



Introduction
Life expectancy for individuals with sickle cell disease (SCD) is significantly lower than that of the general population; however, with advances in medical care, life expectancy has markedly improved over the past three decades in resource-rich countries [1]. Despite these gains, adolescents and adults with SCD still experience higher rates of aging-related morbidity and early mortality [2–4]. Individuals with SCD disproportionately experience functional decline, frailty, and increased rates of premature end-organ damage and malignancies [4–8]. While causes of these aging-related complications have not been elucidated, the presence of these complications suggests that many individuals with SCD experience accelerated aging, particularly in the bone marrow, possibly in response to chronic hypoxia, hemolysis and inflammation associated with a lifetime of ineffective erythropoiesis [3, 9, 10]. Earlier identification of individuals with SCD at greater risk for such age-related (and potentially life-threatening complications) is critical in improving long term outcomes and life expectancy in SCD.
Cellular senescence, a hallmark of biological aging, has been identified as a key contributor to functional decline and onset of aging-related diseases [11, 12]. Cells that undergo senescence are permanently cell-cycle arrested but remain metabolically active, secreting pro-inflammatory signals. Cellular senescence in the immune system has been shown to promote cellular senescence throughout organs and tissues and cause functional decline [13]. Expression of p16INK4a (p16) is a reliable biomarker of cellular senescence [14]. In humans, increased expression of p16 mRNA in peripheral blood T lymphocytes (PBTL) is associated with well-established pro-aging exposures such as tobacco use, physical inactivity, chemotherapy, and radiation, and genetic alterations in the p16 locus have been associated with aging-related conditions in humans such as frailty, cardiovascular disease and type 2 diabetes [14, 15]. In adolescent and young adult survivors of cancer, PBTL p16 expression is markedly increased in a treatment dose-dependent manner and higher expression is associated with frailty and sarcopenia [16]. Taken together, prior work suggests that increased cellular senescence, as measured by PBTL p16 expression, identifies accelerated biological aging [12, 14, 17]. Given SCD is associated with an accelerated aging phenotype [6], we hypothesized that p16 expression would be elevated in people with SCD.
The primary objective of this study was to measure p16 expression in adolescents and young adults (AYAs) living with SCD and compare them to a pre-existing sample of similarly aged individuals without SCD. Our hypothesis was that individuals with SCD would have higher p16 expression compared to the non-SCD comparators.
Results
Participant characteristics
Twenty-one AYAs with SCD were consented for this study, with three participants excluded from the analysis due to failed sample quality control for the p16 assay. Eighteen participants and 27 comparators were included in the final analysis. Participant and available comparator characteristics are summarized in Table 1, and additional SCD patient characteristics are detailed in Supplementary Table 1. Due to the need to protect the privacy of blood donors, there was no available demographic data beyond what is detailed in Table 1.
Table 1. Baseline characteristics of the study population.
| SCD (n = 18) | Non-SCD Comparators (n = 27) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (years), median (range) | 22 (15–27) | 20 (17–29) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gender, n (%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Female | 11 (61%) | 20 (74%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Male | 7 (39%) | 7 (26%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Genotype, n (%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HbSS | 16 (89%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HbSβ0-thalassemia | 2 (11%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| WBC count (109/L), median (IQR) | 11.3 (8.2–13.8) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hemoglobin (g/dL), median (IQR) | 9.1 (8.7–9.6) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Platelet count (109/L), median (IQR) | 356 (282–477) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Absolute reticulocytes (109/L), median (IQR) | 223 (172–281) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Absolute neutrophils (109/L), median (IQR) | 6.5 (4.6–8.2) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Absolute lymphocytes (109/L), median (IQR) | 3.1 (1.9–3.5) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| On hydroxyurea, n (%) | 10 (56%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hydroxyurea dose* (mg/kg) | 19 (16–21) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Time on hydroxyurea* (months) | 119 (64–140) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| On chronic transfusions, n (%) | 8 (44%) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Time on chronic transfusions* (months), median (IQR) | 59 (6–120) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| *Includes only participants receiving this therapy. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Expression of p16
Participants with SCD had significantly higher p16 expression vs. comparators −10.1 vs. 8.7 log2 p16 units. Based on prior analysis of the regression of p16 expression by chronological age, this mean difference of 1.3 log2 units (95% CI 0.79–1.9, p < 0.001) between the SCD and comparator groups may be as much as 43 years (95% CI 23–63 years) of increased biological aging (Figure 1A) [17]. These results were similar when stratifying by chronic transfusion therapy (CTT), a therapy for severe SCD complications such as stroke. The mean difference in p16 expression between the SCD participants on CTT and comparators was 1.3 log2 p16 units (10.0 vs. 8.7 log2 p16 units, p = 0.002), while the difference between the SCD participants not on CTT and comparators was 1.4 (10.1 vs. 8.7 log2 p16 units, p = 0.001) (Figure 1B).
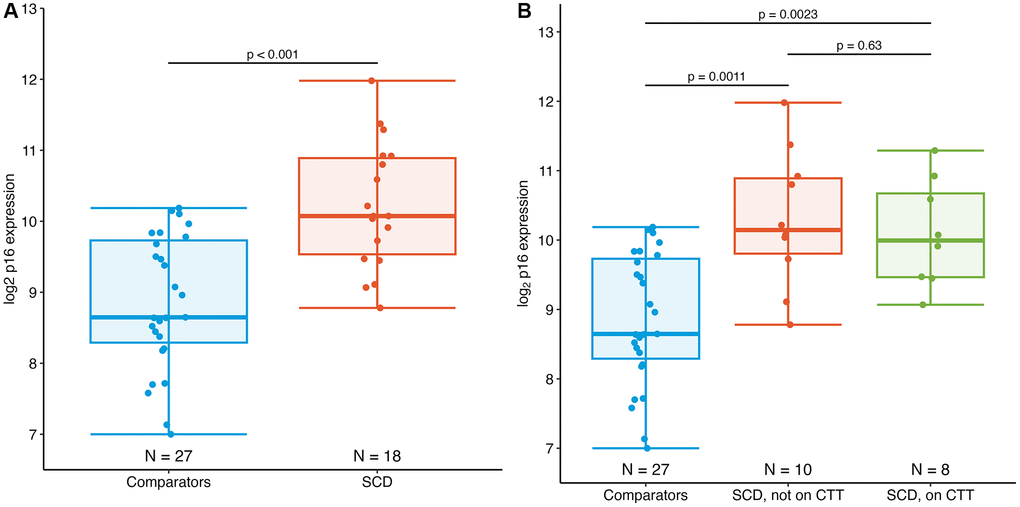
Figure 1. Expression of PBTL p16INK4a in AYAs with SCD. (A) Mean p16 expression in AYAs with SCD (10.1 log2 p16 units) is significantly elevated compared to those without SCD (8.7 log2 p16 units), p < 0.001. (B) Expression of p16 in individuals with SCD did not differ by chronic transfusion therapy (CTT) status.
Exploratory analysis
Within the SCD group, we explored associations between laboratory measures, SCD complications, and PROs. In this small sample, there were no associations between any of these measures and p16 expression (Supplementary Table 2). In the SCD group, p16 expression was higher throughout, regardless of age (Figure 2).
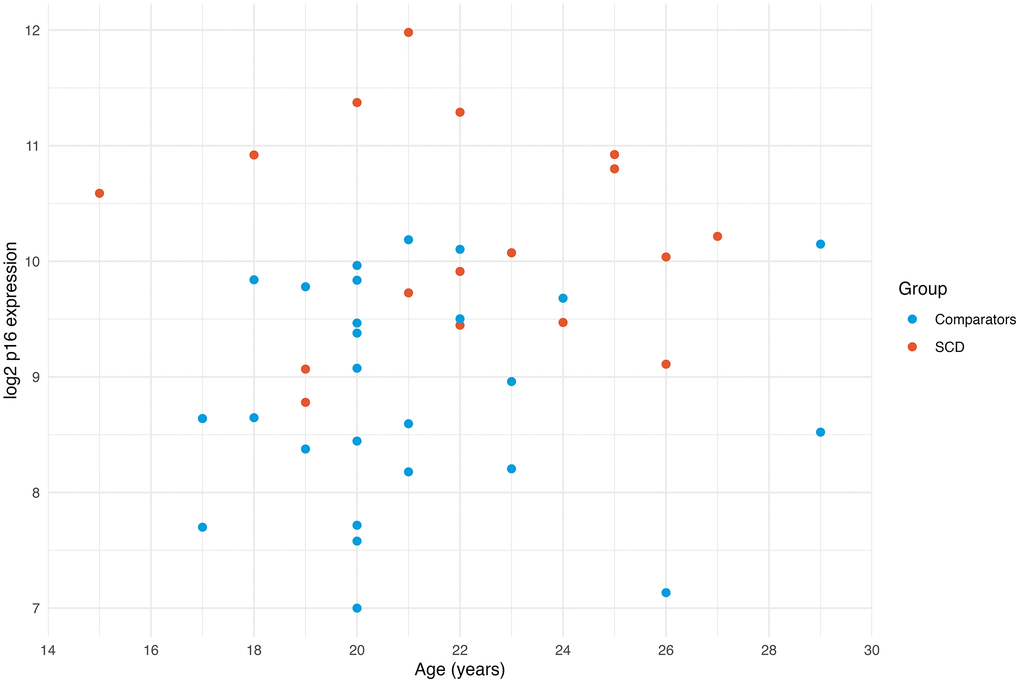
Figure 2. Expression of p16 by age. Scatterplot of p16 expression by age demonstrates that the SCD group generally has higher p16 expression across the entire age range. The youngest SCD participant has higher p16 expression than all the comparators.
Discussion
In this first assessment of PBTL p16 expression in SCD, we demonstrate that p16 expression, a marker of cellular senescence, is strikingly elevated in AYAs with SCD compared to non-SCD age matched comparators. The magnitude of this difference was unexpected, signifying an accelerated aging of up to 43 years in people with SCD, and is even larger than the difference between the AYA cancer survivors treated with pro-aging exposures (cytotoxic chemotherapy) and the same comparator group used in this study [16]. Our youngest participant, a 15-year-old with SCD, had a higher p16 expression than all the comparators, underscoring the early rise of p16 expression in this population. This work complements growing reports on accelerated aging in SCD [6, 18].
The etiology of the marked elevation in p16 expression in SCD is not yet known. Stress and ineffective erythropoiesis due to the hemolysis and increased RBC turnover, tissue hypoxia, anemia and chronic inflammation are all potential stressors to the bone marrow in people with SCD, which may manifest in accumulation of senescent cells and lead to accelerated organismal aging [9, 19].
To protect the privacy of the blood donors, the core of the comparator group, self-identified race of these participants is not available. We explored whether the marked difference in p16 expression observed in this study between the two groups could be partially explained by racial differences, as >90% of individuals with SCD identify as Black/African-American. There is no convincing evidence that p16 expression varies by race. Prior studies have observed no difference in p16 expression between Black/African-American and Caucasian populations [15]. To further explore, we compared p16 expression in the SCD cohort to nine similarly-aged individuals who self-identified as Black/African-American from the previous AYA cancer survivor p16 study, all of whom had received prior chemotherapy [20]. This comparison showed higher mean p16 expression in the SCD cohort (10.1 vs. 8.8 log2 units, p = 0.2), albeit the difference was not statistically significant likely due to the small sample (see Supplementary Figure 1). Subgroup comparisons within the AYA cancer survivor cohort also showed no differences by race. We also observed no overall differences in p16 expression by race after combining participants from multiple datasets from prior p16 studies (see Supplementary Figure 2). Altogether, these findings support that race alone does not account for the surprisingly large differences in p16 expression observed in this study.
Lastly, we explored any association between p16 expression and clinical/PRO measures in SCD participants in this small SCD patient sample to yield insight into potential drivers and clinical significance. As shown in Supplementary Table 2, we did not detect an association between p16 expression and any of the assessed clinical and PRO measures. Additionally, there was no association between disease-modifying therapies such as hydroxyurea, chronic transfusion therapy and p16 expression. There are several potential explanations for these findings: (1) This study was underpowered to detect these associations, (2) the p16 expression observed is a predictive marker of future morbidity and thereby may be more suitable for a longitudinal cohort, or most likely (3) the widespread impact and burden of SCD on cellular senescence itself may be greater than these various markers of clinical severity, which often does not capture disease severity in SCD, which is poorly defined [21]. We anticipate that a minimum sample size of at least 80–90 SCD participants is necessary to have 80% power to detect these associations.
These findings must be interpreted in the setting of study limitations. A cross-sectional design and limited sample size may not capture the relationship between p16 expression and treatments and clinical outcomes. A larger longitudinal study may be able to dissect causes of elevated p16 expression and may lead to interventional studies to address accelerated aging in SCD. Additionally, the bone marrow compartment in people living with SCD may be more prone to the accumulation of senescent cells, making PBTL p16 expression an incomplete reflection of organismal aging in this population. Thus, the age gap we found in this study may be an overestimate. Future studies should also examine the correlation of PBTL p16 expression with other senescence and immune markers from other body compartments in individuals with SCD.
Conclusions
We demonstrate that AYAs with SCD exhibit early and accelerated biological aging with markedly increased p16 expression, a marker of cellular senescence. While further prospective research is needed to establish the clinical implications and predictive utility for elevated p16 expression in patients with SCD, these initial findings provide molecular evidence of biological aging beginning at a young age in individuals living with SCD.
Methods
Study design and participants
The study cross-sectionally measured p16 expression in both individuals with and without SCD. The study protocol was approved by the Institutional Review Board at the University of North Carolina (UNC). AYAs with SCD (HbSS or HbSβ0-thalassemia genotypes only) ages 15 to 29 years receiving care at UNC SCD clinics from 2021–2022 were enrolled following informed consent and assent as appropriate. A 10mL peripheral blood specimen was collected for p16 analysis and participants completed patient-reported outcome (PRO) measures validated for use among people with SCD. We used a convenient sample of AYAs without SCD comparators that was previously recruited from 2018–2020 for an AYA cancer survivor study of p16 expression [16]. These comparators were either recruited from the UNC Platelet Donation Center (healthy individuals) or from the UNC Children’s Hematology Clinic (seen for non-malignant, non-SCD disorders such as iron deficiency anemia) and provided a blood sample similarly to individuals with SCD.
Measures
For all participants, p16 expression was measured as previously described [14–16, 22]. Briefly, CD3+ cells were purified from whole blood and used to isolate total RNA. Expression of p16 was measured by a TaqMan RT-PCR (Sapere Bio, Research Triangle Park, NC, USA) and reported as log2 units. Collection of key patient and treatment data was performed by medical record abstraction. In non-SCD samples where p16 expression was below the limit of detection, the value of 7 log2 p16 units (lower limit of assay detection) was assigned. None of the SCD samples had p16 expression below the limit of detection.
Statistical analysis
Expression of p16 in log2 units was compared among groups using 2-sided Student’s t-tests. Other measures were compared to p16 expression with linear regression. Analysis was performed with R (R Foundation for Statistical Computing, Vienna, Austria). Statistical significance was defined as p-value < 0.05 with 95% confidence intervals (CI) reported for all tests. See Supplementary Methods for additional details.
Data sharing statement
De-identified data from this study will be shared upon reasonable request via email to the corresponding author.
Author Contributions
S.R.W contributed to study design and execution, data analysis and interpretation, and wrote and revised the manuscript. N.M. contributed to p16 expression analysis and assisted in manuscript preparation. V.A.M. contributed to study design and execution, enrollment, and assisted in manuscript revision. A.B.S. and M.A.C. supervised the project and contributed to study conception and design, data analysis and interpretation, and critically revised the manuscript.
Acknowledgments
The authors would like to thank the patients and families who participated in this work. We appreciate Dr. Hy Muss for critical review of the manuscript.
Conflicts of Interest
M.A.C. is currently an employee of Cogent Biosciences, which provided no funding for this work. N.M. is a founder and a shareholder of Sapere Bio. The remaining authors have no other relevant conflicts of interest to disclose.
Ethical Statement and Consent
This study was approved by the Institutional Review Board at UNC, the protocol number IRB# 20-2196. All participants provided informed consent and assent.
Funding
S.R.W. received salary support from the National Heart, Lung, and Blood Institute branch of the National Institutes of Health through Grant Award Number 5T32HL007149-44. Analysis of p16 expression and other work was supported by the National Center for Advancing Translational Sciences (NCATS), NIH, through Grant Award Number UL1TR002489 to M.A.C and A.S., KL2TR002490 to M.A.C. Funders were not involved in study design or data interpretation.
References
- 1. Quinn CT, Rogers ZR, McCavit TL, Buchanan GR. Improved survival of children and adolescents with sickle cell disease. Blood. 2010; 115:3447–52. https://doi.org/10.1182/blood-2009-07-233700 [PubMed]
- 2. DeBaun MR, Ghafuri DL, Rodeghier M, Maitra P, Chaturvedi S, Kassim A, Ataga KI. Decreased median survival of adults with sickle cell disease after adjusting for left truncation bias: a pooled analysis. Blood. 2019; 133:615–7. https://doi.org/10.1182/blood-2018-10-880575 [PubMed]
- 3. Oyedeji C, Strouse JJ, Crawford RD, Garrett ME, Ashley-Koch AE, Telen MJ. A multi-institutional comparison of younger and older adults with sickle cell disease. Am J Hematol. 2019; 94:E115–7. https://doi.org/10.1002/ajh.25405 [PubMed]
- 4. Thein SL, Howard J. How I treat the older adult with sickle cell disease. Blood. 2018; 132:1750–60. https://doi.org/10.1182/blood-2018-03-818161 [PubMed]
- 5. Oyedeji CI, Pieper C, Hall K, Morey M, Whitson H, Strouse JJ. Functional Assessment in Younger and Older Adults with Sickle Cell Disease. Blood. 2020; 136: 26–7. https://doi.org/10.1182/blood-2020-140854
- 6. Idris IM, Botchwey EA, Hyacinth HI. Sickle cell disease as an accelerated aging syndrome. Exp Biol Med (Maywood). 2022; 247:368–74. https://doi.org/10.1177/15353702211068522 [PubMed]
- 7. Brunson A, Keegan THM, Bang H, Mahajan A, Paulukonis S, Wun T. Increased risk of leukemia among sickle cell disease patients in California. Blood. 2017; 130:1597–9. https://doi.org/10.1182/blood-2017-05-783233 [PubMed]
- 8. Seminog OO, Ogunlaja OI, Yeates D, Goldacre MJ. Risk of individual malignant neoplasms in patients with sickle cell disease: English national record linkage study. J R Soc Med. 2016; 109:303–9. https://doi.org/10.1177/0141076816651037 [PubMed]
- 9. El Hoss S, Cochet S, Godard A, Yan H, Dussiot M, Frati G, Boutonnat-Faucher B, Laurance S, Renaud O, Joseph L, Miccio A, Brousse V, Narla M, El Nemer W. Fetal hemoglobin rescues ineffective erythropoiesis in sickle cell disease. Haematologica. 2021; 106:2707–19. https://doi.org/10.3324/haematol.2020.265462 [PubMed]
- 10. Wilson SR, Boynton MH, Carden MA, Ciccone E, Derebail VK, Ataga KI. Age-Associated Decline in Blood Parameters in Individuals with Sickle Cell Disease. Blood. 2021; 138: 970. https://doi.org/10.1182/blood-2021-145919
- 11. He S, Sharpless NE. Senescence in Health and Disease. Cell. 2017; 169:1000–11. https://doi.org/10.1016/j.cell.2017.05.015 [PubMed]
- 12. Borghesan M, Hoogaars WMH, Varela-Eirin M, Talma N, Demaria M. A Senescence-Centric View of Aging: Implications for Longevity and Disease. Trends Cell Biol. 2020; 30:777–91. https://doi.org/10.1016/j.tcb.2020.07.002 [PubMed]
- 13. Yousefzadeh MJ, Flores RR, Zhu Y, Schmiechen ZC, Brooks RW, Trussoni CE, Cui Y, Angelini L, Lee KA, McGowan SJ, Burrack AL, Wang D, Dong Q, et al. An aged immune system drives senescence and ageing of solid organs. Nature. 2021; 594:100–5. https://doi.org/10.1038/s41586-021-03547-7 [PubMed]
- 14. Liu Y, Sanoff HK, Cho H, Burd CE, Torrice C, Ibrahim JG, Thomas NE, Sharpless NE. Expression of p16(INK4a) in peripheral blood T-cells is a biomarker of human aging. Aging Cell. 2009; 8:439–48. https://doi.org/10.1111/j.1474-9726.2009.00489.x [PubMed]
- 15. Sanoff HK, Deal AM, Krishnamurthy J, Torrice C, Dillon P, Sorrentino J, Ibrahim JG, Jolly TA, Williams G, Carey LA, Drobish A, Gordon BB, Alston S, et al. Effect of cytotoxic chemotherapy on markers of molecular age in patients with breast cancer. J Natl Cancer Inst. 2014; 106:dju057. https://doi.org/10.1093/jnci/dju057 [PubMed]
- 16. Smitherman AB, Wood WA, Mitin N, Ayer Miller VL, Deal AM, Davis IJ, Blatt J, Gold SH, Muss HB. Accelerated aging among childhood, adolescent, and young adult cancer survivors is evidenced by increased expression of p16INK4a and frailty. Cancer. 2020; 126:4975–83. https://doi.org/10.1002/cncr.33112 [PubMed]
- 17. Muss HB, Smitherman A, Wood WA, Nyrop K, Tuchman S, Randhawa PK, Entwistle AR, Mitin N, Shachar SS. p16 a biomarker of aging and tolerance for cancer therapy. Transl Cancer Res. 2020; 9:5732–42. https://doi.org/10.21037/tcr.2020.03.39 [PubMed]
- 18. Lê BM, Hatch D, Yang Q, Shah N, Luyster FS, Garrett ME, Tanabe P, Ashley-Koch AE, Knisely MR. Characterizing epigenetic aging in an adult sickle cell disease cohort. Blood Adv. 2024; 8:47–55. https://doi.org/10.1182/bloodadvances.2023011188 [PubMed]
- 19. Tolu SS, Wang K, Yan Z, Zhang S, Roberts K, Crouch AS, Sebastian G, Chaitowitz M, Fornari ED, Schwechter EM, Uehlinger J, Manwani D, Minniti CP, Bouhassira EE. Characterization of Hematopoiesis in Sickle Cell Disease by Prospective Isolation of Stem and Progenitor Cells. Cells. 2020; 9:2159. https://doi.org/10.3390/cells9102159 [PubMed]
- 20. Shachar SS, Deal AM, Reeder-Hayes KE, Nyrop KA, Mitin N, Anders CK, Carey LA, Dees EC, Jolly TA, Kimmick GG, Karuturi MS, Reinbolt RE, Speca JC, et al. Effects of Breast Cancer Adjuvant Chemotherapy Regimens on Expression of the Aging Biomarker, p16INK4a. JNCI Cancer Spectrum. 2020; 4. https://doi.org/10.1093/jncics/pkaa082
- 21. Steinberg MH, Gladwin MT. "Severity" in adult sickle cell disease. Am J Hematol. 2023; 98:1508–11. https://doi.org/10.1002/ajh.27024 [PubMed]
- 22. Wood WA, Krishnamurthy J, Mitin N, Torrice C, Parker JS, Snavely AC, Shea TC, Serody JS, Sharpless NE. Chemotherapy and Stem Cell Transplantation Increase p16INK4a Expression, a Biomarker of T-cell Aging. EBioMedicine. 2016; 11:227–38. https://doi.org/10.1016/j.ebiom.2016.08.029 [PubMed]