



Introduction
As a common chronic disease worldwide, the incidence of hypertension has been increasing annually in recent years, with a population exceeding 1 billion in the world. In China, the morbidity of hypertension is as high as 12% [1], showing the characteristics of high incidence, high disability rate, high mortality rate, low awareness rate, low cure rate, and low control rate. Hypertension is one of the major risk factors for cardiovascular and cerebrovascular diseases [2]. The kidney is one of the target organs in hypertensive damage and damage to renal structure and function caused by essential hypertension is defined as hypertensive renal disease (HRD), the incidence of which is correlated to the degree and duration of blood pressure elevation [3, 4]. Oxidative stress (OS) is observed in the pathophysiological process of hypertension and is characterized by excessive reactive oxygen species (ROS) production and altered Redox status [5]. Multiple transcription factors are activated by the stimulation of the release of ROS, which further facilitates the production of chemokines and cytokines to induce the recruitment of inflammatory and immune cells, thereby promoting cardiovascular and renal inflammation and fibrosis [6]. The kidney is a hypermetabolic organ, and renal mitochondria are rich in oxidative reactions, making them susceptible to OS injuries. The impaired mitochondrial function is triggered by the produced ROS, which eventually results in the impairment of renal cells and tissue function to aggravate renal damage and the progression of chronic kidney disease [7]. Compared to subjects with normal blood pressure, a long-term increase in blood pressure is observed in hypertensive patients, resulting in continuous increases in intravascular pressure to trigger damage to glomerular capillary endothelial cells, renal tubular epithelial cells, and podocytes, further increasing the basement membrane permeability. The renal filtration barrier system is disrupted and proteins are permeated from blood vessels into the urine, which is characterized by microalbuminuria [8]. Urinary protein is the initiating factor of renal interstitial fibrosis and a risk factor for the development of end-stage renal disease [3, 4]. It is claimed that alleviating OS and inflammatory damage of renal tubular epithelial cells shows a significant repair effect on the renal filtration barrier system [9], which might be a new strategy for HRD treatment.
Kruppel-like factor 2 (KLF2) is a member of the zinc finger family of DNA-binding transcription factors that regulate endothelial cell metabolism. KLF2 expression is found to be increased by shear stress in branched points of blood vessels [10], and it has also been found to inhibit endothelial inflammation [11], which is related to endothelial dysfunction, one of the predictors of atherosclerosis development and cardiovascular events [12]. Also, endothelial dysfunction is a mechanism underlying the development of hypertension [13]. Hypertension is associated with endothelial dysfunction. Bae L, et al. have recently reported the renoprotective effect of KLF2 on glomerular endothelial dysfunction in hypertensive nephropathy through angiotensin II-AT1R [14].
Azilsartan medoxomi is developed by Takeda and belongs to a new generation of selective AT1 subtype angiotensin II receptor antagonist, and is the prodrug of Azilsartan. A clinical study has reported the stable and durable antihypertensive effects of Azilsartan [15]. Apart from its antihypertensive effect, other functions claimed for Azilsartan include the prevention of myocardial hypertrophy [16], vascular remodeling [17], and renal function damage [18]. Furthermore, an extremely suppressive property of Azilsartan against OS and inflammatory damage has been illustrated [19, 20].
In this research, we assumed that Azilsartan may have a protective effect in the increase in uterine artery embolization (UAE) in models of hypertension. The underlying mechanism was further explored.
Materials and Methods
Animals and treatments
8-week mice were divided into four groups: Control, angiotensin and high-salt (ANG/HS), ANG/HS + Azilsartan (10 μg/day, Cat#:ajci10796, Amgucam, Shanghai, China), and ANG/HS + Azilsartan (20 μg/day) groups for 3 weeks. To construct the ANG/HS model in mice, animals were implanted with an osmotic minipump to administer 60 ng/min ANG for 14 days. During the administration, the HS diet (8% NaCl) was given to mice. After the modeling, mice were dosed with 10 μg/day or 20 μg/day Azilsartan for 3 weeks. The daily water intake, food intake, and mean arterial pressure were detected during the treatments [21].
Measurement of UAE
Mice were kept in a 24 h metabolic cage for the purpose of collecting urine for 24h while being given free access to water. The collected urine was centrifuged at 3000 rpm for 20 min and the supernatant was retained. The UAE was tested using the ELISA kit (Mlbio, China).
Enzyme-linked immunosorbent assay (ELISA) for the detection of cytokine levels
50 μL samples were diluted at a 1:1 ratio and loaded into the well, followed by introducing 50 μL Biotin-labeled antibody. After 60 min incubation at 37° C, the reagent was removed and 80 μL HRP-loaded secondary antibody, was followed by half an hour culture at 37° C. Then, 50 μL TMB substrates were added and cultured at 37° C for 10 min, followed by loading 50 μL stop solution and detecting the OD value utilizing a microplate reader (MD-Bio, USA).
Immunostaining assay
Collected renal tissues were dehydrated by 70%, 80%, and 90% ethanol solution, successively, followed by dehydration with xylene until transparent, which were then embedded and sliced. Slides were incubated with 10% goat serum overnight and were then introduced with the primary antibody against occludin (1:1000, Affinity, USA) at 4° C for 24 hours. The secondary antibody (1:200, Abcam, UK) was then added for incubation at 4° C for 24 hours, and then rinsed and stained with DAB dye. Lastly, images were taken using the inverted microscope (Nikon, Japan).
Detection of oxidative stress biomarkers
The content of 4,4’-Methylenedianiline (MDA) and Superoxide dismutase (SOD) activity in renal tissues were checked. A commercial kit (Elabscience, USA) was utilized to examine the MDA content in renal tissues using the TBA method according to the kit instructions. The SOD activity in renal tissues was determined with an EnzyChrom Superoxide Dismutase Assay Kit (BioAssay Systems, USA) according to the kit instructions.
Fluorescein-5-isothiocyanate (FITC)-dextran assay
HrGECs were loaded on the transwell to form a monolayer, followed by introducing FITC-dextran (AbMole, USA) in the upper chamber. After 60 min culture, the OD value in the lower chamber at 492/520 nm was detected by a microplate reader (MD-Bio, USA).
Trans-epithelial/endothelial electrical resistance (TEER)
The in vitro endothelial permeability was measured with the TEER assay with a 1600R ECIS System (Applied Biophysics, Australia) according to the method described previously [22]. The data was presented with an average of the resistance values (Ω·cm2) and the average percent change from baseline TEER.
Real-time polymerase chain reaction (PCR)
RNAs were extracted from HrGECs using the TRIzol solution, followed by dissolving in ddH2O and quantified by detecting the absorption at 260 nm. The transcription to cDNA was performed using the reverse transcription kit (QIAGEN, Germany), followed by conducting the PCR reaction utilizing the ABI 7500 Real-time PCR system (Applied Biosystems, USA) with the SYBR green (Sigma, USA). β-actin is used as the housekeeping gene. The gene levels were calculated with the 2-ΔΔCt method. The primer sequences are listed as below: IL-6: Forward 5’- CACCGGGAACGAAAGAGAAG-3’, Reverse, 5’-TCTGAGGTGCCCATGCTACAT-3’; TNF-α : Forward, 5’-CAGAGGGAAGAGTTCCCCAG-3’, Reverse, 5’-CCTTGGTCTGGTAGGAGACG-3’; IL-1β: Forward, 5’-GCTGCTTCCAAACCTTTGAC-3, Reverse, 5’-AGCTTCTCCACAGCCACAAT-3’; occludin: Forward, 5′-TTGGCTACGGAGGTGGCTATGG-3′, Reverse 5′- ACTAAGGAAGCGATGAAGCAGAAGG-3′; KLF2: Forward, 5’-AGACCTACACCAAGAGTTCGCATC-3′, Reverse 5′-ATC GCACAGATGGCACTGGAATG-3′; β-actin: Forward, 5’-CACCCACTCCTCCACCTTTG-3′, Reverse, 5’-CCACCACCCTGTTGCTGTAG-3′.
Western blotting assay
Following isolating proteins from HrGECs, the BCA method was performed for the quantification of proteins, which were further separated with a 12% SDS-PAGE. After transferring onto the PVDF membrane, proteins were blocked with skim milk for 2 hours. Subsequently, the membrane was loaded with primary antibodies against occludin (1:1000, Cat#: sc-133256, SCBT, USA), KLF2 (1:1500, Cat#: ab236507, Abcam, USA), or β-actin (1:10000, Cat#: sc-56459, SCBT, USA) for 12 hours at 4° C, followed by adding the secondary antibody (1:2000, Cat#: sc-2357, SCBT, USA) for 1.5 hours [23]. Lastly, the intensity of the blots was quantified using the Bio-Rad Quantity One software (USA) and analyzed using the GraphPad Prism 9 software.
Statistical analysis
The software GraphPad Prism 9 was used for statistical analysis. Data were presented as mean±standard deviation (S.D.) and the comparison was analyzed using a one-way analysis of variance (ANOVA) method, followed by Bonferroni’s post-hoc test. P<0.05 was taken as a statistically significant difference.
Data availability
The data is available at a reasonable request.
Results
The effects of Azilsartan on ANG/HS-induced hypertension mice
After treatments with Azilsartan, the general condition of mice was measured. The daily average water intake (Figure 1A) was increased from 6.22 ml/d to 9.32 ml/d in the ANG/HS group, then reduced to 7.82 ml/d and 7.05 ml/d by 10 and 20 μg/day Azilsartan, respectively. Moreover, the daily food intake values (Figure 1B) in the control, ANG/HS, ANG/HS+Azilsartan (10 μg/day), and ANG/HS+Azilsartan (20 μg/day) groups were 3.43, 4.85, 4.13, and 3.82 g/d, respectively. The mean arterial pressure (Figure 1C) was elevated from 118.5 mmHg to 162.3 mmHg in the ANG/HS group, then reduced to 136.3 mmHg and 128.8 mmHg by 10 and 20 μg/day Azilsartan, respectively. Hypertension symptoms in mice were alleviated by Azilsartan.
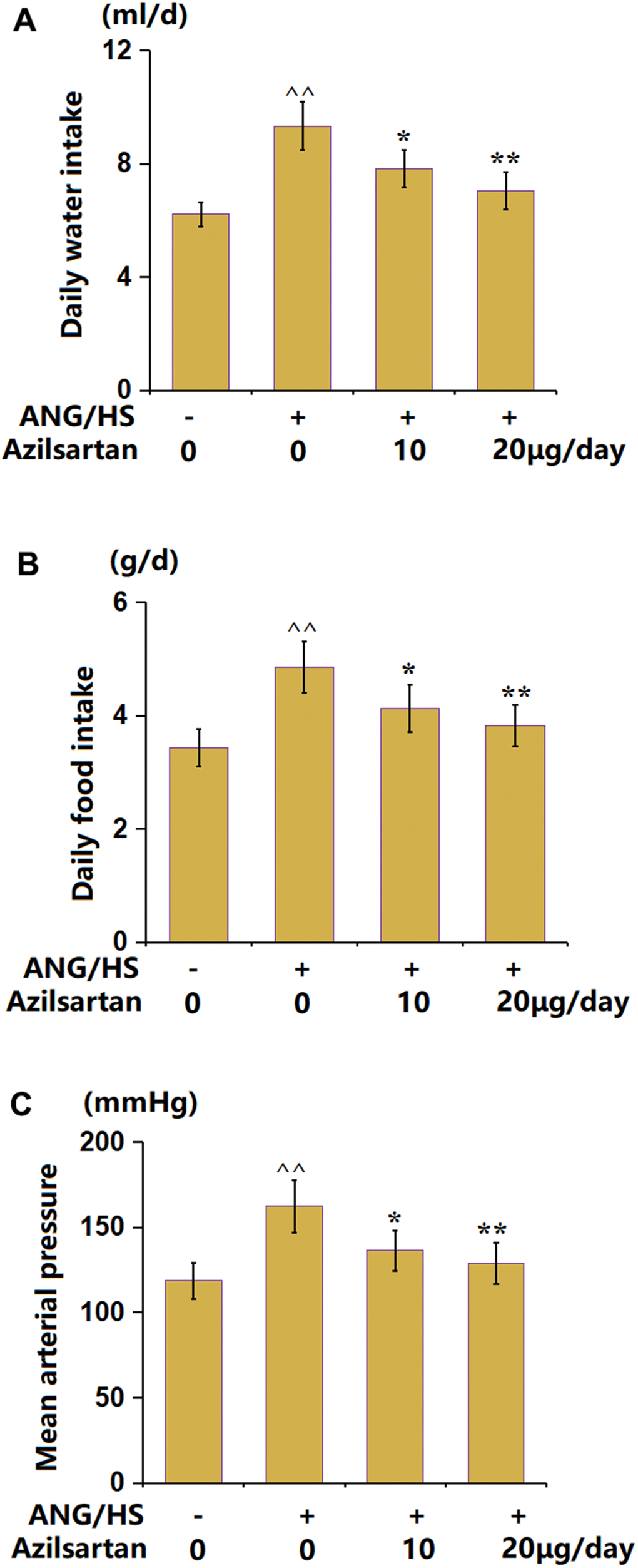
Figure 1. The effects of Azilsartan on ANG/HS-induced hypertension mice. (A) daily water intake; (B) daily food intake; (C) Mean arterial pressure (n=6, ^^, P<0.01 vs. vehicle group; *, **, P<0.05, 0.01 vs. ANG/HS group).
Azilsartan mitigated inflammatory response in renal tissues in ANG/HS- challenged mice
Renal tissues were then extracted for inflammation detection. IL-6, TNF-α, and IL-1β levels were extremely increased in the ANG/HS group, but signally reduced by 10 and 20 μg/day Azilsartan (Figure 2A). Furthermore, the IL-6 level (Figure 2B) in the ANG/HS group was promoted from 1.83 to 3.11 pg/mL, then decreased to 2.38 and 1.78 pg/mL by 10 and 20 μg/day Azilsartan, respectively. Similarly, the TNF-α levels in the control, ANG/HS, ANG/HS+ Azilsartan (10 μg/day), and ANG/HS + Azilsartan (20 μg/day) groups were 1.06, 2.26, 1.76, and 1.37 pg/mL, respectively. Moreover, The expressions of IL-1β in the control, ANG/HS, ANG/HS+ Azilsartan (10 μg/day), and ANG/HS + Azilsartan (20 μg/day) groups were 1.69, 3.58, 2.45, 1.89 pg/mL, respectively. Azilsartan showed a suppressive impact on inflammatory response in ANG/HS-challenged mice.
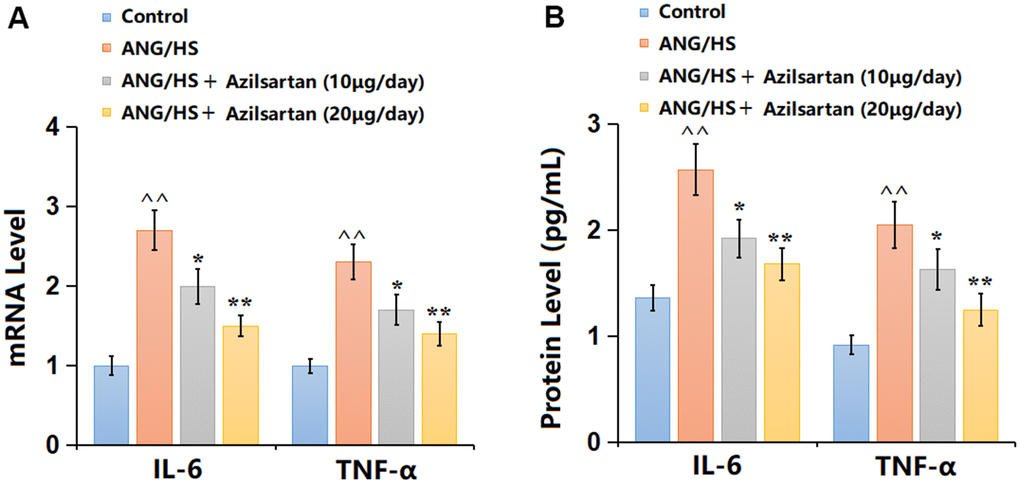
Figure 2. Azilsartan mitigated inflammatory response in renal tissues in ANG/HS-challenged mice. (A) mRNA levels of IL-6, TNF-α and IL-1β; (B) Protein levels of IL-6, TNF-α and IL-1β were measured by ELISA (n=6, ^^, P<0.01 vs. vehicle group; *, **, P<0.05, 0.01 vs. ANG/HS group).
Azilsartan attenuated OS in renal tissues in ANG/HS- challenged mice
The MDA content in ANG/HS- challenged mice was enhanced from 0.85 to 2.56 nmol/mg protein, which was reduced to 1.93 and 1.55 nmol/mg protein by 10 and 20 μg/day Azilsartan, respectively (Figure 3A). In addition, the SOD activity levels (Figure 3B) in the control, ANG/HS, ANG/HS+ Azilsartan (10 μg/day), and ANG/HS+Azilsartan (20 μg/day) groups were 61.3, 36.5, 47.2, and 55.6 U/mg protein, respectively. Azilsartan exerted a suppressive function on OS in ANG/HS-challenged mice.
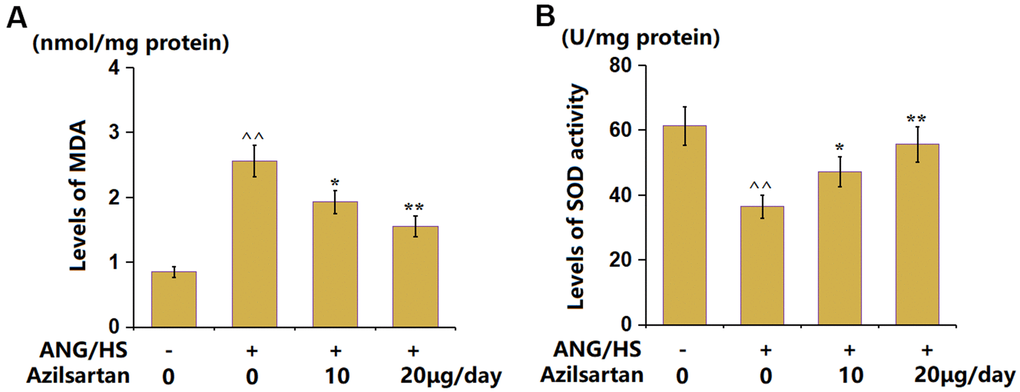
Figure 3. Azilsartan attenuated oxidative stress in renal tissues in ANG/HS-challenged mice. (A) The levels of MDA in renal tissues; (B) The levels of SOD activity in renal tissues (n=5, ^^, P<0.01 vs. vehicle group; *, **, P<0.05, 0.01 vs. ANG/HS group).
The effects of Azilsartan on UAE in ANG/HS-challenged mice
The UAE in ANG/HS-challenged mice was found to be markedly increased from 53.5 mg/d to 276.3 mg/d, which signally declined to 196.5 mg/d and 157.2 mg/d by 10 μg/day and 20 μg/day Azilsartan (Figure 4), implying a repairment by Azilsartan on UAE in ANG/HS-challenged mice.
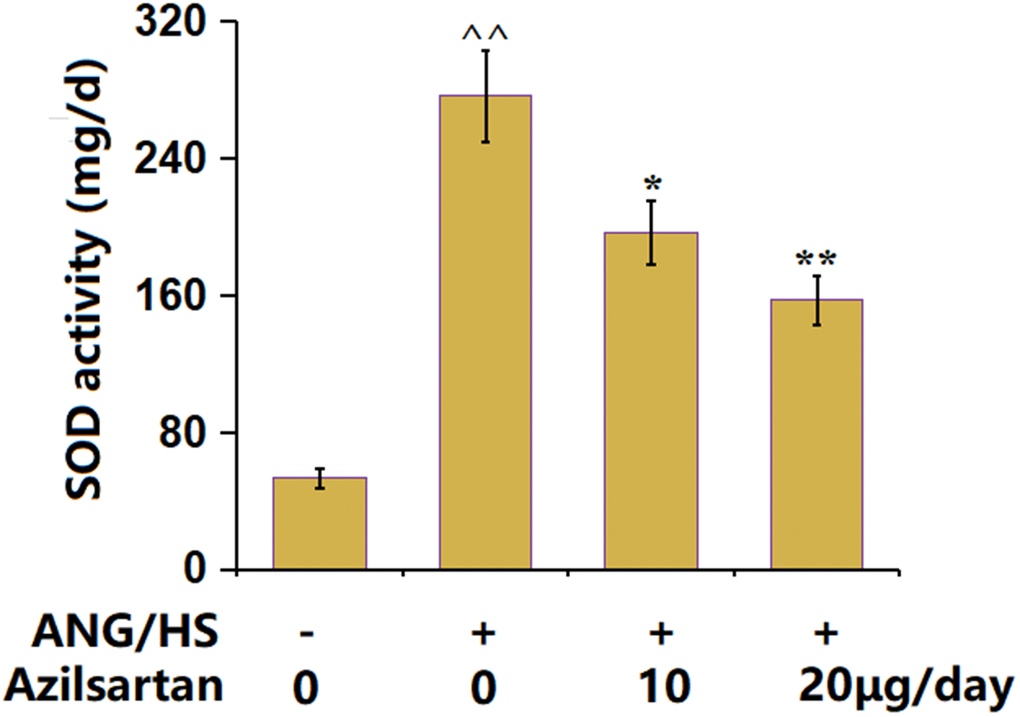
Figure 4. The effects of Azilsartan on urinary albumin excretion in ANG/HS-challenged mice (n=6, ^^, P<0.01 vs. vehicle group; *, **, P<0.05, 0.01 vs. ANG/HS group).
The effects of Azilsartan on the expression of occludin in renal tissues in ANG/HS-challenged mice
Occludin is a critical tight junction (TJ) protein in renal tubular epithelial tissues [24]. Occludin was extremely downregulated in renal tissues of ANG/HS- challenged mice, which was signally rescued by 10 μg/day and 20 μg/day Azilsartan (Figure 5A, 5B), suggesting an elevation property of Azilsartan on the occludin level in renal tissues of ANG/HS-challenged mice.
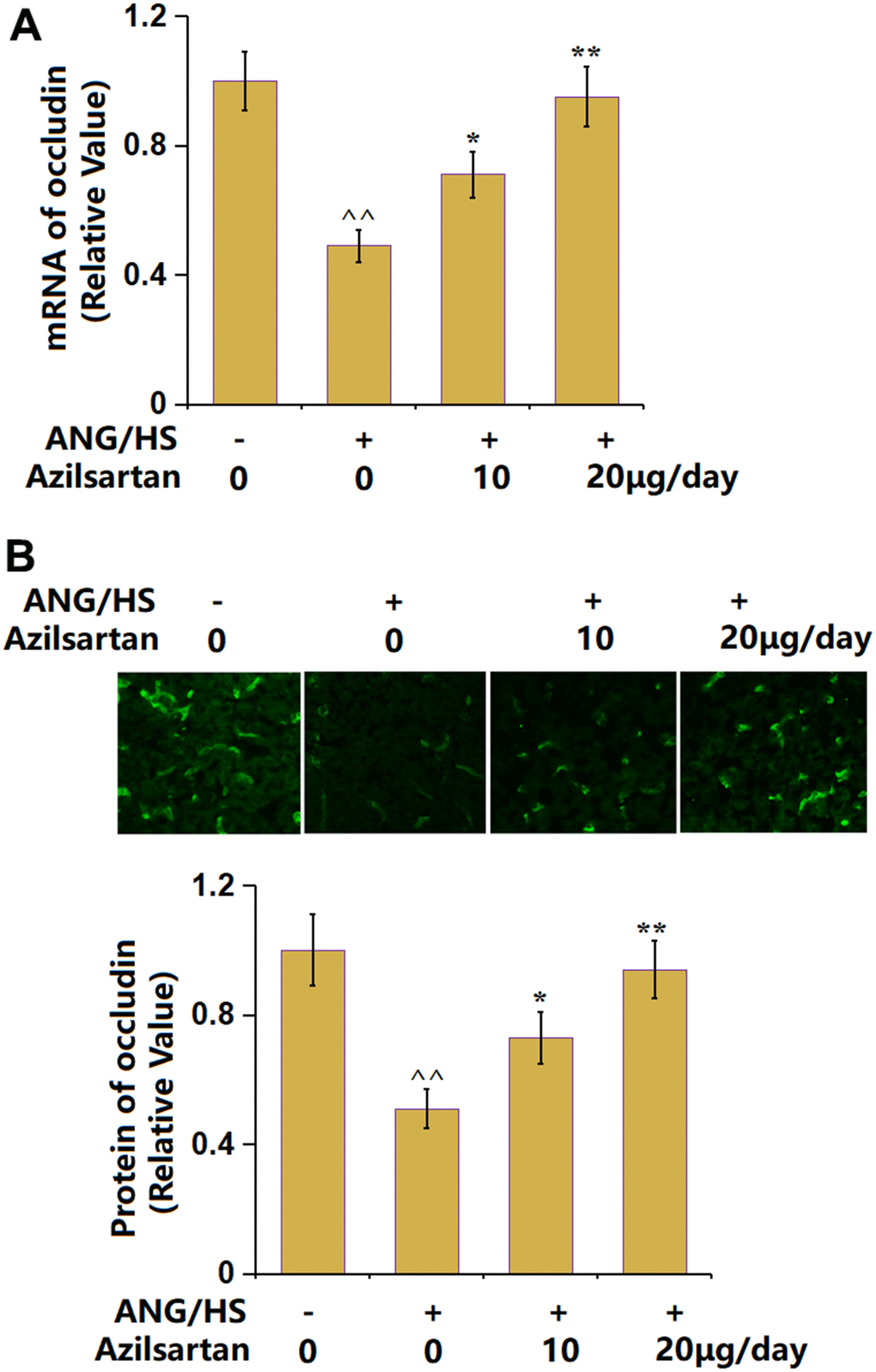
Figure 5. The effects of Azilsartan on the expression of occludin in renal tissues in ANG/HS-challenged mice. (A) mRNA of occludin; (B) Protein of occludin as measured by immunostaining. Scale bar, 100 μm (n=5 or 6, ^^, P<0.01 vs. vehicle group; *, **, P<0.05, 0.01 vs. ANG/HS group).
Azilsartan ameliorated ANG/HS-induced increases in endothelial monolayer permeability in HrGECs
To probe the functional mechanism, an HrGECs monolayer was constructed and treated with ANG/HS with or without Azilsartan (2.5, 5 μM) for 24 hours. The fluorescence intensity of FITC-dextran in the ANG/HS-challenged HrGECs monolayer (Figure 6A) was increased from 16.3% to 45.6%, which was signally decreased to 33.5% and 27.6% by 2.5 and 5 μM Azilsartan, respectively. Furthermore, the TEER values (Figure 6B) in the control, ANG/HS, 2.5 μM Azilsartan, and 5 μM Azilsartan groups were 91.5, 51.2, 69.8, and 83.5 Ωcm2, respectively. Moreover, the occludin level (Figure 6C, 6D) in ANG/HS- challenged HrGECs was markedly repressed, but observably rescued by 2.5 and 5 μM Azilsartan. The increased endothelial monolayer permeability in HrGECs was ameliorated by Azilsartan.
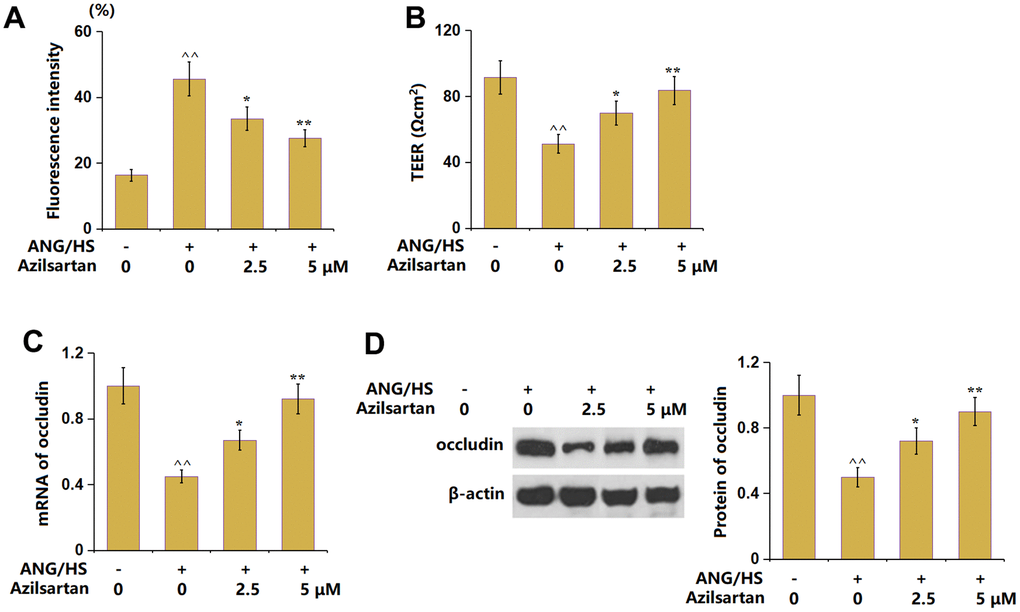
Figure 6. Azilsartan ameliorates ANG/HS-induced increase in endothelial monolayer permeability in human renal glomerular endothelial cells (HrGECs). HrGEC monolayer was treated with ANG/HS with or without Azilsartan (2.5, 5 μM) for 24 hours. (A) Fluorescence intensity of FITC-dextran; (B) Trans-endothelial electrical resistance (TEER) was assayed; (C) mRNA of occludin; (D) Protein levels of occludin (n=5, ^^, P<0.01 vs. vehicle group; *, **, P<0.05, 0.01 vs. ANG/HS group).
Azilsartan prevents ANG/HS-induced reduction in KLF2 in HrGECs
KLF2 is a regulatory transcriptional factor mediating TJ protein levels [25]. KLF2 (Figure 7) was found memorably downregulated in the ANG/HS-challenged HrGECs monolayer but signally upregulated by 2.5 and 5 μM Azilsartan, implying an involvement of KLF2 in the function of Azilsartan.
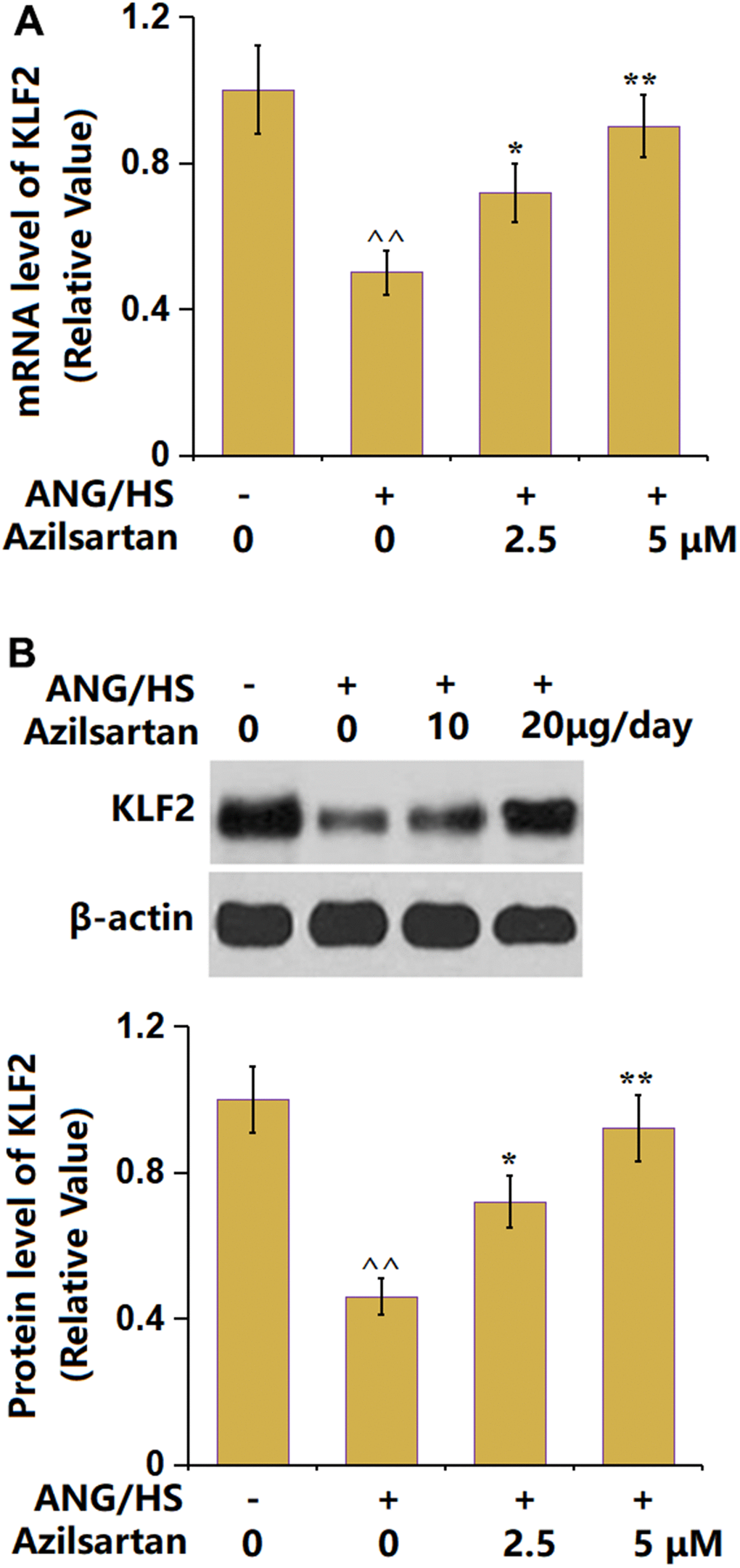
Figure 7. Azilsartan prevents ANG/HS-induced reduction in KLF2 in human renal glomerular endothelial cells (HrGECs). HrGEC monolayer was treated with ANG/HS with or without Azilsartan (2.5, 5 μM) for 24 hours. (A) mRNA level of KLF2; (B) Protein level of KLF2 (n=6, ^^, P<0.01 vs. vehicle group; *, **, P<0.05, 0.01 vs. ANG/HS group).
Silencing of KLF2 abolished the protective effects of Azilsartan against ANG/HS-induced aggravation of endothelial permeability in HrGECs
To provide evidence for the involvement of KLF2 in the function of Azilsartan, the HrGECs monolayer was transduced with lentiviral KLF2 shRNA, followed by stimulation with ANG/HS with or without Azilsartan (5 μM) for 24 hours. The silencing of KLF2 in HrGECs was identified by the results shown in Figure 8A. The declined occludin level observed in ANG/HS- challenged HrGECs was markedly elevated by Azilsartan, which was signally reversed by knocking down KLF2 (Figure 8B). Furthermore, the fluorescence intensity of FITC-dextran in the ANG/HS-challenged HrGECs monolayer (Figure 8C) was promoted from 16.8% to 46.9%, which was notably reduced to 26.6% by Azilsartan. After the knockdown of KLF2, the fluorescence intensity was reversed to 47.2%.
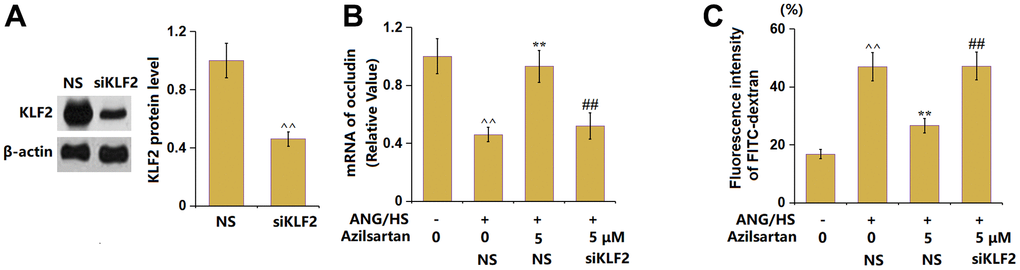
Figure 8. Silencing of KLF2 abolished the protective effects of Azilsartan against ANG/HS-induced aggravation of endothelial permeability in HrGECs. HrGEC monolayer was transduced with lentiviral KLF2 shRNA, followed by stimulation with ANG/HS with or without Azilsartan (5 μM) for 24 hours. (A) Western blot revealed successful knockdown of KLF2; (B) mRNA level of occludin; (C) Fluorescence intensity of FITC-dextran (n=6, ^^, P<0.01 vs. vehicle group; **, P<0.01 vs. ANG/HS group; ##, P<0.01 vs. ANG/HS+ Azilsartan group).
Discussion
Previous studies on HRD mainly focused on the glomerulus. However, recent researches have claimed that the degree of renal tubulointerstitial damage is more associated with renal function [26]. Some scholars have shown that reducing the functional damage of renal tubular epithelial cells is beneficial for the repair of renal damage, thereby reducing or partially inhibiting renal interstitial fibrosis [27]. OS affects hypertension rats by aggravating the severity of renal damage, suggesting that OS shows an intrinsic and inevitable relationship with hypertension [28]. Renal tubular dysfunction is reported to be relevant to OS activation. A sharp decline of SOD activity is triggered by accumulated ROS, while MDA is increased substantially and NO is inactivated, resulting in the reduced bioavailability of NO. After the oxidation of arachidonic acid in the kidney, a large number of prostaglandins are synthesized, which has a direct vasoconstriction effect to further aggravate renal damages [29]. Similar to data reported by Bakri [30], extremely increased water intake, food intake, and arterial pressure were observed in ANG/HS-challenged mice, all of which were signally rescued by Azilsartan, suggesting an alleviative effect of Azilsartan on ANG/HS-induced hypertension in mice. Furthermore, activated OS and enhanced cytokine production were observed in ANG/HS- challenged mice, which were also claimed in cardiac tissues of ANG/HS-challenged mice by Yang [31]. The curative function of Azilsartan on ANG/HS- challenged mice was accompanied by the alleviation of OS and inflammation, implying a correction between the function of Azilsartan and OS inhibition.
Epithelial junction complexes include TJs, adhere junctions (AJs), and gap junctions. TJs are located at the top of all junction complexes, with AJs located underneath for the apical junction complex [32]. TJs are characterized as the banded structure circled epithelial cells, which divide membranes into the apical and basolateral parts. The polarity, proliferation, and differentiation of epithelial cells can be regulated by proteins in TJs [33]. Occludin is the first transmembrane protein discovered in TJs, which participates in maintaining the permeability of TJs and the integrity of the epithelial cell barrier [34]. In ANG/HS- challenged mice, consistent with Bakri’s report [30], an increased UAE was observed, suggesting disruption of the integrity of the epithelial cell barrier. Furthermore, the occludin level was found to be reduced, which explained the increased TJs permeability. The curative function of Azilsartan on ANG/HS-challenged mice was accompanied by reduced UAE and increased occludin levels, implying that the function of Azilsartan was correlated with the integrity of epithelial cell barrier. Our in vitro study revealed that the permeability of the HrGECs monolayer was increased by ANG/HS, also observed in high glucose-stimulated HrGECs [35]. The reparative effect of Azilsartan on the permeability of the HrGECs monolayer was accompanied by the increased occludin level, implying that the function of Azilsartan was correlated with the upregulation of occludin.
KLF2 was discovered by Anderson in 1995 and is mainly expressed in lung tissues [36]. As a member of the KLF family, KLF2 has the common structural features of the family, including a DNA zinc finger binding domain, transcription activation domain, and autoinhibitory domain, among which the autoinhibitory domain is adjacent to the zinc finger domain [37]. Studies have found that KLF2 has specific functions in the maintenance of normal vascular function, adipocyte differentiation, and stemness maintenance of embryonic stem cells [38, 39]. Recently, KLF2 has been claimed to regulate the level of TJ proteins, including occludin [40]. In our research, the KLF2 level was found to be extremely reduced in ANG/HS- challenged HrGECs, then rescued by Azilsartan, implying that the function of Azilsartan was correlated with the upregulation of KLF2. Furthermore, the influence of Azilsartan on the occludin level and endothelial permeability were abrogated by the silencing of KLF2, which further identified the mediation of KLF2 in the function of Azilsartan. In future studies, the direct target of Azilsartan in HrGECs will be probed to fully understand the functional mechanism of Azilsartan in treating HRD.
Collectively, Azilsartan alleviated ANG/HS-induced increase in UAE mediated by the KLF2/occludin axis.
Author Contributions
Jun Cao, Dandan Zhang, and Shaofeng Xie designed the study. Jun Cao, Dandan Zhang, Wenfeng Li, Wenjin Yuan, and Gang Luo performed the experiments. Shaofeng Xie prepared the manuscript. Jun Cao and Dandan Zhang reviewed the manuscript.
Conflicts of Interest
There is no conflicts of interest for all authors.
Ethical Statement
All animal experiments were performed in compliance with the China animal welfare law. Experimental protocols were approved by the Ethical Committee of the People’s Hospital of Ganzhou (Approval No. 180065).
Funding
This study was funded by “People’s Hospital of Ganzhou”.
References
- 1. Rodriguez-Iturbe B, Pons H, Johnson RJ. Role of the Immune System in Hypertension. Physiol Rev. 2017; 97:1127–64. https://doi.org/10.1152/physrev.00031.2016 [PubMed]
- 2. McMaster WG, Kirabo A, Madhur MS, Harrison DG. Inflammation, immunity, and hypertensive end-organ damage. Circ Res. 2015; 116:1022–33. https://doi.org/10.1161/CIRCRESAHA.116.303697 [PubMed]
- 3. Meguid El Nahas A, Bello AK. Chronic kidney disease: the global challenge. Lancet. 2005; 365:331–40. https://doi.org/10.1016/S0140-6736(05)17789-7 [PubMed]
- 4. Rucker AJ, Rudemiller NP, Crowley SD. Salt, Hypertension, and Immunity. Annu Rev Physiol. 2018; 80:283–307. https://doi.org/10.1146/annurev-physiol-021317-121134 [PubMed]
- 5. Touyz RM, Rios FJ, Alves-Lopes R, Neves KB, Camargo LL, Montezano AC. Oxidative Stress: A Unifying Paradigm in Hypertension. Can J Cardiol. 2020; 36:659–70. https://doi.org/10.1016/j.cjca.2020.02.081 [PubMed]
- 6. Cuadrado A, Manda G, Hassan A, Alcaraz MJ, Barbas C, Daiber A, Ghezzi P, León R, López MG, Oliva B, Pajares M, Rojo AI, Robledinos-Antón N, et al. Transcription Factor NRF2 as a Therapeutic Target for Chronic Diseases: A Systems Medicine Approach. Pharmacol Rev. 2018; 70:348–83. https://doi.org/10.1124/pr.117.014753 [PubMed]
- 7. Daenen K, Andries A, Mekahli D, Van Schepdael A, Jouret F, Bammens B. Oxidative stress in chronic kidney disease. Pediatr Nephrol. 2019; 34:975–91. https://doi.org/10.1007/s00467-018-4005-4 [PubMed]
- 8. Ruilope LM, Bakris GL. Renal function and target organ damage in hypertension. Eur Heart J. 2011; 32:1599–604. https://doi.org/10.1093/eurheartj/ehr003 [PubMed]
- 9. Lu MC, Zhao J, Liu YT, Liu T, Tao MM, You QD, Jiang ZY. CPUY192018, a potent inhibitor of the Keap1-Nrf2 protein-protein interaction, alleviates renal inflammation in mice by restricting oxidative stress and NF-κB activation. Redox Biol. 2019; 26:101266. https://doi.org/10.1016/j.redox.2019.101266 [PubMed]
- 10. Bhattacharya R, Senbanerjee S, Lin Z, Mir S, Hamik A, Wang P, Mukherjee P, Mukhopadhyay D, Jain MK. Inhibition of vascular permeability factor/vascular endothelial growth factor-mediated angiogenesis by the Kruppel-like factor KLF2. J Biol Chem. 2005; 280:28848–51. https://doi.org/10.1074/jbc.C500200200 [PubMed]
- 11. SenBanerjee S, Lin Z, Atkins GB, Greif DM, Rao RM, Kumar A, Feinberg MW, Chen Z, Simon DI, Luscinskas FW, Michel TM, Gimbrone MA
Jr , García-Cardeña G, Jain MK. KLF2 Is a novel transcriptional regulator of endothelial proinflammatory activation. J Exp Med. 2004; 199:1305–15. https://doi.org/10.1084/jem.20031132 [PubMed] - 12. Niu N, Xu S, Xu Y, Little PJ, Jin ZG. Targeting Mechanosensitive Transcription Factors in Atherosclerosis. Trends Pharmacol Sci. 2019; 40:253–66. https://doi.org/10.1016/j.tips.2019.02.004 [PubMed]
- 13. Dharmashankar K, Widlansky ME. Vascular endothelial function and hypertension: insights and directions. Curr Hypertens Rep. 2010; 12:448–55. https://doi.org/10.1007/s11906-010-0150-2 [PubMed]
- 14. Bae E, Yu MY, Moon JJ, Kim JE, Lee S, Han SW, Park DJ, Kim YS, Yang SH. Renoprotective Effect of KLF2 on Glomerular Endothelial Dysfunction in Hypertensive Nephropathy. Cells. 2022; 11:762. https://doi.org/10.3390/cells11050762 [PubMed]
- 15. Al-Majed AA, Bakheit AHH, Al-Muhsin A, Al-Kahtani HM, Abdelhameed AS. Azilsartan medoxomil. Profiles Drug Subst Excip Relat Methodol. 2020; 45:1–39. https://doi.org/10.1016/bs.podrm.2019.10.001 [PubMed]
- 16. Hou N, Li LR, Shi YY, Yuan WC, Zhao GJ, Liu XW, Cai SA, Huang Y, Zhan HX, Pan WB, Luo CF. Azilsartan ameliorates ventricular hypertrophy in rats suffering from pressure overload-induced cardiac hypertrophy by activating the Keap1-Nrf2 signalling pathway. J Pharm Pharmacol. 2021; 73:1715–25. https://doi.org/10.1093/jpp/rgab097 [PubMed]
- 17. Abdelsaid M, Coucha M, Ergul A. Cerebrovasculoprotective effects of azilsartan medoxomil in diabetes. Transl Res. 2014; 164:424–32. https://doi.org/10.1016/j.trsl.2014.06.003 [PubMed]
- 18. Hye Khan MA, Neckář J, Haines J, Imig JD. Azilsartan improves glycemic status and reduces kidney damage in zucker diabetic fatty rats. Am J Hypertens. 2014; 27:1087–95. https://doi.org/10.1093/ajh/hpu016 [PubMed]
- 19. Dong Q, Li Y, Chen J, Wang N. Azilsartan Suppressed LPS-Induced Inflammation in U937 Macrophages through Suppressing Oxidative Stress and Inhibiting the TLR2/MyD88 Signal Pathway. ACS Omega. 2020; 6:113–8. https://doi.org/10.1021/acsomega.0c03655 [PubMed]
- 20. Liu H, Mao P, Wang J, Wang T, Xie CH. Azilsartan, an angiotensin II type 1 receptor blocker, attenuates tert-butyl hydroperoxide-induced endothelial cell injury through inhibition of mitochondrial dysfunction and anti-inflammatory activity. Neurochem Int. 2016; 94:48–56. https://doi.org/10.1016/j.neuint.2016.02.005 [PubMed]
- 21. Thomas JM, Ling YH, Huuskes B, Jelinic M, Sharma P, Saini N, Ferens DM, Diep H, Krishnan SM, Kemp-Harper BK, O’Connor PM, Latz E, Arumugam TV, et al. IL-18 (Interleukin-18) Produced by Renal Tubular Epithelial Cells Promotes Renal Inflammation and Injury During Deoxycorticosterone/Salt-Induced Hypertension in Mice. Hypertension. 2021; 78:1296–309. https://doi.org/10.1161/HYPERTENSIONAHA.120.16437 [PubMed]
- 22. Rom S, Dykstra H, Zuluaga-Ramirez V, Reichenbach NL, Persidsky Y. miR-98 and let-7g* protect the blood-brain barrier under neuroinflammatory conditions. J Cereb Blood Flow Metab. 2015; 35:1957–65. https://doi.org/10.1038/jcbfm.2015.154 [PubMed]
- 23. Lin Z, Kumar A, SenBanerjee S, Staniszewski K, Parmar K, Vaughan DE, Gimbrone MA
Jr , Balasubramanian V, García-Cardeña G, Jain MK. Kruppel-like factor 2 (KLF2) regulates endothelial thrombotic function. Circ Res. 2005; 96:e48–57. https://doi.org/10.1161/01.RES.0000159707.05637.a1 [PubMed] - 24. Kim S, Kim GH. Roles of claudin-2, ZO-1 and occludin in leaky HK-2 cells. PLoS One. 2017; 12:e0189221. https://doi.org/10.1371/journal.pone.0189221 [PubMed]
- 25. Li W, Wang C, Zhang D, Zeng K, Xiao S, Chen F, Luo J. Azilsartan ameliorates ox-LDL-induced endothelial dysfunction via promoting the expression of KLF2. Aging (Albany NY). 2021; 13:12996–3005. https://doi.org/10.18632/aging.202973 [PubMed]
- 26. Nangaku M. Mechanisms of tubulointerstitial injury in the kidney: final common pathways to end-stage renal failure. Intern Med. 2004; 43:9–17. https://doi.org/10.2169/internalmedicine.43.9 [PubMed]
- 27. Huang H, Ding R, Chen Z, Yi Z, Wang H, Lv Y, Bao E. Goose nephritic astrovirus infection increases autophagy, destroys intercellular junctions in renal tubular epithelial cells, and damages podocytes in the kidneys of infected goslings. Vet Microbiol. 2021; 263:109244. https://doi.org/10.1016/j.vetmic.2021.109244 [PubMed]
- 28. Beswick RA, Dorrance AM, Leite R, Webb RC. NADH/NADPH oxidase and enhanced superoxide production in the mineralocorticoid hypertensive rat. Hypertension. 2001; 38:1107–11. https://doi.org/10.1161/hy1101.093423 [PubMed]
- 29. Manning RD
Jr , Tian N, Meng S. Oxidative stress and antioxidant treatment in hypertension and the associated renal damage. Am J Nephrol. 2005; 25:311–7. https://doi.org/10.1159/000086411 [PubMed] - 30. Bakri M, Yi Y, Chen LD, Aisa HA, Wang MH. Alkaloids of Nitraria sibirica Pall. decrease hypertension and albuminuria in angiotensin II-salt hypertension. Chin J Nat Med. 2014; 12:266–72. https://doi.org/10.1016/S1875-5364(14)60053-4 [PubMed]
- 31. Yang S, Chen D, Chen F, Zhao X, Zhang Y, Li Z, Jin L, Xu Y, Sanchis D, Ye J. Deletion of protein kinase B2 preserves cardiac function by blocking interleukin-6-mediated injury and restores blood pressure during angiotensin II/high-salt-diet-induced hypertension. J Hypertens. 2018; 36:834–46. https://doi.org/10.1097/HJH.0000000000001613 [PubMed]
- 32. Dokladny K, Zuhl MN, Moseley PL. Intestinal epithelial barrier function and tight junction proteins with heat and exercise. J Appl Physiol (1985). 2016; 120:692–701. https://doi.org/10.1152/japplphysiol.00536.2015 [PubMed]
- 33. Lee DB, Huang E, Ward HJ. Tight junction biology and kidney dysfunction. Am J Physiol Renal Physiol. 2006; 290:F20–34. https://doi.org/10.1152/ajprenal.00052.2005 [PubMed]
- 34. Morrow CM, Mruk D, Cheng CY, Hess RA. Claudin and occludin expression and function in the seminiferous epithelium. Philos Trans R Soc Lond B Biol Sci. 2010; 365:1679–96. https://doi.org/10.1098/rstb.2010.0025 [PubMed]
- 35. Xu L, Shao F. Sitagliptin protects renal glomerular endothelial cells against high glucose-induced dysfunction and injury. Bioengineered. 2022; 13:655–66. https://doi.org/10.1080/21655979.2021.2012550 [PubMed]
- 36. Anderson KP, Kern CB, Crable SC, Lingrel JB. Isolation of a gene encoding a functional zinc finger protein homologous to erythroid Krüppel-like factor: identification of a new multigene family. Mol Cell Biol. 1995; 15:5957–65. https://doi.org/10.1128/MCB.15.11.5957 [PubMed]
- 37. Conkright MD, Wani MA, Lingrel JB. Lung Krüppel-like factor contains an autoinhibitory domain that regulates its transcriptional activation by binding WWP1, an E3 ubiquitin ligase. J Biol Chem. 2001; 276:29299–306. https://doi.org/10.1074/jbc.M103670200 [PubMed]
- 38. Dabravolski SA, Sukhorukov VN, Kalmykov VA, Grechko AV, Shakhpazyan NK, Orekhov AN. The Role of KLF2 in the Regulation of Atherosclerosis Development and Potential Use of KLF2-Targeted Therapy. Biomedicines. 2022; 10:254. https://doi.org/10.3390/biomedicines10020254 [PubMed]
- 39. Jha P, Das H. KLF2 in Regulation of NF-κB-Mediated Immune Cell Function and Inflammation. Int J Mol Sci. 2017; 18:2383. https://doi.org/10.3390/ijms18112383 [PubMed]
- 40. Han J, Tang H, Yao L, Jin E, Pan W, Chen S. Azilsartan protects against hyperglycemia-induced hyperpermeability of the blood-brain barrier. Bioengineered. 2021; 12:3621–33. https://doi.org/10.1080/21655979.2021.1948950 [PubMed]