



Introduction
Receptor-interacting protein kinases (RIPKs) are known as a family of serine/threonine kinases, including RIPK1, RIPK2, RIPK3, RIPK4 and RIPK5/DSTYK [1]. Their functions, regulation, and pathophysiological roles have largely remained a labyrinth. The proteins of RIPKs family have garnered significant interest because of their role in regulating various forms of cell death, inflammation, and innate immunity [2, 3]. Owing to the exploration of cell death mechanisms in recent years, in addition to classical apoptosis and necroptosis via activation of NF-κB signaling or ERK/JNK signaling [4], new forms of cell death, such as ferroptosis, have been shown to be related to the RIPK family [5]. Furthermore, an increasing number of research teams have focused on the effect of RIPK genes on tumor progression and tumor immunity via regulation of cell death and correlated pathways [3, 6].
Lung carcinoma is reported as an aggressive malignancy in humans, with a high rate of mortality worldwide [7]. Approximately 40% of all non-small cell lung carcinomas (NSCLCs) were finally diagnosed as lung adenocarcinoma (LUAD) [8]. Surgery remains the main treatment option for LUAD, accompanied by various modalities of chemotherapy, irradiation therapy, targeted therapy, and even immunotherapy in recent decades [9, 10]. Despite extensive research regarding the underlying mechanism of development and treatment resistance among patients with LUAD, the 5-year survival rate remains unsatisfactory, and most patients still experience relapse, metastasis, and death [11].
In this study, we examined the roles of the RIPK family in LUAD based on specific online databases (Table 1) that estimate the biological significance and potential functions of RIPK proteins in LUAD.
Table 1. The main bioinformatics tools used to analyze the functions of RIPK2 in LUAD.
| Database | Samples | URL |
| GEPIA | Tissues | |
| Wanderer | Tissues | |
| UCLCAN | Tissues | |
| PrognoScan | Tissues | |
| cBioPortal | Tissues | |
| STRING | − | |
| Cytoscape | − | |
| WebGestalt | ||
| TIMER | Tissues | |
| DiseaseMeth | Tissues |
Results
The expression of the RIPK family in LUAD patients
To better understand the RIPK family functions in LUAD patients, we firstly evaluated the mRNA levels of RIPK1, RIPK2, RIPK3, RIPK4 and RIPK5 from databases GEPIA, Wanderer and UCLCAN. When compared to normal tissues, RIPK2 presented coherently upregulated in LUAD tissues among three databases, while RIPK5 (DSTYK) was decreased. Lower expression of RIPK1 were spotted in LUAD tissue from Wanderer and UCLCAN data. GEPIA database illustrated a downregulated RIPK3 in tumor tissue, but no significant change in Wanderer and UCLCAN database. The level of RIPK4 was found increased in LUAD samples base on GEPIA and UCLCAN data, but this trend was not found in Wanderer data (Figure 1A–1C).
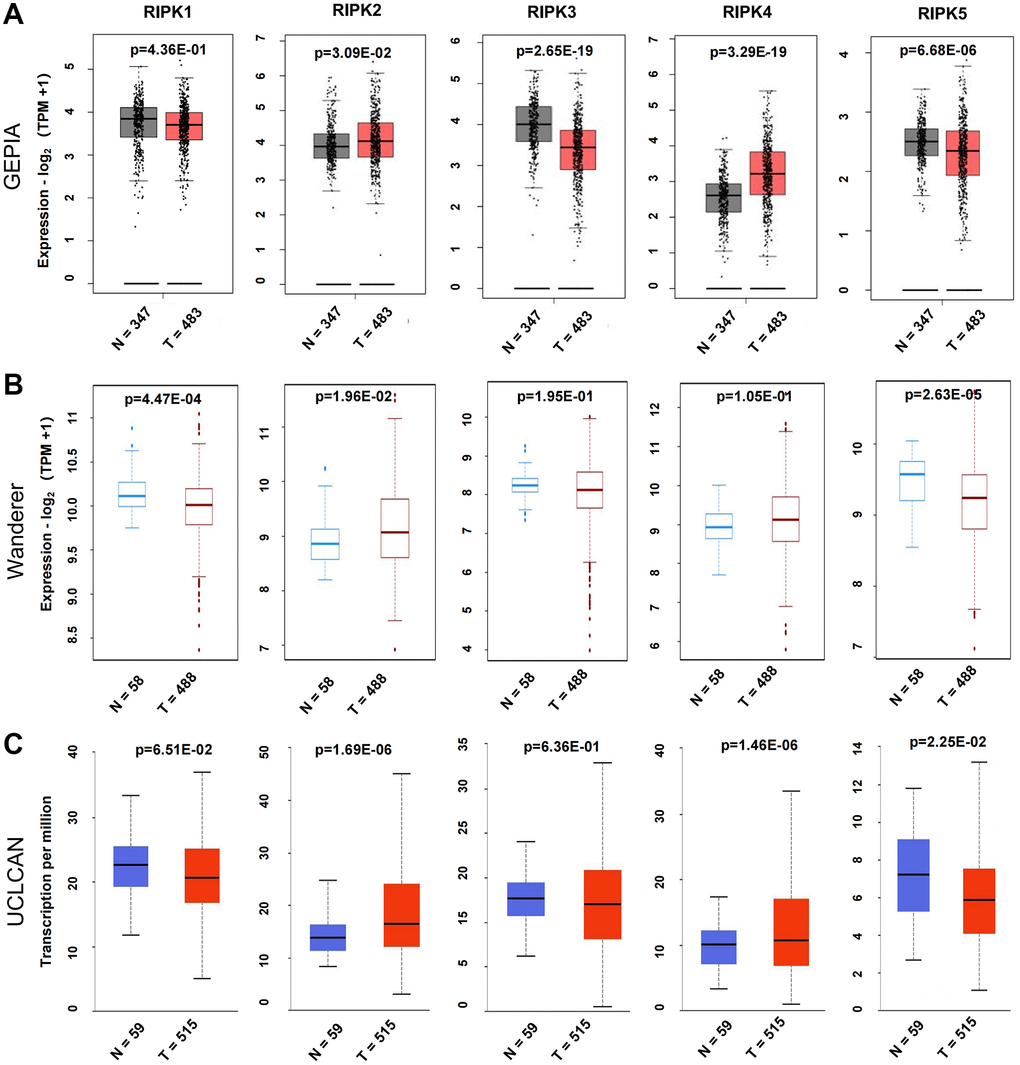
Figure 1. Differential mRNA expression analysis of the RIPK family in LUAD and normal tissues. (A) The expression profiles were collected from the GEPIA databases. (B) The expression profiles were obtained from the Wanderer databases. (C) The expression profiles were analysis via the UALCAN databases.
When evaluating the mRNA levels of the RIPK family in LUAD tissues, RIPK1 and RIPK2 turn out to be the top two expression gene, while RIPK5 expressed lowest (Figure 2A). Further, based on different pathological stages, the mRNAs levels of each RIPK members also analyzed respectively. Only RIPK2 expression positively correlated with pathological stage (p = 0.00055) (Figure 2C). No significant correlation observed in other RIPK family and pathological stages (p > 0.05, Figure 2B, 2D–2F). The data above implied that RIPK genes could participate in LUAD progression.
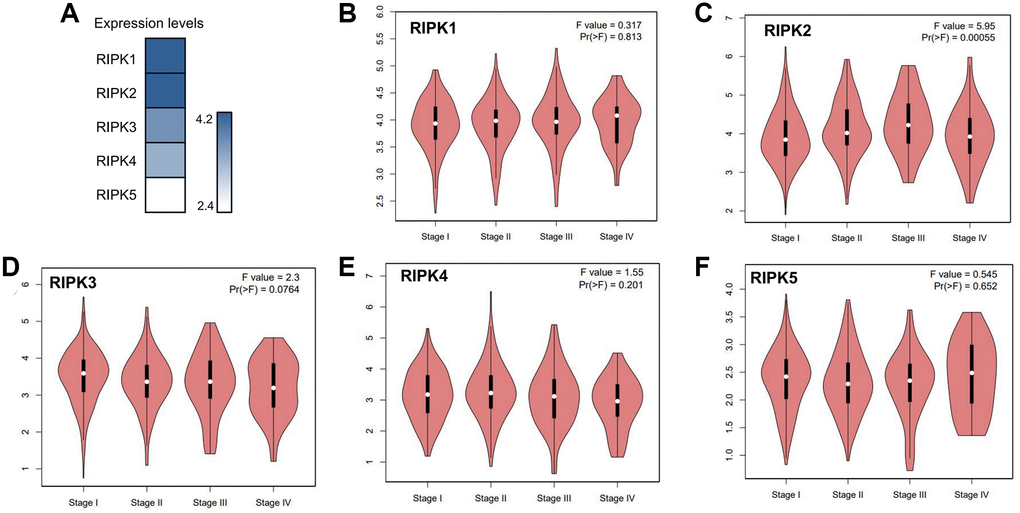
Figure 2. The relative expression levels of the RIPK family in LUAD patients and their correlation to clinic stages. (A) GEPIA databases were used to evaluate the relative expression levels of the RIPK family in LUAD patients. (B–F) the correlation between expression of RIPK1-5 and tumor clinic stage.
The prognostic value of the RIPK family in LUAD patients
Furthermore, we analyzed the impact of RIPK family on the survival of LUAD patients base on the PrognoScan databases. The overall survival (OS) was displayed in Figure 3A in which worse prognosis were found in cases with higher level of RIPK2 (p < 0.00001) and RIPK3 (p = 0.00324), but higher level of RIPK5 suggested a longer survival time (p = 0.00168). In addition, relapse free survival (RFS) data was also estimated according to the RIPK family level. As showed in Figure 3B, high level of RIPK2 indicated a worse RFS (p < 0.00001), as well as the low level of RIPK1 (p = 0.00328), RIPK4 (p = 0.0203) and RIPK5 (p = 0.0068).
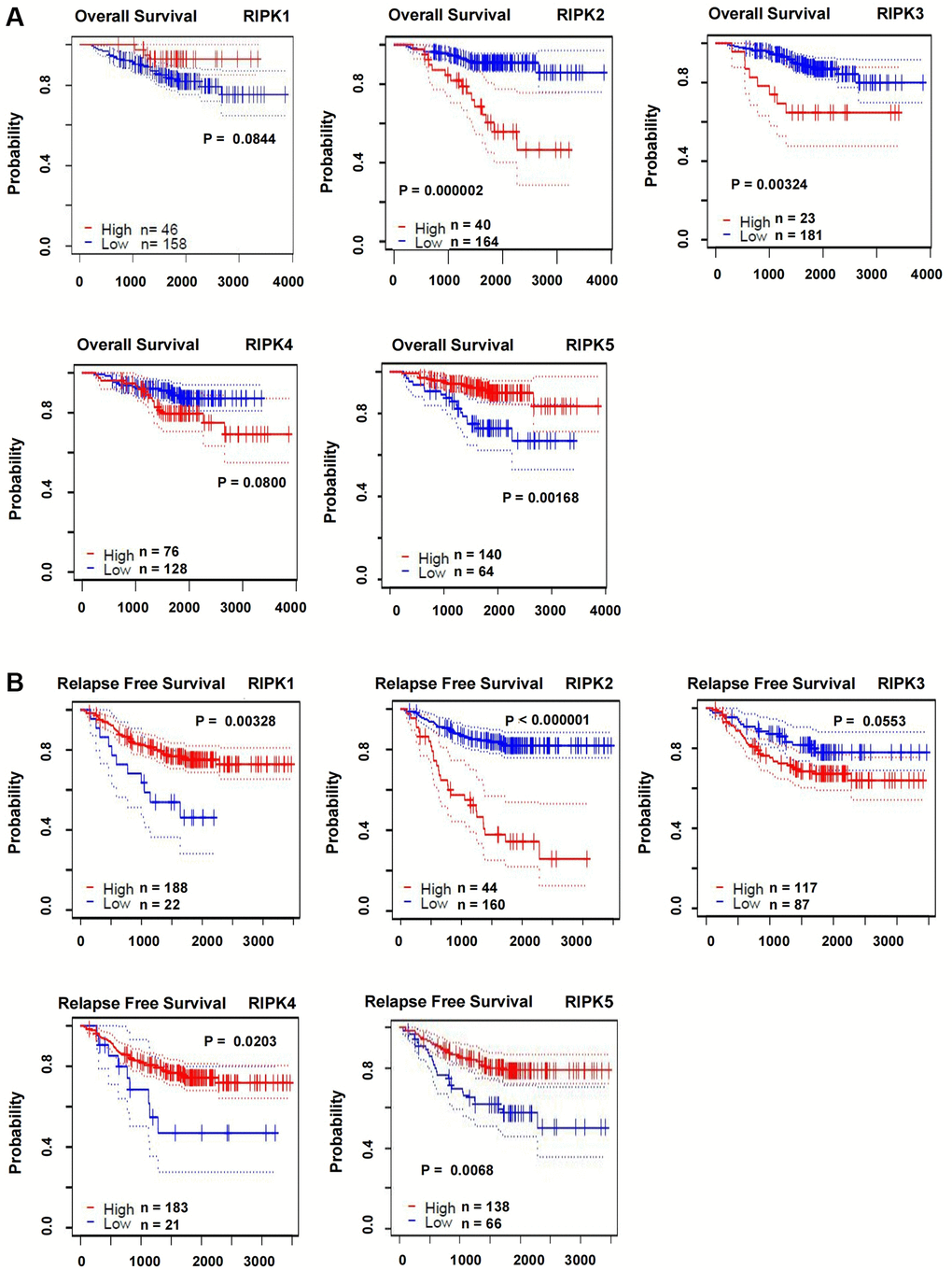
Figure 3. The correlations of RIPK family expression with OS and RSF in LUAD patients. (A) Kaplan-Meier plotter was used to assess the correlation of RIPK family members with the patients’ OS. (B) The correlations of RIPK family expression with RFS in LUAD patients.
Genetic alteration and methylation level of the RIPK family in LUAD patients
Genetic alteration is another pivotal factor resulting in tumor generation and advanced progression. We herein analyzed the alteration profiles of RIPK members by using the cBioPortal database. RIPK2 and RIPK5 present highest alternation ratio reaching at 16% among the LUAD cases, in which mutation was rich in ‘mRNA high’ annotation. The genetic mutations of RIPK1, RIPK3 and RIPK4 were 12%, 5% and 4% respectively (Figure 4A).
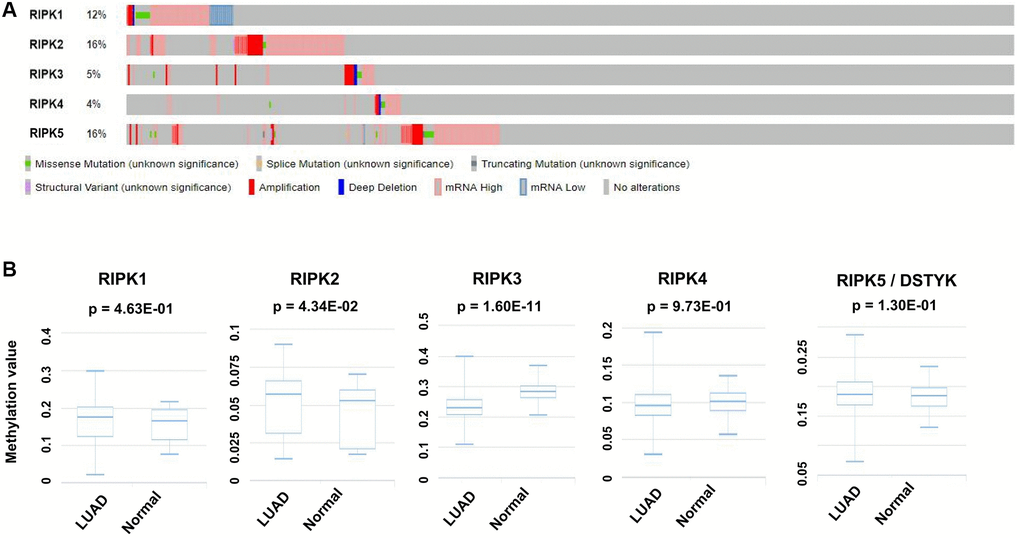
Figure 4. Genetic alteration and methylation level of the RIPK family in LUAD patients. (A) Genetic alteration of the RIPK family in LUAD patients analyzed with cBioPortal. (B) The methylation values of RIPK family members were evaluated using the DiseaseMeth database.
Methylation level was supposed as another regulation mechanism involving in LUAD. From DiseaseMeth database related to LUAD cases, we could confirm a higher methylation level of RIPK2 (p = 0.0434), but lower methylation level of RIPK3 (p = 1.60e-11) when comparing to health people (Figure 4B).
Interaction and functional enrichment analysis of the RIPK family in LUAD patients
We analyzed cBioPortal database and finally extracted 42 most frequently altered genes which mRNA levels were significantly correlated to the RIPK members in LUAD patients. Several hub genes, including CCND1, MYC, MTOR, MAPK1, CDKN1A and CCNB1, were turned out to participate actively in the bio-malignant behaviors of RIPK family modulation in LUAD cases (Figure 5).
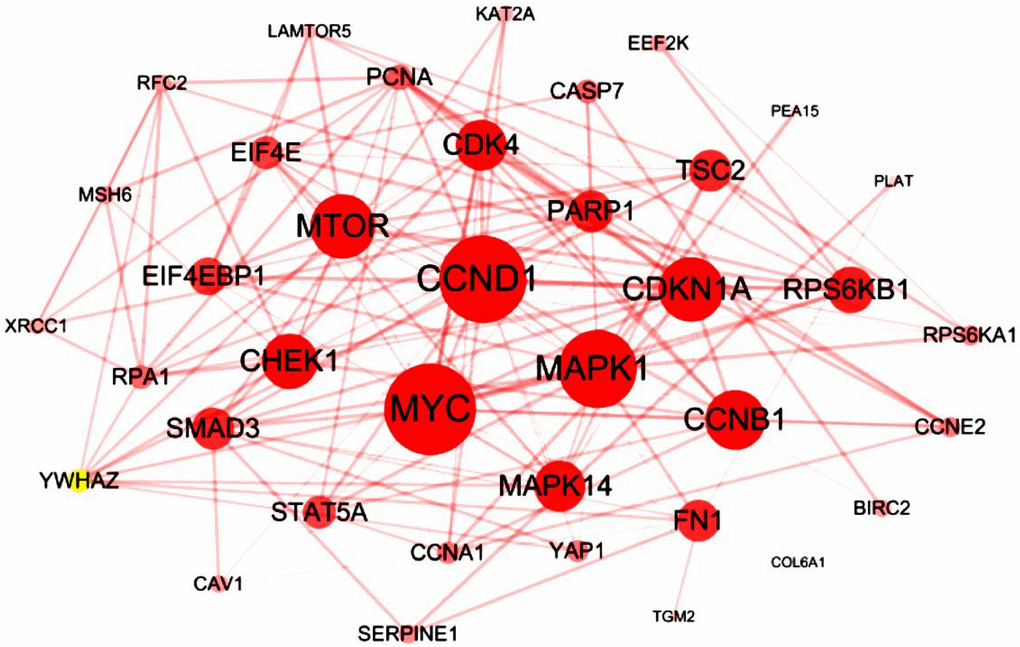
Figure 5. Interaction analysis of the RIPK family in LUAD patients. The 42 most frequently altered genes identified from cBioPortal that are linked to the RIPKs family in LUAD patients.
Next, WebGestalt database was applied to estimate the biological functions of RIPK family based on the strong relevant genes above. As shown in the GO pathways, the top enriched biological processes were metabolic process, response to stimulus, biological regulation and cell communication (Figure 6A). Cellular component categories were highly enriched in cytosol, nucleus protein-containing complex, membrane-enclosed lumen, membrane (Figure 6B). When involving the molecular function categories, the top five related function were protein binding, transferase activity and ion binding (Figure 6C).
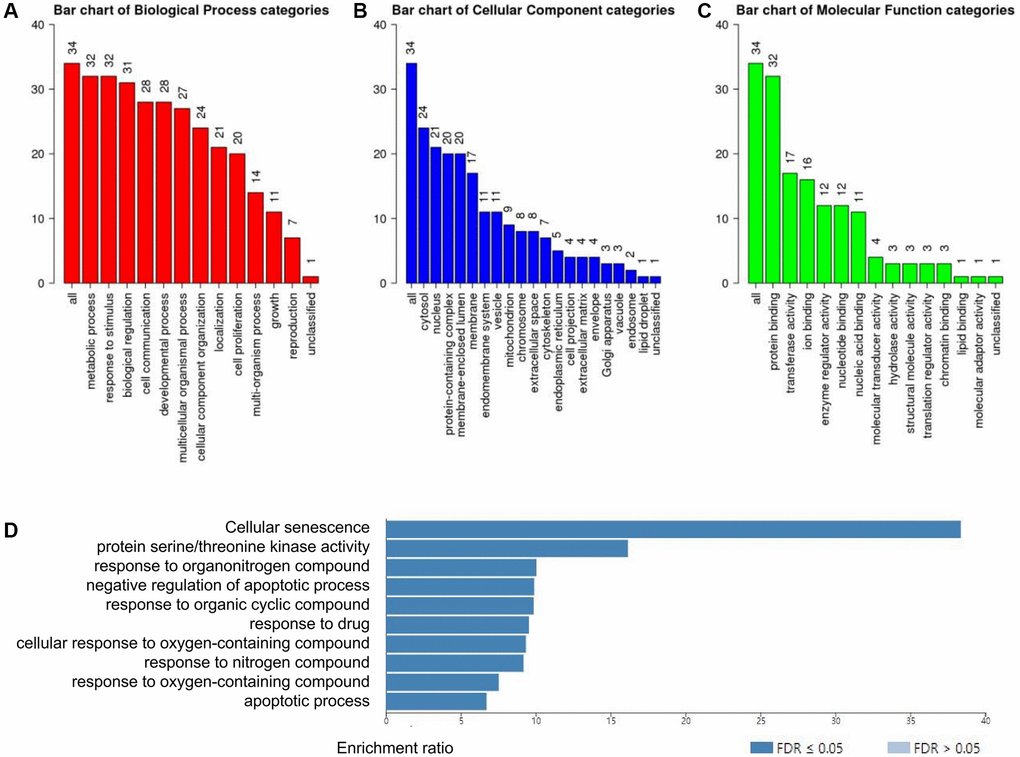
Figure 6. Functional enrichment analysis of the RIPK family in LUAD patients with WebGestalt database. (A–C) Bar plot of GO enrichment in cellular components, biological processes, and molecular functions. (D) The bar plot of KEGG enrichment.
In addition, 10 KEGG pathways was strongly associated to the biological functions of RIPK family in the generation and progression of LUAD, including cellular senescence, protein serine/threonine kinase activity, response to organonitrogen compound, negative regulation of apoptotic process, response to organic cyclic compound, response to drug, cellular response to oxygen-containing compound, response to nitrogen compound, response to oxygen-containing compound and apoptotic process (Figure 6D).
Immune cell infiltration of the RIPK family
Recent study illustrated that the roles of immune cells should be seriously considered in the LAUD progression, as well as their impacts on treatment efficiencies and prognosis. Hence, we identified the connection between RIPK family members and six distinct immune subsets which infiltrated tumor via TIMER database analysis. Intriguingly, RIPK1 was positively linked to the infiltrated B cell (Cor = 0.137, p = 2.50e-03), CD8+ T cell (Cor = 0.129, p = 4.43e-03), CD4+ T cell (Cor = 0.295, p=3.44e-11), macrophage (Cor = 0.166, p=2.45e-04), neutrophil (Cor = 0.281, p = 3.45e-10), dendritic cell (Cor = 0.26, p = 5.84e-09) (Figure 7A). RIPK2 level was positively related to the infiltrated CD8+ T cell (Cor = 0.263, p = 3.58e-09), neutrophil (Cor = 0.322, p = 3.90e-13), dendritic cell (Cor = 0.187, p = 3.27e-05) (Figure 7B). Further, positive correlation was spotted between RIPK3 and B cell (Cor = 0.262, p = 5.14e-09), CD4+ T cell (Cor = 0.438, p = 4.09e-24), macrophage (Cor = 0.247, p = 3.75e-08), neutrophil (Cor = 0.251, p = 2.37e-08), dendritic cell (Cor = 0.339, p = 1.50e-14) (Figure 7C). For RIPK4, we found its mRNA level positively related to infiltrated B cell (Cor = 0.153, p = 7.40e-04), CD4+ T cell (Cor = 0.265, p = 3.189e-09), dendritic cell (Cor = 0.113, p = 1.21e-02) (Figure 7D). Finally, expression of RIPK5(DSTYK) showed its positive relationship to B cell (Cor = 0.13, p = 4.14e-03), CD4+ T cell (Cor = 0.227, p = 4.74e-07), macrophage (Cor = 0.121, p = 7.84e-03), neutrophil (Cor = 0.095, p = 3.78e-02), dendritic cell (Cor = 0.1126, p = 1.37e-02) (Figure 7E).
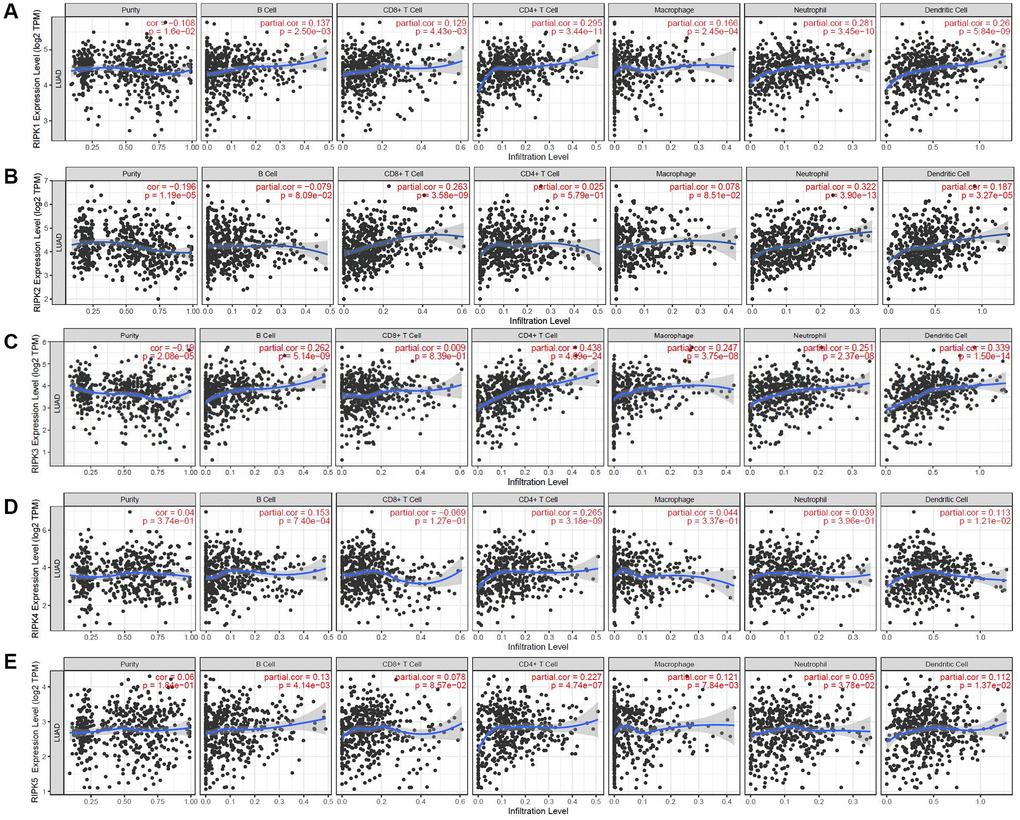
Figure 7. The relationship between immune cell infiltration and the expression of the RIPK family. The TIMER database was used to analyze the effect of (A) RIPK 1, (B) RIPK 2, (C) RIPK 3, (D) RIPK 4, (E) RIPK 5 on the abundance of immune cells in LUAD patients.
The relationship between RIPK family and the infiltration of immune cells were analyzed (Table 2). 12 confounding factors (macrophages, neutrophils, dendritic cells, B cells, CD8+ T cells, CD4+ T cells and five RIPK family members) were put into the Cox proportional hazard model together for further analyzed. From Table 2, we found that B cells (HR = 0.012, p = 0.001), CD4+ T cells (HR = 29.495, p = 0.0170) and RIPK2 (HR = 1.458, p = 0.001) could be considered as independent prognosis factors related to LAUD.
Table 2. The cox proportional hazard model of RIPK family and six types of immune cells in LUAD patients from TIMER database.
| Coef | HR | 95% CI | P value | Sig. | |
| B cell | −4.432 | 0.012 | 0.001−0.161 | 0.001 | ** |
| CD8 T cell | −0.165 | 0.848 | 0.131−5.500 | 0.863 | |
| CD4 T cell | 3.384 | 29.498 | 1.842−472.264 | 0.017 | * |
| Macrophage | 0.664 | 1.943 | 0.143−26.363 | 0.618 | |
| Neutrophil | −2.275 | 0.103 | 0.002−5.513 | 0.263 | |
| Dendritic cell | −0.208 | 0.812 | 0.214−3.077 | 0.759 | |
| RIPK1 | −0.066 | 0.936 | 0.639−1.372 | 0.734 | |
| RIPK2 | 0.377 | 1.458 | 1.168−1.821 | 0.001 | ** |
| RIPK3 | −0.138 | 0.871 | 0.702−1.080 | 0.209 | |
| RIPK4 | −0.066 | 0.937 | 0.772−1.136 | 0.506 | |
| DSTYK | −0.066 | 0.936 | 0.695−1.257 | 0.662 |
Discussion
RIPKs share homologous kinase domains but differ in other functional units, such as identical P-loops, an ionic pair of catalytic Lys and Glu residues in the center of C helix, and an HXD motif in the catalytic loop [1, 4, 12]. Thus, the core kinase domain and these various elements endow these proteins with multiple functional connections and distinct clinical properties in cancer, as supported by the data presented in our study related to LUAD.
RIPK1 were proved as a core protein in a wide range of aberrant physiological processes like neurodegeneration, autoimmune, inflammatory activation and malignancies [13, 14]. Aberrantly expressed RIPK1 has been demonstrated to regulate the cell death pathways, including apoptosis [15] and necrotic cell death [16] as well as ferroptosis [5]. The underlying pathway always leads to activation of MAPKs and NF-κB [17], along with phosphorylation [18] and ubiquitination [19]. RIPK1 participates in various cancers and induces malignancy. Activation of the RIPK1/NF-κB pathway can induce NSCLC cell proliferation and migration [20]. Single-nucleotide polymorphism rs17548629 in RIPK1 may be associated with the formation of lung cancer [21]. However, whether this molecule can be considered a target for tumor therapy is still a matter of dispute. Patel et al. did not find any effect of a RIPK1 inhibitor on tumor growth or metastases, although it did help in inflammatory disease control [22]. Our data revealed that RIPK1 expression was decreased in LUAD and that it increased RFS.
RIPK2 is also known as RIP-like-interacting caspase-like apoptosis-regulatory protein kinase or CARD-containing IL-1b converting enzyme-associated kinase [1, 23]. RIPK2, as a critical protein for the signaling from NOD-like receptors, always triggered MAPK, ERK2, and JNK activation and further affect apoptosis [24]. Thus, target treatment on RIPK2 showed an ideal control of inflammatory diseases, such as cystic fibrosis, asthma, inflammatory bowel disease and pancreatitis [3, 25, 26]. Rare study has documented RIPK2 functions and roles in LUAD. According to our analysis, higher RIPK2 expression was observed in LUAD tissues, suggesting a worse prognosis, and this was positively correlated with the pathological stage. Notably, higher methylation levels of RIPK2 were also found in tumor tissues. Generally, DNA methylation indicates resilience of gene transcription [27]. Here, both RIPK2 expression and methylation levels increased in tumor tissues, which may reflect DNA methylation, not only in the intergenic region or CpG islands to repress gene transcription but also in the non-first exon of the gene body, which is associated with a higher level of gene expression in dividing cells [27]. Surprisingly, the Cox proportional hazard model identified RIPK2 as an independent prognostic factor, as well as B cells and CD4+ T cells. The TIMER database showed that RIPK2 is positively linked to infiltration by B cells, CD8+ T cells, neutrophils, and dendritic cells. This suggests that RIPK2 is a potential target and immunoregulatory protein in LUAD patients.
Similar to RIPK1, RIPK3 play a key function for caspase-independent necrotic cell death via anti-TNF treatment [17, 28]. Phosphorylation of RIPK1 and RIPK3 can stabilize their interaction within the pronecrotic complex inducing kinase activity, triggering downstream ROS (reactive oxygen species) reaction, and resulting in terminal necrotic cell death [29]. However, there are still some divergent opinions regarding its function. Newton et al. reported that the level of NF-kB activation could be induced by TNF in RIPK3-deficient mice [30]. RIPK3 is thought to be a carcinogenic factor in various cancers, such as breast cancer, intestinal and colon cancer, and lung cancer [31]. According to our data, RIPK3 expression negatively correlated with prognosis and positively correlated with various tumor-infiltrating lymphocytes. We also found a lower methylation level of RIPK3 in LUAD patients, which suggests that epigenetic regulation participates in RIPK3-induced biological properties of LUAD.
In addition to sharing a kinase domain, RIPK4 has a C-terminal ankyrin repeats and N-terminal kinase domain part [4]. RIPK4 strongly links to the NF-κB and JNK signaling activation [32] and also regulates the PKC signaling [4]. A recent study revealed that RIPK4 can influence the biological properties of malignancies, such as pancreatic [33], bladder [34], and cervical cancers [35]. It can inhibit STAT3 signaling to sustain LUAD differentiation [36]. Our data revealed that RIPK4 levels are increased in LUAD samples, based on GEPIA and UCLCAN. In addition, RIPK4 was found to positively correlated with infiltrating CD4+ T cells, which has not been reported by other groups.
RIPK5 (encoded by DSTYK) exhibits high sequence homology with RIPK4 (with an overall level of homology of 35%), suggesting similar functions [1]. Increase of RIPK5 could induce cell apoptosis via DNA fragmentation [37]. However, the biological function of RIPK5 remains largely unknown. Our research showed that RIPK5 expression was low in LUAD patients and was indicative of a better prognosis. Additionally, the level of infiltrating B cells and CD4+ T cells were both strongly related to the expression of RIPK5.
In conclusion, we presented the expression profiles of RIPK family members in patients with LUAD. Those findings may facilitated identification of new diagnostic markers and treatment targets, thereby leading to new treatment options for LUAD patients to prevent tumor relapse and prolong their survival time.
Materials and Methods
GEPIA/Wanderer/UALCAN database
With a web server calculation, Gene Expression Profiling Interactive Analysis (GEPIA) database can illustrate us the gene expression difference and interactive relationship between cancer and normal tissue [38]. The Wanderer database can visualize gene expressions according to TCGA data [39]. UALCAN could offer researchers a group of comprehensive and interactive web data via analyzing cancer bio-information data [40]. We used these databases to estimate the expression level of RIPKs genes and also the roles of the RIPK family in clinic stage, prognosis as previous description [41]. P-values was set at 0.05 in our process.
PrognoScan database
We use PrognoScan database [42] to analyze and illustrate the expression RIPK family members and the survival trend in LUAD patients. Here, overall survival (OS) and relapse-free survival (RFS) curves were included to evaluate the prognosis of patients with LUAD. P-value will offer if less than 0.05.
cBioPortal
cBioPortal database includes more than 100 malignancies genomics data. With a series user-friendly calculation strategy, researchers can obtain the genetic alterations and the coexpression profiles conveniently [43]. In this study, we focus on the data related to the RIPK family in LUAD tissue.
STRING
STRING was a kind of website which was always applied to dig out potential protein-protein interactions (PPIs) [44]. In this study, RIPK family member-associated protein-protein interactions relationship data in detail were download and then a PPIs network were constructed with Cytoscape software [45].
DiseaseMeth
DiseaseMeth is one of the database which can visualize the DNA methylation level with gene expression [46]. In our analysis, the relationship between the expression of the RIPK family and their methylation levels were sought out.
WebGestalt
The aims of WebGestalt is providing a better understanding of functions of gene set [47], like Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) rerichment. Here, we analyzed the enrichment pathways associated with the RIPK family in the patients with LUAD.
TIMER
TIMER is a useful database to evaluate the immune cells infiltration status among various cancers, offering an immune strategies and targeting molecules clues [48]. Infiltrated immune cells features and RIPKs expression were drew out via the TIMER database.
Author Contributions
Conception and design: Guo Li, Zhijie Xu, Jinwu Peng and Chencheng Fu. Writing, review, and/or revision of the manuscript: Guo Li, Jinwu Peng and Chencheng Fu. Administrative, technical, or material support: Yuanliang Yan, Yong Liu, Xin Zhang and Yuanzheng Qiu. All authors approved final version of manuscript.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
This study was supported by the Natural Science Foundation of Hunan Province (Nos. 2020JJ5934). The National Natural Science Foundation of China (Nos. 81602684, 81773243, 81772903, 81874133).
Editorial Note
This corresponding author has a verified history of publications using a personal email address for correspondence
References
- 1. Cuny GD, Degterev A. RIPK protein kinase family: Atypical lives of typical kinases. Semin Cell Dev Biol. 2021; 109:96–105. https://doi.org/10.1016/j.semcdb.2020.06.014 [PubMed]
- 2. Humphries F, Yang S, Wang B, Moynagh PN. RIP kinases: key decision makers in cell death and innate immunity. Cell Death Differ. 2015; 22:225–36. https://doi.org/10.1038/cdd.2014.126 [PubMed]
- 3. Kondylis V, Pasparakis M. RIP Kinases in Liver Cell Death, Inflammation and Cancer. Trends Mol Med. 2019; 25:47–63. https://doi.org/10.1016/j.molmed.2018.10.007 [PubMed]
- 4. Zhang D, Lin J, Han J. Receptor-interacting protein (RIP) kinase family. Cell Mol Immunol. 2010; 7:243–9. https://doi.org/10.1038/cmi.2010.10 [PubMed]
- 5. Guerrero-Hue M, García-Caballero C, Palomino-Antolín A, Rubio-Navarro A, Vázquez-Carballo C, Herencia C, Martín-Sanchez D, Farré-Alins V, Egea J, Cannata P, Praga M, Ortiz A, Egido J, et al. Curcumin reduces renal damage associated with rhabdomyolysis by decreasing ferroptosis-mediated cell death. FASEB J. 2019; 33:8961–75. https://doi.org/10.1096/fj.201900077R [PubMed]
- 6. Yu X, Deng Q, Bode AM, Dong Z, Cao Y. The role of necroptosis, an alternative form of cell death, in cancer therapy. Expert Rev Anticancer Ther. 2013; 13:883–93. https://doi.org/10.1586/14737140.2013.811180 [PubMed]
- 7. Barta JA, Powell CA, Wisnivesky JP. Global Epidemiology of Lung Cancer. Ann Glob Health. 2019; 85:8. https://doi.org/10.5334/aogh.2419 [PubMed]
- 8. Anusewicz D, Orzechowska M, Bednarek AK. Lung squamous cell carcinoma and lung adenocarcinoma differential gene expression regulation through pathways of Notch, Hedgehog, Wnt, and ErbB signalling. Sci Rep. 2020; 10:21128. https://doi.org/10.1038/s41598-020-77284-8 [PubMed]
- 9. Kleczko EK, Kwak JW, Schenk EL, Nemenoff RA. Targeting the Complement Pathway as a Therapeutic Strategy in Lung Cancer. Front Immunol. 2019; 10:954. https://doi.org/10.3389/fimmu.2019.00954 [PubMed]
- 10. Santarpia M, Aguilar A, Chaib I, Cardona AF, Fancelli S, Laguia F, Bracht JWP, Cao P, Molina-Vila MA, Karachaliou N, Rosell R. Non-Small-Cell Lung Cancer Signaling Pathways, Metabolism, and PD-1/PD-L1 Antibodies. Cancers (Basel). 2020; 12:1475. https://doi.org/10.3390/cancers12061475 [PubMed]
- 11. Lai YH, Chen WN, Hsu TC, Lin C, Tsao Y, Wu S. Overall survival prediction of non-small cell lung cancer by integrating microarray and clinical data with deep learning. Sci Rep. 2020; 10:4679. https://doi.org/10.1038/s41598-020-61588-w [PubMed]
- 12. He S, Wang X. RIP kinases as modulators of inflammation and immunity. Nat Immunol. 2018; 19:912–22. https://doi.org/10.1038/s41590-018-0188-x [PubMed]
- 13. Degterev A, Ofengeim D, Yuan J. Targeting RIPK1 for the treatment of human diseases. Proc Natl Acad Sci U S A. 2019; 116:9714–22. https://doi.org/10.1073/pnas.1901179116 [PubMed]
- 14. Negroni A, Colantoni E, Cucchiara S, Stronati L. Necroptosis in Intestinal Inflammation and Cancer: New Concepts and Therapeutic Perspectives. Biomolecules. 2020; 10:1431. https://doi.org/10.3390/biom10101431 [PubMed]
- 15. Chen J, Kostrzewa RM, Xu X. Necroptosis, a Potential Therapeutic Target for Neurological Disorders. In: Kostrzewa R. (eds) Handbook of Neurotoxicity. New York, NY: Springer. 2014; 69–93. https://doi.org/10.1007/978-1-4614-5836-4_166
- 16. Yuan J, Amin P, Ofengeim D. Necroptosis and RIPK1-mediated neuroinflammation in CNS diseases. Nat Rev Neurosci. 2019; 20:19–33. https://doi.org/10.1038/s41583-018-0093-1 [PubMed]
- 17. Liang S, Nian Z, Shi K. Inhibition of RIPK1/RIPK3 ameliorates osteoclastogenesis through regulating NLRP3-dependent NF-κB and MAPKs signaling pathways. Biochem Biophys Res Commun. 2020; 526:1028–35. https://doi.org/10.1016/j.bbrc.2020.03.177 [PubMed]
- 18. Dondelinger Y, Delanghe T, Rojas-Rivera D, Priem D, Delvaeye T, Bruggeman I, Van Herreweghe F, Vandenabeele P, Bertrand MJM. MK2 phosphorylation of RIPK1 regulates TNF-mediated cell death. Nat Cell Biol. 2017; 19:1237–47. https://doi.org/10.1038/ncb3608 [PubMed]
- 19. O'Donnell MA, Ting AT. NFκB and ubiquitination: partners in disarming RIPK1-mediated cell death. Immunol Res. 2012; 54:214–26. https://doi.org/10.1007/s12026-012-8321-7 [PubMed]
- 20. Hao S, Li S, Wang J, Zhao L, Yan Y, Wu T, Zhang J, Wang C. C-Phycocyanin Suppresses the In Vitro Proliferation and Migration of Non-Small-Cell Lung Cancer Cells through Reduction of RIPK1/NF-κB Activity. Mar Drugs. 2019; 17:362. https://doi.org/10.3390/md17060362 [PubMed]
- 21. Yang S, Yu F, Lin M, Sun L, Wei J, Lai C, Cai L, Hu Z, He F. Single-nucleotide polymorphism rs17548629 in RIPK1 gene may be associated with lung cancer in a young and middle-aged Han Chinese population. Cancer Cell Int. 2020; 20:143. https://doi.org/10.1186/s12935-020-01215-w [PubMed]
- 22. Patel S, Webster JD, Varfolomeev E, Kwon YC, Cheng JH, Zhang J, Dugger DL, Wickliffe KE, Maltzman A, Sujatha-Bhaskar S, Bir Kohli P, Ramaswamy S, Deshmukh G, et al. RIP1 inhibition blocks inflammatory diseases but not tumor growth or metastases. Cell Death Differ. 2020; 27:161–75. https://doi.org/10.1038/s41418-019-0347-0 [PubMed]
- 23. Morchang A, Yasamut U, Netsawang J, Noisakran S, Wongwiwat W, Songprakhon P, Srisawat C, Puttikhunt C, Kasinrerk W, Malasit P, Yenchitsomanus PT, Limjindaporn T. Cell death gene expression profile: role of RIPK2 in dengue virus-mediated apoptosis. Virus Res. 2011; 156:25–34. https://doi.org/10.1016/j.virusres.2010.12.012 [PubMed]
- 24. Roy AC, Chang G, Ma N, Wang Y, Roy S, Liu J, Aabdin ZU, Shen X. Sodium butyrate suppresses NOD1-mediated inflammatory molecules expressed in bovine hepatocytes during iE-DAP and LPS treatment. J Cell Physiol. 2019; 234:19602–20. https://doi.org/10.1002/jcp.28560 [PubMed]
- 25. Honjo H, Watanabe T, Kamata K, Minaga K, Kudo M. RIPK2 as a New Therapeutic Target in Inflammatory Bowel Diseases. Front Pharmacol. 2021; 12:650403. https://doi.org/10.3389/fphar.2021.650403 [PubMed]
- 26. Poudel B, Gurung P. Allergic asthma: RIPK2 takes the lead. J Leukoc Biol. 2018; 104:441–3. https://doi.org/10.1002/JLB.3CE0718-293 [PubMed]
- 27. Moore LD, Le T, Fan G. DNA methylation and its basic function. Neuropsychopharmacology. 2013; 38:23–38. https://doi.org/10.1038/npp.2012.112 [PubMed]
- 28. Dunai Z, Bauer PI, Mihalik R. Necroptosis: biochemical, physiological and pathological aspects. Pathol Oncol Res. 2011; 17:791–800. https://doi.org/10.1007/s12253-011-9433-4 [PubMed]
- 29. Jia Y, Wang F, Guo Q, Li M, Wang L, Zhang Z, Jiang S, Jin H, Chen A, Tan S, Zhang F, Shao J, Zheng S. Curcumol induces RIPK1/RIPK3 complex-dependent necroptosis via JNK1/2-ROS signaling in hepatic stellate cells. Redox Biol. 2018; 19:375–87. https://doi.org/10.1016/j.redox.2018.09.007 [PubMed]
- 30. Newton K, Dugger DL, Wickliffe KE, Kapoor N, de Almagro MC, Vucic D, Komuves L, Ferrando RE, French DM, Webster J, Roose-Girma M, Warming S, Dixit VM. Activity of protein kinase RIPK3 determines whether cells die by necroptosis or apoptosis. Science. 2014; 343:1357–60. https://doi.org/10.1126/science.1249361 [PubMed]
- 31. Gurol K, Shah S, Degterev A. RIPK1 and RIPK3 - emerging targets in cancer? Mol Cell Ther. 2018; 6:1–13. https://doi.org/10.26781/2052-8426-2018-03
- 32. Meylan E, Martinon F, Thome M, Gschwendt M, Tschopp J. RIP4 (DIK/PKK), a novel member of the RIP kinase family, activates NF-kappa B and is processed during apoptosis. EMBO Rep. 2002; 3:1201–8. https://doi.org/10.1093/embo-reports/kvf236 [PubMed]
- 33. Qi ZH, Xu HX, Zhang SR, Xu JZ, Li S, Gao HL, Jin W, Wang WQ, Wu CT, Ni QX, Yu XJ, Liu L. RIPK4/PEBP1 axis promotes pancreatic cancer cell migration and invasion by activating RAF1/MEK/ERK signaling. Int J Oncol. 2018; 52:1105–16. https://doi.org/10.3892/ijo.2018.4269 [PubMed]
- 34. Liu JY, Zeng QH, Cao PG, Xie D, Chen X, Yang F, He LY, Dai YB, Li JJ, Liu XM, Zeng HL, Zhu YX, Gong L, et al. RIPK4 promotes bladder urothelial carcinoma cell aggressiveness by upregulating VEGF-A through the NF-κB pathway. Br J Cancer. 2018; 118:1617–27. https://doi.org/10.1038/s41416-018-0116-8 [PubMed]
- 35. Azizmohammadi S, Azizmohammadi S, Safari A, Kaghazian M, Sadrkhanlo M, Behnod V, Seifoleslami M. High-Level Expression of RIPK4 and EZH2 Contributes to Lymph Node Metastasis and Predicts Favorable Prognosis in Patients With Cervical Cancer. Oncol Res. 2017; 25:495–501. https://doi.org/10.3727/096504016X14749735594687 [PubMed]
- 36. Kopparam J, Chiffelle J, Angelino P, Piersigilli A, Zangger N, Delorenzi M, Meylan E. RIP4 inhibits STAT3 signaling to sustain lung adenocarcinoma differentiation. Cell Death Differ. 2017; 24:1761–71. https://doi.org/10.1038/cdd.2017.81 [PubMed]
- 37. Zha J, Zhou Q, Xu LG, Chen D, Li L, Zhai Z, Shu HB. RIP5 is a RIP-homologous inducer of cell death. Biochem Biophys Res Commun. 2004; 319:298–303. https://doi.org/10.1016/j.bbrc.2004.04.194 [PubMed]
- 38. Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017; 45:W98–102. https://doi.org/10.1093/nar/gkx247 [PubMed]
- 39. Díez-Villanueva A, Mallona I, Peinado MA. Wanderer, an interactive viewer to explore DNA methylation and gene expression data in human cancer. Epigenetics Chromatin. 2015; 8:22. https://doi.org/10.1186/s13072-015-0014-8 [PubMed]
- 40. Xu J, Zhao Y, Sun H, Xiao Q, Ye P. Identification of Versican as an Independent Prognostic Factor in Uveal Melanoma. Int J Gen Med. 2021; 14:4639–51. https://doi.org/10.2147/IJGM.S325846 [PubMed]
- 41. Cai Y, Wu G, Peng B, Li J, Zeng S, Yan Y, Xu Z. Expression and molecular profiles of the AlkB family in ovarian serous carcinoma. Aging (Albany NY). 2021; 13:9679–92. https://doi.org/10.18632/aging.202716 [PubMed]
- 42. Zheng H, Zhang G, Zhang L, Wang Q, Li H, Han Y, Xie L, Yan Z, Li Y, An Y, Dong H, Zhu W, Guo X. Comprehensive Review of Web Servers and Bioinformatics Tools for Cancer Prognosis Analysis. Front Oncol. 2020; 10:68. https://doi.org/10.3389/fonc.2020.00068 [PubMed]
- 43. Gao J, Aksoy BA, Dogrusoz U, Dresdner G, Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, Cerami E, Sander C, Schultz N. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 2013; 6:pl1. https://doi.org/10.1126/scisignal.2004088 [PubMed]
- 44. Szklarczyk D, Franceschini A, Kuhn M, Simonovic M, Roth A, Minguez P, Doerks T, Stark M, Muller J, Bork P, Jensen LJ, von Mering C. The STRING database in 2011: functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011; 39:D561–8. https://doi.org/10.1093/nar/gkq973 [PubMed]
- 45. Smoot ME, Ono K, Ruscheinski J, Wang PL, Ideker T. Cytoscape 2.8: new features for data integration and network visualization. Bioinformatics. 2011; 27:431–2. https://doi.org/10.1093/bioinformatics/btq675 [PubMed]
- 46. Xiong Y, Wei Y, Gu Y, Zhang S, Lyu J, Zhang B, Chen C, Zhu J, Wang Y, Liu H, Zhang Y. DiseaseMeth version 2.0: a major expansion and update of the human disease methylation database. Nucleic Acids Res. 2017; 45:D888–95. https://doi.org/10.1093/nar/gkw1123 [PubMed]
- 47. Liao Y, Wang J, Jaehnig EJ, Shi Z, Zhang B. WebGestalt 2019: gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019; 47:W199–205. https://doi.org/10.1093/nar/gkz401 [PubMed]
- 48. Li T, Fu J, Zeng Z, Cohen D, Li J, Chen Q, Li B, Liu XS. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020; 48:W509–14. https://doi.org/10.1093/nar/gkaa407 [PubMed]