



Introduction
Aging is broadly defined as a time-dependent functional decline that most living organisms seem to be unable to escape. It is one of the leading risk factors for a number of aging-associated diseases including atherosclerosis, type 2 diabetes, cardiovascular diseases, cancer and Alzheimer's disease [1]. Several lines of evidence suggest that slowing down aging process reduces the incidence of age-related diseases and extends organismal lifespan and health span [2,3]. Most of the mechanistic studies on aging have involved various model organisms, including multicellular worm, fly, mouse, monkey, as well as single-cellular yeast (e.g. Saccharomyces cerevisiae). Two forms of lifespan in yeast S. cerevisiae are usually used to describe aging, namely replicative lifespan (RLS) and chronological lifespan (CLS) [4]. The RLS measures the exact number of daughter cells produced by a single mother cell before cell death [5]. CLS is defined as the length of time that the cells at stationary phase or G0 phase (quiescence state) can be viable and reenter cell cycle upon nutrient availability [6]. The replicative aging models the aging process of dividing cells, while the chronological aging mirrors the aging process of non-dividing (post-mitotic) cells. Numerous studies on yeast chronological aging have been accelerating the pace of revealing the mystery of aging and longevity [4,7–13].
Two evolutionarily conserved nutrient-sensing signaling pathways, the Tor (target of rapamycin)/Sch9 pathway and Ras/PKA pathway, are first demonstrated to modulate CLS in the budding yeast S. cerevisiae [14]. These two signaling pathways are later found to regulate lifespan in higher organisms including mammals as well [2,15–18]. Sch9 is a direct downstream kinase of TORC1 (the target of rapamycin complex 1) [19]. Deletion of SCH9 in yeast inhibits genome instability during chronological aging, and results in significant increase of CLS [14]. The evolutionarily conserved Tor/Sch9 pathway and Ras/PKA pathway also regulate replicative aging [4,20].
Genome instability, one of the hallmarks of aging [1,21], is deleterious for maintaining a long lifespan in many organisms ranging from yeast to mammals [8,21–25]. In S. cerevisiae, the rDNA stability is suggested to regulate both replicative and chronological aging [26–28]. Sir2 and Fob1 play opposite role in rDNA recombination, i.e. Sir2 inhibits, while Fob1 promotes rDNA recombination [29]. Multimer ERC accumulation is significantly enhanced in mutants that have aberrant cell cycle checkpoint control during chronological aging [26]. Sgs1 is a DNA helicase, whose deletion causes much higher gene mutation frequency and gross chromosomal rearrangement (GCR) frequency than wild-type cells during chronological aging [22], but does not result in elevated level of ERCs [30–32]. The phenotypes of genome instability can be suppressed by SCH9 deletion [22]. The Bloom syndrome helicase (BLM) and Werner’s syndrome helicase (WRN) are mammalian orthologs of yeast Sgs1. They are involved in the maintenance of genome stability [22,33]. Mutations in human BLM or WRN may cause pre-mature aging syndrome [34].
Telomeres, the protective DNA-protein structures at the ends of eukaryotic chromosomes, are essential for genome integrity and stability [35]. In S. cerevisiae, telomeric DNA consists of ~350 bp of TG1–3/ C1–3A repeats. The G strand extends beyond its complementary strand to form a single-stranded overhang (called G-overhang) [36]. Telomeric DNA is mainly elongated by a specialized reverse transcriptase called telomerase. Telomerase consists of at least four subunits, the catalytic protein subunit Est2, the RNA template subunit Tlc1 and two accessory subunits Est1 and Est3. Cdc13 is a telomeric single stranded DNA binding protein, which associates with telomeres throughout the cell cycle [37]. While Est1 abundance is cell cycle regulated, which binds telomeres late in S phase . Est1 may interacts with Cdc13 in S phase to convert inactive telomerase to active form [37–40]. The expression of fusion protein Cdc13-Est2 forces telomerase to be constitutively tethered to telomeres, and leads to progressive and over elongation of telomeres [41]. Rif1 and Rif2 are two negative regulators of telomerase [42,43]. Deletion of either RIF1 or RIF2 causes telomere lengthening but rif1∆ cells have much longer telomeres than rif2∆ cells [42,43].
In yeast the absence of telomerase leads to shortening of telomeric TG1-3 DNA at a rate of 2.5-5 base pairs per population doubling [44], and telomerase-null cells eventually cease to divide when telomeres reach critically short length, resulting in replicative senescence [45], which represents a different aging model from the telomere-length indenpendent replicative aging and chronological aging aforementioned. In most mammalian somatic cells, low or no telomerase activity also results in gradual attrition of telomeres and cellular or organismal aging [46–48]. Conversely, experimental telomere lengthening is correlated with increased lifespan in mice [49,50]. Polymorphisms in telomere maintenance factors that lead to longer telomeres are associated with diminished age-related pathology in humans [51]. Additionally, transient overexpression of hTERT in human cells lengthens telomeres and extends RLS [52]. Moreover, overexpression of a telomere binding protein HRP-1 in the nematode Caenorhabditis elegans lengthens lifespan [53]. These findings intuitively suggest that long telomeres might be beneficial to lifespan extension. However, the effect of telomere length on the lifespan regulation remains controversial. Telomere length is stably maintained during replicative aging of yeast cells [54]. The yeast cells that had overlong telomeres displayed a regular RLS [23], while a truncated tlc1 mutant strain that had shorter but stable telomeres exhibited longer RLS than the wild-type strain [55]. Interestingly, the increase of telomere length in mouse tissues of spleen, colon and liver appears to be associated with chronic inflammation accelerated aging [56]. Long telomeres also associated with increased risk for pulmonary hypertension [57]. Nevertheless, how dysregulated telomerase activity influences yeast CLS is still unknown. In this work, we have employed the fusion protein Cdc13-Est2 to force constitutive association of telomerase with telomeres, and explore the effect of constitutive telomerase activity on CLS. Our results suggest that constitutively active telomerase results in genome instability, which accelerates yeast chronological aging.
Results
Genetic strategy to obtain the cells with overlong telomeres
In the budding yeast S. cerevisiae, telomeres are usually 350 ± 75 bp long. The telomere length is mainly determined by two opposite activities: telomerase-mediated elongation and nuclease-mediated degradation. In order to obtain the strains that have overlong telomeres, we employed the CDC13-EST2 fusion gene, whose expression can force enhanced association of telomerase at telomeres and produce long telomeres after successive passages [41]. We transformed either the CEN plasmid pRS316 (control) or the plasmid containing the CDC13-EST2 fusion gene, namely pRS316-CDC13-EST2, into the wild-type BY4742 cells. The individual transformants were passaged on selective plates every 72 hrs for thirteen times. The telomere length was examined by Southern blot using a telomeric TG-probe. The result showed that the ectopic expression of Cdc13-Est2 fusion protein led to gradual elongation of all telomeres (Fig. 1A). At the 5th and 13th streakout of transformants with CDC13-EST2 fusion gene, the size of the telomeric terminal-restriction-fragments (TRFs) digested by restriction endonuclease XhoI reached about 2.0 and 2.6 kb, respectively, which was 0.7 and 1.3 kb longer than that of the vector control, and 0.5 and 1.1 kb longer than that of the 1st streakout. The telomere TRFs longer than 2.0 kb were designated “overlong telomeres”. Because the TRFs contain about 0.95 kb subtelomeric sequence, and the telomeric TG1-3-tracts of wild-type cells are about 0.35 kb, the TG1-3-tracts of the Cdc13-Est2 expressing cells are approximately two (the 5th streakout) to four (the 13th streakout) folds longer.
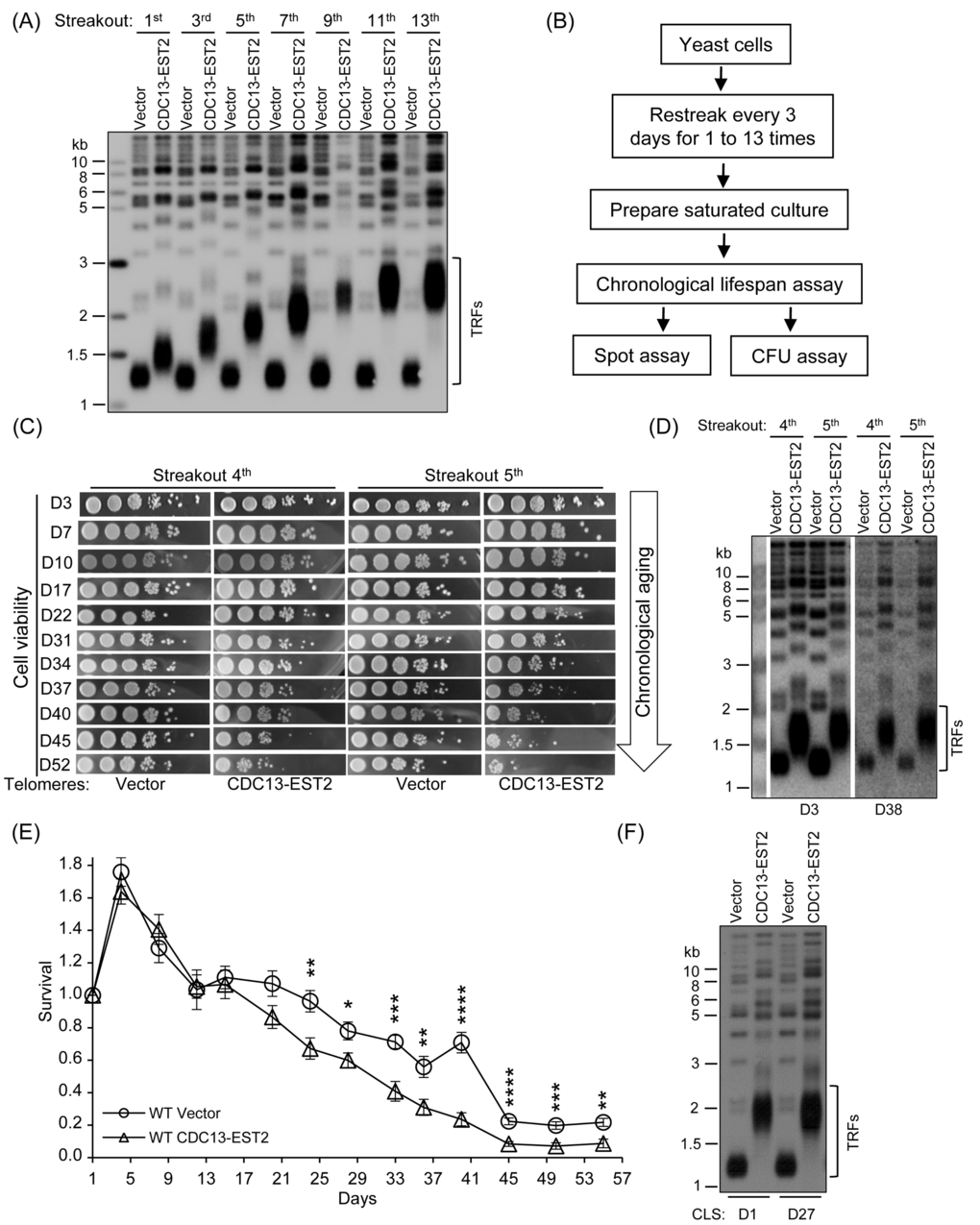
Figure 1. Expression of CDC13-EST2 promotes yeast chronological aging. (A) Telomere length analysis by Southern blot. The cells that contain pRS316 (Vector) or pRS316-CDC13-EST2 plasmid were passaged, and the genomic DNA from cells of different streakouts (labeled on top) was subjected to Southern blot assay using a telomeric TG1-3 probe. (B) Flow chart of CLS assay. (C) Spot assay of CLS. Both the normal-telomere and overlong-telomere cells of streakouts 4th and 5th were used to perform semi-quantitative CLS assay. The time of cultures in the CLS assay was labeled on the left (e.g. D3 means day 3). (D) Telomere length of cells used in (C) at D3 and D38 was examined by Southern blot with telomeric TG1-3 probe. (E) Colony formation unit (CFU) assay of CLS. CLS of cells BY4742-pRS316 (streakout 8th, normal telomeres) and BY4742 -pRS316-CDC13-EST2 (streakout 8th, overlong telomeres) was quantitatively examined. Survival (viable colonies) values at the indicated days of culture were normalized to CLS D1. Values are the averages of 6-10 cultures ± SEM. * denotes p<0.05, ** p<0.01, *** p<0.001 and **** p<0.0001. (F) Telomere length of cells in (E) at CLS D1 and CLS D27 was examined by Southern blot.
Expression of Cdc13-Est2 promotes yeast chronological aging
To evaluate the effect of Cdc13-Est2 expression on CLS, we randomly chose multiple single colonies to perform CLS assay. The procedures for a CLS experiment were schematically shown in Fig. 1B (see Materials and Methods for the details) [58,59]. The results showed that along the extension of culture time (day 3 to day 52), the viability of the cells with Cdc13-Est2 expression dropped much faster than that of the cells without Cdc13-Est2 expression (Fig. 1C). Because the CLS assay involved many steps, as well as many days of culturing (Fig. 1B and C), there was a possibility that telomere length had changed in the long process of culture. Therefore, we examined the telomere length in the cells of day 3 and day 38 samples by Southern blot (D3 and D38 in Fig. 1D). The results showed that the telomeres in the cells of day 38 had not undergone lengthening or shortening (Fig. 1D), and the telomeres in the Cdc13-Est2-expressing cells were substantially longer than that in the cells containing the control-vector. These data suggested that overlong telomeres induced by Cdc13-Est2 expression led to shorter CLS.
To quantify the survival rate of the aging cells, we carried out the colony formation assay (see the Materials and methods for the details) [7,59]. Multiple single colonies of the 8th streakout were used to perform the CLS assay. The survival rate of the cells expressing CDC13-EST2 fusion gene was significantly lower than that of the cells containing the control vector (Fig. 1E). The telomere length in the tested cells did not change during the long-time culture (e.g. 27 days, D27 in Fig. 1F). These results consistently supported the notion that Cdc13-Est2 expression results in overlong telomeres and promotes yeast chronological aging.
Expression of Cdc13-Est2 enhances ERC accumulation
Genome instability is one of the major regulators of CLS. Constitutively active telomerase at telomeres might have led to genome instability that shortens CLS. To test this hypothesis, we firstly examined the recombination activity of rDNA loci because the production of extra-chromosomal rDNA circles (ERCs) is an indication of rDNA stability [27]. The fob1Δ and sir2Δ cells were used as the negative and positive controls, respectively (see Materials and Methods for detail). The Southern blot results showed that deletion of FOB1 and SIR2 reduced and increased the ERCs, respectively (Fig. 2A). Six to ten independent cultures of both the young (CLS D1) and old (CLS D27) cells were subjected to ERC detection (Fig. 2B and C). The ERCs level in the CDC13-EST2 expressing cells is significantly higher than that in the vector-control cells (Fig. 2B and C). One species of multimer ERCs (indicated by solid arrow) in either the young (CLS D1) or old CDC13-EST2 expressing cells (CLS D27) was approximately 60% higher than that in the vector-control cells (Fig. 2B and C). The enhanced ERCs levels in young (CLS D1) and old (CLS D27) cells expressing CDC13-EST2 were comparable, we speculated that ERCs accumulation might take place in the proliferation stage of the culture (Fig. 2B and C). These results suggested that Cdc13-Est2 expression perturbed genome stability.
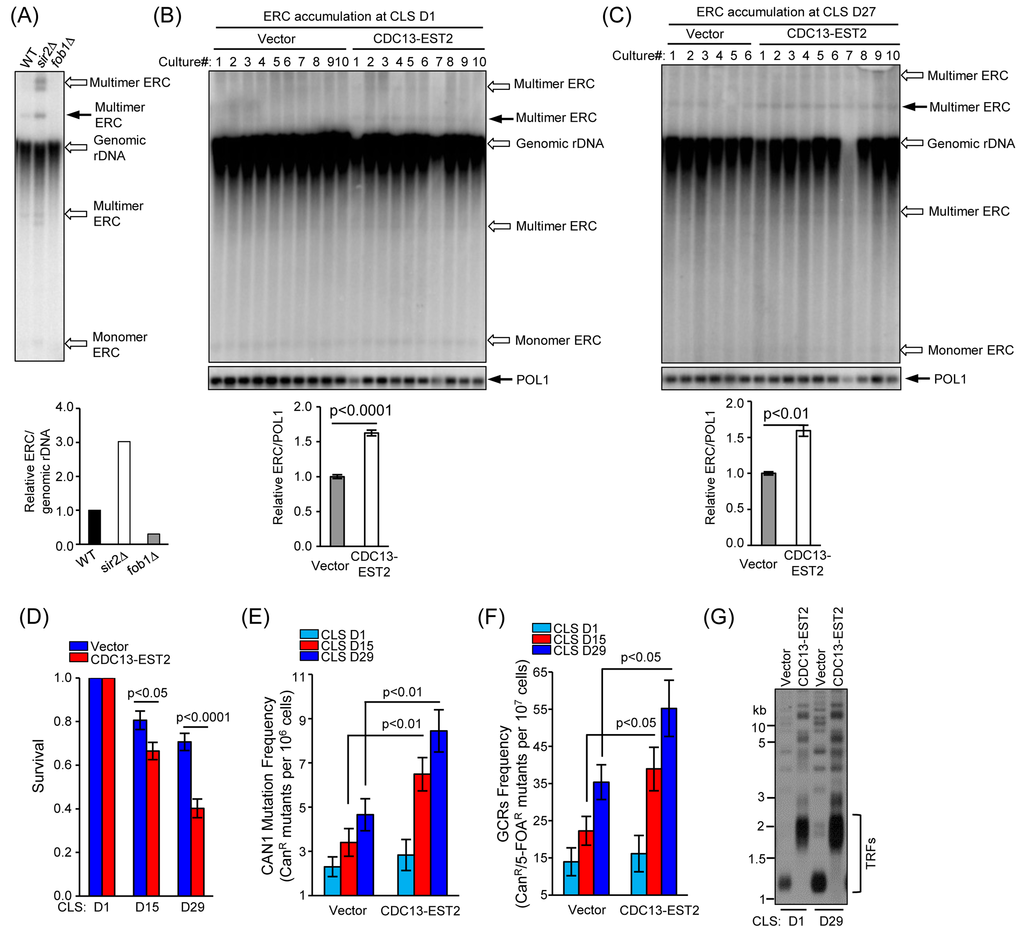
Figure 2. CDC13-EST2 expression enhances genome instability. (A) ERC level in fob1∆ and sir2∆ mutant cells was examined by Southern blot with a probe of 25S rDNA sequence (upper panel). One species of multimer ERCs (indicated by solid arrow) was quantified and normalized to genomic rDNA signal. Lower panel is the quantification of the ERC level in fob1∆ and sir2∆ mutant cells. The ERC level in wild-type cells was set as “1”, and the ERC value in each strain was normalized to the genomic rDNA. (B) and (C) Detection of ERC level in young cells of CLS D1 (B) and old cells of CLS D27 (C) used in Fig. 1E by Southern blot (upper panel). One species of multimer ERC (indicated by solid arrow) was quantified (lower panel). POL1 gene was used as an internal loading control. Values in the quantification are normalized to POL1 level ± SEM. (D) Chronological viability of cells carrying CAN1-URA3 cassette at CLS D1, D15 and D29. The cells of streakout 8th containing pRS315 (control) and pRS315-CDC13-EST2 was examined. Survival (viable colonies) values were normalized to CLS D1. Values are the averages of 6-10 cultures ± SEM. (E) and (F) CAN1 mutation frequency (E) and GCRs frequency (F) in overlong- and normal-telomere cells at CLS D1, D15 and D29 were examined. Values in (E) and (F) are the averages of 6-10 cultures ±SEM. (G) Telomere length analysis of cells at CLS D1 and D29. The cells of streakout 8th that containing pRS315 (normal telomere) and pRS315-CDC13-EST2 (overlong telomere) were examined.
Expression of Cdc13-Est2 increases age-dependent CAN1 marker-gene mutation frequency and GCR frequency
In order to validate further our hypothesis that expression of Cdc13-Est2 promotes yeast chronological aging by increasing genome instability, we next examined the CAN1 marker-gene mutation frequency and gross chromosomal rearrangement (GCR) frequency in the CDC13-EST2 expressing cells. These two assays have been widely used to evaluate age-dependent genome instability in yeast [8,22,60,61]. The CAN1 gene encodes an arginine permease, which controls the uptake of arginine by yeast cells. Detection of the CAN1 mutation frequency uses an arginine analogue, canavanine, which cells intake under arginine deficient conditions. Canavanine is toxic to yeast cells and leads to cell death. Unless CAN1 mutation (such as point mutation, insertion or deletion) inactivates CAN1, cells cannot survive when canavanine is supplemented in the medium lack of arginine [60]. Different from the CAN1 marker-gene mutation frequency assay, the GCR assay detects the loss of both CAN1 and URA3 marker genes simultaneously [60]. To measure GCR, the CAN1 proximal to HXT13 gene was substituted by URA3 marker-gene. 5-fluoroorotic acid (5-FOA) was used to counter-select yeast cells expressing URA3. In the presence of both 5-FOA and canavanine, cells must lose the functions of both CAN1 and URA3 to survive. We firstly examined the survival rate of the cells that contained the CAN1-URA3 marker cassette. The results showed that CDC13-EST2 expressing cells had significantly shortened CLS compared with vector control cells at CLS D15 (p value <0.05, n=6-10) and CLS D29 (p value <0.0001, n=6-10) (Fig. 2D). Consistently, CDC13-EST2 expression significantly increased the CAN1 marker-gene mutation frequency at CLS D15 (p value <0.01, n=6-10) and CLS D29 (p value <0.01, n=6-10) (Fig. 2E), and the GCR frequency at CLS D15 (p value <0.05, n=6-10) and CLS D29 (p value <0.05, n=6-10) (Fig. 2F). The cells used to detect CAN1 mutation frequency and GCR frequency did have overlong telomeres, and their long telomeres were stably maintained during chronological aging (Fig. 2G). Altogether, our results indicated that Cdc13-Est2 expression caused genome instability, and accelerated chronological aging.
Deletion of SCH9 reduces genome instability and suppresses chronological aging mediated by expression of Cdc13-Est2
Previous study has shown that deletion of SCH9 in yeast significantly decreases genome instability during chronological aging [22]. Now that Cdc13-Est2 expression led to enhanced genome instability and resulted in accelerated chronological aging, could calorie restriction (CR) or deletion of SCH9 slow down the rate of chronological aging mediated by Cdc13-Est2 expression? To address this question, we examined whether reducing glucose concentration from 2% to 0.5%, which mimics moderate CR conditions, could restore the CLS of the CDC13-EST2 expressing cells. The results showed that moderate CR did not significantly affect telomere length (Fig. S1A), nor recover the CLS of CDC13-EST2 expressing cells (Fig. S1B and C). This result is consistent with previous report that buffering culture medium to pH 6.0 by citrate/phosphate acts similarly with moderate CR in lengthening CLS [62], and application of moderate CR to synthetic complete medium buffered to pH 6.0 with citrate/phosphate cannot further extend CLS [62].
We next examined the effect of deletion of SCH9 on chronological aging in CDC13-EST2 expressing cells. Deletion of SCH9 had little effect on telomere length in either vector-control or CDC13-EST2 expressing cells (Fig. 3A), but significantly extended CLS in both cells (Fig. 3B). This is consistent with published data that sch9∆ increases CLS [14]. In order to address whether the CLS recovery is attributed to the suppression of genome instability, we examined CAN1 marker-gene mutation frequency, GCR frequency and ERCs accumulation in the sch9Δ cells with CDC13-EST2 expression. The results showed that deletion of SCH9 significantly lowered both the CAN1 marker-gene mutation frequency and the age-dependent GCR frequency mediated by CDC13-EST2 expression during chronological aging (Fig. 4A and B). The cells in the genome instability assay did have overlong telomeres and the telomere length did not exhibit a significant change during chorological aging (e.g. CLS D29) (Fig. 4C). In addition, deletion of SCH9 also lowered ERC accumulation mediated by CDC13-EST2 expression at CLS D1 (p value < 9.2E-07, n=6) and CLS D15 (p value < 8E-05, n=6) (Fig. S2A and B). CDC13-EST2 expressing cells stably maintained overlong telomeres during chronological aging (Fig. S2C). These data suggested that deletion of SCH9 contributed to the maintenance of genome stability in Cdc13-Est2 expressing cells, and inhibition of genome instability by deletion of SCH9 slowed down chronological aging process promoted by constitutively active telomerase.
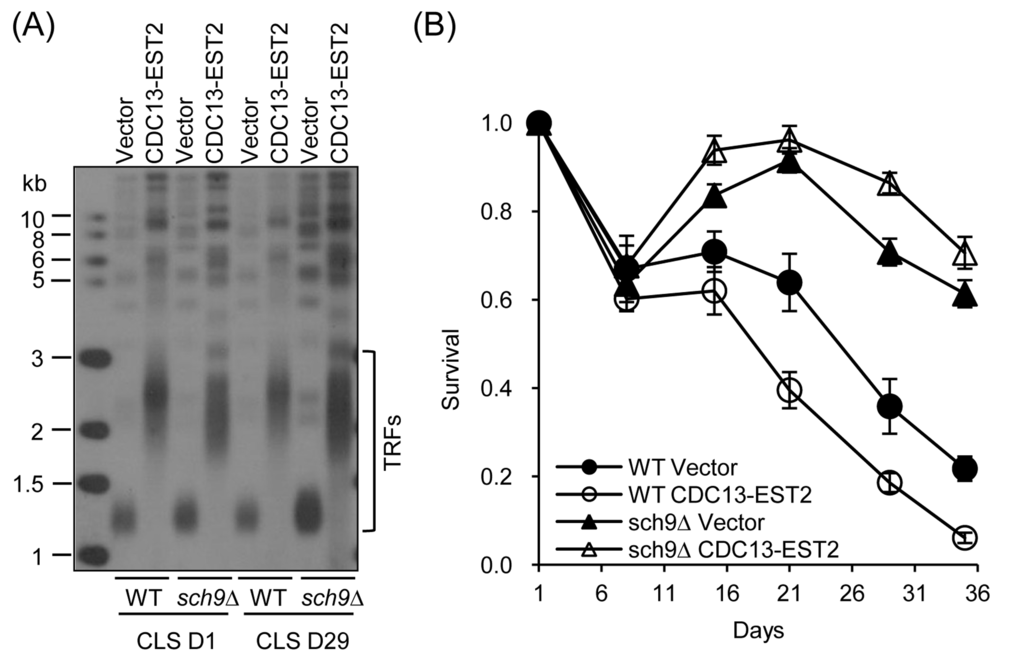
Figure 3. CLS of sch9∆ cells. (A) Telomere length of cells in (B) at CLS D1 and CLS D29 was examined by Southern blot. (B) CLS assay was done using cells WT-pRS316/sch9∆-pRS316 (streakout 13th, normal telomeres), WT-pRS316-CDC13-EST2/sch9∆-pRS316-CDC13-EST2 (streakout 13th, overlong telomeres). Survival (viable colonies) values are normalized to CLS D1 and are the averages of 6-10 cultures ± SEM.
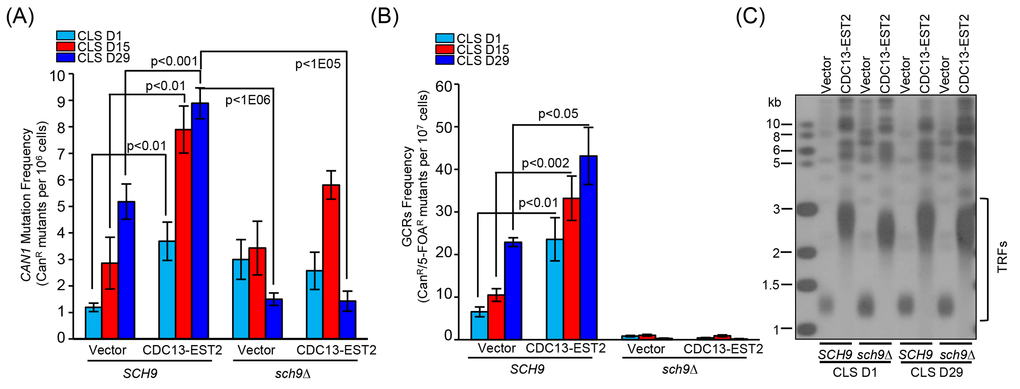
Figure 4. Detection of CAN1 mutation frequency and GCR frequency in sch9∆ cells during chronological aging. (A) Cells of hxt13∆-pRS315/ hxt13∆sch9∆-pRS315 (streakout 14th, normal telomeres), hxt13∆-pRS315-CDC13-EST2/ hxt13∆sch9∆-pRS315-CDC13-EST2 (streakout 14th, overlong telomeres) were used to assay CAN1 marker-gene mutation frequency during chronological aging. (B) The same cells in (A) were used to examine GCRs frequency during chronological aging. The values (viable colonies) are the averages of 6-10 cultures ± SEM. (C) Telomere length of cells of CLS D1 and D29 in (A) and (B) was examined by Southern blot.
Loss of CDC13-EST2 fusion gene restores regular CLS
We used a genetic strategy to force enhanced association of telomerase at telomeres through expression of CDC13-EST2. The expression of Cdc13-Est2 fusion protein resulted in overlong telomeres and shortened CLS (Fig. 1). However, it was unclear whether the shortened CLS was attributed to overlong telomeres or constitutive association of telomerase at telomeres. To discriminate these two possibilities, the pRS316 (control) and pRS316-CDC13-EST2 plasmids in the normal- and overlong-telomere cells were evicted, and CLS assays were performed (See Fig. S3 and S4 for the details). The results showed that the CLS of cells with overlong telomeres was not different from that of cells with normal telomeres (Fig. 5A and S4C). The cells used in the CLS assays maintained either normal or overlong telomeres during their long-term chronological aging (Fig. 5B and Fig. S4D). These results suggested that it was not the overlong telomeres, but rather the constitutive association of telomerase at telomeres that affected CLS. Following the notion that overlong telomeres might not affect CLS, we examined CLS of the rif1∆ mutant cells that have overlong telomeres (Fig.5D) [42,43]. The results showed that the rif1∆ mutant cells exhibited nearly the same chronological aging profile as the wild-type cells (Fig. 5C). The telomere length of wild-type and rif1∆ cells was stably maintained during chronological aging (Fig. 5D). Thus, constitutively tethering telomerase to telomeres causes genome instability and promotes chronological aging.
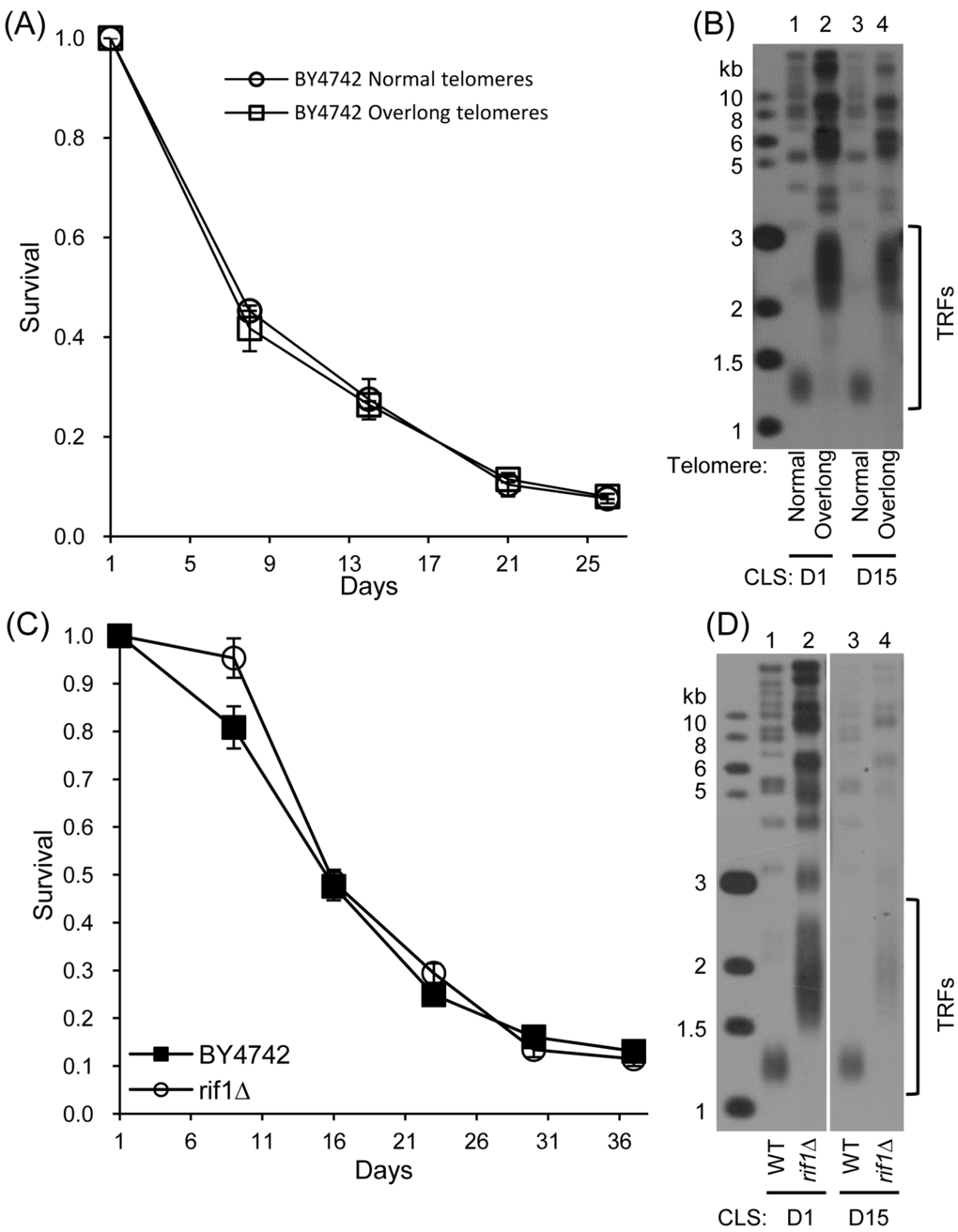
Figure 5. CLS assay of cells with overlong telomeres but no CDC13-EST2 fusion gene (A) CLS of normal- and overlong-telomere cells after plasmids eviction (21st streakout) (B) Southern blot analysis of telomere length using telomeric TG1-3 probe. (C) CLS of wild-type (4th streakout) and rif1∆ (4th streakout). (D) Telomere length detection by Southern blot at CLS D1 and D15.
Discussion
Previous studies have indicated that the replicative aging and the chronological aging are quite different [4,63]. However, numerous lines of evidence have pointed to a consensus that genome instability is one of the major culprits of aging (Fig.6) [1]. Telomeres reside at the end of the chromosomes, and their structural integrity is essential for genome stability [37,64]. Telomerase mediated telomere elongation usually takes place in the late S-phase of the cell cycle, and plays important roles in both telomere length and structure regulation [65,66]. Intact telomeres are required for genome stability (Fig. 6). In the absence of telomerase, following every cell division telomere attrition occurs because of the end replication problem [44,48,67]. When telomeres become critically short, cells could either undergo senescence [45], or repair telomeres through homologous recombination that promote replicative aging (Fig. 6) [23]. Consistently, long telomeres potentiate longer RLS in human somatic cells that have inactive telomerase [52,68]. Therefore, longer telomeres appear to be beneficial to lifespan extension when telomerase is not active. In the presence of telomerase, the telomere length and the RLS seems not to have consistent positive correlation. For examples, the mutant cells containing a truncated tlc1 allele have shortened telomeres but increased RLS, whereas rif1∆ mutant cells with long telomeres had shortened RLS [55]. Additionally, overlong telomeres induced by expressing CDC13-EST2 fusion gene do not affect RLS [23]. In the current work, we have evaluated the effect of overlong telomeres on yeast CLS. Our data suggest that overlong telomeres per se do not affect chronological aging (Fig. 5).
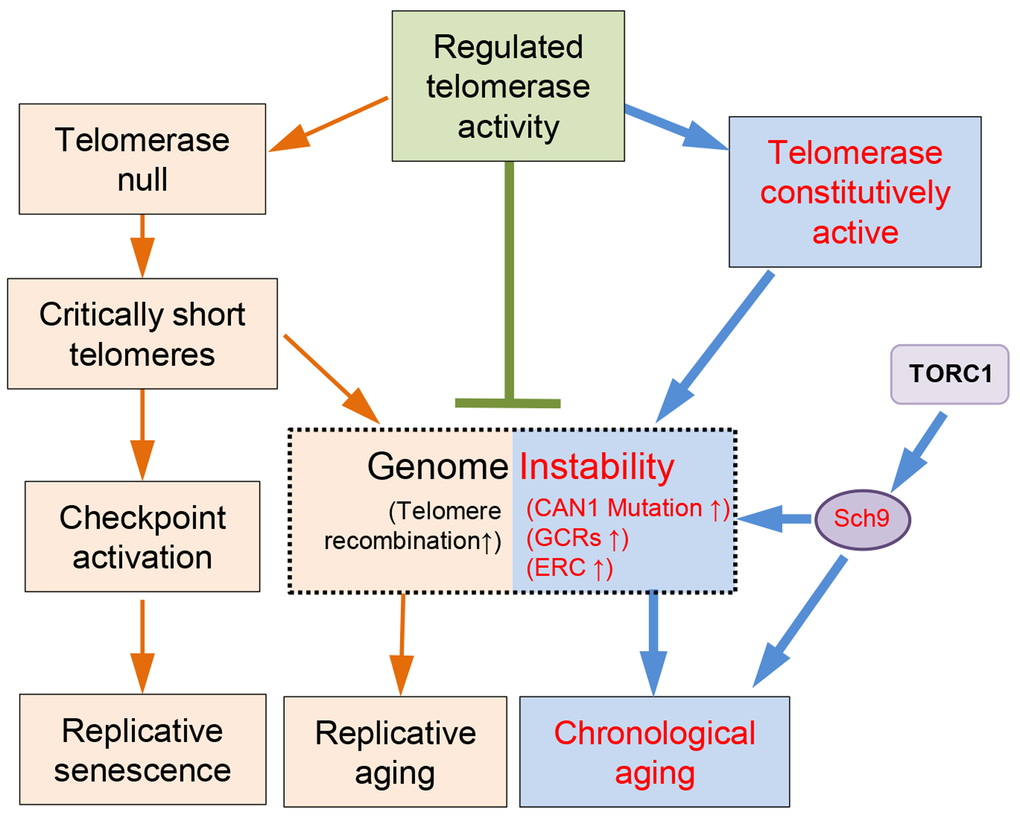
Figure 6. A model of telomere/telomerase associated genome instability affects yeast aging. Wild-type yeast cells have evolved to have regulated telomerase activity to maintain telomere homeostasis. In the absence of telomerase, telomeres gradually shrink to critically short ones that lead to either checkpoint activation and replicative senescence, or telomere recombination and accelerated replicative aging. When telomerase is constitutively active, the genome becomes less stable, and promotes chronological aging, which can be suppressed by down-regulation of TORC1/Sch9 pathway.
Interestingly, constitutively active telomerase mediated by expression of Cdc13-Est2 promotes chronological aging (Fig. 1). The underlying reason appears to be the enhanced genome instability (Fig. 6), exemplified by increased accumulation of ERCs (Fig. 2A-C), enhanced age-dependent spontaneous CAN1 marker-gene mutation frequency and GCR frequency in CDC13-EST2 expressing cells (Fig. 2D-G). How the enhanced association of telomerase at telomeres causes the effect on other genomic loci remains unknown. One possibility is that Cdc13-Est2 fusion protein not only targets to telomeric G-rich overlong, but also spontaneously binds other genomic loci where double strand breaks occur. This speculation is supported by the previous observation that Cdc13 can act on double strand breaks (DSBs) at low frequencies [69,70]. Under normal situation, Est2 binding to DSBs requires the interaction between Cdc13 and Est1 [71]. Cells seem to have evolved mechanisms that prevent the pervasive action of telomerase on DSBs. For example, Mec1 inhibits telomere healing by phosphorylating Cdc13 on its S306 residue, which suppresses Cdc13 accumulation at DSBs [72]. Est1 abundance is cell cycle regulated and peaks in S-phase, which may limit the activation of Cdc13-bound telomerase Est2 [65]. While under the scenario of Cdc13-Est2 fusion, wherever Cdc13 binds, Est2 may pose an effect on these loci. Telomerase mediated chromosomal healing on one hand repairs the DSBs, but on the other hand may endanger genome stability by loss of chromosomal arms.
The genome alterations observed in the CDC13-EST2 expressing cells are the accumulation of ERCs (Fig. 2B and C), enhanced spontaneous CAN1 mutation frequency and GCR frequency (Fig. 2E and F). These changes may exacerbate the chronological aging when the cells are challenged by nutrient depletion (Fig. 1), supporting the argument that genome instability is a direct causal factor for the accelerated chronological aging [1,22]. In agreement with this hypothesis, deletion of SCH9 did not shorten the overlong telomeres (Fig. 3A, 4C and S2C), but significantly suppressed the phenotypes of genome instability (Fig. 4A-B and Fig. S2A-B), and accordingly restored the lifespan of CDC13-ETS2 expressing cells (Fig. 3B). The accelerated chronological aging seen in CDC13-EST2 expressing cells is very similar to the case of sgs1Δ mutant cells. Deletion of SGS1 results in premature age-related changes including reduced CLS, elevated recombinant errors and age-dependent increase in DNA mutations [22]. Additionally, deletion of SCH9 in sgs1∆ mutant cells suppresses recombination and DNA damage, and slows down the chronological aging [22]. Thus, it is significant for the cells to regulate telomerase activity in a properly spatio-temporal manner to avoid genome instability and promote cell longevity.
Materials and Methods
Yeast strains and plasmids
All the yeast strains used in this study were derived from BY4742 (Euroscarf). The isogenic gene knockout mutant was constructed by one-step gene mutagenesis (Table S1) (See Supplemental information for the details).
Genomic DNA isolation and Southern blot analysis of yeast telomere length
Genomic DNA isolation and Southern blot analysis were done as what we did previously with minor modifications [73].
Chronological lifespan assay
Chronological lifespan assay was carried out as previously reported [7,59,62]. Glycerol stocks of yeast strains at -80°C freezer were streaked onto fresh YPD or appropriate selective plate and grown for 2-3 days at 30 °C. To measure chronological viability, 5-6 singe colonies were suspended into 5 ml of SDC-(amino acids auxotroph requirements) citrate/phosphate buffered medium with pH 6.0 for overnight. A calculated quantity of cells was diluted into 25 ml of SDC medium in a 100 ml flask to give an initial concentration of A600nm≈0.005. After 5-7 days of incubation at 30°C with constant shaking at 220 rpm, cells entered into stationary (G0) phase. 50 μl of chronologically aged culture was diluted for 2.5×105 folds for CLS D1. 100 μl of dilution was plated onto fresh YPD plate. Cell colony number was counted after 2-3 days of growth at 30 °C.
ERC level determination
ERC detection during chronological aging was performed as previously described [27]. The probe of 25S rDNA sequence (RDN25) (paired primers RDN25-Fwd and RDN25-Rev) for Southern blot detection of ERC was PCR amplified, and was radio-labeled with 32P-dCTP. The POL1 (YNL102W) was used as an internal loading control [23]. The DNA was separated in a 0.7% agarose gel under constant voltage of 60V (2V/cm) for 24 hours and transferred onto a positive charged nylon membrane (GE Healthcare, Cat#: RPN303B).
CAN1 marker-gene mutation frequency and GCR frequency detection
CAN1 marker-gene mutation and GCR frequency were assayed as previously reported with slight modifications [8,22,60]. (See Supplemental information for the details).
Statistical significance calculation
Statistical significance in this study was calculated by the two-tailed student’s t test in an Excel spreadsheet.
Supplementary Materials
Author Contributions
JL and JQZ developed the ideas and designed the study. JL performed most of experiments. MHH, JP, YMD, YSL, ZW, TG and HTL provided the reagents and carried out some of the experiments. JL and JQZ wrote the manuscript.
Acknowledgements
We thank Dr. V. Lundblad for plasmid expressing fusion gene CDC13-EST2.
Conflicts of Interest
There is no conflict of interest.
Funding
JL was funded by grants from China Postdoctoral Science Foundation (2014M551470), Postdoctoral Research Program of Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences (2014KIP305) and the National Natural Science Foundation of China (NSFC 31501110). The research in JQZ’s lab was supported by the grants from NSFC (31230040/31461143003/31521061) and from Ministry of Science and Technology of China (MoST 2013CB910400). This work was also supported in part by the Joint NSFC-ISF (Israel Science Foundation) Research Program (NSFC 31461143003).
References
- 1. López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013; 153:1194–217. https://doi.org/10.1016/j.cell.2013.05.039 [PubMed]
- 2. Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandez E, Miller RA. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009; 460:392–95. [PubMed]
- 3. Hurez V, Dao V, Liu A, Pandeswara S, Gelfond J, Sun L, Bergman M, Orihuela CJ, Galvan V, Padrón Á, Drerup J, Liu Y, Hasty P, et al. Chronic mTOR inhibition in mice with rapamycin alters T, B, myeloid, and innate lymphoid cells and gut flora and prolongs life of immune-deficient mice. Aging Cell. 2015; 14:945–56. https://doi.org/10.1111/acel.12380 [PubMed]
- 4. Longo VD, Shadel GS, Kaeberlein M, Kennedy B. Replicative and chronological aging in Saccharomyces cerevisiae. Cell Metab. 2012; 16:18–31. https://doi.org/10.1016/j.cmet.2012.06.002 [PubMed]
- 5. Mortimer RK, Johnston JR. Life span of individual yeast cells. Nature. 1959; 183:1751–52. https://doi.org/10.1038/1831751a0 [PubMed]
- 6. Fabrizio P, Longo VD. The chronological life span of Saccharomyces cerevisiae. Aging Cell. 2003; 2:73–81. https://doi.org/10.1046/j.1474-9728.2003.00033.x [PubMed]
- 7. Liu J, Huang X, Withers BR, Blalock E, Liu K, Dickson RC. Reducing sphingolipid synthesis orchestrates global changes to extend yeast lifespan. Aging Cell. 2013; 12:833–41. https://doi.org/10.1111/acel.12107 [PubMed]
- 8. Huang X, Liu J, Dickson RC. Down-regulating sphingolipid synthesis increases yeast lifespan. PLoS Genet. 2012; 8:e1002493. https://doi.org/10.1371/journal.pgen.1002493 [PubMed]
- 9. Kaeberlein M. Lessons on longevity from budding yeast. Nature. 2010; 464:513–19. https://doi.org/10.1038/nature08981 [PubMed]
- 10. Fabrizio P, Wei M. Conserved role of medium acidification in chronological senescence of yeast and mammalian cells. Aging (Albany NY). 2011; 3:1127–29. https://doi.org/10.18632/aging.100412 [PubMed]
- 11. Leontieva OV, Blagosklonny MV. Yeast-like chronological senescence in mammalian cells: phenomenon, mechanism and pharmacological suppression. Aging (Albany NY). 2011; 3:1078–91. https://doi.org/10.18632/aging.100402 [PubMed]
- 12. Kawałek A, Lefevre SD, Veenhuis M, van der Klei IJ. Peroxisomal catalase deficiency modulates yeast lifespan depending on growth conditions. Aging (Albany NY). 2013; 5:67–83. https://doi.org/10.18632/aging.100519 [PubMed]
- 13. Richard VR, Leonov A, Beach A, Burstein MT, Koupaki O, Gomez-Perez A, Levy S, Pluska L, Mattie S, Rafesh R, Iouk T, Sheibani S, Greenwood M, et al. Macromitophagy is a longevity assurance process that in chronologically aging yeast limited in calorie supply sustains functional mitochondria and maintains cellular lipid homeostasis. Aging (Albany NY). 2013; 5:234–69. https://doi.org/10.18632/aging.100547 [PubMed]
- 14. Fabrizio P, Pozza F, Pletcher SD, Gendron CM, Longo VD. Regulation of longevity and stress resistance by Sch9 in yeast. Science. 2001; 292:288–90. https://doi.org/10.1126/science.1059497 [PubMed]
- 15. Bjedov I, Toivonen JM, Kerr F, Slack C, Jacobson J, Foley A, Partridge L. Mechanisms of life span extension by rapamycin in the fruit fly Drosophila melanogaster. Cell Metab. 2010; 11:35–46. https://doi.org/10.1016/j.cmet.2009.11.010 [PubMed]
- 16. Fontana L, Partridge L, Longo VD. Extending healthy life span--from yeast to humans. Science. 2010; 328:321–26. https://doi.org/10.1126/science.1172539 [PubMed]
- 17. Lerner C, Bitto A, Pulliam D, Nacarelli T, Konigsberg M, Van Remmen H, Torres C, Sell C. Reduced mammalian target of rapamycin activity facilitates mitochondrial retrograde signaling and increases life span in normal human fibroblasts. Aging Cell. 2013; 12:966–77. https://doi.org/10.1111/acel.12122 [PubMed]
- 18. Miller RA, Harrison DE, Astle CM, Fernandez E, Flurkey K, Han M, Javors MA, Li X, Nadon NL, Nelson JF, Pletcher S, Salmon AB, Sharp ZD, et al. Rapamycin-mediated lifespan increase in mice is dose and sex dependent and metabolically distinct from dietary restriction. Aging Cell. 2014; 13:468–77. https://doi.org/10.1111/acel.12194 [PubMed]
- 19. Urban J, Soulard A, Huber A, Lippman S, Mukhopadhyay D, Deloche O, Wanke V, Anrather D, Ammerer G, Riezman H, Broach JR, De Virgilio C, Hall MN, Loewith R. Sch9 is a major target of TORC1 in Saccharomyces cerevisiae. Mol Cell. 2007; 26:663–74. https://doi.org/10.1016/j.molcel.2007.04.020 [PubMed]
- 20. Kaeberlein M, Powers RW
3rd , Steffen KK, Westman EA, Hu D, Dang N, Kerr EO, Kirkland KT, Fields S, Kennedy BK. Regulation of yeast replicative life span by TOR and Sch9 in response to nutrients. Science. 2005; 310:1193–96. https://doi.org/10.1126/science.1115535 [PubMed] - 21. Vijg J, Suh Y. Genome instability and aging. Annu Rev Physiol. 2013; 75:645–68. https://doi.org/10.1146/annurev-physiol-030212-183715 [PubMed]
- 22. Madia F, Gattazzo C, Wei M, Fabrizio P, Burhans WC, Weinberger M, Galbani A, Smith JR, Nguyen C, Huey S, Comai L, Longo VD. Longevity mutation in SCH9 prevents recombination errors and premature genomic instability in a Werner/Bloom model system. J Cell Biol. 2008; 180:67–81. https://doi.org/10.1083/jcb.200707154 [PubMed]
- 23. Peng J, He M-H, Duan Y-M, Liu Y-T, Zhou J-Q. Inhibition of telomere recombination by inactivation of KEOPS subunit Cgi121 promotes cell longevity. PLoS Genet. 2015; 11:e1005071. https://doi.org/10.1371/journal.pgen.1005071 [PubMed]
- 24. Chen X-F, Meng F-L, Zhou J-Q. Telomere recombination accelerates cellular aging in Saccharomyces cerevisiae.. PLoS Genet. 2009; 5:e1000535. https://doi.org/10.1371/journal.pgen.1000535 [PubMed]
- 25. Mostoslavsky R, Chua KF, Lombard DB, Pang WW, Fischer MR, Gellon L, Liu P, Mostoslavsky G, Franco S, Murphy MM, Mills KD, Patel P, Hsu JT, et al. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell. 2006; 124:315–29. https://doi.org/10.1016/j.cell.2005.11.044 [PubMed]
- 26. Lewinska A, Miedziak B, Kulak K, Molon M, Wnuk M. Links between nucleolar activity, rDNA stability, aneuploidy and chronological aging in the yeast Saccharomyces cerevisiae. Biogerontology. 2014; 15:289–316. https://doi.org/10.1007/s10522-014-9499-y [PubMed]
- 27. Sinclair DA, Guarente L. Extrachromosomal rDNA circles--a cause of aging in yeast. Cell. 1997; 91:1033–42. https://doi.org/10.1016/S0092-8674(00)80493-6 [PubMed]
- 28. Saka K, Ide S, Ganley AR, Kobayashi T. Cellular senescence in yeast is regulated by rDNA noncoding transcription. Curr Biol. 2013; 23:1794–98. https://doi.org/10.1016/j.cub.2013.07.048 [PubMed]
- 29. Kobayashi T, Horiuchi T, Tongaonkar P, Vu L, Nomura M. SIR2 regulates recombination between different rDNA repeats, but not recombination within individual rRNA genes in yeast. Cell. 2004; 117:441–53. https://doi.org/10.1016/S0092-8674(04)00414-3 [PubMed]
- 30. Heo S-J, Tatebayashi K, Ohsugi I, Shimamoto A, Furuichi Y, Ikeda H. Bloom’s syndrome gene suppresses premature ageing caused by Sgs1 deficiency in yeast. Genes Cells. 1999; 4:619–25. https://doi.org/10.1046/j.1365-2443.1999.00288.x [PubMed]
- 31. McVey M, Kaeberlein M, Tissenbaum HA, Guarente L. The short life span of Saccharomyces cerevisiae sgs1 and srs2 mutants is a composite of normal aging processes and mitotic arrest due to defective recombination. Genetics. 2001; 157:1531–42. [PubMed]
- 32. Merker RJ, Klein HL. hpr1Delta affects ribosomal DNA recombination and cell life span in Saccharomyces cerevisiae. Mol Cell Biol. 2002; 22:421–29. https://doi.org/10.1128/MCB.22.2.421-429.2002 [PubMed]
- 33. Klein HL, Symington LS. Sgs1--the maestro of recombination. Cell. 2012; 149:257–59. https://doi.org/10.1016/j.cell.2012.03.020 [PubMed]
- 34. Brosh RM
Jr , Bohr VA. Human premature aging, DNA repair and RecQ helicases. Nucleic Acids Res. 2007; 35:7527–44. https://doi.org/10.1093/nar/gkm1008 [PubMed] - 35. Kupiec M. Biology of telomeres: lessons from budding yeast. FEMS Microbiol Rev. 2014; 38:144–71. https://doi.org/10.1111/1574-6976.12054 [PubMed]
- 36. Wellinger RJ, Wolf AJ, Zakian VA. Saccharomyces telomeres acquire single-strand TG1-3 tails late in S phase. Cell. 1993; 72:51–60. https://doi.org/10.1016/0092-8674(93)90049-V [PubMed]
- 37. Taggart AK, Teng S-C, Zakian VA. Est1p as a cell cycle-regulated activator of telomere-bound telomerase. Science. 2002; 297:1023–26. https://doi.org/10.1126/science.1074968 [PubMed]
- 38. Wu Y, Zakian VA. The telomeric Cdc13 protein interacts directly with the telomerase subunit Est1 to bring it to telomeric DNA ends in vitro. Proc Natl Acad Sci USA. 2011; 108:20362–69. https://doi.org/10.1073/pnas.1100281108 [PubMed]
- 39. Qi H, Zakian VA. The Saccharomyces telomere-binding protein Cdc13p interacts with both the catalytic subunit of DNA polymerase α and the telomerase-associated est1 protein. Genes Dev. 2000; 14:1777–88. [PubMed]
- 40. Pennock E, Buckley K, Lundblad V. Cdc13 delivers separate complexes to the telomere for end protection and replication. Cell. 2001; 104:387–96. https://doi.org/10.1016/S0092-8674(01)00226-4 [PubMed]
- 41. Evans SK, Lundblad V. Est1 and Cdc13 as comediators of telomerase access. Science. 1999; 286:117–20. https://doi.org/10.1126/science.286.5437.117 [PubMed]
- 42. Hardy CF, Sussel L, Shore D. A RAP1-interacting protein involved in transcriptional silencing and telomere length regulation. Genes Dev. 1992; 6:801–14. https://doi.org/10.1101/gad.6.5.801 [PubMed]
- 43. Wotton D, Shore D. A novel Rap1p-interacting factor, Rif2p, cooperates with Rif1p to regulate telomere length in Saccharomyces cerevisiae. Genes Dev. 1997; 11:748–60. https://doi.org/10.1101/gad.11.6.748 [PubMed]
- 44. Soudet J, Jolivet P, Teixeira MT. Elucidation of the DNA end-replication problem in Saccharomyces cerevisiae. Mol Cell. 2014; 53:954–64. https://doi.org/10.1016/j.molcel.2014.02.030 [PubMed]
- 45. Lundblad V, Szostak JW. A mutant with a defect in telomere elongation leads to senescence in yeast. Cell. 1989; 57:633–43. https://doi.org/10.1016/0092-8674(89)90132-3 [PubMed]
- 46. Blasco MA, Lee H-W, Hande MP, Samper E, Lansdorp PM, DePinho RA, Greider CW. Telomere shortening and tumor formation by mouse cells lacking telomerase RNA. Cell. 1997; 91:25–34. https://doi.org/10.1016/S0092-8674(01)80006-4 [PubMed]
- 47. Weng NP, Levine BL, June CH, Hodes RJ. Human naive and memory T lymphocytes differ in telomeric length and replicative potential. Proc Natl Acad Sci USA. 1995; 92:11091–94. https://doi.org/10.1073/pnas.92.24.11091 [PubMed]
- 48. Chang E, Harley CB. Telomere length and replicative aging in human vascular tissues. Proc Natl Acad Sci USA. 1995; 92:11190–94. https://doi.org/10.1073/pnas.92.24.11190 [PubMed]
- 49. Bernardes de Jesus B, Schneeberger K, Vera E, Tejera A, Harley CB, Blasco MA. The telomerase activator TA-65 elongates short telomeres and increases health span of adult/old mice without increasing cancer incidence. Aging Cell. 2011; 10:604–21. https://doi.org/10.1111/j.1474-9726.2011.00700.x [PubMed]
- 50. Bernardes de Jesus B, Vera E, Schneeberger K, Tejera AM, Ayuso E, Bosch F, Blasco MA. Telomerase gene therapy in adult and old mice delays aging and increases longevity without increasing cancer. EMBO Mol Med. 2012; 4:691–704. https://doi.org/10.1002/emmm.201200245 [PubMed]
- 51. Codd V, Nelson CP, Albrecht E, Mangino M, Deelen J, Buxton JL, Hottenga JJ, Fischer K, Esko T, Surakka I, Broer L, Nyholt DR, Mateo Leach I, et al, and CARDIoGRAM consortium. Identification of seven loci affecting mean telomere length and their association with disease. Nat Genet. 2013; 45:422–27, e1–2. https://doi.org/10.1038/ng.2528 [PubMed]
- 52. Rubio MA, Kim S-H, Campisi J. Reversible manipulation of telomerase expression and telomere length. Implications for the ionizing radiation response and replicative senescence of human cells. J Biol Chem. 2002; 277:28609–17. https://doi.org/10.1074/jbc.M203747200 [PubMed]
- 53. Joeng KS, Song EJ, Lee K-J, Lee J. Long lifespan in worms with long telomeric DNA. Nat Genet. 2004; 36:607–11. https://doi.org/10.1038/ng1356 [PubMed]
- 54. Smeal T, Claus J, Kennedy B, Cole F, Guarente L. Loss of transcriptional silencing causes sterility in old mother cells of S. cerevisiae. Cell. 1996; 84:633–42. https://doi.org/10.1016/S0092-8674(00)81038-7 [PubMed]
- 55. Austriaco NR
Jr , Guarente LP. Changes of telomere length cause reciprocal changes in the lifespan of mother cells in Saccharomyces cerevisiae. Proc Natl Acad Sci USA. 1997; 94:9768–72. https://doi.org/10.1073/pnas.94.18.9768 [PubMed] - 56. Jurk D, Wilson C, Passos JF, Oakley F, Correia-Melo C, Greaves L, Saretzki G, Fox C, Lawless C, Anderson R, Hewitt G, Pender SL, Fullard N, et al. Chronic inflammation induces telomere dysfunction and accelerates ageing in mice. Nat Commun. 2014; 2:4172. https://doi.org/10.1038/ncomms5172 [PubMed]
- 57. Izikki M, Hoang E, Draskovic I, Mercier O, Lecerf F, Lamrani L, Liu W-Y, Guignabert C, Fadel E, Dorfmuller P, Humbert M, Londoño-Vallejo A, Eddahibi S. Telomere Maintenance Is a Critical Determinant in the Physiopathology of Pulmonary Hypertension. J Am Coll Cardiol. 2015; 66:1942–43. https://doi.org/10.1016/j.jacc.2015.08.869 [PubMed]
- 58. Burtner CR, Murakami CJ, Kennedy BK, Kaeberlein M. A molecular mechanism of chronological aging in yeast. Cell Cycle. 2009; 8:1256–70. https://doi.org/10.4161/cc.8.8.8287 [PubMed]
- 59. Fabrizio P, Longo V. (2007). The Chronological Life Span of Saccharomyces cerevisiae. Biological Aging: Humana Press), pp. 89-95.
- 60. Wei M, Madia F, Longo VD. Studying age-dependent genomic instability using the S. cerevisiae chronological lifespan model. J Vis Exp. 2011e3030. [PubMed]
- 61. Huang X, Liu J, Withers BR, Samide AJ, Leggas M, Dickson RC. Reducing signs of aging and increasing lifespan by drug synergy. Aging Cell. 2013; 12:652–60. https://doi.org/10.1111/acel.12090 [PubMed]
- 62. Murakami CJ, Wall V, Basisty N, Kaeberlein M. Composition and acidification of the culture medium influences chronological aging similarly in vineyard and laboratory yeast. PLoS One. 2011; 6:e24530. https://doi.org/10.1371/journal.pone.0024530 [PubMed]
- 63. Lin S-J, Austriaco N. Aging and cell death in the other yeasts, Schizosaccharomyces pombe and Candida albicans. FEMS Yeast Res. 2014; 14:119–35. https://doi.org/10.1111/1567-1364.12113 [PubMed]
- 64. Feng J, Funk WD, Wang SS, Weinrich SL, Avilion AA, Chiu CP, Adams RR, Chang E, Allsopp RC, Yu J, al. The RNA component of human telomerase. Science. 1995; 269:1236–41. https://doi.org/10.1126/science.7544491 [PubMed]
- 65. Mitton-Fry RM, Anderson EM, Hughes TR, Lundblad V, Wuttke DS. Conserved structure for single-stranded telomeric DNA recognition. Science. 2002; 296:145–47. https://doi.org/10.1126/science.1068799 [PubMed]
- 66. Gravel S, Larrivée M, Labrecque P, Wellinger RJ. Yeast Ku as a regulator of chromosomal DNA end structure. Science. 1998; 280:741–44. https://doi.org/10.1126/science.280.5364.741 [PubMed]
- 67. Lingner J, Cooper JP, Cech TR. Telomerase and DNA end replication: no longer a lagging strand problem? Science. 1995; 269:1533–34. https://doi.org/10.1126/science.7545310 [PubMed]
- 68. Ramunas J, Yakubov E, Brady JJ, Corbel SY, Holbrook C, Brandt M, Stein J, Santiago JG, Cooke JP, Blau HM. Transient delivery of modified mRNA encoding TERT rapidly extends telomeres in human cells. FASEB J. 2015; 29:1930–39. https://doi.org/10.1096/fj.14-259531 [PubMed]
- 69. Stellwagen AE, Haimberger ZW, Veatch JR, Gottschling DE. Ku interacts with telomerase RNA to promote telomere addition at native and broken chromosome ends. Genes Dev. 2003; 17:2384–95. https://doi.org/10.1101/gad.1125903 [PubMed]
- 70. Bertuch AA, Lundblad V. Which end: dissecting Ku’s function at telomeres and double-strand breaks. Genes Dev. 2003; 17:2347–50. https://doi.org/10.1101/gad.1146603 [PubMed]
- 71. Bianchi A, Negrini S, Shore D. Delivery of yeast telomerase to a DNA break depends on the recruitment functions of Cdc13 and Est1. Mol Cell. 2004; 16:139–46. https://doi.org/10.1016/j.molcel.2004.09.009 [PubMed]
- 72. Zhang W, Durocher D. De novo telomere formation is suppressed by the Mec1-dependent inhibition of Cdc13 accumulation at DNA breaks. Genes Dev. 2010; 24:502–15. https://doi.org/10.1101/gad.1869110 [PubMed]
- 73. Meng FL, Hu Y, Shen N, Tong XJ, Wang J, Ding J, Zhou JQ. Sua5p a single-stranded telomeric DNA-binding protein facilitates telomere replication. EMBO J. 2009; 28:1466–78. https://doi.org/10.1038/emboj.2009.92 [PubMed]