



I. Circadian rhythms, well-being, and life span
Organisms on earth evolved to restrict their activity to the night or day, being nocturnal or diurnal, respectively. By developing an endogenous circadian (circa - about; dies - day) clock, which can be entrained to external stimuli, primarily light, animals and plants ensure that physiological processes are performed at the appropriate, optimal time of day or night [1]. Adaptation to external conditions through clock entrainment imparts a survival advantage, as the organism can predict environmental changes [1-3]. The clock core machinery is self sustained, so that in the absence of external cues, e.g., in constant darkness, the endogenous rhythms free-run, generating cycles of approximately but not exactly 24 hours.
In mammals, the circadian clock influences nearly all aspects of physiology and behavior, such as sleep-wake cycles, cardiovascular activity, endocrine system, body temperature, renal activity, physiology of the gastrointestinal tract, and hepatic metabolism [1,2]. Epidemiological studies indicate that myocardial infarction, pulmonary edema, hypertensive crises, and asthma and allergic rhinitis attacks, all peak at certain times during the day [4-6]. Disruption of circadian coordination in humans or animals is manifested by hormone imbalance, some aspects of disease, and reduced life span [2,7-12].For instance, psychological and sleep disorders [2] and cardiovascular diseases [13,14] can be associated with irregular or dysfunctional circadian clock. Disruption of circadian coordination can also accelerate cancer proneness and malignant growth in animals and humans, suggesting that the circadian clock controls tumor progression [8-10]. In addition, symptoms seen in jet lagged travelers, e.g., fatigue, disorientation, and insomnia, or in shift workers, e.g., altered hormone profiles and morbidity, result from the constant need to extend wakefulness or to repeatedly invert the normal sleep-wake cycle [10,15,16]. Also, chronic reversal of the external light-dark cycle at weekly intervals resulted in a significant decrease in the survival time of cardiomyopathic hamsters [7]. Importantly, circadian rhythms change with normal aging in animals and humans, including a shift in the phase and decrease in amplitude [15-18]. By using a more direct approach, it was shown thatlongevity was diminished in golden hamsters carrying a 20 h-period mutation, tau, raised in 24 h light-dark cycles [19]. On the contrary, life span was extended in aged animals given fetal suprachiasmatic implants that restore higher amplitude rhythms [19-21]. Altogether, it seems that circadian disruption is associated with multiple negative manifestations, whereas resetting of circadian rhythms could lead to increased longevity. These findings, although largely correlative, point to a critical role for the circadian clock in maintaining normal peripheral physiology.
II. The circadian clock
A. The location of the mammalian circadian clock. In mammals, the central circadian clock is located in the suprachiasmatic nuclei (SCN), a distinct bilateral group of cells located in the anterior hypothalamus in the brain. Similar clock oscillators have been found in many peripheral tissues, such as the liver, intestine, heart, adipose tissue, retina and in various regions of the brain [2,22-24]. The SCN clock is composed of multiple, intracellular circadian oscillators, which, when synchronized, generate coordinated circadian outputs that regulate overt rhythms [25-28]. SCN oscillation is not exactly 24 h and it is necessaryto entrain the circadian pacemaker each day to the external light-dark cycle to prevent drifting (or free-running) out of phase. Light perceived primarily by melanopsin-expressing retinal ganglion cells transmit signals to the SCN via the retinohypothalamic tract (RHT) [2,29,30]. As a result, vasoactive intestinal polypeptide (VIP), an intrinsic SCN factor, acutely activates and synchronizes SCN neurons [31,32]. Synchronization among SCN neurons leads to the sending of signals to peripheral oscillators to prevent the dampening of circadian rhythms in these tissues. The SCN accomplishes this task via neuronal connections or circulating humoral factors [33] although the mechanisms are not fully understood (Figure 1). Several humoral factors expressed cyclically by the SCN, such as transforming growth factor α (TGFα) [34], prokineticin 2 (PK2) [35], and cardiotrophin-like cytokine (CLC) [36], have been shown to affect peripheral clocks. Their intracerebroventricular injection inhibits nocturnal locomotor activity, an SCN output. Complete electrical destruction of SCN neurons abolishes overall circadian rhythmicity in SCN-controlled tissues, because of the loss of synchrony among individual cells in the periphery and damping of the rhythm at the population level [37]. However, at the cellular level each cell oscillates, but with a different phase [37,38]. The fraction of cyclically expressed transcripts in each peripheral tissue ranges between 5-20% of the total population and the vast majority of these genes are tissue-specific [24,39-47]. These findings emphasize the circadian control over a large portion of the transcriptomes in peripheral tissues. Considering the circadian gene expression in peripheral tissues, it is difficult to determine whether the SCN clock drives these rhythmic patterns directly or indirectly by driving rhythmic feeding, activity, and/or body temperature, which, in turn, contribute to rhythms in gene expression in the periphery. It has been shown that for a peripheral tissue, such as the liver, signals both from the SCN clock or the local endogenous clock may control rhythmic gene expression [48,49].
B. The biological clock at the molecular level. Genetic analysis of mutations affecting the clock in Neurospora, Drosophila, Cyanobacteria, Arabidopsis, and, recently, the mouse have paved the way for the identification of clock genes. In mammals, the clock is an intracellular mechanism sharing the same molecular components in SCN neurons and peripheral cells [3]. Generation of circadian rhythms is dependent on the concerted co-expression of specific clock genes. Transcriptional-translational feedback loops lie at the very heart of the core clock mechanism. Many clock gene products function as transcription factors, which possess PAS (PER, ARNT, SIM) and basic helix-loop-helix (bHLH) domains involved in protein-protein and protein-DNA interactions, respectively. These factors ultimately activate or repress their own expression and, thus, constitute self-sustained transcriptional feedback loops. Changes in concentration, subcellular localization, post-transcriptional microRNA regulation, posttranslational modifications (phosphorylation, acetylation, deacetylation, SUMOylation), and delays between transcription and translation are crucial in order to achieve a 24-h cycle [1,2,50-52].
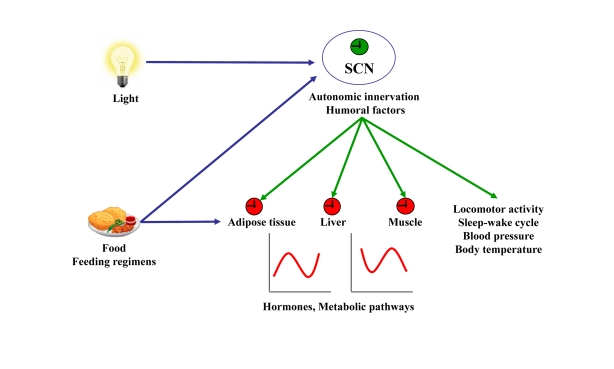
Figure 1. Resetting signals of the central and peripheral clocks.
The SCN resets peripheral oscillators via
humoral factors or autonomic innervation leading to circadian hormone
expression and secretion and rhythmic activity of metabolic pathways. In
addition, the SCN dictates rhythms of locomotor activity, sleep-wake cycle,
blood pressure, and body temperature. Light, food, and feeding regimens
affect either the central clock in the SCN or peripheral clocks. Input to
central or peripheral clocks are in blue. Outputs from the central clock to
the periphery are in green.
In the mouse, the first clock gene identified, encodes the transcription factor CLOCK (Circadian Locomotor Output Cycles Kaput) [53], which dimerizes with BMAL1 (brain and muscle ARNT-like protein 1) to activate transcription. CLOCK and BMAL1, two bHLH-PAS transcription factors, are capable of activating transcription upon binding to E-box (5'- CACGTG -3') and E-box-like promoter sequences [2]. BMAL1 can also dimerize with other CLOCK homologs, such as neuronal PAS domain protein 2 (NPAS2), to activate transcription and sustain rhythmicity [54,55]. Amongst the regulatory targets of CLOCK:BMAL1 are the three Period genes (Per1, Per2, and Per3), which encode PAS domain factors, and two Cryptochrome genes (Cry1 and Cry2). PERs and CRYs function as negative regulators, blocking CLOCK:BMAL1-mediated transcriptional activation [2,56] (Figure 2A).Thus, CLOCK:BMAL1 heterodimers bind to E-box sequences and mediate transcription of a large number of genes including those of the negative feedback loop Pers and Crys. When PERs and CRYs are produced in the cytoplasm, they oligomerize after reaching an appropriate concentrationand translocate to the nucleus to inhibit CLOCK:BMAL1-mediated transcription. All the aforementioned clock genes exhibit a 24-h oscillation in SCN cells and peripheral tissues, except for Clock that has been shown not to oscillate in the SCN [50]. Recent studies have demonstrated that CLOCK has intrinsic histone acetyltransferase activity, suggesting that rhythmic activation of chromatin remodeling may underlie the clock transcriptional network [57,58]. Indeed, cyclic histone acetylation and methylation have been observed on the promoters of several clock genes [58-63]. In addition, CLOCK also acetylates several proteins of the core clock apparatus, thus, enabling cycles of acetylation and deacetylation, the latter activity involving SIRT1 will be discussed below (Figure 2).
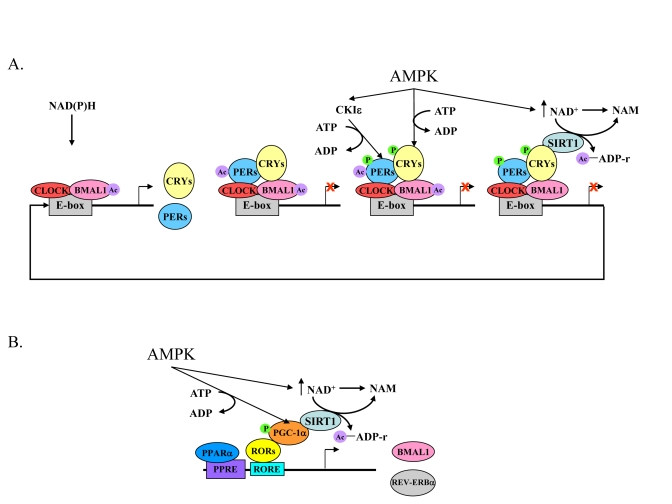
Figure 2. The core mechanism of the mammalian circadian clock and its link to energy metabolism.
(A) High NAD(P)H levels promote
CLOCK:BMAL1 binding to E-box sequences leading to the acetylation of BMAL1
and expression of Pers, Crys, and other clock-controlled genes. The
negative feedback loop, PERs:CRYs, binds to CLOCK:BMAL1 and consequently
PERs are acetylated. Activated AMPK leads to a rise in NAD+ levels,
phosphorylation of CRYs, and phosphorylation of CKI?, which then phosphorylates
the PERs. As a result of increased NAD+ levels, SIRT1 deacetylates PERs and BMAL1.
This and the destabilization of phosphorylated PERs and CRYs relieves PERs:CRYs
repression and another cycle starts. (B) Expression of Bmal1 and Rev-erbα
genes are controlled by PPARα and binding of RORs to RORE sequences. RORs
need a co-activator, PGC-1α, which is phosphorylated by activated AMPK. In
parallel, AMPK activation leads to an increase in NAD+ levels, which, in turn
activate SIRT1. SIRT1 activation leads to PGC-1α deacetylation and activation.
Acetyl adenosine diphosphate ribose (Ac-ADP-r) and nicotinamide (NAM) are released
after deacetylation by SIRT1.
Several other players appear to be important to sustain clock function. Casein kinase I epsilon (CKIε) phosphorylates the PER proteins and, thereby, enhances their instability and degradation [50,64-66]. CKIε also phosphorylates and partially activates the transcription factor BMAL1 [67]. Bmal1 expression is nega�tively regulated by the transcription factor reverse erythroblastosis virus α (REV-ERBα) [68], which recruits histone deacetylase (HDAC) complexes [69]. Bmal1 expression is positively regulated by retinoic acid receptor-related orphan receptor α (RORα) and RORγ [70]via the ROR response element (RORE) [71]. Thus, Bmal1 oscillation is driven by a rhythmic change in RORE occupancy by RORs and REV-ERBα. This alternating occupancy occurs due to the robust rhythmic levels of REV-ERBα, a result of direct transcriptional activation of the Rev-erbα gene by the heterodimer CLOCK:BMAL1 [68]. Indeed, mice deficient in RORα or REV-ERBα have impaired circadian rhythms of locomotor activity and clock gene expression [68,70] (Figure 2B).
III. The biological clock and energy homeostasis
A. Circadian rhythms and metabolism
The circadian clock has been reported to regulate metabolism and energy homeostasis in peripheral tissues [72,73]. This is achieved by mediating the expression and/or activity of certain metabolic enzymes and transport systems [74,75] involved in metabolic pathways [76-80]. In addition, a large number of nuclear receptors involved in lipid and glucose metabolism has been found to exhibit circadian expression [81]. Many hormones involved in metabolism, such as insulin [76], glucagon [82], adiponectin [83], corticosterone [84], leptin, and ghrelin [85,86], have been shown to exhibit circadian oscillation. Leptin, an adipocyte-derived circulating hormone, acts at specific receptors in the hypothalamus to suppress appetite and increase catabolism. Leptin exhibits striking circadian patterns in both gene expression and protein secretion, with peaks during the sleep phase in humans [87]. Neither feeding time nor adrenalectomy affected the rhythmicity of leptin release. However, ablation of the SCN has been shown to eliminate leptin circadian rhythmicity in rodents, suggesting that the central circadian clock regulates leptin expression [88]. Receptors for leptin and ghrelin are present on SCN cells [89-91], so it is possible that these hormones bind directly to SCN neurons, similarly to their effect on the orexigenic neuropeptide Y (NPY) and agouti-related protein (AgRP) neurons. Indeed, exogenous leptin was reported to phase-advance rhythms of neuronal firing in rat SCN slices [92].Activation of ventromedial arcuate nucleus (vmARC) neurons by systemic administration of the ghrelin mimetic growth hormone-releasing peptide 6 combined with SCN tracing showed that vmARC neurons transmit feeding-related signals to the SCN [90]. This injection induced Fos in the vmARC and resulted in attenuation of light-induced phase delay in mice and light-induced Fos expression in the SCN in rats [93]. Administration of ghrelin in vitro to SCN slices or SCN explants from Per2::luc transgenic mice caused phase shifts in Per2::luc reporter gene expression. However, intra-peritoneal injection of ghrelin to wild type mice caused phase shifts only after 30 h of food deprivation, but not when the mice were fed ad libitum [94]. Thus, it seems that ghrelin and leptin may affect the SCN directly or through their effect on the ARC, which is then relayed to the SCN.
Experiments using cultured cells have suggested that the cellular redox state is capable of influencing rhythms [95]. CLOCKand itshomolog NPAS2 can bind efficiently to BMAL1 andconsequently to E-box sequences in the presence of reduced nicotinamide adenine dinucleotides (NADH and NADPH) (Figure 2A). On the other hand, the oxidized forms of the nicotinamide adenine dinucleotides (NAD+ and NADP+) inhibit DNA binding of CLOCK:BMAL1 or NPAS2:BMAL1 [95,96]. As the NAD(P)+/NAD(P)Hredox equilibrium depends on the metabolicstate of the cell, this ratio could dictate the binding of CLOCK/NPAS2:BMAL1 to E-boxes and result in phase-shifting of cyclic gene expression [74,95,96]. NAD+ is also required for the activation of SIRT1, a deacetylase involved in clock modulation, as will be discussed below.
B. The circadian clock and crucial metabolic factors are tightly linked
BMAL1 : Circadian clocks have been shown to be present in several fat tissues, including inguinal white adipose tissue, epididymal white adipose tissue, and brown adipose tissue [45,97,98]. Recent transcriptome studies revealed rhythmic expression of clock and adipokine genes, such as resistin, adiponectin, and visfatin, in visceral fat tissue [83]. Recent molecular studies established the involvement of BMAL1 activity in the control of adipogenesis and lipid metabolism in mature adipocytes. Embryonic fibroblasts from Bmal1-/- knockout mice failed to differentiate into adipocytes. Loss of BMAL1 expression led to a significant decrease in adipogenesis and gene expression of several key adipogenic/lipogenic factors. Furthermore, over-expression of BMAL1 in adipocytes increased lipid synthesis activity. Thus, BMAL1, a master regulator of circadian rhythms, plays important roles in the regulation of adipose differentiation and lipogenesis in mature adipocytes [99].
REV-ERBα : Another important candidate to link the circadian clock with lipid metabolism is REV-ERBα. This pro-adipogenic transcription factor, whose levels increase dramatically during adipocyte differentiation [100], exhibits striking diurnal variations in expression in murine adipose tissue [101] and rat liver [102]. During adipocyte differentiation, REV-ERBα acts downstream of the differentiation factor peroxisome proliferator receptor-γ (PPARγ) by facilitating gene expression of PPARγ target genes [103,104]. Ectopic REV-ERBα expression in 3T3L1 pre-adipocytes promotes their differentiation into mature adipocytes [103]. In addition to its role in lipid metabolism and adipocyte differentiation, REV-ERBα is a component of the core clock apparatus, as mentioned above (Figure 2B). It acts as a negative regulator of Bmal1 expression, and its encoding gene, Rev-erbα, is directly activated by the CLOCK:BMAL1 heterodimer [68].
PPARα : Peroxisome proliferator-activated receptor α (PPARα) is a member of the nuclear receptor family that plays a unique role at the intersection of circadian and lipid metabolic pathways. The CLOCK:BMAL heterodimer mediates the expression of PPARα, which subsequently binds to the peroxisome-proliferator response element (PPRE) and activates transcription of Bmal1 [105-107] (Figure 2B). PPARα also regulates transcription of genes involved in lipid and glucose metabolism upon binding of endogenous free fatty acids [108,109]. Thus the circadian rhythmicity of PPARα provides an example of a reciprocal link between circadian and lipid metabolic processes.
PPARγ coactivator (PGC-1α) : PGC-1α, a transcriptional co-activator that regulates energy metabolism, is rhythmically expressed in the liver and skeletal muscle of mice. PGC-1α stimulates the expression of Bmal1 and Rev-erbα, through co-activation of the ROR family of orphan nuclear receptors [110,111] (Figure 2B). Mice lacking PGC-1α show abnormal diurnal rhythms of activity, body temperature, and metabolic rate, due to aberrant expression of clock genes and those involved in energy metabolism. Analyses of PGC-1α-deficient fibroblasts and mice with liver-specific knockdown of PGC-1α indicate that it is required for cell-autonomous clock function [110].
AMP-activated protein kinase (AMPK) : AMPK could be another important link that integrates the circadian clock with metabolism. AMPK is a sensor of the energy status within cells, which upon activation acts to restore energy balance [112,113]. This is done in part by modulating NAD+ levels and SIRT1 activity [114,115]. AMPK has been found to directly phosphorylate Ser-389 of CKIε in Rat-1 fibroblasts, resulting in increased CKIε activity and degradation of mPER2 (Figure 2A). mPER2 degradation led to a phase advance in the circadian expression pattern of clock genes [116]. AMPK has also been shown to phosphorylate and destabilize mCRY1 in mouse fibroblasts, leading to altered circadian rhythms [117] (Figure 2A). In addition, the expression profile of clock-related genes, such as Per1 and Cry2 in skeletal muscle, as well as the diurnal shift in energy utilization, is impaired in AMPKγ3 subunit knockout mice in response to 5-amino-4-imidazole-carboxamide riboside (AICAR), an AMPK activator [118]. In addition to its intracellular role, AMPK is involved in whole body energy metabolism by regulating the response to feeding in the hypothalamus [112]. In this brain area, AMPK activation is inhibited by leptin and insulin, hormones which suppress feeding, whereas it is activated under starvation by ghrelin, a hormone primarily produced by the stomach that leads to increased food intake [119-122].
SIRT1 : Another protein recently found to link metabolism with the circadian clock is SIRT1. This is the mammalian ortholog of yeast Sir2, an NAD+-dependent histone deacetylase involved in transcriptional silencing and genome stability in yeast [123,124]. Sir2 or its ortholog enzymes are involved in life span extension and the response to caloric restriction in yeast, Caenorhabditis elegans, Drosophila [123,125], and mice [115,126]. The dependence on NAD+ as a cofactor for catalysis is thought to link SIRT1 activity to the energy state of the cell [127]. Non-histone substrates of SIRT1, as found in C2C12 myotubes, include regulatory molecules that modulate energy metabolism, such as PPARγ and PGC-1α [114], key factors that regulate the core molecular clock (Figure 2). Recent studies showthat SIRT1 interacts directly with CLOCK and deacetylates BMAL1 and PER2 in cultured fibroblasts [128,129] (Figure 2A). It seems that after binding to E-box, CLOCK and CBP/p300 acetylate histones H3 and H4 [57] and BMAL1 [130]. BMAL1 acetylation potentiates its binding by the repressive PER/CRY complex [130] and, as a result, PER2 is acetylated [128]. When acetylated, PER2 [128] and possibly BMAL1 [129] are more stable. SIRT1 then becomes activated and deacetylates BMAL1, PER2, and histones [131]. Deacetylated PER2 is further phosphorylated and degraded and a new cycle begins (Figure 2A). It has also been shown that the CLOCK:BMAL1 heterodimer regulates the circadian expression of NAMPT (nicotinamide phosphoribosyl-transferase), a rate-limiting enzyme in the NAD+ salvage pathway. SIRT1 is recruited to the Nampt promoter and contributes to the circadian synthesis of its own coenzyme [132]. Most recently, it has been shown that AMPK enhances SIRT1 activity by increasing cellular NAD+ levels, resulting in the deacetylation and modulation of the activity of downstream SIRT1 targets [114]. Thus, the levels of NAD+ together with the cycling of SIRT1 can determine the activity and robustness of clock gene transcription at least in cultured cells.
C. Clock mutations and metabolic disorders
The most compelling linkage between metabolic disorders and the circadian clock is demonstrated by the phenotypes of clock gene mutants and knockouts. Homozygous C57BL/6J ClockΔ19 mice, with a truncated exon 18 and deleted exon 19 of the Clock gene, have a greatly attenuated diurnal feeding rhythm, are hyperphagicand obese, and develop a metabolic syndrome of hyperleptinemia,hyperlipidemia, hepatic steatosis, and hyperglycemia [133]. Loss of circadian rhythms in ClockΔ19 mutant mice was accompanied by attenuated expression of hypothalamic peptides associated with energy balance, such as ghrelin and orexin [133]. Insulin administration caused significantlygreater hypoglycemia in ClockΔ19 mutant mice than in wildtype mice [134]. Increased insulin sensitivity was also seen in ClockΔ19 mutant mice of the BALB/c/CBA background that preserve rhythmicity in melatonin production [135]. In ClockΔ19 on an Jcl:ICR background, serum levels of triglyceride and free fatty acids were significantly lower than in wild type control mice, whereas total cholesterol and glucose, insulin, and leptin levels did not differ [136]. Unlike C57BL/6J ClockΔ19 mutant mice, Jcl:ICR ClockΔ19 mutantmice were not obese, had low or normal fasting plasmaglucose, low plasma free fatty acids, and normal plasma leptin. However, in Jcl:ICR ClockΔ19 mutant mice, high fat diet amplified the diurnal variation in glucose tolerance and insulin sensitivity, and obesity was attenuated through impaired dietary fat absorption [136]. Although the effects on metabolism were variable due to strain differences, the overall picture is that disruption of the clock gene leads to disruption of metabolic pathways.
Bmal1-/- knockout mice, similarly to C57BL/6J ClockΔ19mutant mice, exhibited suppressed diurnal variations in glucose and triglycerides as well as abolished gluconeogenesis. Liver-specific deletion of Bmal1 showed a direct effect of the liver clock on glucose metabolism, as exhibited by hypoglycemia during fasting, exaggerated glucose clearance, and loss of rhythmic expression of hepatic glucose regulatory genes [137]. Although recovery from insulin-induced hypoglycemia was impaired in C57BL/6J ClockΔ19 mutant and Bmal1-/- knockout mice, the counter-regulatory responses of corticosterone and glucagon were retained [134].
Mutation in another central clock gene, Per2 (mPer2-/- mice), exhibited no glucocorticoid rhythm even though the corticosterone response to hypoglycemia was intact. In addition, the diurnal feeding rhythm was absent in these mice. Although food consumption was similar during the light and dark periods on high fat diet, mPer2-/- mice developed significant obesity [138].
IV. Effect of feeding regimens on circadian rhythms
In addition to light, feeding regimens have been reported to affect the clocks in the SCN and/or the periphery.
A. Restricted feeding (RF)
RF limits the time and duration of food availability with no calorie reduction [3,74,139].Animals, which receive food ad libitum everyday at the same time for only a few hours, adjust to the feeding period within a few days [49] and can consume their daily food intake during that limited time [140,141]. Restricting food to a particular time of day has profound effects on the behavior and physiology of animals. Many physiological activities that are normally dictated by the master clock in the SCN are altered by RF, such as hepatic P450 activity, body temperature, locomotor activity, and heart rate [142-145]. 2-4 h before the meal, the animals display food anticipatory activity (FAA), which is typifiedby an increase in locomotor activity, body temperature, corticosterone secretion,gastrointestinal motility, and activity of digestive enzymes [140,146-148], all are known output systems of the biological clock. RF is dominant over the SCN and drives rhythms in arrhythmic and clock mutant mice and animals with lesioned SCN, regardless of the lighting conditions [142,143,148-151]. In most incidents, RF affects the core clock apparatus in peripheral tissues, such as liver (Figure 3), kidney, heart, and pancreas, with no effect on the central pacemaker in the SCN [3,74,139,143,150,152,153], causing uncoupling from the central pacemaker in the SCN. This suggests that nutritional regulation of clock oscillators in peripheral tissues may play a direct role in coordinating metabolic oscillations [154]. As soon as food availability returns to normal, the SCN clock, whosephase remains unaffected,resets the peripheral oscillators [152]. The location of this food-entrainable oscillator (FEO) has been elusive. Lesions in the dorsomedial hypothalamic nucleus (DMH) [155-158], the brain stem parabrachial nuclei (PBN) [156,159], and the core and shell regions of nucleus accumbens [160,161] revealed that these brain regions may be involved in FEO output, but they cannot fully account for the oscillation [162]. Neither vagal signals nor leptin are critical for the entrainment [163,164].CLOCK [165] or BMAL1 [166] and other clock genes [167] have been shown not to be necessary for food anticipatory activity. However, it has recently been demonstrated that mPer2 mutant mice did not exhibit wheel-running food anticipation [168,169]. Recently, the FEO was suggested to be localized, in part, in ghrelin-secreting cells in the stomach [170]. Clearly, the localization and nature of the FEO and the effect of RF on circadian rhythms warrants further study.
The effect of RF on ageing and longevity has never been studied. Interestingly, the survival of Glasgow osteosarcoma-inoculated mice was prolonged under an RF regimen during the light period compared to those under the dark period or those fed ad libitum [171]. Also, RF modified the expression of genes involved in carcinogenesis and tumor progression, such as c-myc and p53 [172]. It remains to be determined whether RF feeding affects life span.
B. Calorie restriction (CR)
CR, sometimes denoted dietary restriction (DR), refers to a dietary regimen low in calories without malnutrition, that restricts the daily amount of calories derived from carbohydrates, fats, or proteins usually to 60-75% of ad libitum-fed animals. CR extends the life span of diverse species, such as C. elegans, Drososphila, rodents [125,173,174], and recently monkeys [175]. CR in mice, rats, and monkeys prevents or delays the onset of major age-related diseases, such as cancer, diabetes, kidney disease, and cataracts [173,176]. In humans, long-termed CR results in sustained beneficial effects on major risk factors for atherosclerosis, type 2 diabetes, and inflammation [177]. The reduction of energy intake is considered to be the critical beneficial factor in the CR regimen [173]. Theories on how CR modulates aging and longevity abound, but the exact mechanism is still unclear [178]. For a longtime, the most prevalent explanation was related to the widely acceptable theory on aging, the Free Radical Theory. This theory attributes the aging process to the continuousaccumulation of oxidative damage in macromolecules generated by reactive oxygen species (ROS) produced in the mitochondria [179]. A later variation of this theory, the Oxidative Stress Theory, attributes the oxidative damage to the imbalance between preoxidant and antioxidant components, and CR was suggested to increase the resistance to oxidative stress [180]. Recently, this explanation was put into question, at least for rodents, as increasing oxidative stress by several genetic alterations increased aging-related diseases, such as cancer, but did not diminish life span [181-183]. ROS, primarily H2O2, have recently been suggested to promote aging as activators of the TOR (target of rapamycin) pathway [184]. This signaling pathway acts as a sensor of the nutritional and energetic state in the cell and transmits anabolic signals to regulate cell size, growth, and metabolism. Mammalian TOR (mTOR) could play an important role in the regulation of life span, as indicated by findings showing that CR attenuated mTOR signaling in several tissues in mice [185], and mice deficient of ribosomal protein S61 kinase 1, a central component in mTOR signaling, or mice treated with rapamycin, an inhibitor of the mTORC1 component, exhibited increased life span [186,187]. Interestingly, mTOR has also been recently linked to the circadian clock as a light-activated signaling cascade in the SCN of mice [188].
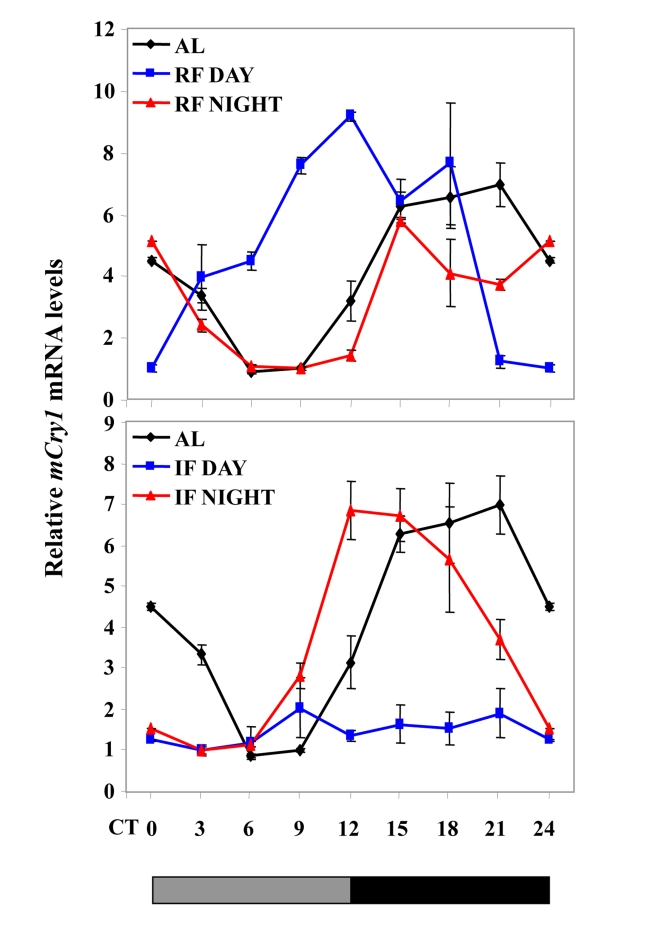
Figure 3. Effect of night vs. day RF and night vs. day IF on clock gene expression. Expression
of a representative clock gene mCry1 was measured in the liver of
C57BL mice during
ad libitum (AL), day and night RF, and day and night IF.Total
RNA extracted from liver tissue collected every 3 h around the circadian
cycle (mean ± SEM; n=3 for each time-point and each mouse group) was
reverse transcribed and analyzed by quantitative real time PCR. Clock gene levels
were normalized using Gapdh as the reference gene. The grey and
black bars designate the subjective light and dark cycles, respectively.
CR-fed animals resemble RF-treated animals, as they usually consume all or most of their food within a short period of time. While anticipating for food, calorically restricted animals show a rise in body temperature [189]. Thus, due to the temporal component of food intake, it is possible that CR, similarly to RF, synchronizes peripheral clocks and influences clock-controlled output systems, such as the anticipatory behavior and body temperature. As opposed to RF, CR entrains the clock in the SCN [190-192]. Under light-dark conditions and daytime feeding, calorically restricted mice showed strong FAA but with a phase advance of the nocturnal pattern of activity, a direct output of the SCN clock. When mice were transferred to dark-dark conditions, i.e. under free-running conditions, and fed ad libitum, the onset of the nocturnal period of locomotor activity occurred significantly earlier (1.3 h) in the calorically restricted than in ad libitum-fed animals, indicating an SCN effect. The period, however, did not differ between calorically restricted and ad libitum-fed mice [190]. Also, when SCN clock gene expression was tested, slight changes in gene expression were observed [191,192]. Overall, these results suggest that CR during the daytime affects the temporal organization of the SCN clockwork and circadian outputs in mice under light-dark cycle. In addition, CR affects photic responses of the circadian system, as measured by light pulses, suggesting that energy metabolism modulates gating of photic inputs in mammals [192]. It is noteworthy that microarray data comparing gene expression in seven different tissues under CR identified circadian rhythms among the three most over-expressed biological processes, with Per2 being the most up-regulated gene [193]. Collectively, these findings suggest that synchronization of peripheral oscillators during CR could be achieved directly due to the temporal feeding, as has been reported for RF [143,152,153], or by synchronizing the SCN [190-192], which, in turn, sends humoral or neuronal signals to entrain the peripheral tissues [194,195]. It is not known whether there is dominancy or harmony between the central pacemaker and peripheral oscillators under CR.
C. Intermittent fasting (IF)
During IF, also denoted alternate day fasting (ADF), food is available ad libitum every other day. IF-treated mice eat on the day they have access to food approximately twice as much as those having continuous access to food [196-198]. Similarly to calorically restricted animals, IF-fed animals exhibit increased life span in comparison with the ad libitum-fed control, even if there is little or no overall decrease in calories [199,200]. IF-fed animals also exhibit improved glucose metabolism, cardio-protection, neuro-protection [196,201-205], and increased resistance to cancer [197,200]. IF may also decrease the risk for cardiovascular diseases in humans [206].The IF-induced beneficial effects are thought to occur independently of the overall caloric intake, but the underlying mechanisms are still unknown. One suggested mechanism is stimulation of cellular stress pathways induced by the IF regimen [196,207,208]. Brain-derived neurotrophic factor (BDNF), normally involved in brain development and plasticity, is elevated in IF animals, and is causally linked to the protective effect of the IF regimen against neuronal damage inflicted by the neurotoxin kainic acid [209]. It must be noted, however, that BDNF could not be linked to the neuro-protective effects in the brain of calorically restricted rats [210,211], but increased levels of another neurotrophic factor, glial cell line-derived factor (GDNF), were correlated with neuro-protection of a calorically restricted primate model of Parkinson's disease [212]. Interestingly, BDNF is also a component of the hypothalamic melanocortin pathway that controls food intake and body weight in adult mice [213], and it has been implicated in the regulation of energy metabolism [214]. Heterozygousknockout BDNF (BDNF+/-) mice exhibit metabolic abnormalities, hyperphagia, obesity, and insulin resistance that could be significantly reversed by IF, indicating that BDNF is indeed involved in the beneficial effects induced by IF [214]. Interestingly,the BDNF+/- mice resemble circadian Clock mutant mice [133] in metabolic abnormalities. In addition, BDNF and its cognate receptor TrkB were suggested to play a role in circadian modulation of the SCN pacemaker sensitivity to light [215,216]. These data point to the possibility that IF could affect the SCN and, as a result, peripheral clocks, at least via elevating BDNF levels.
Recently, we have shown that, under an IF protocol,when food was introduced during the light period, mice exhibited almost arrhythmicity in clock gene expression in the liver. Unlike daytime feeding, nighttime feeding yielded rhythms similar to those generated during ad libitum feeding [198] (Figure 3). Furthermore, rhythms were maintained when daytime IF occurred under disruptive light, suggesting that SCN signals were involved in inducing the arrhythmic state in the periphery [198]. Thus, the fact that IF can affect circadian rhythms differently depending on the timing of food availability and light conditions suggests that this regimen affects the SCN clock, similarly to CR. We assume that SCN resetting by IF and CR could be involved in the health benefits conferred by these regimens [195].
The effects of IF are in contrast to those of restricted feeding (RF) that dictates peripheral rhythms in arrhythmic and mutant mice and animals with lesioned SCN regardless of the lighting conditions [142,143,148-151] (Figure 3). It, thus, appears that IF is not as dominant as RF in dictating peripheral rhythms. Never-theless, this feeding regimen exhibits some similarities with RF, as reflected by the anticipatory feeding behavior that preceded food availability and restoration of circadian rhythms under disruptive light conditions, due most likely to the effect on the food entrainable oscillator (FEO) [198]. Thus, under daytime IF, clock gene expression in the periphery would be controlled by the SCN, which responds to both light-dark cycle and IF, as well as directly by the temporal feeding via the FEO. Co-activation of both the FEO and the SCN would yield rhythms at two opposite phases leading to overall arrhythmicity. In contrast, under nighttime IF, normal rhythms are generated, as both the FEO and the SCN work in synchrony to dictate peripheral rhythms [198].
V. The circadian clock as a possible mediator in CR- or IF-induced increased longevity
A. Long-lived, spontaneously calorically restricted αMUPA mice
αMUPA mice carry as a transgene, the urokinase-type plasminogen activator (uPA) [217], an extracellular fibrinolytic serine protease implicated in tissue remodelling [218] and brain development and plasticity [219-224]. αMUPA mice spontaneously eat less (20-30%) compared to their wild type (WT) FVB/N control mice when fed ad libitum, indicating that their appetite is genetically suppressed. The mechanism linking transgenic uPA to reduced hunger is not yet clear. It could be related to uPA over-expression in the brain stem, as was found in two transgenic lines showing reduced food intake [225]. The transgenic effect is likely to be developmental, similarly to the remodelling effect recently detected in αMUPA developing incisor teeth [226]. αMUPA mice live longer (median, 16%; 10th decile, 15%) than WT mice [227], thus resembling calorically restricted animals in showing an inverse relation betweenfood intake and life span.αMUPA mice exhibit additional similarities with calorically restricted mice, such as reduced body weight, reduced levels of serum IGF-1 or glucose, enhanced capacity to conduct apoptosis, and reduced incidence of tumors[225,227-230].
B. αMUPA mice, circadian rhythms, and aging
Recent data show that αMUPA mice exhibit higher amplitude in the circadian expression of several clock genes in the liver compared with FVB/N WT mice. This change coincides with higher amplitude rhythms of food intake and body temperature [194]. Since circadian patterns of food intake and body temperature constituteclock-controlled output systems, it is conceivable that their alteration in the transgenic mice stems from the higher amplitude of clock gene expression in the periphery, and possibly also in the central biological clock in the SCN. Higher amplitude of circadian rhythms have been previously associated with young age [15] and extended life span [19]. Support for a linkage between circadian rhythms and attenuation of aging in αMUPA mice is provided by comparing young vs. old mice. When tested for circadian food intake, an SCN output system, 18-month-old WT control mice exhibit a 4-6-h shift in circadian food intake compared to 5-month-old mice [195]. This behaviour is consistent with literature data showing that aging can alter the amplitude and/or phase of circadian rhythms [15,16,18]. In contrast to WT mice, adult and young αMUPA mice show similar circadian food intake, indicating that at least some aspects of circadian behavior maintain a youthful pattern at an old age in these mice. At an old age, αMUPA mice maintain a young and healthier appearance, they look lean, and their fur is shiny, whereas WT mice are sluggish and they look old (Figure 4). In addition, αMUPA mice do not become obese throughout their life-time, whereas about one third of the WT mice show severe obesity (Figure 4). The major difference in body weight between αMUPA and WT mice stems from the fact that the quantitative difference in food intake between αMUPA and WT mice is maintained at the old age.
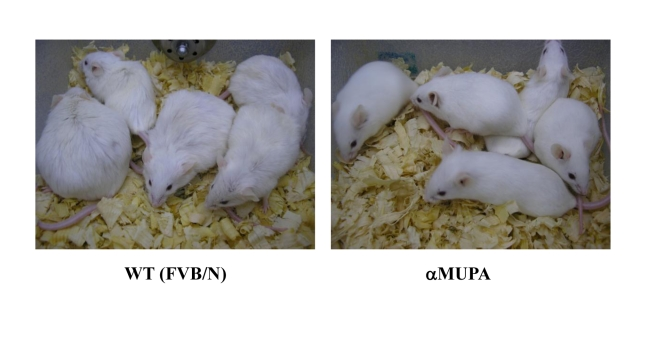
Figure 4. 18-month-old αMUPA and FVB/N WT mice. αMUPA
mice maintain a youthful and healthy appearance, whereas WT mice look old.
C. αMUPA mice reveal effectsof feeding regimens on circadian rhythms
It is difficult to eliminate the effect of temporal food consumption in calorically restricted animals, as mice consume their food within a few hours. αMUPA mice spontaneously consume reduced calories (20-30% reduction) compared with WT mice under different feeding regimens, i.e. AL, RF and IF, suggesting that these mice can be utilized as a model for CR in the absence of the imposed temporal food consumption under ad libitum feeding, and a model for imposed temporal CR under RF or IF conditions. Therefore, the transgenic mouse model αMUPA [225] has recently been used to investigate the contribution of calorie reduction per se vs. timed feeding to clock adaptation [49]. Under light-dark conditions and ad libitum feeding, αMUPA mice show high amplitude, appropriately reset circadian rhythms in peripheral clock genes [194] (Figure 5). This finding could reflect the effect of the reduced calorie intake on the SCN in αMUPA mice, as has been previously reported for calorically restricted animals [190-192].
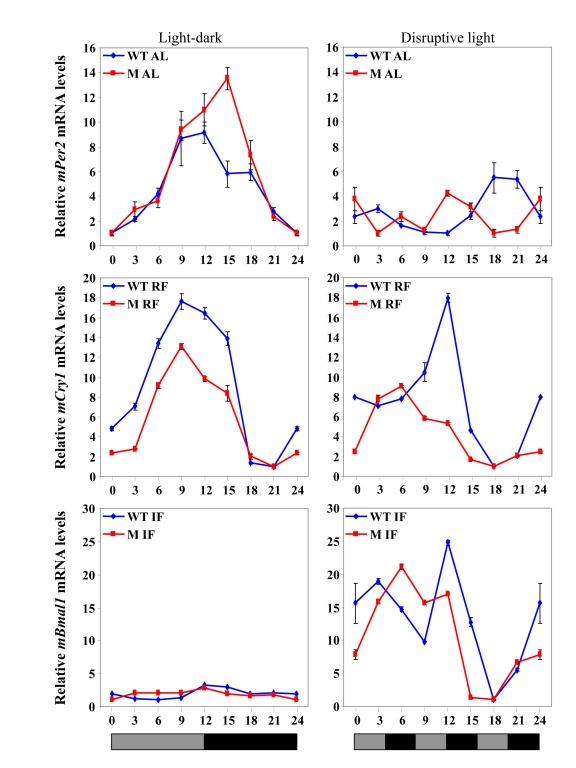
Figure 5. Clock gene expression in the liver under various feeding and lighting conditions in αMUPA (M) and WT mice. Expression levels
of the following clock genes are presented under light-dark or disruptive
light conditions:mPer2during ad
libitum (AL) feeding, mCry1 under RF, and Bmal1 under IF.Total
RNA extracted from liver tissue collected every 3 h around the circadian
cycle (mean ± SEM; n=3 for each time-point and each mouse group) was
reverse transcribed and analyzed by quantitative real time PCR. Clock gene levels
were normalized using Gapdh as the reference gene.
Under light-dark conditions, RF advanced the expression phase of all clock genes in the liver in a similar manner in both WT mice and αMUPA mice, and in some cases increased the amplitude [49](Figure 5). These findings were in concert with previous results in the literature, as is mentioned above. Nevertheless, an effect of the SCN on clock-controlled output systems under RF could be seen when the phases of peripheral clock gene expression in WT mice vs. αMUPA mice were compared under arrhythmicity imposed by disruptive light. Whereas the pattern of clock gene expression did not change in WT mice, a phase shift was seen in several genes in αMUPA mice (Figure 5), suggesting an SCN effect. Again, this effect of the SCN could be attributed to the reduced calorie intake of αMUPA mice. Altogether, these findings suggest that both the reduced calories and the light-dark cycle work in synchrony on the central biological clock of αMUPA mice to generate rhythms in the periphery. However, it seems that in WT mice, as has been found for other mouse strains, RF is dominant over the SCN in dictating rhythms in the periphery regardless of the lighting conditions. In αMUPA mice, RF dictates the phase of clock gene expression under light-dark; but under disruptive light conditions, as the SCN is under the influence of calorie restriction, it becomes dominant in dictating the phase of clock gene expression.
Unlike RF, the effect of IF on circadian rhythms in αMUPA mice was similar to that of WT mice under light-dark or disruptive light conditions, and it resembled also that on C57BL mice (Figure 3, Figure 5). Thus, in all mice tested, daytime IF caused arrhythmia in clock gene expression in the liver under light-dark, whereas rhythmicity was restored under disruptive light [198] (Figure 3). These observations suggest that IF, similarly to CR, may affect the SCN clock. This effect could possibly be mediated through a metabolic state generated by the day of fast during IF regardless of the calories consumed, as discussed earlier.
Altogether, the findings in αMUPA mice suggest that reduced calories affect the SCN so it becomes dominant over RF in the periphery only under disruptive light conditions. In addition, IF affects peripheral rhythms depending on the timing of food availability and light conditions, but regardless of the total daily calorie consumption, suggesting that this regimen induces a metabolic state that affects the SCN.
D. Temporal vs. quantitative food consumption and circadian rhythms
Previous publications have dealt with the issue of timed feeding and life span, reporting that calorically restricted mice showed increased longevity whether fed twice a day at daytime, once a day at daytime or nighttime, several times a day at nighttime [231,232], or three meals a week [233]. In these studies, the low-calorie feeding was practically timed and confined to the day or night, similarly to MUPA, or introduced in large intervals and continued throughout life time allowing appropriate adaptation. As a result, timed feeding was suggested to lead to high amplitude circadian rhythms and increased life span [232,234]. However, others rejected any contribution of timed feeding to CR-induced longevity [174,231]. The uncoupling of timed meals from reduced calories could practically be achieved only with animals, such as αMUPA, that spontaneously eat less. The results obtained with αMUPA indicate that temporal and quantitative aspects of food intake can be separately controlled. The timing of food intake is controlled by the central biological clock, whereas a separate mechanism appears to dictate the amount of food or calorie intake, that, in turn, could entrain the SCN clock, as has experimentally been shown for calorically restricted animals [191,192]. The results achieved with IF suggest that IF can be beneficial when food is given during the activity period of the animal, as explained above. Indeed, neuro- and cardio-protection alongside increased fatty acid oxidation and improved stress resistance have been induced after weeks of IF treatment when food was introduced at the beginning of the activity period [205,235-237]. It is noteworthy that cardio- and neuro-protection and life span extension were also seen when food was introduced during the day, but after many months of IF treatment [196,199], so that the animals could adjust after such a prolonged treatment. In light of these findings, we assume that the effect of IF on the SCN through a metabolic change, as mentioned above, alongside the timed feeding might affect the SCN to yield better-reset rhythms.
E. Differences between αMUPA mice and calorically restricted rodents
Although αMUPA mice exhibit reduced calorie intake and body fat, they show remarkable differences in energy metabolism compared with CR-treated animals. In particular, calorically restricted animals exhibit high levels of ghrelin [238,239], but low levels of leptin [240,241]and insulin [173], indicating an overall state of hunger. It is noteworthy that leptin-deficient animals are long-lived under CR feeding regimen, suggesting that leptin is not necessary for the CR-mediated benefits [242,243]. In addition, CR-treated mice exhibit high expression levels of PGC-1α and no change in PPARγ levels in the liver [244,245]. All these findings are in sharp contrast with those found in αMUPA mice that have low levels of ghrelin and high levels of leptin and insulin, suggesting that αMUPA mice eat less because their metabolism is of satiated rather than hungry animals (unpublished data). Nevertheless, one aspect found to be common to both αMUPA mice and calorically restricted animals are the low SIRT1 expression levels in the liver [246] and high levels in the brain [247]. It is noteworthy that the information regarding SIRT1 levels in the hypothalamus of calorically restricted animals is still lacking, and data for peripheral tissues is sometimes contradictory [124,246]. Results obtained with SIRT1-null micehave recently suggested that this enzyme could be required for the in vivo response to CR [248], and transgenic mice over-expressing SIRT1 show a phenotype resembling calorically restricted animals [249]. We assume that, in calorically restricted mice, SIRT1 activity could be elevated in the brain, possibly in the hypothalamus and SCN, through AMPK activation, as AMPK can be activated in the hypothalamus under starvation conditions and high ghrelin [112,121]. In αMUPA mice, that show low AMPK levels in the hypothalamus, the high leptin levels could lead to SIRT1 elevation, as leptin is required for the increase in SIRT1 protein levels in the hypothalamus under starvation [250]. Thus, although stimulated through different pathways in αMUPA and calorically restricted mice, SIRT1 could act as a common factor modulating the SCN clock and, as a result, longevity.
αMUPA mice share also some similarities with those of Lou/C rats, both obesity-resistant long-lived rodents.
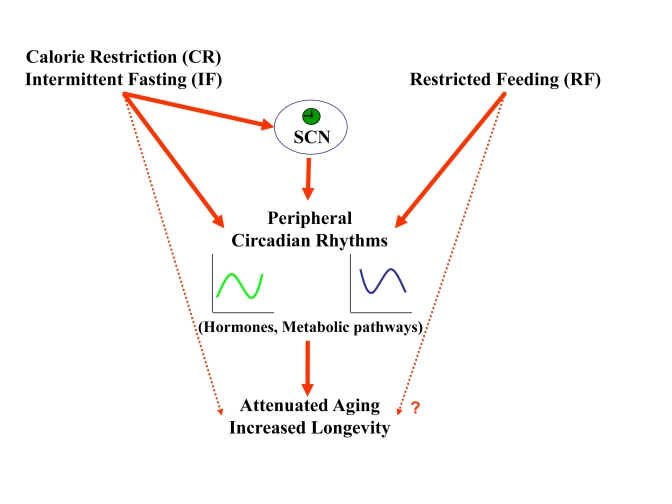
Figure 6. A schematic model describing the effect of feeding regimens on longevity through peripheral and SCN clock resetting. CR and IF reset
circadian rhythms in the periphery and the SCN. The synchronized, robust
circadian rhythms could be the mediator though which these feeding regimens
lead to aging attenuation and life span extension. RF resets circadian
rhythms only in the periphery, but its effect on life span is not known.
However, there are some fundamental differences in Lou/C rats compared with αMUPA mice, such as increased levels of PGC-1α and SIRT1 in the liver, increased levels of ghrelin, and reduced levels of leptin and insulin in the serum, although with some improved sensitivity for the latter two hormones [251,252].Overall, it seems that αMUPA mice are metabolically different from Lou/C rats.
F. Role of circadian rhythms in CR on health and longevity
The capacity of CR to reset the SCN clock, as previously reported [191,192],and the pronounced circadian rhythms seen in the long-lived αMUPA mice, pose the biological clock as a possible major factor determining longevity of calorically restricted mice [194,195]. The beneficial effect induced by CR on health and longevity can be achieved by appropriately resetting and synchronising a variety of hormonal, biochemical, and physiological functions. In turn, some of these functions can feedback to the biological clock in the periphery [143,153] and the SCN [191,192] and help sustain the rhythms. Indeed, the redox state affects the dimerization of the two clock proteins CLOCK and BMAL1 in vitro [96]. As SIRT1 has been linked to life span and suggested to mediate CR-induced effects [115,126,253], it could be a candidate that modulates the clock of calorically restricted animals, as discussed above. Clock resetting could lead to robust circadian rhythms that are associated with young ages and extended life span [15,16,19,195].
VI. Conclusions
RF entrains peripheral clocks due to temporal food consumption, whereas CR and IF appear to synchronize the central pacemaker in the SCN, suggesting a role for a metabolicstate imposed by low calories in central clock entrainment. In αMUPA mice, reduced calories alone were found not to be sufficient to sustain rhythms, unless feeding was spontaneously timed at night, or timed at day through a restricted feeding protocol. Therefore, it appears that when reduced calories are timed, as always occurs during CR and IF regimens, clock adjustment can influence a wide variety of output systems, so that cellular and physiological systems perform in a more synchronised and appropriately reset manner. We assume that SIRT1 could be a key mediator in clock synchronization at least under CR. Robust circadian rhythms can ensure a better tissue and body homeostasis, and could constitute an important mediator in aging attenuation and longevity extension (Figure 6).
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Panda S , Hogenesch JB and Kay SA. Circadian rhythms from flies to human. Nature. 2002; 417: 329 -335. [PubMed] .
- 2. Reppert SM and Weaver DR. Coordination of circadian timing in mammals. Nature. 2002; 418: 935 -941. [PubMed] .
- 3. Schibler U , Ripperger J and Brown SA. Peripheral circadian oscillators in mammals: time and food. J Biol Rhythms. 2003; 18: 250 -260. [PubMed] .
- 4. Maron BJ , Kogan J , Proschan MA , Hecht GM and Roberts WC. Circadian variability in the occurrence of sudden cardiac death in patients with hypertrophic cardiomyopathy. J Am Coll Cardiol. 1994; 23: 1405 -1409. [PubMed] .
- 5. Staels B When the Clock stops ticking, metabolic syndrome explodes. Nat Med. 2006; 12: 54 -55. [PubMed] .
- 6. Burioka N , Fukuoka Y , Takata M , Endo M , Miyata M , Chikumi H , Tomita K , Kodani M , Touge H , Takeda K , Sumikawa T , Yamaguchi K , Ueda Y , Nakazaki H , Suyama H , Yamasaki A , Sano H , Igishi T and Shimizu E. Circadian rhythms in the CNS and peripheral clock disorders: function of clock genes: influence of medication for bronchial asthma on circadian gene. J Pharmacol Sci. 2007; 103: 144 -149. [PubMed] .
- 7. Penev PD , Kolker DE , Zee PC and Turek FW. Chronic circadian desynchronization decreases the survival of animals with cardiomyopathic heart disease. Am J Physiol. 1998; 275: H2334 -H2337. [PubMed] .
- 8. Fu L , Pelicano H , Liu J , Huang P and Lee C. The circadian gene Period2 plays an important role in tumor suppression and DNA damage response in vivo. Cell. 2002; 111: 41 -50. [PubMed] .
- 9. Filipski E , King VM , Li X , Granda TG , Mormont MC , Claustrat B , Hastings MH and Levi F. Disruption of circadian coordination accelerates malignant growth in mice. Pathol Biol. 2003; 51: 216 -219. [PubMed] .
- 10. Davis S and Mirick DK. Circadian disruption, shift work and the risk of cancer: a summary of the evidence and studies in Seattle. Cancer Causes Control. 2006; 17: 539 -545. [PubMed] .
- 11. Kondratov RV , Kondratova AA , Gorbacheva VY , Vykhovanets OV and Antoch MP. Early aging and age-related pathologies in mice deficient in BMAL1, the core component of the circadian clock. Genes Dev. 2006; 20: 1868 -1873. [PubMed] .
- 12. Kondratov RV A role of the circadian system and circadian proteins in aging. Ageing Res Rev. 2007; 6: 12 -27. [PubMed] .
- 13. Montagnana M , Salvagno GL and Lippi G. Circadian variation within hemostasis: an underrecognized link between biology and disease. Semin Thromb Hemost. 2009; 35: 23 -33. [PubMed] .
- 14. Anea CB , Zhang M , Stepp DW , Simkins GB , Reed G , Fulton DJ and Rudic RD. Vascular disease in mice with a dysfunctional circadian clock. Circulation. 2009; 119: 1510 -1517. [PubMed] .
- 15. Hofman MA and Swaab DF. Living by the clock: the circadian pacemaker in older people. Ageing Res Rev. 2006; 5: 33 -51. [PubMed] .
- 16. Gibson EM , Williams WP 3rd and Kriegsfeld LJ. Aging in the circadian system: considerations for health, disease prevention and longevity. Exp Gerontol. 2009; 44: 51 -56. [PubMed] .
- 17. Scarbrough K , Losee-Olson S , Wallen EP and Turek FW. Aging and photoperiod affect entrainment and quantitative aspects of locomotor behavior in Syrian hamsters. Am J Physiol. 1997; 272: R1219 -R1225. [PubMed] .
- 18. Yamazaki S , Straume M , Tei H , Sakaki Y , Menaker M and Block GD. Effects of aging on central and peripheral mammalian clocks. Proc Natl Acad Sci U S A. 2002; 99: 10801 -10806. [PubMed] .
- 19. Hurd MW and Ralph MR. The significance of circadian organization for longevity in the golden hamster. J Biol Rhythms. 1998; 13: 430 -436. [PubMed] .
- 20. Hurd MW , Zimmer KA , Lehman MN and Ralph MR. Circadian locomotor rhythms in aged hamsters following suprachiasmatic transplant. Am J Physiol. 1995; 269: R958 -968. [PubMed] .
- 21. Li H and Satinoff E. Fetal tissue containing the suprachiasmatic nucleus restores multiple circadian rhythms in old rats. Am J Physiol. 1998; 275: R1735 -1744. [PubMed] .
- 22. Lee C , Etchegaray JP , Cagampang FR , Loudon AS and Reppert SM. Posttranslational mechanisms regulate the mammalian circadian clock. Cell. 2001; 107: 855 -867. [PubMed] .
- 23. Froy O and Chapnik N. Circadian oscillation of innate immunity components in mouse small intestine. Mol Immunol. 2007; 44: 1954 -1960. [PubMed] .
- 24. Young ME The circadian clock within the heart: potential influence on myocardial gene expression, metabolism, and function. Am J Physiol Heart Circ Physiol. 2006; 290: H1 -H16. [PubMed] .
- 25. Welsh DK , Logothetis DE , Meister M and Reppert SM. Individual neurons dissociated from rat suprachiasmatic nucleus express independently phased circadian firing rhythms. Neuron. 1995; 14: 697 -706. [PubMed] .
- 26. Liu C , Weaver DR , Strogatz SH and Reppert SM. Cellular construction of a circadian clock: period determination in the suprachiasmatic nuclei. Cell. 1997; 91: 855 -860. [PubMed] .
- 27. Herzog ED , Takahashi JS and Block GD. Clock controls circadian period in isolated suprachiasmatic nucleus neurons. Nat Neurosci. 1998; 1: 708 -713. [PubMed] .
- 28. Reppert SM and Weaver DR. Molecular analysis of mammalian circadian rhythms. Annu Rev Physiol. 2001; 63: 647 -676. [PubMed] .
- 29. Gooley JJ , Lu J , Chou TC , Scammell TE and Saper CB. Melanopsin in cells of origin of the retinohypothalamic tract. Nat Neurosci. 2001; 4: 1165 [PubMed] .
- 30. Lucas RJ , Freedman MS , Lupi D , Munoz M , David-Gray ZK and Foster RG. Identifying the photoreceptive inputs to the mammalian circadian system using transgenic and retinally degenerate mice. Behav Brain Res. 2001; 125: 97 -102. [PubMed] .
- 31. Harmar AJ , Marston HM , Shen S , Spratt C , West KM , Sheward WJ , Morrison CF , Dorin JR , Piggins HD , Reubi JC , Kelly JS , Maywood ES and Hastings MH. The VPAC(2) receptor is essential for circadian function in the mouse suprachiasmatic nuclei. Cell. 2002; 109: 497 -508. [PubMed] .
- 32. Maywood ES , Reddy AB , Wong GK , O'Neill JS , O'Brien JA , McMahon DG , Harmar AJ , Okamura H and Hastings MH. Synchronization and maintenance of timekeeping in suprachiasmatic circadian clock cells by neuropeptidergic signaling. Curr Biol. 2006; 16: 599 -605. [PubMed] .
- 33. Le Minh N , Damiola F , Tronche F , Schutz G and Schibler U. Glucocorticoid hormones inhibit food-induced phase-shifting of peripheral circadian oscillators. EMBO J. 2001; 20: 7128 -7136. [PubMed] .
- 34. Kramer A , Yang FC , Snodgrass P , Li X , Scammell TE , Davis FC and Weitz CJ. Regulation of daily locomotor activity and sleep by hypothalamic EGF receptor signaling. Science. 2001; 294: 2511 -2515. [PubMed] .
- 35. Cheng MY , Bullock CM , Li C , Lee AG , Bermak JC , Belluzzi J , Weaver DR , Leslie FM and Zhou QY. Prokineticin 2 transmits the behavioural circadian rhythm of the suprachiasmatic nucleus. Nature. 2002; 417: 405 -410. [PubMed] .
- 36. Kraves S and Weitz CJ. A role for cardiotrophin-like cytokine in the circadian control of mammalian locomotor activity. Nat Neurosci. 2006; 9: 212 -219. [PubMed] .
- 37. Yoo SH , Yamazaki S , Lowrey PL , Shimomura K , Ko CH , Buhr ED , Siepka SM , Hong HK , Oh WJ , Yoo OJ , Menaker M and Takahashi JS. PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc Natl Acad Sci U S A. 2004; 101: 5339 -5346. [PubMed] .
- 38. Welsh DK , Yoo SH , Liu AC , Takahashi JS and Kay SA. Bioluminescence imaging of individual fibroblasts reveals persistent, independently phased circadian rhythms of clock gene expression. Curr Biol. 2004; 14: 2289 -2295. [PubMed] .
- 39. Kornmann B , Preitner N , Rifat D , Fleury-Olela F and Schibler U. Analysis of circadian liver gene expression by ADDER, a highly sensitive method for the display of differentially expressed mRNAs. Nucleic Acids Res. 2001; 29: E51 [PubMed] .
- 40. Akhtar RA , Reddy AB , Maywood ES , Clayton JD , King VM , Smith AG , Gant TW , Hastings MH and Kyriacou CP. Circadian cycling of the mouse liver transcriptome, as revealed by cDNA microarray, is driven by the suprachiasmatic nucleus. Curr Biol. 2002; 12: 540 -550. [PubMed] .
- 41. Duffield GE , Best JD , Meurers BH , Bittner A , Loros JJ and Dunlap JC. Circadian programs of transcriptional activation, signaling, and protein turnover revealed by microarray analysis of mammalian cells. Curr Biol. 2002; 12: 551 -557. [PubMed] .
- 42. Panda S , Antoch MP , Miller BH , Su AI , Schook AB , Straume M , Schultz PG , Kay SA , Takahashi JS and Hogenesch JB. Coordinated transcription of key pathways in the mouse by the circadian clock. Cell. 2002; 109: 307 -320. [PubMed] .
- 43. Storch KF , Lipan O , Leykin I , Viswanathan N , Davis FC , Wong WH and Weitz CJ. Extensive and divergent circadian gene expression in liver and heart. Nature. 2002; 417: 78 -83. [PubMed] .
- 44. Kita Y , Shiozawa M , Jin W , Majewski RR , Besharse JC , Greene AS and Jacob HJ. Implications of circadian gene expression in kidney, liver and the effects of fasting on pharmacogenomic studies. Pharmacogenetics. 2002; 12: 55 -65. [PubMed] .
- 45. Zvonic S , Ptitsyn AA , Conrad SA , Scott LK , Floyd ZE , Kilroy G , Wu X , Goh BC , Mynatt RL and Gimble JM. Characterization of peripheral circadian clocks in adipose tissues. Diabetes. 2006; 55: 962 -970. [PubMed] .
- 46. Reddy AB , Karp NA , Maywood ES , Sage EA , Deery M , O'Neill JS , Wong GK , Chesham J , Odell M , Lilley KS , Kyriacou CP and Hastings MH. Circadian orchestration of the hepatic proteome. Curr Biol. 2006; 16: 1107 -1115. [PubMed] .
- 47. McCarthy JJ , Andrews JL , McDearmon EL , Campbell KS , Barber BK , Miller BH , Walker JR , Hogenesch JB , Takahashi JS and Esser KA. Identification of the circadian transcriptome in adult mouse skeletal muscle. Physiol Genomics. 2007; 31: 86 -95. [PubMed] .
- 48. Kornmann B , Schaad O , Bujard H , Takahashi JS and Schibler U. System-driven and oscillator-dependent circadian transcription in mice with a conditionally active liver clock. PLoS Biol. 2007; 5: e34 [PubMed] .
- 49. Froy O , Chapnik N and Miskin R. The suprachiasmatic nuclei are involved in determining circadian rhythms during restricted feeding. Neuroscience. 2008; 155: 1152 -1159. [PubMed] .
- 50. Dunlap JC Molecular bases for circadian clocks. Cell. 1999; 96: 271 -290. [PubMed] .
- 51. Cardone L , Hirayama J , Giordano F , Tamaru T , Palvimo JJ and Sassone-Corsi P. Circadian clock control by SUMOylation of BMAL1. Science. 2005; 309: 1390 -1394. [PubMed] .
- 52. Nagel R , Clijsters L and Agami R. The miRNA-192/194 cluster regulates the Period gene family and the circadian clock. Febs J. 2009; 276: 5447 -5455. [PubMed] .
- 53. Vitaterna MH , King DP , Chang AM , Kornhauser JM , Lowrey PL , McDonald JD , Dove WF , Pinto LH , Turek FW and Takahashi JS. Mutagenesis and mapping of a mouse gene, Clock, essential for circadian behavior. Science. 1994; 264: 719 -725. [PubMed] .
- 54. Asher G and Schibler U. A CLOCK-less clock. Trends Cell Biol. 2006; 16: 547 -549. [PubMed] .
- 55. Debruyne JP , Noton E , Lambert CM , Maywood ES , Weaver DR and Reppert SM. A clock shock: mouse CLOCK is not required for circadian oscillator function. Neuron. 2006; 50: 465 -477. [PubMed] .
- 56. Froy O , Chang DC and Reppert SM. Redox potential: differential roles in dCRY and mCRY1 functions. Curr Biol. 2002; 12: 147 -152. [PubMed] .
- 57. Doi M , Hirayama J and Sassone-Corsi P. Circadian regulator CLOCK is a histone acetyltransferase. Cell. 2006; 125: 497 -508. [PubMed] .
- 58. Nakahata Y , Grimaldi B , Sahar S , Hirayama J and Sassone-Corsi P. Signaling to the circadian clock: plasticity by chromatin remodeling. Curr Opin Cell Biol. 2007; 19: 230 -237. [PubMed] .
- 59. Etchegaray JP , Lee C , Wade PA and Reppert SM. Rhythmic histone acetylation underlies transcription in the mammalian circadian clock. Nature. 2003; 421: 177 -182. [PubMed] .
- 60. Curtis AM , Seo SB , Westgate EJ , Rudic RD , Smyth EM , Chakravarti D , FitzGerald GA and McNamara P. Histone acetyltransferase-dependent chromatin remodeling and the vascular clock. J Biol Chem. 2004; 279: 7091 -7097. [PubMed] .
- 61. Naruse Y , Oh-hashi K , Iijima N , Naruse M , Yoshioka H and Tanaka M. Circadian and light-induced transcription of clock gene Per1 depends on histone acetylation and deacetylation. Mol Cell Biol. 2004; 24: 6278 -6287. [PubMed] .
- 62. Ripperger JA and Schibler U. Rhythmic CLOCK-BMAL1 binding to multiple E-box motifs drives circadian Dbp transcription and chromatin transitions. Nat Genet. 2006; 38: 369 -374. [PubMed] .
- 63. Etchegaray JP , Yang X , DeBruyne JP , Peters AH , Weaver DR , Jenuwein T and Reppert SM. The polycomb group protein EZH2 is required for mammalian circadian clock function. J Biol Chem. 2006; 281: 21209 -21215. [PubMed] .
- 64. Whitmore D , Cermakian N , Crosio C , Foulkes NS , Pando MP , Travnickova Z and Sassone-Corsi P. A clockwork organ. Biol Chem. 2000; 381: 793 -800. [PubMed] .
- 65. Eide EJ and Virshup DM. Casein kinase I: another cog in the circadian clockworks. Chronobiol Int. 2001; 18: 389 -398. [PubMed] .
- 66. Eide EJ , Woolf MF , Kang H , Woolf P , Hurst W , Camacho F , Vielhaber EL , Giovanni A and Virshup DM. Control of mammalian circadian rhythm by CKIepsilon-regulated proteasome-mediated PER2 degradation. Mol Cell Biol. 2005; 25: 2795 -2807. [PubMed] .
- 67. Eide EJ , Kang H , Crapo S , Gallego M and Virshup DM. Casein kinase I in the mammalian circadian clock. Methods Enzymol. 2005; 393: 408 -418. [PubMed] .
- 68. Preitner N , Damiola F , Lopez-Molina L , Zakany J , Duboule D , Albrecht U and Schibler U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell. 2002; 110: 251 -260. [PubMed] .
- 69. Yin L and Lazar MA. The orphan nuclear receptor Rev-erbalpha recruits the N-CoR/histone deacetylase 3 corepressor to regulate the circadian Bmal1 gene. Mol Endocrinol. 2005; 19: 1452 -1459. [PubMed] .
- 70. Sato TK , Panda S , Miraglia LJ , Reyes TM , Rudic RD , McNamara P , Naik KA , FitzGerald GA , Kay SA and Hogenesch JB. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron. 2004; 43: 527 -537. [PubMed] .
- 71. Ueda HR , Hayashi S , Chen W , Sano M , Machida M , Shigeyoshi Y , Iino M and Hashimoto S. System-level identification of transcriptional circuits underlying mammalian circadian clocks. Nat Genet. 2005; 37: 187 -192. [PubMed] .
- 72. Froy O The relationship between nutrition and circadian rhythms in mammals. Front Neuroendocrinol. 2007; 28: 61 -71. [PubMed] .
- 73. Green CB , Takahashi JS and Bass J. The meter of metabolism. Cell. 2008; 134: 728 -742. [PubMed] .
- 74. Hirota T and Fukada Y. Resetting mechanism of central and peripheral circadian clocks in mammals. Zoolog Sci. 2004; 21: 359 -368. [PubMed] .
- 75. Kohsaka A and Bass J. A sense of time: how molecular clocks organize metabolism. Trends Endocrinol Metab. 2007; 18: 4 -11. [PubMed] .
- 76. La Fleur SE , Kalsbeek A , Wortel J and Buijs RM. A suprachiasmatic nucleus generated rhythm in basal glucose concentrations. J Neuroendocrinol. 1999; 11: 643 -652. [PubMed] .
- 77. La Fleur SE Daily rhythms in glucose metabolism: suprachiasmatic nucleus output to peripheral tissue. J Neuroendocrinol. 2003; 15: 315 -322. [PubMed] .
- 78. Davidson AJ , Castanon-Cervantes O and Stephan FK. Daily oscillations in liver function: diurnal vs circadian rhythmicity. Liver Int. 2004; 24: 179 -186. [PubMed] .
- 79. Ramsey KM , Marcheva B , Kohsaka A and Bass J. The clockwork of metabolism. Annu Rev Nutr. 2007; 27: 219 -240. [PubMed] .
- 80. Froy O Metabolism and Circadian Rhythms--Implications for Obesity. Endocr Rev. 2009; In press .
- 81. Yang X , Downes M , Yu RT , Bookout AL , He W , Straume M , Mangelsdorf DJ and Evans RM. Nuclear receptor expression links the circadian clock to metabolism. Cell. 2006; 126: 801 -810. [PubMed] .
- 82. Ruiter M , La Fleur SE , van Heijningen C , van der Vliet J , Kalsbeek A and Buijs RM. The daily rhythm in plasma glucagon concentrations in the rat is modulated by the biological clock and by feeding behavior. Diabetes. 2003; 52: 1709 -1715. [PubMed] .
- 83. Ando H , Yanagihara H , Hayashi Y , Obi Y , Tsuruoka S , Takamura T , Kaneko S and Fujimura A. Rhythmic messenger ribonucleic acid expression of clock genes and adipocytokines in mouse visceral adipose tissue. Endocrinology. 2005; 146: 5631 -5636. [PubMed] .
- 84. De Boer SF and Van der Gugten J. Daily variations in plasma noradrenaline, adrenaline and corticosterone concentrations in rats. Physiol Behav. 1987; 40: 323 -328. [PubMed] .
- 85. Ahima RS , Prabakaran D and Flier JS. Postnatal leptin surge and regulation of circadian rhythm of leptin by feeding. Implications for energy homeostasis and neuroendocrine function. J Clin Invest. 1998; 101: 1020 -1027. [PubMed] .
- 86. Bodosi B , Gardi J , Hajdu I , Szentirmai E , Obal F Jr and Krueger JM. Rhythms of ghrelin, leptin, and sleep in rats: effects of the normal diurnal cycle, restricted feeding, and sleep deprivation. Am J Physiol Regul Integr Comp Physiol. 2004; 287: R1071 -R1079. [PubMed] .
- 87. Kalra SP , Bagnasco M , Otukonyong EE , Dube MG and Kalra PS. Rhythmic, reciprocal ghrelin and leptin signaling: new insight in the development of obesity. Regul Pept. 2003; 111: 1 -11. [PubMed] .
- 88. Kalsbeek A , Fliers E , Romijn JA , La Fleur SE , Wortel J , Bakker O , Endert E and Buijs RM. The suprachiasmatic nucleus generates the diurnal changes in plasma leptin levels. Endocrinology. 2001; 142: 2677 -2685. [PubMed] .
- 89. Guan XM , Hess JF , Yu H , Hey PJ and van der Ploeg LH. Differential expression of mRNA for leptin receptor isoforms in the rat brain. Mol Cell Endocrinol. 1997; 133: 1 -7. [PubMed] .
- 90. Yi CX , van der Vliet J , Dai J , Yin G , Ru L and Buijs RM. Ventromedial arcuate nucleus communicates peripheral metabolic information to the suprachiasmatic nucleus. Endocrinology. 2006; 147: 283 -294. [PubMed] .
- 91. Zigman JM , Jones JE , Lee CE , Saper CB and Elmquist JK. Expression of ghrelin receptor mRNA in the rat and the mouse brain. J Comp Neurol. 2006; 494: 528 -548. [PubMed] .
- 92. Prosser RA and Bergeron HE. Leptin phase-advances the rat suprachiasmatic circadian clock in vitro. Neurosci Lett. 2003; 336: 139 -142. [PubMed] .
- 93. Yi CX , Challet E , Pevet P , Kalsbeek A , Escobar C and Buijs RM. A circulating ghrelin mimetic attenuates light-induced phase delay of mice and light-induced Fos expression in the suprachiasmatic nucleus of rats. Eur J Neurosci. 2008; 27: 1965 -1972. [PubMed] .
- 94. Yannielli PC , Molyneux PC , Harrington ME and Golombek DA. Ghrelin effects on the circadian system of mice. J Neurosci. 2007; 27: 2890 -2895. [PubMed] .
- 95. Rutter J , Reick M , Wu LC and McKnight SL. Regulation of clock and NPAS2 DNA binding by the redox state of NAD cofactors. Science. 2001; 293: 510 -514. [PubMed] .
- 96. Rutter J , Reick M and McKnight SL. Metabolism and the control of circadian rhythms. Annu Rev Biochem. 2002; 71: 307 -331. [PubMed] .
- 97. Zvonic S , Floyd ZE , Mynatt RL and Gimble JM. Circadian rhythms and the regulation of metabolic tissue function and energy homeostasis. Obesity (Silver Spring). 2007; 15: 539 -543. [PubMed] .
- 98. Loboda A , Kraft WK , Fine B , Joseph J , Nebozhyn M , Zhang C , He Y , Yang X , Wright C , Morris M , Chalikonda I , Ferguson M , Emilsson V , Leonardson A , Lamb J , Dai H , Schadt E , Greenberg HE and Lum PY. Diurnal variation of the human adipose transcriptome and the link to metabolic disease. BMC Med Genomics. 2009; 2: 7 [PubMed] .
- 99. Shimba S , Ishii N , Ohta Y , Ohno T , Watabe Y , Hayashi M , Wada T , Aoyagi T and Tezuka M. Brain and muscle Arnt-like protein-1 (BMAL1), a component of the molecular clock, regulates adipogenesis. Proc Natl Acad Sci U S A. 2005; 102: 12071 -12076. [PubMed] .
- 100. Chawla A and Lazar MA. Induction of Rev-ErbA alpha, an orphan receptor encoded on the opposite strand of the alpha-thyroid hormone receptor gene, during adipocyte differentiation. J Biol Chem. 1993; 268: 16265 -16269. [PubMed] .
- 101. Bray MS and Young ME. Circadian rhythms in the development of obesity: potential role for the circadian clock within the adipocyte. Obes Rev. 2007; 8: 169 -181. [PubMed] .
- 102. Torra IP , Tsibulsky V , Delaunay F , Saladin R , Laudet V , Fruchart JC , Kosykh V and Staels B. Circadian and glucocorticoid regulation of Rev-erbalpha expression in liver. Endocrinology. 2000; 141: 3799 -3806. [PubMed] .
- 103. Fontaine C , Dubois G , Duguay Y , Helledie T , Vu-Dac N , Gervois P , Soncin F , Mandrup S , Fruchart JC , Fruchart-Najib J and Staels B. The orphan nuclear receptor Rev-Erbalpha is a peroxisome proliferator-activated receptor (PPAR) gamma target gene and promotes PPARgamma-induced adipocyte differentiation. J Biol Chem. 2003; 278: 37672 -37680. [PubMed] .
- 104. Duez H and Staels B. Rev-erb alpha gives a time cue to metabolism. FEBS Lett. 2008; 582: 19 -25. [PubMed] .
- 105. Oishi K , Shirai H and Ishida N. CLOCK is involved in the circadian transactivation of peroxisome-proliferator-activated re-ceptor alpha (PPARalpha) in mice. Biochem J. 2005; 386: 575 -581. [PubMed] .
- 106. Inoue I , Shinoda Y , Ikeda M , Hayashi K , Kanazawa K , Nomura M , Matsunaga T , Xu H , Kawai S , Awata T , Komoda T and Katayama S. CLOCK/BMAL1 is involved in lipid metabolism via transactivation of the peroxisome proliferator-activated receptor (PPAR) response element. J Atheroscler Thromb. 2005; 12: 169 -174. [PubMed] .
- 107. Canaple L , Rambaud J , Dkhissi-Benyahya O , Rayet B , Tan NS , Michalik L , Delaunay F , Wahli W and Laudet V. Reciprocal regulation of brain and muscle Arnt-like protein 1 and peroxisome proliferator-activated receptor alpha defines a novel positive feedback loop in the rodent liver circadian clock. Mol Endocrinol. 2006; 20: 1715 -1727. [PubMed] .
- 108. Kersten S , Desvergne B and Wahli W. Roles of PPARs in health and disease. Nature. 2000; 405: 421 -424. [PubMed] .
- 109. Lefebvre P , Chinetti G , Fruchart JC and Staels B. Sorting out the roles of PPAR alpha in energy metabolism and vascular homeostasis. J Clin Invest. 2006; 116: 571 -580. [PubMed] .
- 110. Liu C , Li S , Liu T , Borjigin J and Lin JD. Transcriptional coactivator PGC-1alpha integrates the mammalian clock and energy metabolism. Nature. 2007; 447: 477 -481. [PubMed] .
- 111. Grimaldi B and Sassone-Corsi P. Circadian rhythms: metabolic clockwork. Nature. 2007; 447: 386 -387. [PubMed] .
- 112. Carling D AMP-activated protein kinase: balancing the scales. Biochimie. 2005; 87: 87 -91. [PubMed] .
- 113. Hardie DG , Hawley SA and Scott JW. AMP-activated protein kinase--development of the energy sensor concept. J Physiol. 2006; 574: 7 -15. [PubMed] .
- 114. Canto C , Gerhart-Hines Z , Feige JN , Lagouge M , Noriega L , Milne JC , Elliott PJ , Puigserver P and Auwerx J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature. 2009; 458: 1056 -1060. [PubMed] .
- 115. Canto C and Auwerx J. Caloric restriction, SIRT1 and longevity. Trends Endocrinol Metab. 2009; 20: 325 -331. [PubMed] .
- 116. Um JH , Yang S , Yamazaki S , Kang H , Viollet B , Foretz M and Chung JH. Activation of 5'-AMP-activated kinase with diabetes drug metformin induces casein kinase Iepsilon (CKIepsilon)-dependent degradation of clock protein mPER2. J Biol Chem. 2007; 282: 20794 -20798. [PubMed] .
- 117. Lamia KA , Sachdeva UM , DiTacchio L , Williams EC , Alvarez JG , Egan DF , Vasquez DS , Juguilon H , Panda S , Shaw RJ , Thompson CB and Evans RM. AMPK regulates the circadian clock by cryptochrome phosphorylation and degradation. Science. 2009; 326: 437 -440. [PubMed] .
- 118. Vieira E , Nilsson EC , Nerstedt A , Ormestad M , Long YC , Garcia-Roves PM , Zierath JR and Mahlapuu M. Relationship between AMPK and the transcriptional balance of clock-related genes in skeletal muscle. Am J Physiol Endocrinol Metab. 2008; 295: E1032 -E1037. [PubMed] .
- 119. Martin TL , Alquier T , Asakura K , Furukawa N , Preitner F and Kahn BB. Diet-induced obesity alters AMP kinase activity in hypothalamus and skeletal muscle. J Biol Chem. 2006; 281: 18933 -18941. [PubMed] .
- 120. Kola B Role of AMP-activated protein kinase in the control of appetite. J Neuroendocrinol. 2008; 20: 942 -951. [PubMed] .
- 121. Kola B and Korbonits M. Shedding light on the intricate puzzle of ghrelin's effects on appetite regulation. J Endocrinol. 2009; 202: 191 -198. [PubMed] .
- 122. Minokoshi Y , Shiuchi T , Lee S , Suzuki A and Okamoto S. Role of hypothalamic AMP-kinase in food intake regulation. Nutrition. 2008; 24: 786 -790. [PubMed] .
- 123. Blander G and Guarente L. The Sir2 family of protein deacetylases. Annu Rev Biochem. 2004; 73: 417 -435. [PubMed] .
- 124. Dali-Youcef N , Lagouge M , Froelich S , Koehl C , Schoonjans K and Auwerx J. Sirtuins: the 'magnificent seven', function, metabolism and longevity. Ann Med. 2007; 39: 335 -345. [PubMed] .
- 125. Mair W and Dillin A. Aging and survival: the genetics of life span extension by dietary restriction. Annu Rev Biochem. 2008; 77: 727 -754. [PubMed] .
- 126. Haigis MC and Guarente LP. Mammalian sirtuins--emerging roles in physiology, aging, and calorie restriction. Genes Dev. 2006; 20: 2913 -2921. [PubMed] .
- 127. Imai SI "Clocks" in the NAD World: NAD as a metabolic oscillator for the regulation of metabolism and aging. Biochim Biophys Acta. 2009; In press .
- 128. Asher G , Gatfield D , Stratmann M , Reinke H , Dibner C , Kreppel F , Mostoslavsky R , Alt FW and Schibler U. SIRT1 regulates circadian clock gene expression through PER2 deacetylation. Cell. 2008; 134: 317 -328. [PubMed] .
- 129. Nakahata Y , Kaluzova M , Grimaldi B , Sahar S , Hirayama J , Chen D , Guarente LP and Sassone-Corsi P. The NAD+-dependent deacetylase SIRT1 modulates CLOCK-mediated chromatin remodeling and circadian control. Cell. 2008; 134: 329 -340. [PubMed] .
- 130. Hirayama J , Sahar S , Grimaldi B , Tamaru T , Takamatsu K , Nakahata Y and Sassone-Corsi P. CLOCK-mediated acetylation of BMAL1 controls circadian function. Nature. 2007; 450: 1086 -1090. [PubMed] .
- 131. Belden WJ and Dunlap JC. SIRT1 is a circadian deacetylase for core clock components. Cell. 2008; 134: 212 -214. [PubMed] .
- 132. Nakahata Y , Sahar S , Astarita G , Kaluzova M and Sassone-Corsi P. Circadian Control of the NAD+ Salvage Pathway by CLOCK-SIRT1. Science. 2009; 324: 654 -657. [PubMed] .
- 133. Turek FW , Joshu C , Kohsaka A , Lin E , Ivanova G , McDearmon E , Laposky A , Losee-Olson S , Easton A , Jensen DR , Eckel RH , Takahashi JS and Bass J. Obesity and metabolic syndrome in circadian Clock mutant mice. Science. 2005; 308: 1043 -1045. [PubMed] .
- 134. Rudic RD , McNamara P , Curtis AM , Boston RC , Panda S , Hogenesch JB and Fitzgerald GA. BMAL1 and CLOCK, two essential components of the circadian clock, are involved in glucose homeostasis. PLoS Biol. 2004; 2: e377 [PubMed] .
- 135. Kennaway DJ , Owens JA , Voultsios A , Boden MJ and Varcoe TJ. Metabolic homeostasis in mice with disrupted Clock gene expression in peripheral tissues. Am J Physiol Regul Integr Comp Physiol. 2007; 293: R1528 -R1537. [PubMed] .
- 136. Oishi K , Atsumi G , Sugiyama S , Kodomari I , Kasamatsu M , Machida K and Ishida N. Disrupted fat absorption attenuates obesity induced by a high-fat diet in Clockmutant mice. FEBS Lett. 2006; 580: 127 -130. [PubMed] .
- 137. Lamia KA , Storch KF and Weitz CJ. Physiological significance of a peripheral tissue circadian clock. Proc Natl Acad Sci U S A. 2008; 105: 15172 -15177. [PubMed] .
- 138. Yang S , Liu A , Weidenhammer A , Cooksey RC , McClain D , Kim MK , Aguilera G , Abel ED and Chung JH. The role of mPer2 clock gene in glucocorticoid and feeding rhythms. Endocrinology. 2009; 150: 2153 -2160. [PubMed] .
- 139. Cassone VM and Stephan FK. Central and peripheral regulation of feeding and nutrition by the mammalian circadian clock: implications for nutrition during manned space flight. Nutrition. 2002; 18: 814 -819. [PubMed] .
- 140. Honma KI , Honma S and Hiroshige T. Critical role of food amount for prefeeding corticosterone peak in rats. Am J Physiol. 1983; 245: R339 -R344. [PubMed] .
- 141. Grasl-Kraupp B , Bursch W , Ruttkay-Nedecky B , Wagner A , Lauer B and Schulte-Hermann R. Food restriction eliminates preneoplastic cells through apoptosis and antagonizes carcinogenesis in rat liver. Proc Natl Acad Sci U S A. 1994; 91: 9995 -9999. [PubMed] .
- 142. Mistlberger RE Circadian food-anticipatory activity: formal models and physiological mechanisms. Neurosci Biobehav Rev. 1994; 18: 171 -195. [PubMed] .
- 143. Hara R , Wan K , Wakamatsu H , Aida R , Moriya T , Akiyama M and Shibata S. Restricted feeding entrains liver clock without participation of the suprachiasmatic nucleus. Genes Cells. 2001; 6: 269 -278. [PubMed] .
- 144. Boulamery-Velly A , Simon N , Vidal J , Mouchet J and Bruguerolle B. Effects of three-hour restricted food access during the light period on circadian rhythms of temperature, locomotor activity, and heart rate in rats. Chronobiol Int. 2005; 22: 489 -498. [PubMed] .
- 145. Hirao J , Arakawa S , Watanabe K , Ito K and Furukawa T. Effects of restricted feeding on daily fluctuations of hepatic functions including p450 monooxygenase activities in rats. J Biol Chem. 2006; 281: 3165 -3171. [PubMed] .
- 146. Saito M , Murakami E and Suda M. Circadian rhythms in disaccharidases of rat small intestine and its relation to food intake. Biochim Biophys Acta. 1976; 421: 177 -179. [PubMed] .
- 147. Comperatore CA and Stephan FK. Entrainment of duodenal activity to periodic feeding. J Biol Rhythms. 1987; 2: 227 -242. [PubMed] .
- 148. Stephan FK The "other" circadian system: food as a Zeitgeber. J Biol Rhythms. 2002; 17: 284 -292. [PubMed] .
- 149. Stephan FK , Swann JM and Sisk CL. Anticipation of 24-hr feeding schedules in rats with lesions of the suprachiasmatic nucleus. Behav Neural Biol. 1979; 25: 346 -363. [PubMed] .
- 150. Oishi K , Miyazaki K and Ishida N. Functional CLOCK is not involved in the entrainment of peripheral clocks to the restricted feeding: entrainable expression of mPer2and Bmal1 mRNAs in the heart of Clock mutant mice on Jcl:ICR background. Biochem Biophys Res Commun. 2002; 298: 198 -202. [PubMed] .
- 151. Horikawa K , Minami Y , Iijima M , Akiyama M and Shibata S. Rapid damping of food-entrained circadian rhythm of clock gene expression in clock-defective peripheral tissues under fasting conditions. Neuroscience. 2005; 134: 335 -343. [PubMed] .
- 152. Damiola F , Le Minh N , Preitner N , Kornmann B , Fleury-Olela F and Schibler U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev. 2000; 14: 2950 -2961. [PubMed] .
- 153. Stokkan KA , Yamazaki S , Tei H , Sakaki Y and Menaker M. Entrainment of the circadian clock in the liver by feeding. Science. 2001; 291: 490 -493. [PubMed] .
- 154. Lin JD , Liu C and Li S. Integration of energy metabolism and the mammalian clock. Cell Cycle. 2008; 7: 453 -457. [PubMed] .
- 155. Mieda M , Williams SC , Richardson JA , Tanaka K and Yanagisawa M. The dorsomedial hypothalamic nucleus as a putative food-entrainable circadian pacemaker. Proc Natl Acad Sci U S A. 2006; 103: 12150 -12155. [PubMed] .
- 156. Gooley JJ , Schomer A and Saper CB. The dorsomedial hypothalamic nucleus is critical for the expression of food-entrainable circadian rhythms. Nat Neurosci. 2006; 9: 398 -407. [PubMed] .
- 157. Landry GJ , Simon MM , Webb IC and Mistlberger RE. Persistence of a behavioral food-anticipatory circadian rhythm following dorsomedial hypothalamic ablation in rats. Am J Physiol Regul Integr Comp Physiol. 2006; 290: R1527 -R1534. [PubMed] .
- 158. Landry GJ , Yamakawa GR , Webb IC , Mear RJ and Mistlberger RE. The dorsomedial hypothalamic nucleus is not necessary for the expression of circadian food-anticipatory activity in rats. J Biol Rhythms. 2007; 22: 467 -478. [PubMed] .
- 159. Davidson AJ , Cappendijk SL and Stephan FK. Feeding-entrained circadian rhythms are attenuated by lesions of the parabrachial region in rats. Am J Physiol Regul Integr Comp Physiol. 2000; 278: R1296 -R1304. [PubMed] .
- 160. Mistlberger RE and Mumby DG. The limbic system and food-anticipatory circadian rhythms in the rat: ablation and dopamine blocking studies. Behav Brain Res. 1992; 47: 159 -168. [PubMed] .
- 161. Mendoza J , Angeles-Castellanos M and Escobar C. Differential role of the accumbens Shell and Core subterritories in food-entrained rhythms of rats. Behav Brain Res. 2005; 158: 133 -142. [PubMed] .
- 162. Davidson AJ Search for the feeding-entrainable circadian oscillator: a complex proposition. Am J Physiol Regul Integr Comp Physiol. 2006; 290: R1524 -R1526. [PubMed] .
- 163. Comperatore CA and Stephan FK. Effects of vagotomy on entrainment of activity rhythms to food access. Physiol Behav. 1990; 47: 671 -678. [PubMed] .
- 164. Mistlberger RE and Marchant EG. Enhanced food-anticipatory circadian rhythms in the genetically obese Zucker rat. Physiol Behav. 1999; 66: 329 -335. [PubMed] .
- 165. Pitts S , Perone E and Silver R. Food-entrained circadian rhythms are sustained in arrhythmic Clk/Clk mutant mice. Am J Physiol Regul Integr Comp Physiol. 2003; 285: R57 -R67. [PubMed] .
- 166. Pendergast JS , Nakamura W , Friday RC , Hatanaka F , Takumi T and Yamazaki S. Robust food anticipatory activity in BMAL1-deficient mice. PLoS ONE. 2009; 4: e4860 [PubMed] .
- 167. Storch KF and Weitz CJ. Daily rhythms of food-anticipatory behavioral activity do not require the known circadian clock. Proc Natl Acad Sci U S A. 2009; 106: 6808 -6813. [PubMed] .
- 168. Feillet CA , Ripperger JA , Magnone MC , Dulloo A , Albrecht U and Challet E. Lack of food anticipation in Per2 mutant mice. Curr Biol. 2006; 16: 2016 -2022. [PubMed] .
- 169. Mistlberger RE Circadian rhythms: perturbing a food-entrained clock. Curr Biol. 2006; 16: R968 -R969. [PubMed] .
- 170. LeSauter J , Hoque N , Weintraub M , Pfaff DW and Silver R. Stomach ghrelin-secreting cells as food-entrainable circadian clocks. Proc Natl Acad Sci U S A. 2009; 106: 13582 -13587. [PubMed] .
- 171. Wu MW , Li XM , Xian LJ and Levi F. Effects of meal timing on tumor progression in mice. Life Sci. 2004; 75: 1181 -1193. [PubMed] .
- 172. Filipski E , Innominato PF , Wu M , Li XM , Iacobelli S , Xian LJ and Levi F. Effects of light and food schedules on liver and tumor molecular clocks in mice. J Natl Cancer Inst. 2005; 97: 507 -517. [PubMed] .
- 173. Masoro EJ Overview of caloric restriction and ageing. Mech Ageing Dev. 2005; 126: 913 -922. [PubMed] .
- 174. Spindler SR Caloric restriction: From soup to nuts. Ageing Res Rev. 2009; In press .
- 175. Colman RJ , Anderson RM , Johnson SC , Kastman EK , Kosmatka KJ , Beasley TM , Allison DB , Cruzen C , Simmons HA , Kemnitz JW and Weindruch R. Caloric restriction delays disease onset and mortality in rhesus monkeys. Science. 2009; 325: 201 -204. [PubMed] .
- 176. Roth GS , Mattison JA , Ottinger MA , Chachich ME , Lane MA and Ingram DK. Aging in rhesus monkeys: relevance to human health interventions. Science. 2004; 305: 1423 -1426. [PubMed] .
- 177. Fontana L Modulating human aging and age-associated diseases. Biochim Biophys Acta. 2009; 1790: 1133 -1138. [PubMed] .
- 178. Masoro EJ Caloric restriction-induced life extension of rats and mice: a critique of proposed mechanisms. Biochim Biophys Acta. 2009; 1790: 1040 -1048. [PubMed] .
- 179. Harman D Aging: A theory based on free radical and radiation chemistry. J Gerontol. 1956; 11: 298 -300. [PubMed] .
- 180. Sohal RS and Weindruch R. Oxidtive stress, caloric restriction, and aging. Science. 1996; 273: 59 -63. [PubMed] .
- 181. Van Remmen H , Ikeno Y , Hamilton M , Pahlavani M , Wolf N , Thorpe SR , Alderson NL , Baynes JW , Epstein CJ , Huang TT , Nelson J , Strong R and Richardson A. Life-long reduction in MnSOD activity results in increased DNA damage and higher incidence of cancer but does not accelerate aging. Physiol Genomics. 2003; 16: 29 -37. [PubMed] .
- 182. Perez VI , Bokov A , Van Remmen H , Mele J , Ran Q , Ikeno Y and Richardson A. Is the oxidative stress theory of aging dead. Biochim Biophys Acta. 2009; 1790: 1005 -1014. [PubMed] .
- 183. Salmon AB , Perez VI , Bokov A , Jernigan A , Kim G , Zhao H , Levine RL and Richardson A. Lack of methionine sulfoxide reductase A in mice increases sensitivity to oxidative stress but does not diminish life span. FASEB J. 2009; 23: 3601 -3608. [PubMed] .
- 184. Blagosklonny MV Aging: ROS or TOR. Cell Cycle. 2008; 7: 3344 -3354. [PubMed] .
- 185. Jiang W , Zhu Z and Thompson HJ. Dietary energy restriction modulates the activity of AMP-activated protein kinase, Akt, and mammalian target of rapamycin in mammary carcinomas, mammary gland, and liver. Cancer Res. 2008; 68: 5492 -5499. [PubMed] .
- 186. Selman C , Tullet JM , Wieser D , Irvine E , Lingard SJ , Choudhury AI , Claret M , Al-Qassab H , Carmignac D , Ramadani F , Woods A , Robinson IC , Schuster E , Batterham RL , Kozma SC , Thomas G , Carling D , Okkenhaug K , Thornton JM , Partridge L , Gems D and Withers DJ. Ribosomal protein S6 kinase 1 signaling regulates mammalian life span. Science. 2009; 326: 140 -144. [PubMed] .
- 187. Harrison DE , Strong R , Sharp ZD , Nelson JF , Astle CM , Flurkey K , Nadon NL , Wilkinson JE , Frenkel K , Carter CS , Pahor M , Javors MA , Fernandez E and Miller RA. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009; 460: 392 -395. [PubMed] .
- 188. Cao R , Lee B , Cho HY , Saklayen S and Obrietan K. Photic regulation of the mTOR signaling pathway in the suprachiasmatic circadian clock. Mol Cell Neurosci. 2008; 38: 312 -324. [PubMed] .
- 189. Duffy PH , Feuers RJ , Leakey JA , Nakamura K , Turturro A and Hart RW. Effect of chronic caloric restriction on physiological variables related to energy metabolism in the male Fischer 344 rat. Mech Ageing Dev. 1989; 48: 117 -133. [PubMed] .
- 190. Challet E , Solberg LC and Turek FW. Entrainment in calorie-restricted mice: conflicting zeitgebers and free-running conditions. Am J Physiol. 1998; 274: R1751 -R1761. [PubMed] .
- 191. Challet E , Caldelas I , Graff C and Pevet P. Synchronization of the molecular clockwork by light- and food-related cues in mammals. Biol Chem. 2003; 384: 711 -719. [PubMed] .
- 192. Mendoza J , Graff C , Dardente H , Pevet P and Challet E. Feeding cues alter clock gene oscillations and photic responses in the suprachiasmatic nuclei of mice exposed to a light/dark cycle. J Neurosci. 2005; 25: 1514 -1522. [PubMed] .
- 193. Swindell WR Comparative analysis of microarray data identifies common responses to caloric restriction among mouse tissues. Mech Ageing Dev. 2008; 129: 138 -153. [PubMed] .
- 194. Froy O , Chapnik N and Miskin R. Long-lived alphaMUPA transgenic mice exhibit pronounced circadian rhythms. Am J Physiol Endocrinol Metab. 2006; 291: E1017 -E1024. [PubMed] .
- 195. Froy O and Miskin R. The interrelations among feeding, circadian rhythms and ageing. Prog Neurobiol. 2007; 82: 142 -150. [PubMed] .
- 196. Anson RM , Guo Z , de Cabo R , Iyun T , Rios M , Hagepanos A , Ingram DK , Lane MA and Mattson MP. Intermittent fasting dissociates beneficial effects of dietary restriction on glucose metabolism and neuronal resistance to injury from calorie intake. Proc Natl Acad Sci U S A. 2003; 100: 6216 -6220. [PubMed] .
- 197. Descamps O , Riondel J , Ducros V and Roussel AM. Mitochondrial production of reactive oxygen species and incidence of age-associated lymphoma in OF1 mice: effect of alternate-day fasting. Mech Ageing Dev. 2005; 126: 1185 -1191. [PubMed] .
- 198. Froy O , Chapnik N and Miskin R. Effect of intermittent fasting on circadian rhythms in mice depends on feeding time. Mech Ageing Dev. 2009; 130: 154 -160. [PubMed] .
- 199. Goodrick CL , Ingram DK , Reynolds MA , Freeman JR and Cider N. Effects of intermittent feeding upon body weight and lifespan in inbred mice: interaction of genotype and age. Mech Ageing Dev. 1990; 55: 69 -87. [PubMed] .
- 200. Mattson MP and Wan R. Beneficial effects of intermittent fasting and caloric restriction on the cardiovascular and cerebrovascular systems. J Nutr Biochem. 2005; 16: 129 -137. [PubMed] .
- 201. Contestabile A and Ciani E. Dietary restriction differentially protects from neurodegeneration in animal models of excitotoxicity. Brain Res. 2004; 1002: 162 -166. [PubMed] .
- 202. Mattson MP Energy intake, meal frequency, and health: a neurobiological perspective. Annu Rev Nutr. 2005; 25: 237 -260. [PubMed] .
- 203. Sharma S and Kaur G. Neuroprotective potential of dietary restriction against kainate-induced excitotoxicity in adult male Wistar rats. Brain Res Bull. 2005; 67: 482 -491. [PubMed] .
- 204. Ahmet I , Wan R , Mattson MP , Lakatta EG and Talan M. Cardioprotection by intermittent fasting in rats. Circulation. 2005; 112: 3115 -3121. [PubMed] .
- 205. Mager DE , Wan R , Brown M , Cheng A , Wareski P , Abernethy DR and Mattson MP. Caloric restriction and intermittent fasting alter spectral measures of heart rate and blood pressure variability in rats. FASEB J. 2006; 20: 631 -637. [PubMed] .
- 206. Varady KA and Hellerstein MK. Alternate-day fasting and chronic disease prevention: a review of human and animal trials. Am J Clin Nutr. 2007; 86: 7 -13. [PubMed] .
- 207. Mattson MP , Duan W , Wan R and Guo Z. Prophylactic activation of neuroprotective stress response pathways by dietary and behavioral manipulations. NeuroRx. 2004; 1: 111 -116. [PubMed] .
- 208. Mattson MP Dietary factors, hormesis and health. Ageing Res Rev. 2008; 7: 43 -48. [PubMed] .
- 209. Duan W , Lee J , Guo Z and Mattson MP. Dietary restriction stimulates BDNF production in the brain and thereby protects neurons against excitotoxic injury. J Mol Neurosci. 2001; 16: 1 -12. [PubMed] .
- 210. Newton IG , Forbes ME , Legault C , Johnson JE , Brunso-Bechtold JK and Riddle DR. Caloric restriction does not reverse aging-related changes in hippocampal BDNF. Neurobiol Aging. 2005; 26: 683 -688. [PubMed] .
- 211. Andrade JP , Mesquita R , Assuncao M and Pereira PA. Effects of food restriction on synthesis and expression of brain-derived neurotrophic factor and tyrosine kinase B in dentate gyrus granule cells of adult rats. Neurosci Lett. 2006; 399: 135 -140. [PubMed] .
- 212. Maswood N , Young J , Tilmont E , Zhang Z , Gash DM , Gerhardt GA , Grondin R , Roth GS , Mattison J , Lane MA , Carson RE , Cohen RM , Mouton PR , Quigley C , Mattson MP and Ingram DK. Caloric restriction increases neurotrophic factor levels and attenuates neurochemical and behavioral deficits in a primate model of Parkinson's disease. Proc Natl Acad Sci U S A. 2004; 101: 18171 -18176. [PubMed] .
- 213. Xu B , Goulding EH , Zang K , Cepoi D , Cone RD , Jones KR , Tecott LH and Reichardt LF. Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor. Nat Neurosci. 2003; 6: 736 -742. [PubMed] .
- 214. Duan W , Guo Z , Jiang H , Ware M and Mattson MP. Reversal of behavioral and metabolic abnormalities, and insulin resistance syndrome, by dietary restriction in mice deficient in brain-derived neurotrophic factor. Endocrinology. 2003; 144: 2446 -2453. [PubMed] .
- 215. Liang FQ , Sohrabji F , Miranda R , Earnest B and Earnest D. Expression of brain-derived neurotrophic factor and its cognate receptor, TrkB, in the rat suprachiasmatic nucleus. Exp Neurol. 1998; 151: 184 -193. [PubMed] .
- 216. Liang FQ , Allen G and Earnest D. Role of brain-derived neurotrophic factor in the circadian regulation of the suprachiasmatic pacemaker by light. J Neurosci. 2000; 20: 2978 -2987. [PubMed] .
- 217. Miskin R , Axelrod JH , Griep AE , Lee E , Belin D , Vassalli JD and Westphal H. Human and murine urokinase cDNAs linked to the murine alpha A-crystallin promoter exhibit lens and non-lens expression in transgenic mice. Eur J Biochem. 1990; 190: 31 -38. [PubMed] .
- 218. Mondino A and Blasi F. uPA and uPAR in fibrinolysis, immunity and pathology. Trends Immunol. 2004; 25: 450 -455. [PubMed] .
- 219. Soreq H and Miskin R. Plasminogen activator in the developing rat cerebellum: biosynthesis and localization in granular neurons. Brain Res. 1983; 313: 149 -158. [PubMed] .
- 220. Sumi Y , Dent MA , Owen DE , Seeley PJ and Morris RJ. The expression of tissue and urokinase-type plasminogen activators in neural development suggests different modes of proteolytic involvement in neuronal growth. Development. 1992; 116: 625 -637. [PubMed] .
- 221. Masos T and Miskin R. mRNAs encoding urokinase-type plaminogen activator and plasminogen activator inhibitor-1 are elevated in the mouse brain following kainate-mediated excitation. Brain Res Mol Brain Res. 1997; 47: 157 -169. [PubMed] .
- 222. Bahi A , Boyer F , gumy C , Kafri T and Dreyer JL. In vivo gene delivery of urokinase-type plasminogen activator with regulatable lentivirus induces behavioural changes in chronic cocaine administration. Eur J Neurosci. 2004; 20: 3473 -3488. [PubMed] .
- 223. Bahi A and Dreyer JL. Overexpression of plasminogen activators in the nucleus accumbens enhances cocaine-, amphetamine- and morphine-induced reward and behavioral sensitization. Genes Brain Behav. 2008; 7: 244 -256. [PubMed] .
- 224. Lahtinen L , Lukasiuk K and Pitkanen A. Increased expression and activity of urokinase-type plasminogen activator during epileptogenesis. Eur J Neurosci. 2006; 24: 1935 -1945. [PubMed] .
- 225. Miskin R , Tirosh O , Pardo M , Zusman I , Schwartz B , Yahav S , Dubnov G and Kohen R. alphaMUPA mice: a transgenic model for longevity induced by caloric restriction. Mech Ageing Dev. 2005; 126: 255 -261. [PubMed] .
- 226. Miskin R , Masos T , Shoham Z and Williams-Simons L. Urokinase-type plasminogen activator mRNA is expressed in normal developing teeth and leads to abnormal incisor enamel in alpha MUPA transgenic mice. Transgenic Res. 2006; 15: 241 -254. [PubMed] .
- 227. Miskin R and Masos T. Transgenic mice overexpressing urokinase-type plasminogen activator in the brain exhibit reduced food consumption, body weight and size, and increased longevity. J Gerontol A Biol Sci Med Sci. 1997; 52: B118 -B124. [PubMed] .
- 228. Miskin R , Masos T , Yahav S , Shinder D and Globerson A. alphaMUPA mice: a transgenic model for increased life span. Neurobiol Aging. 1999; 20: 555 -564. [PubMed] .
- 229. Tirosh O , Aronis A , Zusman I , Kossoy G , Yahav S , Shinder D , Abramovitz R and Miskin R. Mitochondrion-mediated apoptosis is enhanced in long-lived alphaMUPA transgenic mice and calorically restricted wild-type mice. Exp Gerontol. 2003; 38: 955 -963. [PubMed] .
- 230. Tirosh O , Pardo M , Schwartz B and Miskin R. Long-lived alphaMUPA transgenic mice show reduced SOD2 expression, enhanced apoptosis and reduced susceptibility to the carcinogen dimethylhydrazine. Mech Ageing Dev. 2005; 126: 1262 -1273. [PubMed] .
- 231. Masoro EJ , Shimokawa I , Higami Y , McMahan CA and Yu BP. Temporal pattern food intake not a factor in the retardation of aging processes by dietary restriction. J Gerontol A Biol Sci Med Sci. 1995; 50A: B48 -53. [PubMed] .
- 232. Nelson W and Halberg F. Meal-timing, circadian rhythms and life span of mice. J Nutr. 1986; 116: 2244 -2253. [PubMed] .
- 233. Cheney KE , Liu RK , Smith GS , Meredith PJ , Mickey MR and Walford RL. The effect of dietary restriction of varying duration on survival, tumor patterns, immune function, and body temperature in B10C3F1 female mice. J Gerontol. 1983; 38: 420 -430. [PubMed] .
- 234. Nelson W and Halberg F. Schedule-shifts, circadian rhythms and lifespan of freely feeding and meal-fed mice. Physiol Behav. 1986; 38: 781 -788. [PubMed] .
- 235. Heilbronn LK , Civitarese AE , Bogacka I , Smith SR , Hulver M and Ravussin E. Glucose tolerance and skeletal muscle gene expression in response to alternate day fasting. Obes Res. 2005; 13: 574 -581. [PubMed] .
- 236. Heilbronn LK , Smith SR , Martin CK , Anton SD and Ravussin E. Alternate-day fasting in nonobese subjects: effects on body weight, body composition, and energy metabolism. Am J Clin Nutr. 2005; 81: 69 -73. [PubMed] .
- 237. Aksungar FB , Topkaya AE and Akyildiz M. Interleukin-6, C-reactive protein and biochemical parameters during prolonged intermittent fasting. Ann Nutr Metab. 2007; 51: 88 -95. [PubMed] .
- 238. Beck B and Richy S. Dietary modulation of ghrelin and leptin and gorging behavior after weight loss in the obese Zucker rat. J Endocrinol. 2009; 202: 29 -34. [PubMed] .
- 239. Yang H , Youm YH , Nakata C and Dixit VD. Chronic caloric restriction induces forestomach hypertrophy with enhanced ghrelin levels during aging. Peptides. 2007; 28: 1931 -1936. [PubMed] .
- 240. Gonzalez AA , Kumar R , Mulligan JD , Davis AJ , Weindruch R and Saupe KW. Metabolic adaptations to fasting and chronic caloric restriction in heart, muscle, and liver do not include changes in AMPK activity. Am J Physiol Endocrinol Metab. 2004; 287: E1032 -E1037. [PubMed] .
- 241. Das M , Gabriely I and Barzilai N. Caloric restriction, body fat and ageing in experimental models. Obes Rev. 2004; 5: 13 -19. [PubMed] .
- 242. Harrison DE and Archer JR. Genetic differences in effects of food restriction on aging in mice. J Nutr. 1987; 117: 376 -382. [PubMed] .
- 243. Allison DB , Miller RA , Austad SN , Bouchard C , Leibel R , Klebanov S , Johnson T and Harrison DE. Genetic variability in responses to caloric restriction in animals and in regulation of metabolism and obesity in humans. J Gerontol A Biol Sci Med Sci. 2001; 56 Spec No 1: 55 -65. [PubMed] .
- 244. Bordone L and Guarente L. Calorie restriction, SIRT1 and metabolism: understanding longevity. Nat Rev Mol Cell Biol. 2005; 6: 298 -305. [PubMed] .
- 245. Masternak MM , Al-Regaiey KA , Del Rosario Lim MM , Jimenez-Ortega V , Panici JA , Bonkowski MS , Kopchick JJ and Bartke A. Effects of caloric restriction and growth hormone resistance on the expression level of peroxisome proliferator-activated receptors superfamily in liver of normal and long-lived growth hormone receptor/binding protein knockout mice. J Gerontol A Biol Sci Med Sci. 2005; 60: 1394 -1398. [PubMed] .
- 246. Chen D , Bruno J , Easlon E , Lin SJ , Cheng HL , Alt FW and Guarente L. Tissue-specific regulation of SIRT1 by calorie restriction. Genes Dev. 2008; 22: 1753 -1757. [PubMed] .
- 247. Cohen HY , Miller C , Bitterman KJ , Wall NR , Hekking B , Kessler B , Howitz KT , Gorospe M , de Cabo R and Sinclair DA. Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase. Science. 2004; 305: 390 -392. [PubMed] .
- 248. Boily G , Seifert EL , Bevilacqua L , He XH , Sabourin G , Estey C , Moffat C , Crawford S , Saliba S , Jardine K , Xuan J , Evans M , Harper ME and McBurney MW. SirT1 regulates energy metabolism and response to caloric restriction in mice. PLoS One. 2008; 3: e1759 [PubMed] .
- 249. Bordone L , Cohen D , Robinson A , Motta MC , van Veen E , Czopik A , Steele AD , Crowe H , Marmor S , Luo J , Gu W and Guarente L. SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell. 2007; 6: 759 -767. [PubMed] .
- 250. Ramadori G , Lee CE , Bookout AL , Lee S , Williams KW , Anderson J , Elmquist JK and Coppari R. Brain SIRT1: anatomical distribution and regulation by energy availability. J Neurosci. 2008; 28: 9989 -9996. [PubMed] .
- 251. Taleux N , De Potter I , Deransart C , Lacraz G , Favier R , Leverve XM , Hue L and Guigas B. Lack of starvation-induced activation of AMP-activated protein kinase in the hypothalamus of the Lou/C rats resistant to obesity. Int J Obes (Lond). 2008; 32: 639 -647. [PubMed] .
- 252. Veyrat-Durebex C , Montet X , Vinciguerra M , Gjinovci A , Meda P , Foti M and Rohner-Jeanrenaud F. The Lou/C rat: a model of spontaneous food restriction associated with improved insulin sensitivity and decreased lipid storage in adipose tissue. Am J Physiol Endocrinol Metab. 2009; 296: E1120 -1132. [PubMed] .
- 253. Guarente L and Picard F. Calorie restriction--the SIR2 connection. Cell. 2005; 120: 473 -482. [PubMed] .