



Introduction
Acute myeloid leukemia (AML) is a highly malignant disease with a poor survival rate despite continuous advances in treatment [1]. Advances in molecular biology and multiomics technology have increased the knowledge of how genes control physiological functions involved in the modulation of specific phenotypes and drug efficacy, and provide highly informative data for studying the molecular state of disease biology [2, 3]. Increasingly omics data have promoted the exploration of the interconnections across molecular layers as well as investigation of AML pathogenesis and identification of underlying diagnostic and therapeutic molecules.
Heat shock factors (HSFs) are a class of evolutionarily conserved transcription factors, that regulate the heat shock response (HSR) by mediating the expression pattern of heat shock proteins (HSPs) [4, 5]. HSPs are observed to be increased in a wide variety of cancers and are involved in almost every aspect of cancer development [6, 7]. HSP90 overexpression has been reported in AML, and HSP90 inhibitors have been confirmed to be effective agents against primary AML blasts [8]. The transcriptional activation of the HSPs is under the control of HSFs. Four HSFs HSF1, HSF2, HSF4 and HSF5, exist in humans, and HSF1 is the primary activator in human tissues [9]. HSF1, localized to chromosome 8q24.3, prevents cancer cell death when exposed to stressors, which indispensable for various malignant transformation–related signaling pathways, thereby facilitating cancer cell proliferation and migration and affecting the tumor microenvironment [10]. Additionally, significant correlations between HSF1 and immune checkpoint genes and the levels of B cells, CD8+ T cells, CD4+ T cells, macrophages, neutrophils, and dendritic cells have been detected in multiple kinds of cancers [11]. There are studies showing that AML molecular features correlate with clinicopathological characteristics and prognosis [12]. Emerging evidence has demonstrated that the upregulation of HSF1 is associated with poor overall survival (OS) in multiple cancers, suggesting that HSF1 has the potential to serve as an independent prognostic biomarker [13–15].
Previous research on HSP expression from both the stress response and cancer fields involves HSF1. A study confirmed that HSF1 is elevated in AML cell lines including HL–60 and OCI–AML3, acting as a promoter of cell proliferation [16]. Additionally, the expression level of HSF1 was observed to be enhanced in chronic lymphocytic leukemia (CLL) and decreased in patients responding to in the rituximab–bendamustine protocol [17], which supports its potential to be a biomarker for efficacy prediction. These results demonstrated that HSF1 is involved in leukemogenesis and has the potential to be a novel biomarker but its clinical significance and potential function in AML have not been fully investigated.
In this study, the expression level, diagnostic efficiency, and prognostic significance of HSF1 in AML, as well as the correlations between HSF1 and clinical parameters, were investigated and validated based on The Cancer Genome Atlas (TCGA), Genotype–Tissue Expression (GTEx), Gene Expression Omnibus (GEO), Cancer Cell Line Encyclopedia (CCLE), and Beat AML data viewer (Vizome). Then, the potential mechanism of HSF1 in AML was explored based on annotation of transcriptome and single–cell sequencing data. The molecular regulatory association between HSF1 and its related genes was explored by constructing a protein–protein interaction network. Additionally, the correlation between HSF1 expression and immune cell subset abundance was further explored. This study aimed to elucidate the role of HSF1 in AML at a systems level in order to explore the clinical implications of HSF1 and the relevant molecular mechanisms in AML pathogenesis.
Results
HSF1 expression is elevated in AML and indicates poor prognosis
The HSF family in humans includes HSF1, HSF2, HSF4 and HSF5. According to data from AML based on TCGA and healthy controls derived from GTEx, HSF1 and HSF5 were elevated, while HSF2 and HSF3 were decreased in AML compared to healthy controls (Figure 1A, 1D, 1J and 1G). Additionally, HSF1 was highly expressed in most of 21 leukemia cell lines, especially the acute myeloid leukemia cell line KG–1 (Figure 1M). The receiver operating characteristic (ROC) curve analyses showed good discriminative ability of HSF1, area under the curve (AUC) is 0.723, (Figure 1C), HSF2 (AUC = 0.737), HSF4 (AUC = 0.889), and HSF5 (AUC = 0.998) (Figure 1F, 1I, and 1L). There were statistically significant changes in the expression of all HSF members, but only HSF1 had marked prognostic significance (Figure 1B, 1E, 1H and 1K). A high expression level of HSF1 predicts poor OS, which supports the idea that HSF1 is a prognostic marker for poor outcomes. The expression level and diagnostic efficacy were validated in replication datasets. In GSE9476, HSF1 was validated to be highly expressed in 26 AML patients compared to 38 healthy controls, and AUC was 0.756 (Figure 2A and 2B). The association of high HSF1 expression with a poor prognosis for AML was validated using GSE12417, and data from 163 patients were included (Figure 2C).
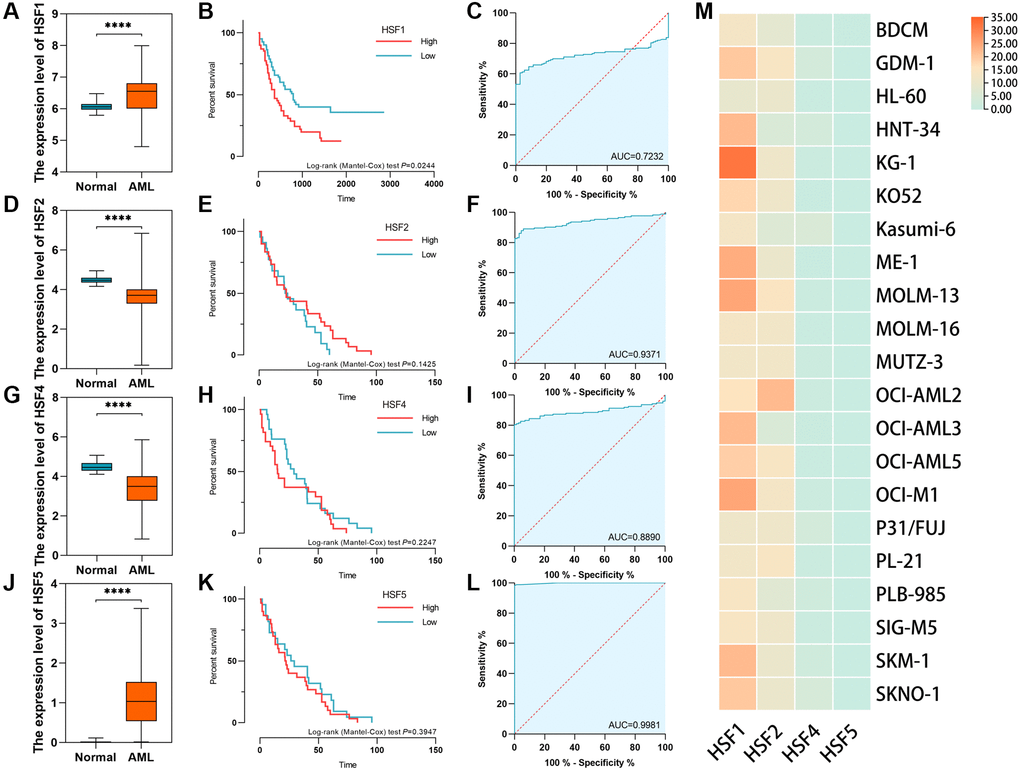
Figure 1. The expression level, diagnostic efficacy and prognostic significance of HSF family in acute myeloid leukemia (AML) based on data from the cancer genome atlas (TCGA) and the genotype–tissue expression (GETx) processed using the TOIL pipeline. The validation datasets were downloaded from the Gene Expression Omnibus (GEO), and Cancer Cell Line Encyclopedia (CCLE). (A–C) The expression level, diagnostic efficacy and prognostic significance of HSF1 in AML. (D–F) The expression level, diagnostic efficacy and prognostic significance of HSF2 in AML. (G–I) The expression level, diagnostic efficacy and prognostic significance of HSF4 in AML. (J–L) The expression level, diagnostic efficacy and prognostic significance of HSF5 in AML. (M) The expression level of HSFs examined in leukemia cell lines. ****P < 0.001.
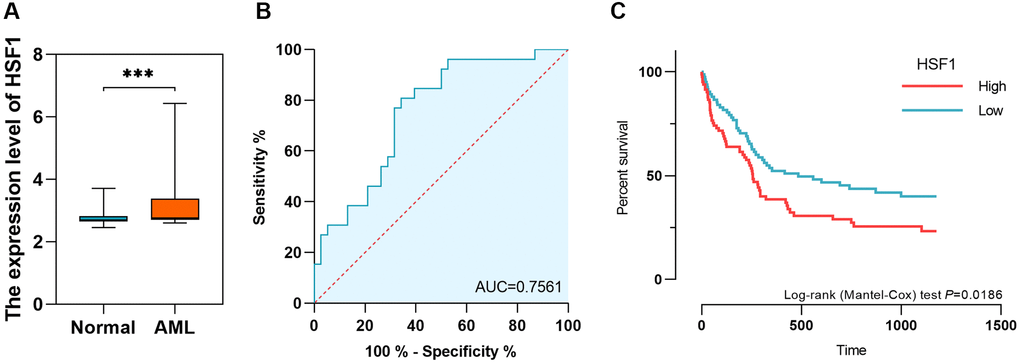
Figure 2. The expression level, diagnostic value and prognostic significance of HSF1 in AML based on data from gene expression omnibus (GEO). (A and B) The expression level and diagnostic value of HSF1 in AML based on data from GSE9476. (C) The prognostic significance of HSF1 based on data from GSE12417. ***P < 0.01.
Clinical significance of HSF1 expression in AML
A logistic model was constructed to explore the association between HSF1 expression and clinical parameters of AML patients, and the results showed that HSF1 is significantly associated with patient age and peripheral blood (PB) blast proportion based on data from TCGA. Evidence from the Vizome indicated that HSF1 is significantly associated with patient age (Table 1). In both TCGA and Vizome, age was a significant associated factor for HSF1, but HSF1 bore no relationship to survival in the age subgroup. In subgroup of bone marrow (BM) blast proportion (%) >20, HSF1 was significantly associated with patient survival based on data from TCGA, which was also validated in the replication cohort based on data from Vizome (Figure 3).
Table 1. Association between HSF1 expression and clinical parameters.
| Characteristics | TCGA (n = 149) | LAML (n = 445) | ||
| OR | P value | OR | P value | |
| Gender | 1.140 (0.600–2.171) | 0.689 | 1.104 (0.650–1.875) | 0.713 |
| Age | 2.000 (1.041–3.894) | 0.039 | 1.027 (1.011,1.043) | 0.001 |
| WBC count (×109/L) | 1.306 (0.688–2.491) | 0.414 | 1.002 (0.996–1.007) | 0.573 |
| BM blast proportion (%) | 1.276 (0.664–2.461) | 0.465 | 0.994 (0.982–1.007) | 0.364 |
| PB blast proportion (%) | 2.722 (1.419–5.316) | 0.003 | 1.002 (0.990–1.014) | 0.740 |
| Cytogenetic risk (Poor vs. Favorable and Intermediate) | 1.843 (0.865–4.035) | 0.117 | 0.786 (0.433–1.427) | 0.429 |
| FLT3 mutation (Positive vs. Negative) | 1.755 (0.866–3.621) | 0.122 | 0.825 (0.433–1.573) | 0.560 |
| NPM1 mutation (Positive vs. Negative) | 1.044 (0.481–2.277) | 0.912 | 0.757 (0.386–1.486) | 0.429 |
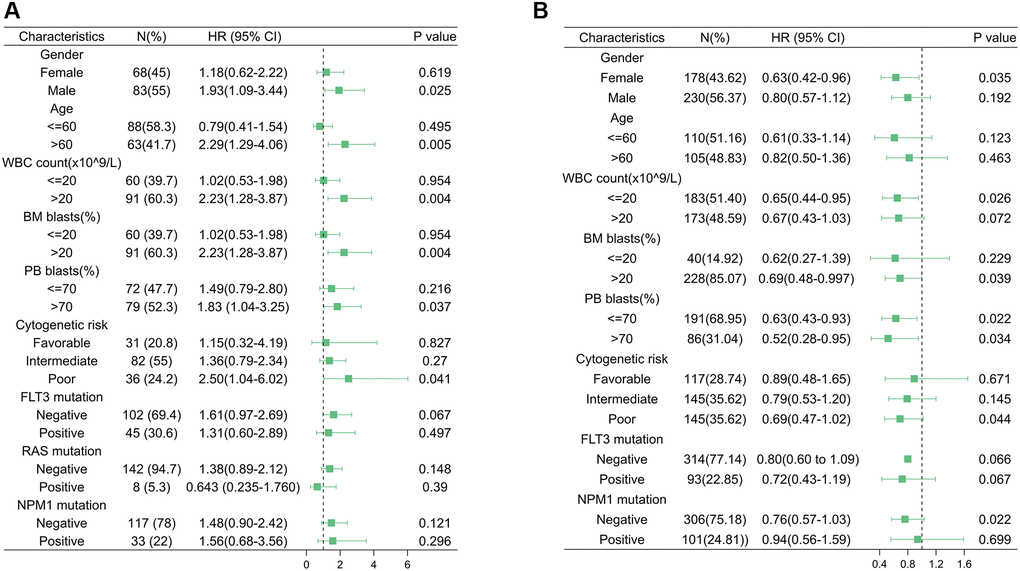
Figure 3. Acute myeloid leukemia subgroup overall survival analyses. Forest plot of Cox analyses of HSF1 for different subgroups based on data from The Cancer Genome Atlas (TCGA) (A) and Vizome (B).
Functional analyses of HSF1 in AML
The top 50 negatively and positively correlated genes related to HSF1 expression in AML were obtained from linkedomics (Figure 4A). Correlation analyses of HSF1–associated genes were conducted using GeneMANIA, and the results showed that 71.16% are annotated as co–expressed, 16.68% are annotated as physical interactions, 4.44% are annotated as genetic interactions, 1.77% are annotated as co–localization and 0.11% of genes are shared protein domains (Figure 4B). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses of HSF1–associated genes showed that HSF1 is involved in the AMPK signaling pathway, Hippo signaling pathway, NF–kappa B signaling pathway, TGF–βsignaling pathway, and TNF signaling pathway (Figure 5B). CancerSEA analyses showed that the functional phenotypes of HSF1 in AML were significantly positively correlated with metastasis, angiogenesis, and differentiation inflammation and negatively correlated with DNA repair (Figure 6). The Gene Set Enrichment Analysis (GSEA) results showed that HSF1 was significantly related to HALLMARK INFLAMMATORY RESPONSE, HALLMARK INTERFERON GAMMA RESPONSE, HALLMARK ANGIOGENESIS, and HALLMARK KRAS SIGNALING in AML (Figure 5A).
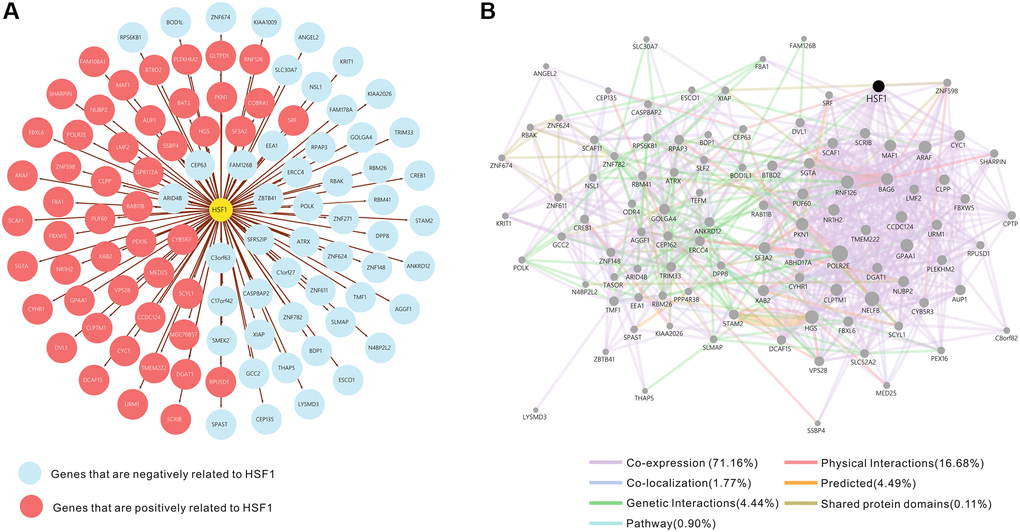
Figure 4. The fifty genes with the strongest positive and negative correlations with HSF1. (A) Network of HSF1 and correlated genes. Positively correlated genes are marked in red. Negatively correlated genes are marked in blue. (B) Network analysis of HSF1 and correlated genes using GeneMANIA.
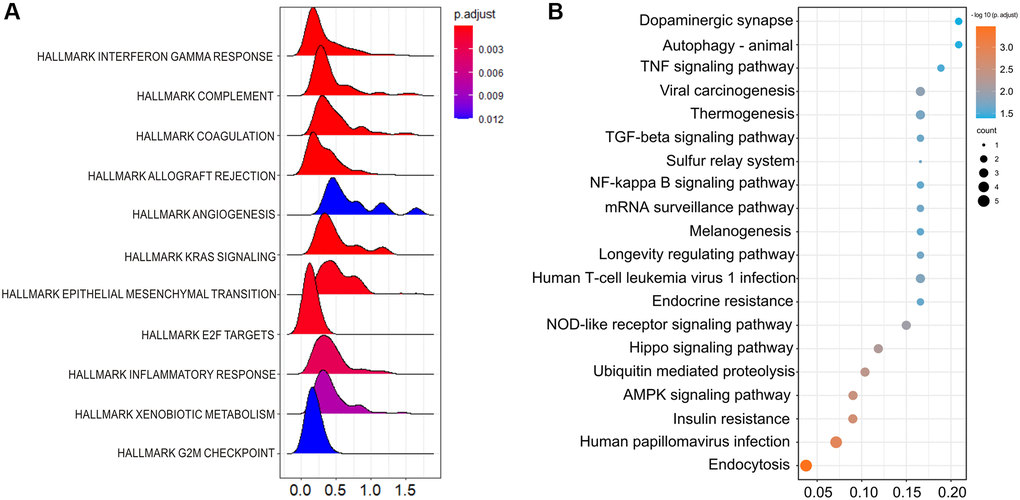
Figure 5. The underlying mechanisms related to HSF1 are shown. (A) Gene Set Enrichment Analysis (GSEA) of the deferentially expressed genes between the HSF1 high–expression versus HSF1 low–expression group in AML. (B) Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis of HSF1 related genes.
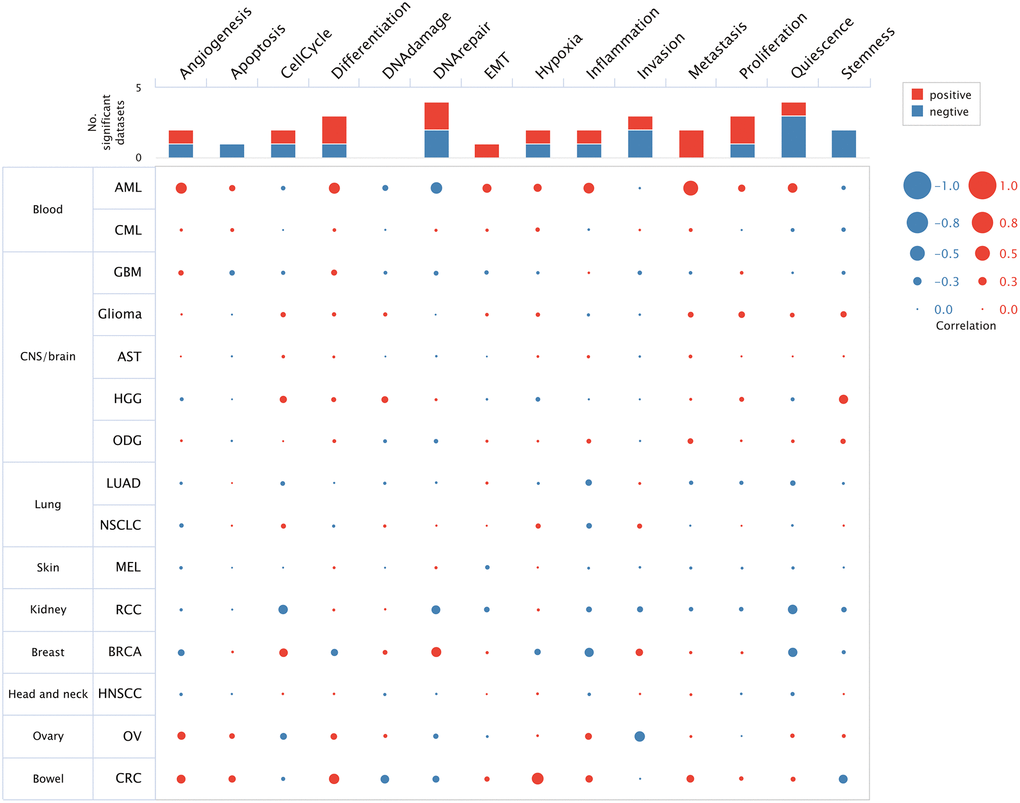
Figure 6. The function of HSF1 was analyzed at single–cell level based on the CancerSEA database.
Immune infiltration analyses
Based on single–sample Gene Set Enrichment Analysis (ssGSEA), correlation between HSF1 expression and immune cell infiltration was analyzed. HSF1 expression was significantly related to the abundance of infiltrating immune cells in AML including the T cell population, nature killer (NK) cells, B cells, and immature dendritic cells (iDCs) (Figure 7A). There were significantly correlations between HSF1 expression and infiltrating levels of NK CD56dim cells (Figure 7B), Tregs (Figure 7C), iDCs (Figure 7D), B cells (Figure 7E), T helper (Th) cells (Figure 7F), T central memory (Tcm) cells (Figure 7G) and Th2 cells (Figure 7H).
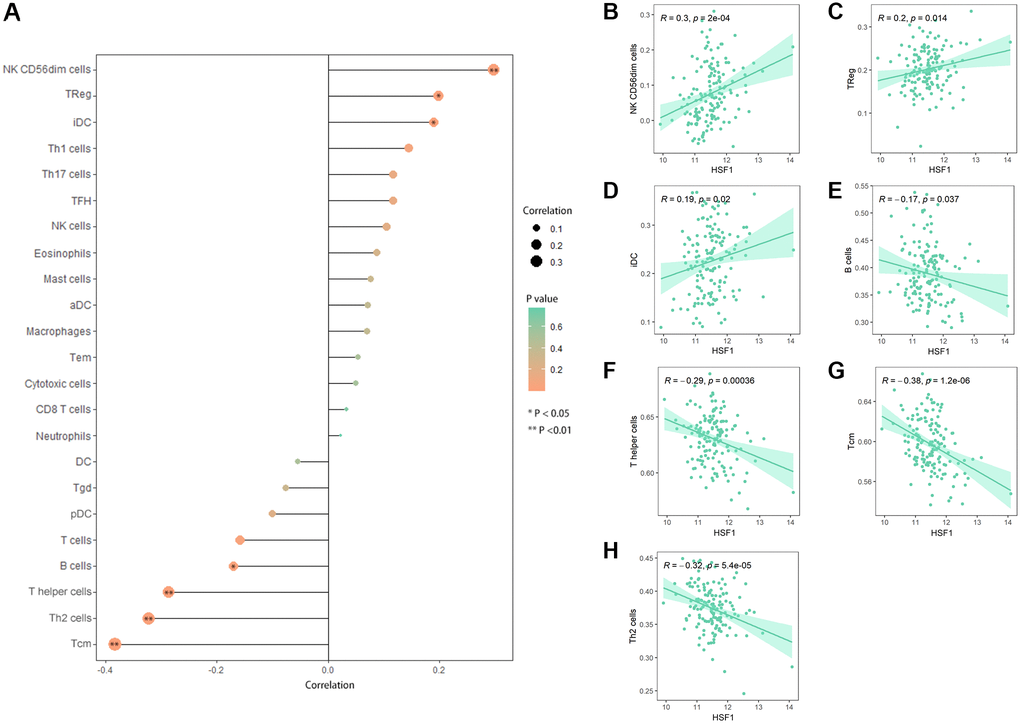
Figure 7. Correlation analyses between HSF1 and immune infiltration. (A) Lollipop plot showing the expression of HSF1 exhibits a strong correlation with the infiltration abundance of NK CD56dim cells (B), Tregs (C), immature DCs (iDCs) (D), B cells (E), T helper (Th) cells (F), T central memory (Tcm) cells (G) and Th2 cells (H). *P < 0.05, **P < 0.01.
Discussion
HSF–induced upregulation of HSPs is a key part of HSR. Among members of the HSF family, HSF1 is reported to be the major regulator of increased HSPs in cancer, and switches on during the development of cancer [18]. Generally, elevated expression of HSF1 has been described in various cancer types [11]. In hematological malignancies, the role of HSF1 has also been partially explored. HSF1 is elevated in Hodgkin lymphoma, multiple myeloma, CLL and AML [16, 19–21]. In multiple myeloma, HSF1 expression has been demonstrated to be related to poor prognosis [19]. Additionally, overexpression of HSF1 in CLL patients is associated with poor prognosis and localized in the nucleus of leukemic B cells [20]. In addition to being highly expressed in AML patient blood samples and AML cell lines, HSF1–pSer326 has been reported to predict the response to bortezomib–containing chemotherapy in pediatric AML [22].
HSF family members expressed in humans include HSF1, HSF2, HSF4 and HSF5, of which expression pattern, diagnostic efficiency and prognostic significance were investigated in this study. HSF1 and HSF5 were highly expressed, while HSF2 and HSF4 were decreased in AML compared to healthy controls, and elevated expression of HSF1 has prognostic significance, indicating a worse survival probability, which has the potential to be a prognosis–predicted biomarker. Further logistic analysis showed that HSF1 expression is correlated to patient age. Previous studies have suggested that elevated HSF1 increases both stress tolerance and lifespan of C. elegans [23, 24]. HSF1 can affect the aging process by upregulating HSP genes, modulating other stress/repair pathways, and influencing signaling systems [25]. Although HSF1 appears to be intimately connected to senescence pathways, the prognostic significance of HSF1 in elderly AML patients remains to be established. Additionally, survival analyses demonstrated that HSF1 predict poor prognosis in BM blast proportion (%) >20 subgroups. Leukemia is characterized by over production of abnormal blast cells in the bone marrow that often spill out into other hematopoietic tissues. The BM blast proportion reflects the malignancy of AML. The result indicated HSF1 might be a prognostic marker in higher malignancy.
It is known that HSF1 exerts its pro–oncogenic effects by mediating HSR–related biological processes in the oncogenic network. To further explore the underlying role of HSF1 in AML pathogenesis, multi-omics analyses were integrated in this study. In the results from single–cell sequencing data form CancerSEA, HSF1 was significantly related to various aspects of leukemogenesis, and a remarkable association was observed for HSF1 and leukemia stem cell metastasis in AML. The correlation between HSF1 expression and metastatic potential has been reported in breast, colon, lung, prostate, hepatocellular tumors and malignant melanoma, indicating that HSF1 supports anchorage–independent growth [26]. In AML, HSF1 may contribute to leukemia stem cells metastasis and is associated with malignant characteristic. Additionally, the functional phenotype of HSF1 is highly implicated in inflammation–related signaling pathways according to the results from the CancerSEA, GSEA and KEGG analyses. Evidence from previous studies has shown that HSF1 is widely involved in immune modulation. HSF1 works as an innate repressor of human immunodeficiency virus–induced inflammation by regulating NF–κB [27]. HSF1 was remarkably correlated with the levels of infiltrating cells and immune checkpoint genes in various cancers [11]. The clinical data indicate that AML risk was significantly associated with multiple autoimmune diseases, and inflammation may impact prognosis in AML patients [28, 29]. Additionally, inflammatory signaling pathways including TNFa/NF–κB, nod–like receptor signaling, TGF–β signaling and the interleukin family have been noted in AML and associated with leukemogenesis [30]. To further explore the correlations between the inflammation state and HSF1, ssGSEA was used to evaluate infiltrated immune cells and HSF1 expression in AML, and the results showed that HSF is significantly correlated with NK cells and T cells. Tumor cells are under the surveillance of immune cells, and disorders of NK cells and T cells–mediated immune responses contribute to leukemogenesis [31, 32]. HSF1 is necessary for optimal T cell proliferation at normal temperatures in the presence of robust stimuli and serves as the predominant regulator of the heat shock response, which represses the cytotoxicity of human NK cells in vitro and in vivo [33, 34]. HSF1 may affect immune cells in order to regulate the immune state in AML and influence leukemia cell growth, metastasis, and AML patient survival, but its mechanisms remain to be validated in vitro and in vivo.
In the present study, we integrate data from several sources of publicly available databases and aimed to systematically investigate the expression and functional roles of HSF1 in AML as well as its clinical implications. The results analyzed based on TCGA were validated using independent datasets sourced from GEO and Vizome. The credibility of the prediction was improved, but some inconsistency exists between different cohorts. Validated results provide an important perspective for understanding the potential role of HSF1 in AML, and contradictory or conflicting results represent open questions for future studies. Additionally, datasets are limited in diversity and sample sizes, large–scale prospective cohort studies should be implemented to warrant the screening value of HSF1 for clinical purposes.
In summary, HSF1 was screened as a potential diagnostic and prognostic biomarker for AML patients. The leukemogenesis mechanisms of HSF1 are a multifactor regulatory process that is involved in inflammatory states, leukemia stem cell metastasis and related signaling pathways. This study comprehensively elucidated the clinical significance and potential functions of HSF1, which provides a perspective to understand the complex molecular network of leukemogenesis.
Materials and Methods
Data sources
In this study, mRNA sequencing data for AML and healthy controls were downloaded from UCSC XENA (https://xenabrowser.net/datapages/), which were sourced from the TCGA and GETx databases and processed using the Toil pipeline to minimize batch effects [35]. The validation datasets were downloaded from the GEO, CCLE (https://www.broadinstitute.org/ccle) [36], and Vizome (http://vizome.org/) [37].
HSF expression, diagnostic, and prognostic analyses
The HSF family includes HSF1, HSF2, HSF4 and HSF5. The expression level, diagnostic efficacy and prognostic significance of HSFs in AML were detected based on the data sourced from TCGA and GTEx. Expression levels were analyzed for normality using the Kolmogorov–Smirnov test. An unpaired t test was used for independent samples, and the Mann–Whitney test was used if the assumptions of normality for the t tests were not fulfilled. The expression levels of HSF1, HSF2, HSF4, and HSF5 in AML cell lines was further examined in CCLE. The ROC curve was then generated to evaluate the diagnostic value of HSF1 in AML based on data from TCGA. An AUC closer to 1 means better classification of the samples, and indicates a test of perfect diagnostic value, whereas an AUC of 0.5 indicated no diagnostic value.
High (top 50%) and low (bottom 50%) HSF1 expression groups were divided according to the median expression of HSF1. OS of the high expression group and the low expression group was assessed using the Kaplan-Meier method by log-rank test based on data sourced from TCGA. Subsequently, AML patients were divided into different subgroups based on clinical variables, and the prognostic value of HSF1 among different subgroups was assessed to further elucidate the prognostic significance of HSF1 in AML patient subgroups.
HSF1 expression level and diagnostic efficacy were validated using GSE9476, and the prognostic significance was validated using GSE12417 and Vizome. Statistical analyses and visualization were performed using GraphPad Prism (version 8.0, GraphPad Software Inc., La Jolla, CA, USA), and a P-value < 0.05 was considered to be statistically significant.
Evaluation of correlation between HSF1 and clinical parameters in AML
The association of HSF1 expression with clinical parameters including patient gender, age, BM blast proportion, PB blast proportion, white blood cell (WBC) counts, cytogenetic risks, NPM1 mutation and FLT3 mutation was estimated using logistic regression models based on TCGA data, which was validated using data sourced from the Vizome database. This analysis was conducted using IBM SPSS Statistics (version 26.0, SPSS Inc., Chicago, IL, USA) by taking HSF1 as an independent variable, and relevant clinical parameters were set as dependent variables. Clinical parameters with a threshold P-value < 0.05 were identified as independent clinical factors associated with HSF1 expression.
CancerSEA analyses
CancerSEA (http://biocc.hrbmu.edu.cn/CancerSEA/) is an integrative database depicts functional states of cancer cells at a single–cell resolution, covering 14 functional phenotypes of 41,900 cancer single cells from 25 cancer types [38]. In order to explore the underlying role of HSF1 in leukemogenesis, CancerSEA was used for functional analysis based on single–cell level. The correlation between HSF1 and functional states in AML, including invasion, metastasis, proliferation, angiogenesis, apoptosis, stemness, cell cycle, differentiation, DNA damage, DNA repair, hypoxia, epithelial–mesenchymal transition, inflammation and quiescence was investigated.
Linkedomics analyses
LinkedOmics (http://www.linkedomics.org/login.php), a database containing multiomics data and clinical data for 32 cancer types and a total of 11,158 patients from TCGA project [39]. Genes related to HSF1 expression in AML were screened in linkedomics using a Pearson's correlation. The top 50 genes positively and negatively associated with HSF1 expression were collected for further analysis. Cytoscape software (version 3.9.1) was used for visualization [40].
GeneMANIA analyses
GeneMANIA (http://www.genemania.org) is a tool for generating hypotheses about gene function, analyzing gene lists and prioritizing genes for functional assays [41]. Positively and negatively correlated genes were uploaded to GeneMANIA to determine protein–protein interactions, gene regulation, coexpression and pathway association.
Functional enrichment analyses
To understand the functional implications of HSF1 in AML. KEGG analyses of positively and negatively correlated genes of HSF1 were performed based on KOBAS–i (http://kobas.cbi.pku.edu.cn/) [42]. Differentially expressed genes between high (top 50%) and low (bottom 50%) HSF1 expression groups were screen based on R package “DESeq2” (version 1.26.0). Based on the differential analyses of high and low HSF1 expression groups, the differentially expressed genes (DEGs) were sorted according to log fold change and enriched to carry out GSEA based on the R package “clusterProfiler” (version 3.14.3). The Hallmark list “h.all.v7.2.symbols.gmt” from MSigDB collections (version 5) was used as the gene set references. A Benjamini–Hochberg false discovery rate (FDR)-value < 0.05 and P. adjust value <0.05 were considered to be significantly enriched.
Immune infiltration analyses
HSF1 expression with immune cell infiltration levels in AML was further explored. The gene markers of tumor–infiltrating immune cell markers are referenced in prior studies [43]. The infiltration levels of 23 immune cell types including DCs, immature DCs, activated DCs, plasmacytoid DCs, NK cells, NK CD56dim cells, B cells, T cells, CD8 T cells, Th cells, Tcm cells , T effector memory (Tem) cells, T follicular helper (TFH) cells, T gamma delta (Tgd) cells, Th1 cells, Th2 cells, Th17 cells, Treg cells, cytotoxic cells, eosinophils, macrophages, mast cells, and neutrophils, were quantified as enrichment scores using ssGSEA based on the R package “GSVA” (version 3.15), and the enrichment score represents the infiltration abundance of immune cells. A Pearson’s correlation was used to investigate the correlation between HSF1 expression and immune cell infiltration. A P-value < 0.05 was considered to be a significant association.
Data availability
All R code used for the pre-processing of the data is available at Open Science Framework (OSF), https://osf.io/nw23m/. The data supporting the conclusions of this article are available upon request to the corresponding author.
Author Contributions
R.X. and C.L. conceived the study. Q.W. and C.L. contributed to the data acquisition and analyses. T.W., X.Y., Z.L., C.G., K.L., S.C. and Z.W. helped to perform the data analyses. C.L. drafted the manuscript. C.L. and R.X. revised the manuscript. R.X. and Y.W. helped in funding acquisition. All of the authors have reviewed and approved the final manuscript.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
This work was supported by the National Natural Science Foundation of China (No. 81974547).
References
- 1. Newell LF, Cook RJ. Advances in acute myeloid leukemia. BMJ. 2021; 375:n2026. https://doi.org/10.1136/bmj.n2026 [PubMed]
- 2. Muench DE, Olsson A, Ferchen K, Pham G, Serafin RA, Chutipongtanate S, Dwivedi P, Song B, Hay S, Chetal K, Trump-Durbin LR, Mookerjee-Basu J, Zhang K, et al. Mouse models of neutropenia reveal progenitor-stage-specific defects. Nature. 2020; 582:109–14. https://doi.org/10.1038/s41586-020-2227-7 [PubMed]
- 3. Dwivedi P, Muench DE, Wagner M, Azam M, Grimes HL, Greis KD. Time resolved quantitative phospho-tyrosine analysis reveals Bruton's Tyrosine kinase mediated signaling downstream of the mutated granulocyte-colony stimulating factor receptors. Leukemia. 2019; 33:75–87. https://doi.org/10.1038/s41375-018-0188-8 [PubMed]
- 4. Lindquist S, Craig EA. The heat-shock proteins. Annu Rev Genet. 1988; 22:631–77. https://doi.org/10.1146/annurev.ge.22.120188.003215 [PubMed]
- 5. Feder ME, Hofmann GE. Heat-shock proteins, molecular chaperones, and the stress response: evolutionary and ecological physiology. Annu Rev Physiol. 1999; 61:243–82. https://doi.org/10.1146/annurev.physiol.61.1.243 [PubMed]
- 6. Yun CW, Kim HJ, Lim JH, Lee SH. Heat Shock Proteins: Agents of Cancer Development and Therapeutic Targets in Anti-Cancer Therapy. Cells. 2019; 9:60. https://doi.org/10.3390/cells9010060 [PubMed]
- 7. Calderwood SK, Gong J. Heat Shock Proteins Promote Cancer: It's a Protection Racket. Trends Biochem Sci. 2016; 41:311–23. https://doi.org/10.1016/j.tibs.2016.01.003 [PubMed]
- 8. Reikvam H, Ersvaer E, Bruserud O. Heat shock protein 90 - a potential target in the treatment of human acute myelogenous leukemia. Curr Cancer Drug Targets. 2009; 9:761–76. https://doi.org/10.2174/156800909789271486 [PubMed]
- 9. Sanghera SS, Skitzki JJ. Targeting the heat shock response in cancer: tipping the balance in transformed cells. Surg Oncol Clin N Am. 2013; 22:665–84. https://doi.org/10.1016/j.soc.2013.06.004 [PubMed]
- 10. Mendillo ML, Santagata S, Koeva M, Bell GW, Hu R, Tamimi RM, Fraenkel E, Ince TA, Whitesell L, Lindquist S. HSF1 drives a transcriptional program distinct from heat shock to support highly malignant human cancers. Cell. 2012; 150:549–62. https://doi.org/10.1016/j.cell.2012.06.031 [PubMed]
- 11. Chen F, Fan Y, Cao P, Liu B, Hou J, Zhang B, Tan K. Pan-Cancer Analysis of the Prognostic and Immunological Role of HSF1: A Potential Target for Survival and Immunotherapy. Oxid Med Cell Longev. 2021; 2021:5551036. https://doi.org/10.1155/2021/5551036 [PubMed]
- 12. Brodská B, Otevřelová P, Šálek C, Fuchs O, Gašová Z, Kuželová K. High PD-L1 Expression Predicts for Worse Outcome of Leukemia Patients with Concomitant NPM1 and FLT3 Mutations. Int J Mol Sci. 2019; 20:2823. https://doi.org/10.3390/ijms20112823 [PubMed]
- 13. Puustinen MC, Sistonen L. Molecular Mechanisms of Heat Shock Factors in Cancer. Cells. 2020; 9:1202. https://doi.org/10.3390/cells9051202 [PubMed]
- 14. Dai C. The heat-shock, or HSF1-mediated proteotoxic stress, response in cancer: from proteomic stability to oncogenesis. Philos Trans R Soc Lond B Biol Sci. 2018; 373:20160525. https://doi.org/10.1098/rstb.2016.0525 [PubMed]
- 15. Wang G, Cao P, Fan Y, Tan K. Emerging roles of HSF1 in cancer: Cellular and molecular episodes. Biochim Biophys Acta Rev Cancer. 2020; 1874:188390. https://doi.org/10.1016/j.bbcan.2020.188390 [PubMed]
- 16. Li G, Song Y, Zhang Y, Wang H, Xie J. miR-34b Targets HSF1 to Suppress Cell Survival in Acute Myeloid Leukemia. Oncol Res. 2016; 24:109–16. https://doi.org/10.3727/096504016X14611963142254 [PubMed]
- 17. Frezzato F, Raggi F, Martini V, Severin F, Trimarco V, Visentin A, Scomazzon E, Accordi B, Bresolin S, Piazza F, Facco M, Basso G, Semenzato G, Trentin L. HSP70/HSF1 axis, regulated via a PI3K/AKT pathway, is a druggable target in chronic lymphocytic leukemia. Int J Cancer. 2019; 145:3089–100. https://doi.org/10.1002/ijc.32383 [PubMed]
- 18. Prince TL, Lang BJ, Guerrero-Gimenez ME, Fernandez-Muñoz JM, Ackerman A, Calderwood SK. HSF1: Primary Factor in Molecular Chaperone Expression and a Major Contributor to Cancer Morbidity. Cells. 2020; 9:1046. https://doi.org/10.3390/cells9041046 [PubMed]
- 19. Fok JHL, Hedayat S, Zhang L, Aronson LI, Mirabella F, Pawlyn C, Bright MD, Wardell CP, Keats JJ, De Billy E, Rye CS, Chessum NEA, Jones K, et al. HSF1 Is Essential for Myeloma Cell Survival and A Promising Therapeutic Target. Clin Cancer Res. 2018; 24:2395–407. https://doi.org/10.1158/1078-0432.CCR-17-1594 [PubMed]
- 20. Ganguly S, Home T, Yacoub A, Kambhampati S, Shi H, Dandawate P, Padhye S, Saluja AK, McGuirk J, Rao R. Targeting HSF1 disrupts HSP90 chaperone function in chronic lymphocytic leukemia. Oncotarget. 2015; 6:31767–79. https://doi.org/10.18632/oncotarget.5167 [PubMed]
- 21. Santón A, García-Cosío M, Cristóbal E, Pascual A, Muriel A, García-Laraña J. Expression of heat shock proteins in classical Hodgkin lymphoma: correlation with apoptotic pathways and prognostic significance. Histopathology. 2011; 58:1072–80. https://doi.org/10.1111/j.1365-2559.2011.03803.x [PubMed]
- 22. Hoff FW, van Dijk AD, Qiu Y, Ruvolo PP, Gerbing RB, Leonti AR, Jenkins GN, Gamis AS, Aplenc R, Kolb EA, Alonzo TA, Meshinchi S, de Bont ESJM, et al. Heat shock factor 1 (HSF1-pSer326) predicts response to bortezomib-containing chemotherapy in pediatric AML: a COG report. Blood. 2021; 137:1050–60. https://doi.org/10.1182/blood.2020005208 [PubMed]
- 23. Hsu AL, Murphy CT, Kenyon C. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science. 2003; 300:1142–5. https://doi.org/10.1126/science.1083701 [PubMed]
- 24. Morley JF, Morimoto RI. Regulation of longevity in Caenorhabditis elegans by heat shock factor and molecular chaperones. Mol Biol Cell. 2004; 15:657–64. https://doi.org/10.1091/mbc.e03-07-0532 [PubMed]
- 25. Barna J, Csermely P, Vellai T. Roles of heat shock factor 1 beyond the heat shock response. Cell Mol Life Sci. 2018; 75:2897–916. https://doi.org/10.1007/s00018-018-2836-6 [PubMed]
- 26. Vydra N, Toma A, Widlak W. Pleiotropic role of HSF1 in neoplastic transformation. Curr Cancer Drug Targets. 2014; 14:144–55. https://doi.org/10.2174/1568009614666140122155942 [PubMed]
- 27. Pan X, Lin J, Zeng X, Li W, Wu W, Lu WZ, Liu J, Liu S. Heat shock factor 1 suppresses the HIV-induced inflammatory response by inhibiting nuclear factor-κB. Cell Immunol. 2018; 327:26–35. https://doi.org/10.1016/j.cellimm.2018.01.015 [PubMed]
- 28. Lacourt TE, Kavelaars A, Galloway-Peña JR, Sahasrabhojane PV, Shah ND, Futreal A, Kontoyiannis DP, Shelburne SA, Heijnen CJ. Associations of inflammation with symptom burden in patients with acute myeloid leukemia. Psychoneuroendocrinology. 2018; 89:203–8. https://doi.org/10.1016/j.psyneuen.2018.01.018 [PubMed]
- 29. Récher C. Clinical Implications of Inflammation in Acute Myeloid Leukemia. Front Oncol. 2021; 11:623952. https://doi.org/10.3389/fonc.2021.623952 [PubMed]
- 30. Hemmati S, Haque T, Gritsman K. Inflammatory Signaling Pathways in Preleukemic and Leukemic Stem Cells. Front Oncol. 2017; 7:265. https://doi.org/10.3389/fonc.2017.00265 [PubMed]
- 31. Li Z, Philip M, Ferrell PB. Alterations of T-cell-mediated immunity in acute myeloid leukemia. Oncogene. 2020; 39:3611–9. https://doi.org/10.1038/s41388-020-1239-y [PubMed]
- 32. Lion E, Willemen Y, Berneman ZN, Van Tendeloo VF, Smits EL. Natural killer cell immune escape in acute myeloid leukemia. Leukemia. 2012; 26:2019–26. https://doi.org/10.1038/leu.2012.87 [PubMed]
- 33. Gandhapudi SK, Murapa P, Threlkeld ZD, Ward M, Sarge KD, Snow C, Woodward JG. Heat shock transcription factor 1 is activated as a consequence of lymphocyte activation and regulates a major proteostasis network in T cells critical for cell division during stress. J Immunol. 2013; 191:4068–79. https://doi.org/10.4049/jimmunol.1202831 [PubMed]
- 34. Harada H, Murakami T, Tea SS, Takeuchi A, Koga T, Okada S, Suico MA, Shuto T, Kai H. Heat shock suppresses human NK cell cytotoxicity via regulation of perforin. Int J Hyperthermia. 2007; 23:657–65. https://doi.org/10.1080/02656730701822087 [PubMed]
- 35. Vivian J, Rao AA, Nothaft FA, Ketchum C, Armstrong J, Novak A, Pfeil J, Narkizian J, Deran AD, Musselman-Brown A, Schmidt H, Amstutz P, Craft B, et al. Toil enables reproducible, open source, big biomedical data analyses. Nat Biotechnol. 2017; 35:314–6. https://doi.org/10.1038/nbt.3772 [PubMed]
- 36. Nusinow DP, Szpyt J, Ghandi M, Rose CM, McDonald ER
3rd , Kalocsay M, Jané-Valbuena J, Gelfand E, Schweppe DK, Jedrychowski M, Golji J, Porter DA, Rejtar T, et al. Quantitative Proteomics of the Cancer Cell Line Encyclopedia. Cell. 2020; 180:387–402.e16. https://doi.org/10.1016/j.cell.2019.12.023 [PubMed] - 37. Tyner JW, Tognon CE, Bottomly D, Wilmot B, Kurtz SE, Savage SL, Long N, Schultz AR, Traer E, Abel M, Agarwal A, Blucher A, Borate U, et al. Functional genomic landscape of acute myeloid leukaemia. Nature. 2018; 562:526–31. https://doi.org/10.1038/s41586-018-0623-z [PubMed]
- 38. Yuan H, Yan M, Zhang G, Liu W, Deng C, Liao G, Xu L, Luo T, Yan H, Long Z, Shi A, Zhao T, Xiao Y, Li X. CancerSEA: a cancer single-cell state atlas. Nucleic Acids Res. 2019; 47:D900–8. https://doi.org/10.1093/nar/gky939 [PubMed]
- 39. Vasaikar SV, Straub P, Wang J, Zhang B. LinkedOmics: analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2018; 46:D956–63. https://doi.org/10.1093/nar/gkx1090 [PubMed]
- 40. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003; 13:2498–504. https://doi.org/10.1101/gr.1239303 [PubMed]
- 41. Warde-Farley D, Donaldson SL, Comes O, Zuberi K, Badrawi R, Chao P, Franz M, Grouios C, Kazi F, Lopes CT, Maitland A, Mostafavi S, Montojo J, et al. The GeneMANIA prediction server: biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010; 38:W214–20. https://doi.org/10.1093/nar/gkq537 [PubMed]
- 42. Bu D, Luo H, Huo P, Wang Z, Zhang S, He Z, Wu Y, Zhao L, Liu J, Guo J, Fang S, Cao W, Yi L, et al. KOBAS-i: intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021; 49:W317–25. https://doi.org/10.1093/nar/gkab447 [PubMed]
- 43. Bindea G, Mlecnik B, Tosolini M, Kirilovsky A, Waldner M, Obenauf AC, Angell H, Fredriksen T, Lafontaine L, Berger A, Bruneval P, Fridman WH, Becker C, et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity. 2013; 39:782–95. https://doi.org/10.1016/j.immuni.2013.10.003 [PubMed]