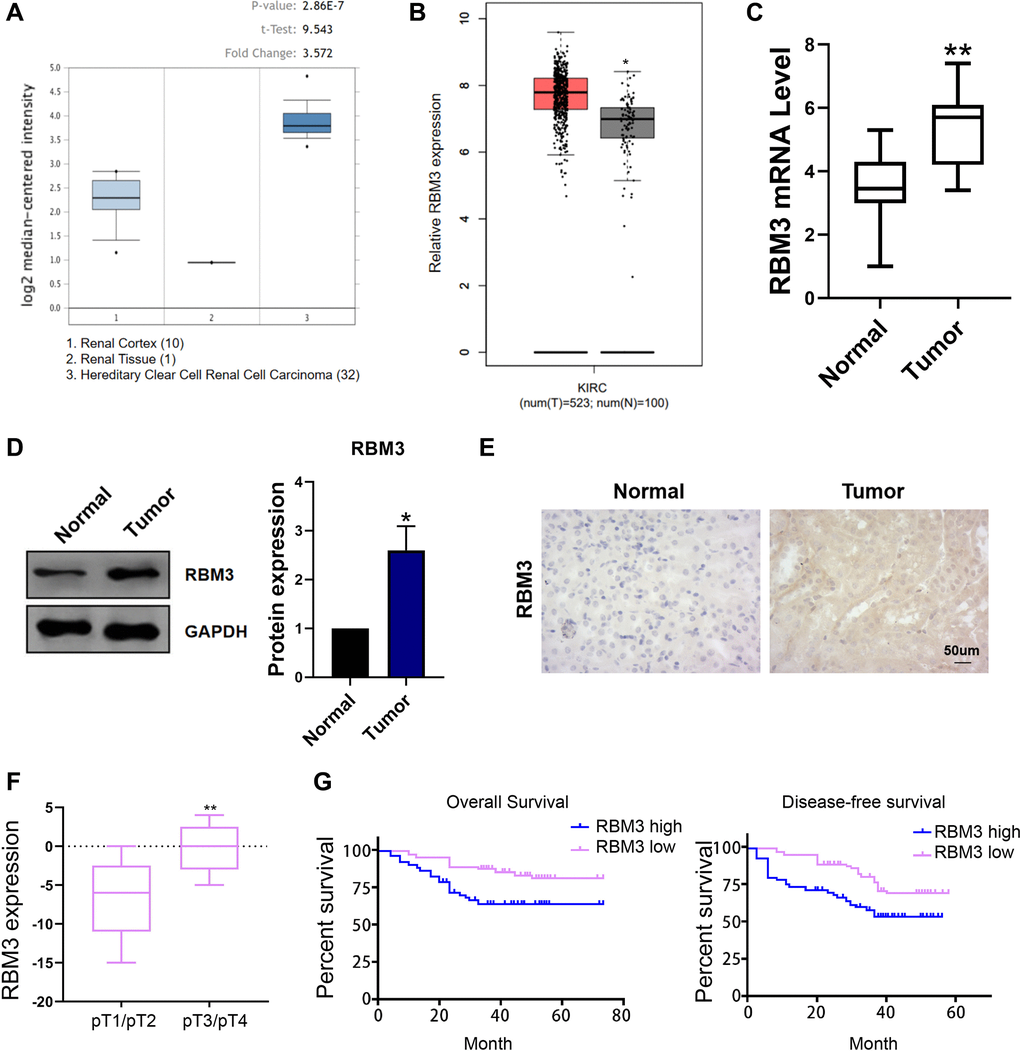
Figure 1. RBM3 level in ccRCC. (A) The expression of RBM3 was predicted based on the Oncomine database. (B) The expression of RBM3 was predicted based on the GEPIA database. (C) The expression of RBM3 in carcinoma and paracancerous tissues of 30 patients with ccRCC was detected by qRT-PCR. n = 30. (D) RBM3 protein in ccRCC and paracancerous tissues was detected using western blot. Representative image was shown on the left. Quantification data represented mean ± SD (normalized to adjacent tissue). n = 6. (E) Immunohistochemistry was used to detect RBM3 protein in renal cell carcinoma and paracancerous tissues. (F) The expression of RBM3 was analyzed by qPCR in the different stage of the patients. (G) The correlation of RBM3 expression with overall survival and disease-free survival of the patients was analyzed. Representative image was shown. *p < 0.05, **p < 0.01 vs. normal.
RBM3 promoted glycolysis in ccRCC cells
In order to analyze the effect of RBM3 on glucose consumption and lactic acid production of renal cancer cells, we constructed stable RBM3 overexpression renal cancer cell lines, and the RBM3 protein level increased steadily (p < 0.05) (Figure 2A). Compared with the control cells, the glucose consumption (p < 0.05) (Figure 2B) and lactic acid production (p < 0.05) (Figure 2C) were significantly increased. On the other hand, we used two independent siRNA targeting RBM3 to silence the RBM3 in 769-P and OSRC-2 cells (p < 0.01) (Figure 2D). The decrease of RBM3 significantly reduced the glucose consumption and lactic acid production of the two cell lines (p < 0.01) (Figure 2E–2F). In summary, these data suggested that RBM3 promotes glycolysis in renal cancer cells.
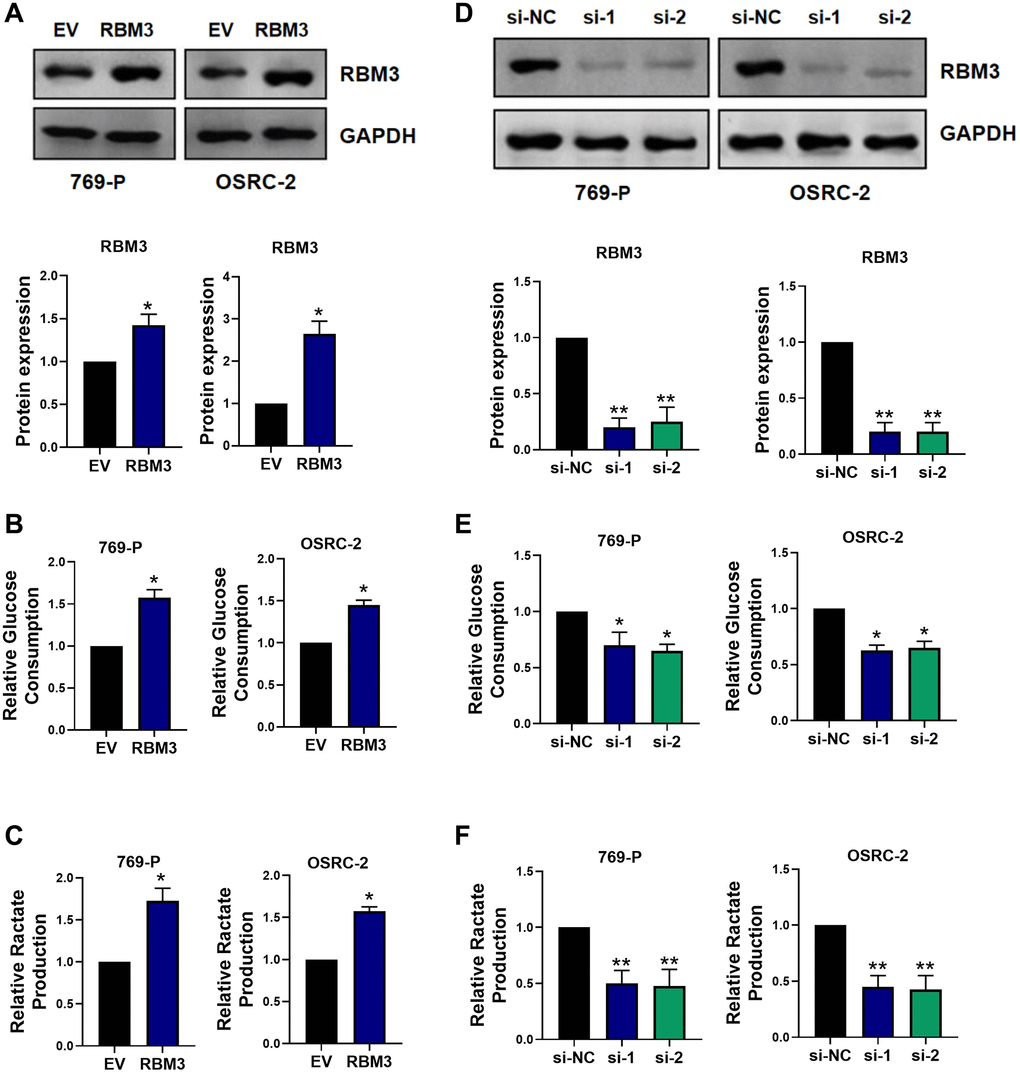
Figure 2. RBM3 facilitated aerobic glycolysis in ccRCC cells. (A) Protein levels of RBM3 was analyzed using western blot in 769-P and OSRC-2 cells. EV indicated the empty vector control; RBM3 indicated the RBM3 overexpression vector. Glucose consumption (B) and lactate production (C) were detected in both 769-P and OSRC-2 cells. n = 6. (D–F) RBM3 protein levels in 769-P and OSRC-2 cells were detected by western blot after transfection with si-RBM3-1 or si-RBM3-2 or negative control. n = 6. *p < 0.05, **p < 0.01 vs. EV or si-NC.
RBM3 regulated expression of HK2
To further investigate the regulatory function of RBM3 on glycolysis of renal clear cell carcinoma, we measured the expression of key genes related to glycolysis, such as glucose transporter 1 (Glu_1), glucose transporter 4 (Glu_4), lactate dehydrogenase A (Ldha), lactate dehydrogenase B (LdhB), HK2, pyruvate kinase M2 (PkM_2), hypoxia inducible factor-1 α (HIF-1 α) and pyruvate dehydrogenase kinase 1 (Pdk_1). We found that HK2, a well-known glycolysis mediator, was differentially expressed in 769-P or OSRC-2 cells with RBM3 overexpression (Figure 3A). On the contrary, Western blot analysis showed that RBM3 knockdown significantly decreased the HK2 protein level in 769-P or OSRC-2 cells, while the overexpression of RBM3 led to a significant increase of HK2 protein (p < 0.05) (Figure 3B). Therefore, our results show that RBM3 regulates the expression of HK2.
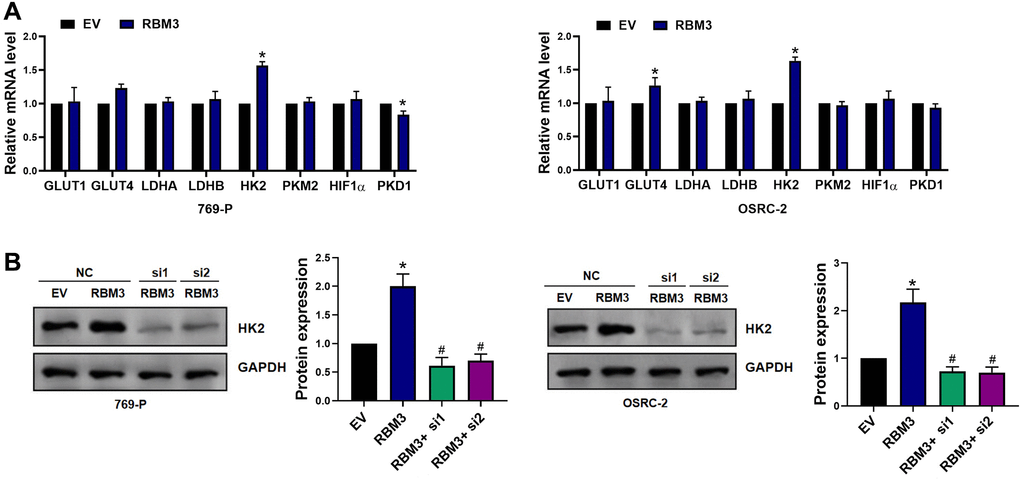
Figure 3. RBM3 promoted the expression of HK2 in ccRCC cells. (A) RT-qPCR was used to measure glycolysis-related genes in both 769-P or OSRC-2 cells, n = 6. (B) The HK2 protein level in the 769-P and OSRC-2 cells after treated with si-NC or RBM3 siRNA were tested by western blot, n = 4. *p < 0.05 vs. EV, #p < 0.05, ##p < 0.01 vs. RBM3.
High expression of HK2 in ccRCC
Oncomine database predicted the high expression of HK2 in ccRCC (p < 0.05) (Figure 4A). The GEPIA database analysis shows the same results (p < 0.05) (Figure 4B). In ccRCC tissues, the expression of HK2 was positively correlated with that of RBM3 (Figure 4C). The RT-PCR results of clinical samples revealed that HK2 was upregulated in tumor tissues (p < 0.01) (Figure 4D). Western blot and immunohistochemical experiments showed that the HK2 protein was increased in tumors (p < 0.05) (Figure 4E–4F).
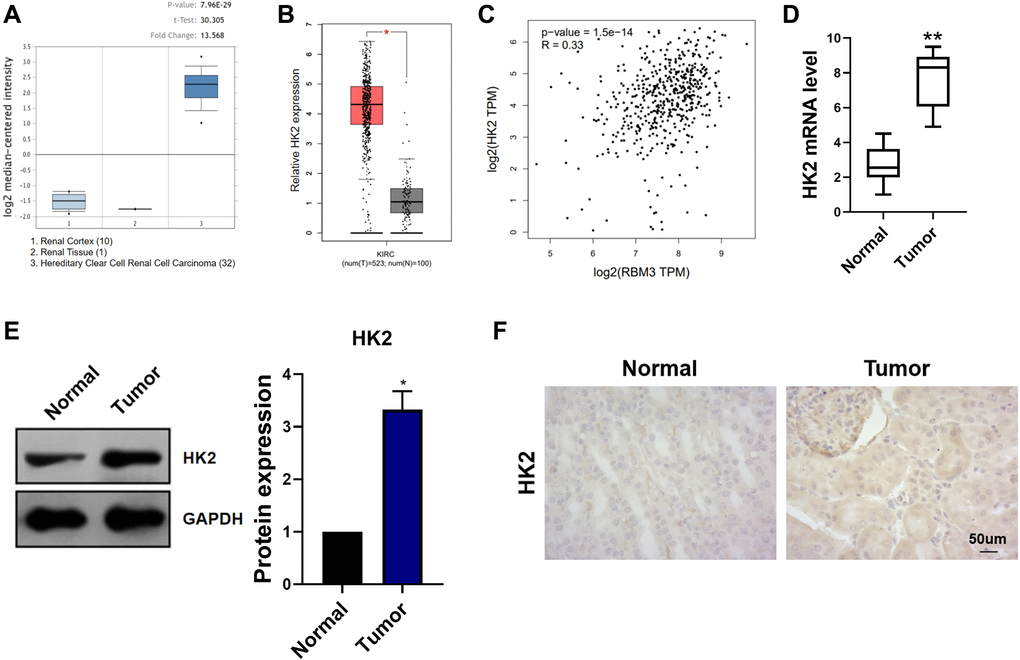
Figure 4. Expression of HK2 in ccRCC. (A) The expression of RBM3 was predicted based on the Oncomine database. (B) The expression of RBM3 was predicted based on the GEPIA database. (C) GEPIA database predicted the correlation between RBM3 and HK2 expression. (D) The expression of RBM3 in carcinoma and paracancerous tissues of 30 patients with renal cell carcinoma was detected by qRT-PCR. (E) The relative amount of the HK2 protein in ccRCC and paracancerous tissues tested by western blot, n = 4. (F) Representative immunohistochemistry image of the HK2 protein in ccRCC and paracancerous tissues. *p < 0.05, **p < 0.01 vs. normal.
RBM3 increased ccRCC cell chemoresistance via upregulating HK2
In addition to accelerating tumorigenesis, the activity of aerobic glycolysis may promote the chemoresistance of cancer cells. Previous studies have reported that knockout of HK2 enhanced the sensitivity of pleomorphic glioblastoma cells to chemotherapeutics drugs [19]. Here, we chose HK2 siRNA (25 nM) or 5 mM HK2 inhibitor 2-DG to evaluate whether HK2 plays a key role in RBM3-regulated cancer chemoresistance.
Colony formation assay was used to detect the effect of RBM3 on chemotherapy resistance of renal cell carcinoma cells. The overexpression of RBM3 s ignificantly enhanced the invasion and colony formation ability of 769-P and OSRC-2 when exposed to 5-FU (p < 0.05) (Figure 5A–5B). Apoptosis detection showed that RBM3 reduced 5-FU-induced apoptosis (Figure 5C). In addition, MTT showed that RBM3 overexpression enhanced the resistance of 769-P or OSRC-2 to 5-FU (Figure 5D). In order to verify whether the overexpression of RBM3 leads to the resistance to chemotherapeutic drugs through the increase of glucose metabolism, 769-P and OSRC-2 cells with overexpression of RBM3 were treated with HK2 siRNA or 2-DG. We found that the resistance of 769-P and OSRC-2 cells overexpressing RBM3 to 5-FU could be reversed by si-HK2 or 2-DG (p < 0.05) (Figure 5D).
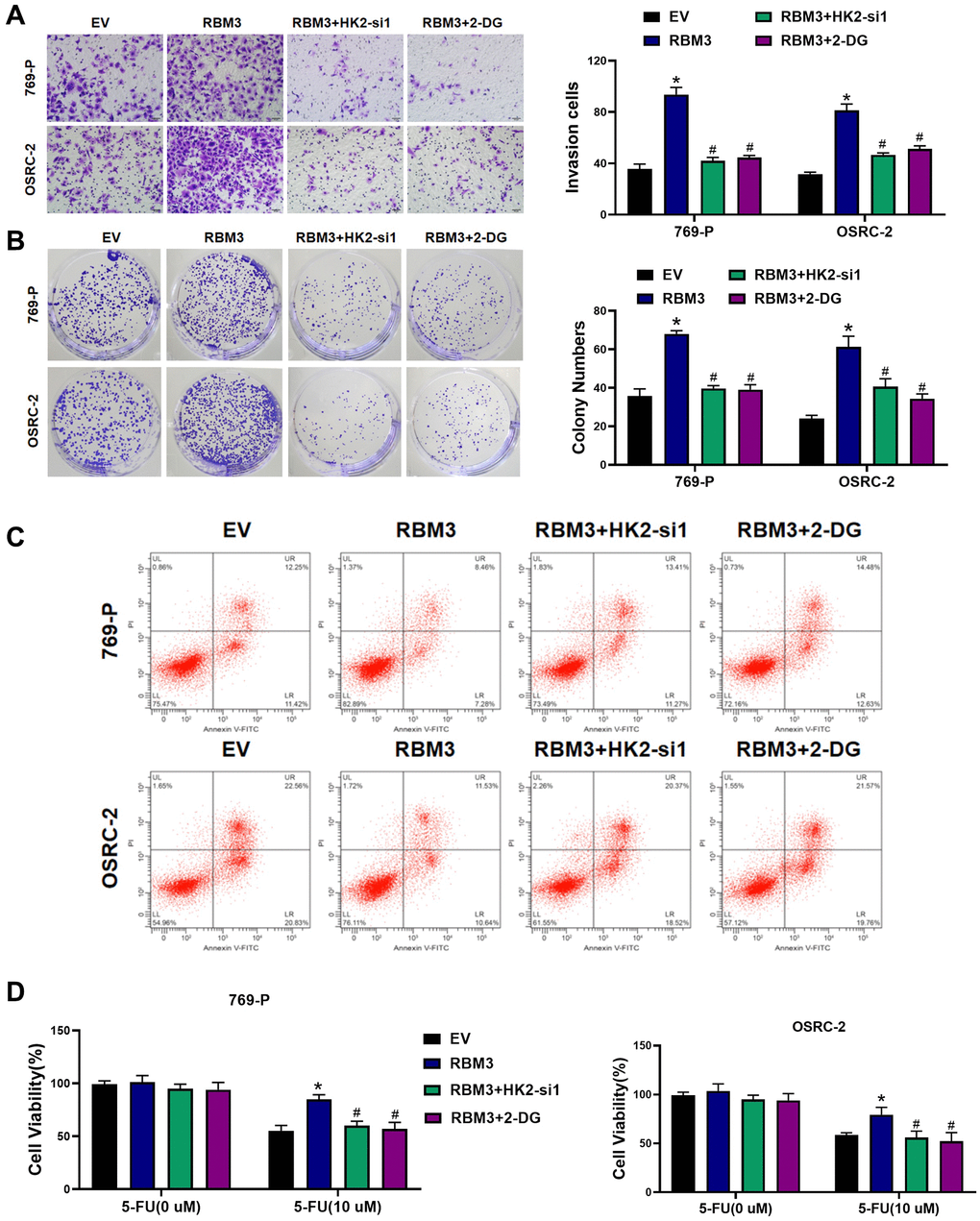
Figure 5. RBM3 regulated the chemoresistance of ccRCC cells by HK2 in vitro. (A) Cells invasion of the 769-P and OSRC-2 cells exposed to 5-FU. Representative images were shown on the left. Bar graphs were mean ± SD. n = 5. (B) Colony formation of the 769-P and OSRC-2 cells exposed to 5-FU, n = 3. (C) The effect of RBM3 on cell apoptosis was abolished by HK2 siRNA or 2-DG. (D) MTT was used to measure cell viability, n = 10. *p < 0.05 vs. EV; #p < 0.05 vs. RBM3.
RBM3 regulated HK2 induced ccRCC chemoresistance in vivo
In order to confirm the chemotherapeutics resistance function of RBM3 to renal cell carcinoma, the transplanted tumor models of OSRC-2 cells overexpressing RBM3 was established in nude mice, and 5-FU were given at different time points (Figure 6A). Tumor volume and weight were compared among different treatment groups. We found that the mass and volume of OSRC-2 tumors with overexpression of RBM3 were significantly higher than those of the control group (Figure 6B), indicating RBM3 overexpression promotes 5-FU resistance. In order to confirm whether the drug resistance of renal cell carcinoma regulated by RBM3 is dependent on HK2 in vivo, the mouse model of transplanted tumor was injected with 2-DG (p < 0.05) (Figure 6B–6E). The expression of RBM3 and HK2 was validated in the model (Figure 6F). The result showed that 2-DG significantly reversed the 5-FU resistance of tumors in vivo. The results suggest that RBM3-induced glycolysis promotes the chemotherapy resistance of renal cell carcinoma cells, and interfering with this process can reverse the chemotherapeutic resistance of ccRCC.
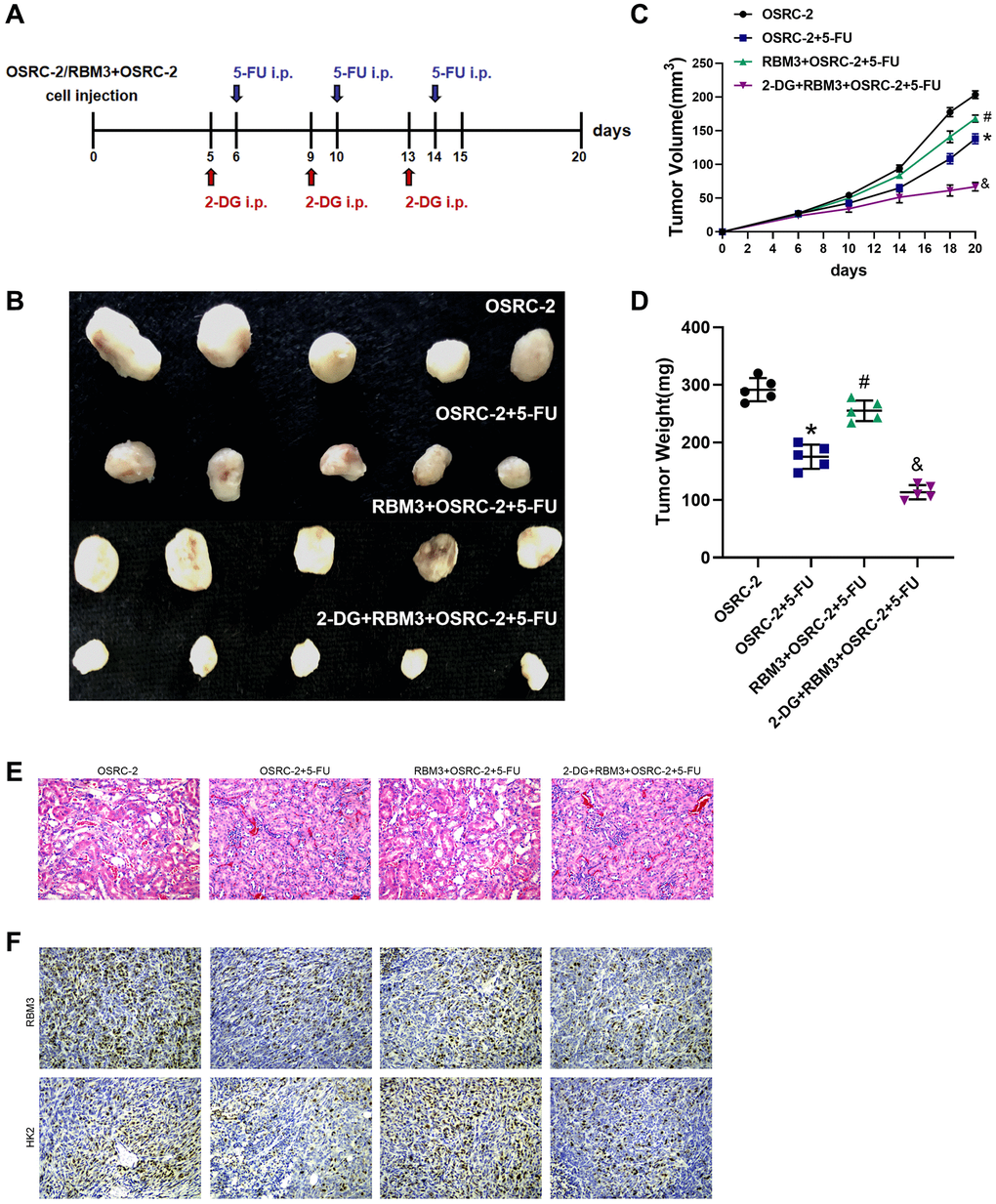
Figure 6. RBM3 dependent on HK2 to regulate the chemoresistance of ccRCC in vivo. (A) Schematic overview of the 5-FU/2-DG treatment and xenograft tumor model. (B) A representative image of RBM3-overexpressing OSRC-2 tumors in nude mice. (C) The volumes of the RBM3-overexpressing OSRC-2 tumors with or without 5-FU treatment. (D) The weights of the RBM3-overexpressing OSRC-2 tumors in nude mice with or without 5-FU treatment. (E) The injury was detected by hematoxylin and eosin staining. (F) The levels of RBM3 and HK2 were analyzed by immunohistochemical staining. n = 5, *p < 0.05 vs. OSRC-2; #p < 0.05 vs. OSRC-2+5FU; &p < 0.05 vs. RBM3+OSRC-2+5FU.
Discussion
CcRCC is one of the most common urological tumors, with high morbidity and mortality and poor prognosis. At present, the treatment for advanced ccRCC is very limited, and the main reason of its poor efficacy is due to the insensitive to adjuvant therapy, such as radiotherapy and chemotherapy. At present, targeted antiangiogenic drugs are mainly used for adjuvant therapy, but they are only effective in certain patients, and most patients display drug resistance in varying degrees [20]. Many ccRCC patients are in advanced stage when they are first diagnosed, and 20–30% of them may have recurrence and metastasis after operation. Therefore, it is of great clinical significance to study the mechanism of carcinogenesis of ccRCC and to find reliable and effective molecular biomarkers for the early diagnosis, prognosis prediction and clinical treatment of ccRCC.
RBM3 has been found to be highly expressed in some other invasive and malignant tumors. The expression of RBM3 is related to tumor pathological stage, and contributes to the invasion and proliferation, but the mechanism is not clear. It was speculated that RBM3 may promote the progression of early tumorigenesis by affecting the DNA damage repair and the chromosome stability, thus increasing the chance of malignancy [21, 22]. Meanwhile, it has been found that RBM3 contributes to cell proliferation of hepatocellular carcinoma [23]. The effects of RBM3 expression on ccRCC energy metabolism and chemoresistance has been unclear.
In this study, we reported the regulatory effect of RBM3 on glycolysis and chemotherapy resistance of renal clear cell carcinoma. Bioinformatics has been widely used in biomedical research [24, 25]. By analyzing the data set of RBM3 expression in ccRCC and normal renal tissue in Oncomine database and GEPIA database, we found that RBM3 gene was highly expressed in ccRCC compared with normal renal tissue, which suggested that RBM3 is a potential diagnostic biomarker of ccRCC. Meanwhile, 5-FU-based chemotherapy is not considered as standard therapy for patients with ccRCC and the effect of RBM3 on other standard chemotherapy resistance in ccRCC should be explored in future investigations. In addition, it has been reported that RBM3 is highly expressed in the samples of surgically treated colorectal lung metastases and paired primary tumors and is a promising clinical prognostic factor [26]. A meta-analysis also demonstrates the prognostic and clinicopathological significance of RBM3 in differect cancer types [27]. In our clinical analysis, we found that RBM3 was elevated in clinical ccRCC samples and the high expression of RBM3 was positive correlated with the poor survival of ccRCC patients. The prognostic and clinical significance of RBM3 deserved to more exploration in future investigations.
Using ccRCC tissues and adjacent tissues from patients, we found that RBM3 was upregulated in cancer tissues. Overexpression of RBM3 in cancer cell lines enhanced glucose consumption, while downregulation of RBM3 showed the opposite effect. Through screening of a panel of genes, we found the elevated glucose consumption was mediated by enhanced HK2 expression. The upregulation of HK2 was confirmed in cancer tissues as well. 5-FU is one of the most commonly used conventional anti-metabolites chemotherapy drugs. We used 5-FU as the chemoresistance drug model of ccRCC in our study. Downregulation of HK2 by siRNA of inhibitor reduced the chemoresistance, invasion and cell viability of cancer cells in vitro. Importantly, inhibiting the glycolysis reduced the chemoresistance of tumor volume and weight in nude mice tumor model.
Tumor cells promote and maintain the ability of continuous proliferation, invasion and metastasis through the reconstruction of energy metabolism system. HK2 is rarely distributed in normal human body. However, studies have confirmed the high expression of HK2 in glioma, cervical cancer, metastatic carcinoma, thymoma and many other tumor tissues. HK2 overexpression has been correlated with malignant stage and enhanced glycolysis of cancer cells [28]. We confirmed that HK2 was higher expressed in ccRCC and regulated by RBM3. Our data suggested that RBM3 was an important gene in the energy metabolism of ccRCC, and RBM3 affected the drug resistance of ccRCC through modulating HK2 activity. Overall, RBM3 is a promising therapeutic target of clear cell renal cell carcinoma.
The expression of some proteins such as hERG1 and CA IX are associated with disease recurrence in surgically resected ccRCC [26]. In our study, due to the low sample number of ccRCC patients, we did not observe a potential correlation between the level of RBM3 expression and the recurrence and prognosis. In addition, although we showed RBM3 affected the HK2 activity and chemoresistance, the potential regulating roles of other unidentified factors could not be ruled out. More detailed study with large sample number is needed to address these questions in the future.
Conclusions
RBM3 was overexpressed in ccRCC cells and it promoted the malignancy by upregulation of HK2. Our data suggested that RBM3 is a promising therapeutic target of clear cell renal cell carcinoma.
Methods
Tissue specimen
The surgical specimens of 30 patients with ccRCC from the hospital included ccRCC tissue and adjacent normal tissue, which were used for this study. All the cases were clear cell carcinoma in this study, which were confirmed after nephrectomy. The clinical information of the samples was shown in Supplementary Table 1. The experiment was permitted by the Ethics Review Committee of the Third Hospital of Jilin University and the patients signed informed consent.
Cell culture
769-P, ORSC-2 cell lines and RBM3 stable-transfected cells were purchased from CHI Scientific, Inc (Jiangsu, China). The cells were cultured in a complete 1640 medium including 10% FBS from Biological Industries (Beit-Haemek, Israel), and maintained in a humidified incubator with 37°C and 5% of CO2. The concentration of 5-FU was 15 nmol/mL [29].
Animals
Animal experiments were permitted by the Animal Protection and Ethics Committee of the Third Hospital of Jilin University. BALB/c nude mice (6–8 weeks) were purchased from Beijing Weitong Lihua Experimental Animal Technology Co., Ltd. (Beijing, China). For the experiment of Xenograft, OSRC-2 cells (5 × 106) were suspended in 200 μl normal saline and injected subcutaneously. Tumor volume (mm3): V (mm3) = S2 (mm2) × L (mm)/2. In order to determine the resistance of RBM3 and HK2 to 5-FU, mice were randomly divided into empty vector OSRC-2 group, 5-FU chemotherapy group (5-FU-OSRC-2), 5-FU chemotherapy group with overexpression of RBM3 (RBM3+OSRC-2+5-FU) and HK2 inhibitor treatment group (2-DG+RBM3+OSRC-2+5-FU). The 5-FU (10 mg/kg; 6, 10, 14 days) [29] and 2-DG (500 mg/kg; 5, 9, 13 days) (Sigma) [30] were intraperitoneal injected after tumor implantation. The hematoxylin and eosin staining was performed to assess the injury. The levels of RBM3 and HK2 were analyzed by immunohistochemical staining in the model.
Colony formation experiment
About 300 cells were inoculated into a 10-cm dish. After the cells were evenly distributed, they were cultured in a cell incubator for 14 ~ 21 days. The medium was abandoned and 4% paraformaldehyde was added to fix the cells for 15 min. After abandoning the fixed solution, Giemsa was added to stain the cells for 15 min. Rinse the dish with running water and dry. The number of colony was counted under microscope.
Glucose consumption and lactic acid production test
The glucose quantification kit (glucose oxidase-peroxidase method) was bought from Rongsheng Biopharmaceutical Co., Ltd. (Shanghai, China), the lactic acid detection kit was bought from Jikai Co., Ltd. (Jiangsu, China). The experiment was performed according to the product manual.
qRT-PCR
RNA extraction was performed using Trizol reagent. NanoDrop 8000 (Thermo Scientific, Waltham, MA, USA) was used to measure the concentration and purity of RNA. The single-stranded cDNAs were synthesized from total RNA using TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix (Transgen). The expression of mRNAs and miRNAs were quantified by RT-PCR with SYBR Green I (Thermo Fisher Scientific, Inc).
Matrigel transwell assay
24-well matrigel transwell (Corning, USA) were used to investigated cell invasion. 2 × 105 769-P or ORSC-2 cells were seeded in insert precoated with 1 μg/μL Matrigel (BD Biosciences, USA). Medium include 20% FBS was used to stimulate invasion in the bottom of wells. After 48 h, the invaded cells were stained with a 0.1% crystal violet.
Immunohistochemical staining
Paraffin sections of carcinoma or adjacent tissue were dewaxing to water in xylene and descending series of ethanol. Sections were penetrated using 0.5% Triton X-100. After 3 times wash, sections were blocked with 50% goat serum. Then, sections were incubated with RBM3 or HK2 primary antibody overnight. Sections were incubated with secondary antibody followed by DAPI staining. The sections were photographed under an IX73 fluorescence microscope (Olympus, Valley, PA, USA) and analyzed by Image J software.
Transfection
Plasmid of HK2 or small interfering RNA (siRNA) of RBM3 and HK2 were constructed by Genechem (Shanghai, China). The 769-P or ORSC-2 cells were transfected using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA).
MTT assay
The cell viability was detected by MTT assay. The 769-P and ORSC-2 cells were conducted with indicated treatment, and placed in 96-well plates at a density of 4 × 103 cells per well to growth for 48 hours. Then 0.5 mg/mL of MTT reagent was added in each well to incubate for another 4 hours. At the end time point, the medium with MTT was discarded and 150 μL DMSO was added in each well. The absorbance values were detected at 490 nm using a microplate reader (PerkinElmer, CA, USA).
Detection of apoptosis
The Annexin V-FITC/PI apoptosis kit was purchased from Solarbio (Beijing, China). The 769-P or ORSC-2 cells were suspended with 500 ul of binding buffer and mixed with 5 ul of annexin V-FITC and PI, respectively, and placed at 25°C for 15 min. The apoptosis rate was determined by flow cytometry.
Western blot
After RIPA lysis, total proteins were extracted from clinical tumor tissues, adjacent non-tumor tissues, and 769-P and ORSC-2 cells and measured with BCA method. After SDS-PAGE separation, protein binds were transferred onto PVDF membrane and blocked using 5% non-fat milk. The primary antibody of RBM3 (1:500) and HK2 (1:1000), and HRP-anticonjugated secondary antibodies were purchased from Affinity Biosciences (Jiangsu, China). The expression of the protein was expressed by the gray value.
Statistical analysis
Statistical analysis of the data was performed using GraphPad Prism 8.0 software. The results were expressed in the form of mean ± standard deviation [23]. Two-tailed Student's t-test was used to compare data of the two groups, and one-way ANOVA analysis was used to evaluate the differences among multiple groups. P < 0.05 was considered that the difference was statistically significant.
Supplementary Materials
Author Contributions
All authors contributed to the study conception and design. Material preparation, experiments performed, data collection and analysis were performed by Siqi Zhang, Xiaohua Xu and Qianyu Li. The first draft of the manuscript was written by Na Li, Jing Yu, Feng Liu and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflicts of interest.
References
- 1. Cronin KA, Ries LA, Edwards BK. The Surveillance, Epidemiology, and End Results (SEER) Program of the National Cancer Institute. Cancer. 2014 (Suppl 23); 120:3755–7. https://doi.org/10.1002/cncr.29049 [PubMed]
- 2. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018; 68:7–30. https://doi.org/10.3322/caac.21442 [PubMed]
- 3. King SC, Pollack LA, Li J, King JB, Master VA. Continued increase in incidence of renal cell carcinoma, especially in young patients and high grade disease: United States 2001 to 2010. J Urol. 2014; 191:1665–70. https://doi.org/10.1016/j.juro.2013.12.046 [PubMed]
- 4. Keegan KA, Schupp CW, Chamie K, Hellenthal NJ, Evans CP, Koppie TM. Histopathology of surgically treated renal cell carcinoma: survival differences by subtype and stage. J Urol. 2012; 188:391–7. https://doi.org/10.1016/j.juro.2012.04.006 [PubMed]
- 5. Dabestani S, Thorstenson A, Lindblad P, Harmenberg U, Ljungberg B, Lundstam S. Renal cell carcinoma recurrences and metastases in primary non-metastatic patients: a population-based study. World J Urol. 2016; 34:1081–6. https://doi.org/10.1007/s00345-016-1773-y [PubMed]
- 6. Lee SY, Ju MK, Jeon HM, Lee YJ, Kim CH, Park HG, Han SI, Kang HS. Oncogenic Metabolism Acts as a Prerequisite Step for Induction of Cancer Metastasis and Cancer Stem Cell Phenotype. Oxid Med Cell Longev. 2018; 2018:1027453. https://doi.org/10.1155/2018/1027453 [PubMed]
- 7. Porporato PE, Payen VL, Baselet B, Sonveaux P. Metabolic changes associated with tumor metastasis, part 2: Mitochondria, lipid and amino acid metabolism. Cell Mol Life Sci. 2016; 73:1349–63. https://doi.org/10.1007/s00018-015-2100-2 [PubMed]
- 8. Zhang C, Zhang Z, Zhu Y, Qin S. Glucose-6-phosphate dehydrogenase: a biomarker and potential therapeutic target for cancer. Anticancer Agents Med Chem. 2014; 14:280–9. https://doi.org/10.2174/18715206113136660337 [PubMed]
- 9. Zhong J, Rajaram N, Brizel DM, Frees AE, Ramanujam N, Batinic-Haberle I, Dewhirst MW. Radiation induces aerobic glycolysis through reactive oxygen species. Radiother Oncol. 2013; 106:390–6. https://doi.org/10.1016/j.radonc.2013.02.013 [PubMed]
- 10. Mou JJ, Peng J, Shi YY, Li N, Wang Y, Ke Y, Zhou YF, Zhou FX. Mitochondrial DNA content reduction induces aerobic glycolysis and reversible resistance to drug-induced apoptosis in SW480 colorectal cancer cells. Biomed Pharmacother. 2018; 103:729–37. https://doi.org/10.1016/j.biopha.2018.04.099 [PubMed]
- 11. Jiang Z, Liu Z, Li M, Chen C, Wang X. Increased glycolysis correlates with elevated immune activity in tumor immune microenvironment. EBioMedicine. 2019; 42:431–42. https://doi.org/10.1016/j.ebiom.2019.03.068 [PubMed]
- 12. Robertson-Tessi M, Gillies RJ, Gatenby RA, Anderson AR. Impact of metabolic heterogeneity on tumor growth, invasion, and treatment outcomes. Cancer Res. 2015; 75:1567–79. https://doi.org/10.1158/0008-5472.CAN-14-1428 [PubMed]
- 13. Li W, Qiu Y, Hao J, Zhao C, Deng X, Shu G. Dauricine upregulates the chemosensitivity of hepatocellular carcinoma cells: Role of repressing glycolysis via miR-199a:HK2/PKM2 modulation. Food Chem Toxicol. 2018; 121:156–65. https://doi.org/10.1016/j.fct.2018.08.030 [PubMed]
- 14. Tech K, Deshmukh M, Gershon TR. Adaptations of energy metabolism during cerebellar neurogenesis are co-opted in medulloblastoma. Cancer Lett. 2015; 356:268–72. https://doi.org/10.1016/j.canlet.2014.02.017 [PubMed]
- 15. Dong W, Dai ZH, Liu FC, Guo XG, Ge CM, Ding J, Liu H, Yang F. The RNA-binding protein RBM3 promotes cell proliferation in hepatocellular carcinoma by regulating circular RNA SCD-circRNA 2 production. EBioMedicine. 2019; 45:155–67. https://doi.org/10.1016/j.ebiom.2019.06.030 [PubMed]
- 16. Ma R, Zhao LN, Yang H, Wang YF, Hu J, Zang J, Mao JG, Xiao JJ, Shi M. RNA binding motif protein 3 (RBM3) drives radioresistance in nasopharyngeal carcinoma by reducing apoptosis via the PI3K/AKT/Bcl-2 signaling pathway. Am J Transl Res. 2018; 10:4130–40. [PubMed]
- 17. Grupp K, Wilking J, Prien K, Hube-Magg C, Sirma H, Simon R, Steurer S, Budäus L, Haese A, Izbicki J, Sauter G, Minner S, Schlomm T, Tsourlakis MC. High RNA-binding motif protein 3 expression is an independent prognostic marker in operated prostate cancer and tightly linked to ERG activation and PTEN deletions. Eur J Cancer. 2014; 50:852–61. https://doi.org/10.1016/j.ejca.2013.12.003 [PubMed]
- 18. Sureban SM, Ramalingam S, Natarajan G, May R, Subramaniam D, Bishnupuri KS, Morrison AR, Dieckgraefe BK, Brackett DJ, Postier RG, Houchen CW, Anant S. Translation regulatory factor RBM3 is a proto-oncogene that prevents mitotic catastrophe. Oncogene. 2008; 27:4544–56. https://doi.org/10.1038/onc.2008.97 [PubMed]
- 19. Zhang J, Chen G, Gao Y, Liang H. HOTAIR/miR-125 axis-mediated Hexokinase 2 expression promotes chemoresistance in human glioblastoma. J Cell Mol Med. 2020; 24:5707–17. https://doi.org/10.1111/jcmm.15233 [PubMed]
- 20. van der Mijn JC, Mier JW, Broxterman HJ, Verheul HM. Predictive biomarkers in renal cell cancer: insights in drug resistance mechanisms. Drug Resist Updat. 2014; 17:77–88. https://doi.org/10.1016/j.drup.2014.10.003 [PubMed]
- 21. Ehlén Å, Nodin B, Rexhepaj E, Brändstedt J, Uhlén M, Alvarado-Kristensson M, Pontén F, Brennan DJ, Jirström K. RBM3-regulated genes promote DNA integrity and affect clinical outcome in epithelial ovarian cancer. Transl Oncol. 2011; 4:212–21. https://doi.org/10.1593/tlo.11106 [PubMed]
- 22. Ehlén A, Brennan DJ, Nodin B, O'Connor DP, Eberhard J, Alvarado-Kristensson M, Jeffrey IB, Manjer J, Brändstedt J, Uhlén M, Pontén F, Jirström K. Expression of the RNA-binding protein RBM3 is associated with a favourable prognosis and cisplatin sensitivity in epithelial ovarian cancer. J Transl Med. 2010; 8:78. https://doi.org/10.1186/1479-5876-8-78 [PubMed]
- 23. Vidarsdottir H, Siesing C, Nodin B, Jönsson P, Eberhard J, Jirström K, Brunnström H. Clinical significance of RBM3 expression in surgically treated colorectal lung metastases and paired primary tumors. J Surg Oncol. 2021; 123:1144–56. https://doi.org/10.1002/jso.26375 [PubMed]
- 24. Xu T, Ruan H, Song Z, Cao Q, Wang K, Bao L, Liu D, Tong J, Yang H, Chen K, Zhang X. Identification of CXCL13 as a potential biomarker in clear cell renal cell carcinoma via comprehensive bioinformatics analysis. Biomed Pharmacother. 2019; 118:109264. https://doi.org/10.1016/j.biopha.2019.109264 [PubMed]
- 25. Jardillier R, Chatelain F, Guyon L. Bioinformatics Methods to Select Prognostic Biomarker Genes from Large Scale Datasets: A Review. Biotechnol J. 2018; 13:e1800103. https://doi.org/10.1002/biot.201800103 [PubMed]
- 26. Miao X, Zhang N. Role of RBM3 in the regulation of cell proliferation in hepatocellular carcinoma. Exp Mol Pathol. 2020; 117:104546. https://doi.org/10.1016/j.yexmp.2020.104546 [PubMed]
- 27. Gao G, Shi X, Yao Z, Shen J, Shen L. Identification of lymph node metastasis-related microRNAs in breast cancer using bioinformatics analysis. Medicine (Baltimore). 2020; 99:e22105. https://doi.org/10.1097/MD.0000000000022105 [PubMed]
- 28. Garcia SN, Guedes RC, Marques MM. Unlocking the Potential of HK2 in Cancer Metabolism and Therapeutics. Curr Med Chem. 2019; 26:7285–322. https://doi.org/10.2174/0929867326666181213092652 [PubMed]
- 29. Xu T, Guo P, Pi C, He Y, Yang H, Hou Y, Feng X, Jiang Q, Wei Y, Zhao L. Synergistic Effects of Curcumin and 5-Fluorouracil on the Hepatocellular Carcinoma In vivo and vitro through regulating the expression of COX-2 and NF-κB. J Cancer. 2020; 11:3955–64. https://doi.org/10.7150/jca.41783 [PubMed]
- 30. Shi T, Ma Y, Cao L, Zhan S, Xu Y, Fu F, Liu C, Zhang G, Wang Z, Wang R, Lu H, Lu B, Chen W, Zhang X. B7-H3 promotes aerobic glycolysis and chemoresistance in colorectal cancer cells by regulating HK2. Cell Death Dis. 2019; 10:308. https://doi.org/10.1038/s41419-019-1549-6 [PubMed]