



Introduction
Sepsis is defined as life-threatening organ dysfunction caused by a dysregulated immune response to infection, which means that sepsis is diagnosed when the body’s response to infection results in life-threatening damage to its tissues or organs [1]. Sepsis is now the leading cause of mortality among patients in intensive care units [2]. The most significant and common complication of sepsis is Acute kidney injury (AKI) [3]. Sepsis-associated AKI (SA-AKI) accounts for 45–70% of all AKI cases in foreign intensive care units, with a mortality rate of 60–80% in AKI patients requiring dialysis [4]. Therefore, preventing and controlling the occurrence of SA-AKI and reducing the morbidity and mortality of septic patients is another challenge is another important public health concern. It has been shown that in sepsis, inflammatory mediators originating from pathogens and activated immune cells can act on renal tubular epithelial cells via a Toll-like receptor 4 (TLR4)-dependent pathway, leading to mitochondrial dysfunction and tubular epithelial cell injury [5]. Therefore, it is presumed that the inflammatory response, tubular epithelial cell injury, and renal microcirculatory disorders are all involved in the development of SA-AKI. However, the pathogenesis of SA-AKI remains unknown.
Astragalus membranaceus (AM) has historically been utilized primarily as a Qi-tonifying in traditional Chinese medicine. The active ingredients contained in Astragalus membranaceus and related plants have been investigated extensively [6]. Astragalus membranaceus is distributed across China, with the medicinal Astragalus most widely used Shanxi, Heilongjiang, and Inner Mongolia. Astragalus is rich in chemical constituents and much research has been conducted to isolate and purify the saponin, polysaccharide, and flavonoid components of Astragalus as well as their pharmacological effects [7, 8]. Astragalus has more than fifty saponins that exist in a discrete state. Astragaloside I/IV, Mauroisoflavone glycosides and Astragaloside Isoflavan are all found in abundance in Astragaloside membranaceus [9]. Astragaloside membranaceus possesses favorable pharmacological properties and plays a unique role in anti-inflammation, cardiovascular improvement, and neuroprotection [9]. Additionally, Astragalus contains trace elements such as proteins, amino acids, monosaccharides, and folic acid in trace levels. However, research on these trace elements is scarce [10]. Although Astragalus is of the most extensively used herbal remedies for the treatment of kidney diseases, the material basis for its action and mechanism of action are unknown. Certain herbal remedies have demonstrated significant benefits and high efficacy in inhibiting the inflammatory response, programmed cell death, i.e., apoptosis and oxidative stress. The therapeutic effects of herbal medicine have been extensively validated in animal models and even in humans [11]. However, the effects and specific mechanisms of Astragalus in the treatment of SA-AKI have not been reported.
The theory of network pharmacology was proposed by Hopkins in 2007 based on the scope of systems biology and pharmacology. The concept is based on multi-target research, which involves collecting and organizing data on target drugs, constructing bio-visualization networks, and analyzing the interactions between drugs and genes, targets, and diseases to reveal the mechanism of action from a systematic and holistic manner [12]. The mechanism of action of Chinese medicines in regulating various signaling pathways has now been applied to the study of the material basis for drug efficacy and mechanism of action of Chinese medicines. In recent years, as multi-omics integration research has advanced, the acquisition of targets in network pharmacology has largely relied on directly related experimental data, significantly improving the accuracy of network pharmacology mechanisms and pharmacodynamic substance bases. Additionally, as big data analysis and molecular docking technology advance, the accuracy of the network pharmacology will improve further [13]. In this study, we investigated the mechanism of Astragalus membranaceus’s anti-SA-AKI effect using network pharmacology. This was achieved by first investigating the active components, targets of action, and mechanisms of Astragalus membranaceus against SA-AKI.
Materials and Methods
Chemicals and reagents
LPS (E. coli 0111: B4) was obtained from Millipore Sigma. Astragalus membranaceus was purchased from Kang Qiao Traditional Chinese Medicine Decoction Pieces Co. Ltd. Astragaloside IV (purity above 98%) was purchased from Xi’an Sobeo Pharmaceutical Technology Co., Ltd. The antibodies used for Western blotting, PI3K (#4255) and Phospho-PI3K p85 (Tyr458) (#17366) were purchased from Cell Signaling Technology. AKT (A17909) and Phospho-AKT (Ser473) (AP0637) were purchased from ABclonal.
Water extract of Astragalus membranaceus
Astragalus slices (500 g) were weighed, distilled water was added in a ration of 1:10, soaked for 2–3 h, then decocted and extracted 3 times at 90°C for 1.5 h. The decoction was filtered twice with gauze and the filtrate combined. The filtrate was concentrated to a viscous liquid using a rotary evaporator at 50°C, poured into trays, and dried in a water bath set at 65°C to form an infusion. The infusion was frozen for two days at −80°C and then lyophilized to form a powder in a freeze dryer.
Animals
Male C57BL/6J mice (18–22 g) were purchased from the Changzhou Cavens Laboratory Animal Co., China. All mice were housed in an automated 12-hour dark-light cycle at a controlled temperature of 22°C ± 2°C and relative humidity of 50–60%, with free access to standard dry feed and tap water. All animal experiments were conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals (National Academies Press, 2011) and were approved by the Naval Medical University Committee on Animal Care (EC11-055).
Sepsis model
Sepsis was induced by CLP as previously described [14]. Isoflurane was used to completely anesthetize the mice, and a midline abdominal incision was made. The ileocecal valve was ligated on the distal 3/4 of the cecum. in a sterile environment, the cecum was perforated twice with a 21-gauge needle, and a droplet of feces was extruded from the perforations to induce polymicrobial peritonitis. Sutures were placed in two layers along the abdomen wall and 1 ml of a 0.9% sodium chloride solution was administered for fluid resuscitation. The sham group underwent laparotomy and bowel manipulation without ligation or perforation. Following recovery from anesthesia, all mice had unrestricted access to food and water. There were three groups of six animals each for Astragalus membranaceus: the sham-operated group (Sham group), the sepsis model group (CLP group), and treatment groups with Astragalus membranaceus (250 mg/kg). Astragalus membranaceus was administrated orally every day before CLP. There were four groups of six animals each in the AS-IV study, a sham group, a CLP group, and two treatment groups: AS-IV low dose (5 mg/kg) and AS-IV high dose (10 mg/kg). AS-IV was administrated orally every day before CLP.
Kidney injury study
Kidney tissues were cut into 5 μm sections and stained with H&E, PAS, and nitrotyrosine reagents, as well as KIM-1 antibody and Phospho-AKT antibody. The kidney injury score is as follows: 0, normal; 1, <10%; 2, 10–25%; 3, 25–50%; 4, 50–75%; 5, 75–100% of affected kidney areas obtained from mice exposed to sham or CLP surgery. Tubular atrophy or dilatation, loss of the brush border, vacuolization, epithelial cell shedding, and denuded tubular basement membrane were some of the criteria.
Cell culture
HK-2 cells (CRL-2190, ATCC, Rockefeller, MD, USA) were maintained in RPMI1640 (31800022, Gibco, NY, USA) supplemented with 10% v/v FBS, 100 U/ml penicillin, and 100 μg/ml streptomycin (Thermo Fisher Scientific, Waltham, MA, USA) in humidified incubators at 37°C and 5% CO2.
Bioinformatics analysis
The TCMSP server consisted of a system-level pharmacology database with a flexible, user-friendly web interface. To gain a better understanding of the complex, network targets genes and diseases were constructed and analyzed using Cytoscape 3.0.
Western blotting
The protein samples were electrophoresed in 8–12% SDS-PAGE gels and transferred onto nitrocellulose membranes (Amersham). The membranes were blocked with 5% BSA in PBS for 1 hour at room temperature and then probed overnight at 4°C with primary antibodies (1:1000 dilution). The membranes were then incubated with secondary antibodies (1:10,000 dilution) for 50 min at room temperature. Finally, Odyssey (LI-COR, Lincoln, NE, USA) and the relative protein expression were calculated from gray-scale blots using ImageJ software.
Statistics
GraphPad Prism 9.0.1 was used to perform statistical analysis and data were expressed as mean ± SEM. Statistical significance was determined using Student’s t-test or ANOVA. P < 0.05 was considered statistically significant.
Data statement
Our data are available. Please contact the corresponding author for requirement. More detailed Materials and Methods are in the Supplementary Materials and Methods.
Results
Astragalus membranaceus protects against renal injury in CLP-induced sepsis. In this study, the CLP model, a widely accepted mouse model of sepsis, was used to induce SA-AKI in C57BL/6J mice. Histopathological examination revealed that 24 hours after CLP surgery, the kidney tissue interstitial edema in the CLP mice was obvious, with diffuse neutrophil infiltration and severe glomerular wrinkling, while the tubular lumen was occluded, some cells were swollen and vacuolated, and the tubular cystic lumen was dilated. Further treatment with Astragalus membranaceus (AM) improved focal tubular damage, tubular swelling, and vacuolar degeneration as well as inflammatory cell infiltration and interstitial edema to varying degrees in mice kidney tissues (Figure 1A and 1B). Consistently, septic mice after AM treatment exhibited a markedly reduced kidney injury score (Figure 1C). Immunohistochemical analysis of kidney sections from treated septic mice revealed decreased staining for kidney injury molecule-1 (KIM-1) at the ends of renal tubular epithelial cells (Figure 1D). Furthermore, blood urea nitrogen (BUN) and creatinine levels, indicative of kidney dysfunction, were markedly decreased in mice of the treatment groups, suggesting preserved renal function for AM (Figure 1E and 1F). ELISA results confirmed that IL-6 and IL-1β concentrations in serum were deceased after AM treatment (Figure 1G).
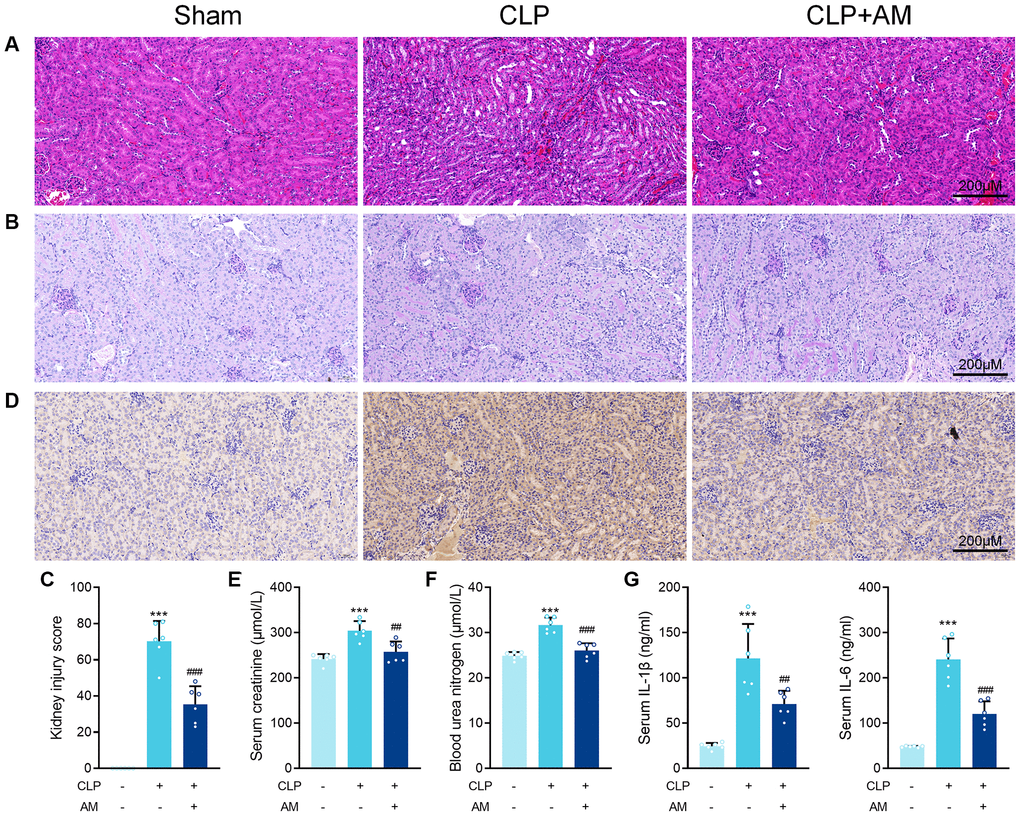
Figure 1. Astragalus membranaceus protects against renal injury in CLP-induced sepsis. (A) Representative images of H&E staining of kidney sections obtained at 24 hours after CLP surgery following treatment with (250 mg/kg) Astragalus membranaceus. (B) Representative images of periodic acid Schiff’s staining reagent depicting basement membrane and brush border in kidney sections. (C) Semiquantitative analysis of tubular injury. (D) Representative images showing KIM-1 immunohistochemical staining results in kidney tissues. Serum creatinine (E) and Blood urea nitrogen (F) in mice were measured. (G) Serum IL-6 and IL-1β concentrations were measured by ELISA. Data are presented as the mean ± SEM (n = 6 per group, ***P < 0.001 compared to Sham group, ##P < 0.01, ###P < 0.001 compared to CLP group).
The effect of Astragalus membranaceus active ingredients on LPS-induced damage on HK-2 cell line
Given that sepsis’s primary target is proximal tubular epithelial cells, HK-2 cells were treated with LPS to mimic Gram (–) induced sepsis-associated tubular injury. A total of 144 Astragalus-related compounds were identified through a database search. The TCMSP database was examined for OB and DL values of each compound, and 42 compounds (28.6%) had OB values greater than 30%, 50 compounds (40%) had DL values greater than 0.18, and 22 compounds (31.4%) had optimum OB and DL values. Astragalus saponins were excluded from the study due to their failure to meet the OB and DL thresholds. However, because these components are the most often reported active ingredients in the pharmacological action of Astragalus, they were included in this study. To screen for anti-SA-AKI activity, the final 26 monomers from Astragalus were collected (Table 1). We discovered nine compounds, including isorhamnetin, kaempferol, mangosteen, and astragaloside, that have been reported in “sepsis” or “acute kidney injury” and have been verified by the official website of the China Academy of Food and Drug Control. The nine compounds were screened further using LPS-induced HK-2 cells. The MTT assay was used to examine the effects of isorhamnetin (Iso), kaempferol (Kae), formononetin (Form), astragaloside I (AS-I), astragaloside II (AS-II), astragaloside IV (AS-IV), Mairin, Jaranol, and Calycosin (Cal) on LPS-induced HK-2 cell injury in vitro (Figure 2).
Table 1. Active ingredients of Astragalus membranaceus.
| No. | Compound | MOL ID | OB | DL |
| C1 | Mairin | MOL000211 | 55.38 | 0.78 |
| C2 | kaempferol | MOL000422 | 41.88 | 0.24 |
| C3 | Jaranol | MOL000239 | 50.83 | 0.29 |
| C4 | isorhamnetin | MOL000354 | 49.60 | 0.31 |
| C5 | isomucronulatol-7,2′-di-O-glucosiole | MOL000439 | 49.28 | 0.62 |
| C6 | isoflavanone | MOL000398 | 109.99 | 0.30 |
| C7 | hederagenin | MOL000296 | 36.91 | 0.75 |
| C8 | formononetin | MOL000392 | 69.67 | 0.21 |
| C9 | astragaloside IV | MOL000407 | 22.50 | 0.15 |
| C10 | astragaloside I | MOL000401 | 46.79 | 0.11 |
| C11 | astragaloside II | MOL000403 | 46.06 | 0.13 |
| C12 | astragaloside III | MOL000405 | 31.83 | 0.10 |
| C13 | Quercetin | MOL000098 | 46.43 | 0.28 |
| C14 | FA | MOL000433 | 68.96 | 0.71 |
| C15 | Calycosin | MOL000417 | 47.75 | 0.24 |
| C16 | Bifendate | MOL000387 | 31.10 | 0.67 |
| C17 | 9,10-dimethoxypterocarpan-3-O-β-D-glucoside | MOL000379 | 36.74 | 0.92 |
| C18 | MOL000378 | 74.69 | 0.30 | |
| C19 | 5′-hydroxyiso-muronulatol-2′,5′-di-O-glucoside | MOL000374 | 41.72 | 0.69 |
| C20 | 3,9-di-O-methylnissolin | MOL000371 | 53.74 | 0.48 |
| C21 | (6aR,11aR)-9,10-dimethoxy-6a,11a-dihydro-6H-benzofurano [3,2-c] chromen-3-ol | MOL000380 | 64.26 | 0.42 |
| C22 | (3R)-3-(2-hydroxy-3,4-dimethoxyphenyl) chroman-7-ol | MOL000438 | 67.67 | 0.26 |
| C23 | 1,7-Dihydroxy-3,9-dimethoxy pterocarpene | MOL000442 | 39.05 | 0.48 |
| C24 | (3S,8S,9S,10R,13R,14S,17R)-10,13-dimethyl-17-[(2R,5S)-5-propan-2-yloctan-2-yl]-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol | MOL000033 | 36.23 | 0.78 |
| C25 | β-sitosterol | MOL001987 | 33.94 | 0.70 |
| C26 | (+)-Medicarpin | MOL011076 | 60.46 | 0.34 |
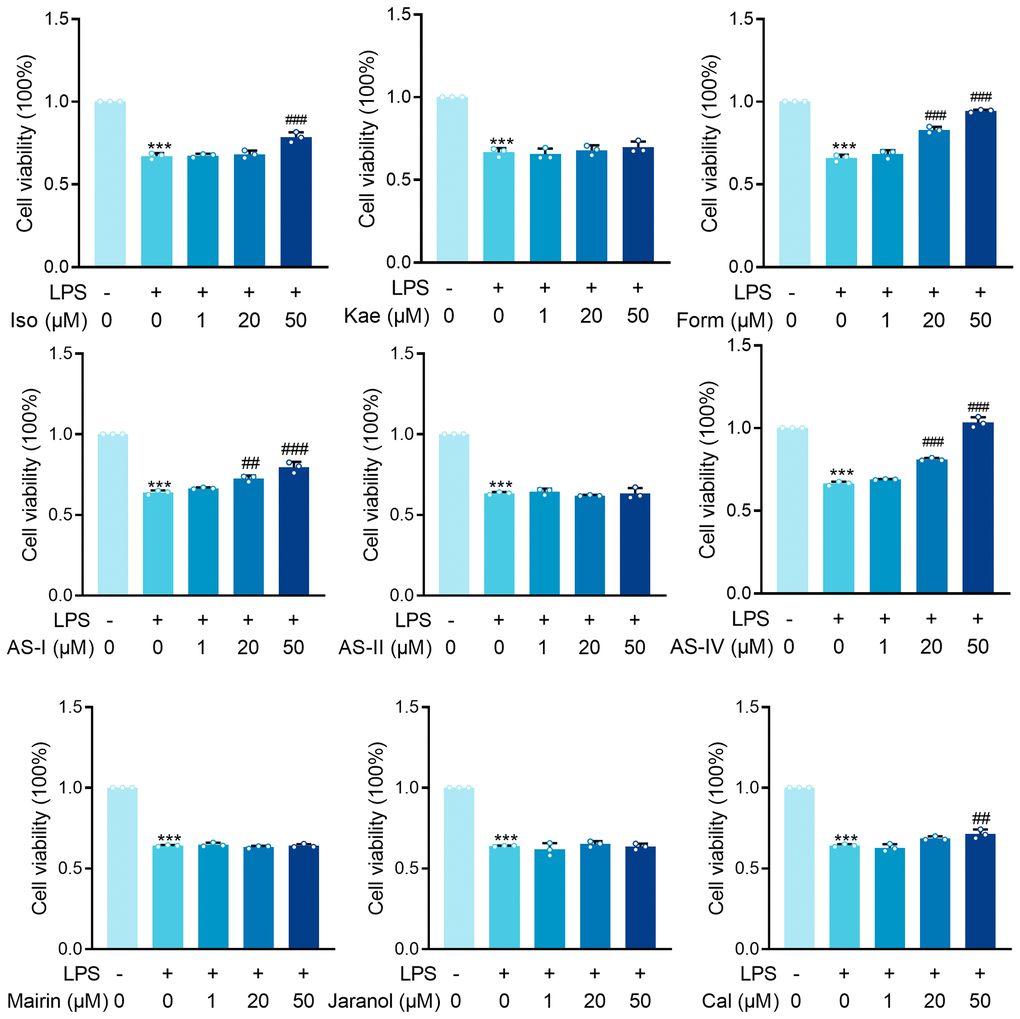
Figure 2. MTT results for isorhamnetin (Iso), kaempferol (Kae), formononetin (Form), Astragaloside I (AS-I), Astragaloside II (AS-II), Astragaloside IV (AS-IV), Mairin, Jaranol and Calycosin (Cal) treatment on renal epithelial cells HK-2 cells incubated with LPS (1 μg/ml) for 6 hours. Data are presented as the mean ± SEM (n = 3 per group, ***P < 0.001 compared to Control group, ##P < 0.01, ###P < 0.001 compared to LPS group).
The effect of LPS on HK-2 cells
LPS significantly inhibited the survival rate of HK-2 cells (P < 0.001). Astragaloside I (AS-I) and Astragaloside IV (AS-IV) demonstrated dose-dependent protective effects at 20 μM versus 50 μM concentrations (AS-I: P < 0.01, P < 0.001; Form: P < 0.001, P < 0.001; AS-IV: P < 0.001, P < 0.001). At 50 μM Iso and Cal showed significant protection (P < 0.001). In contrast, treatment of HK-2 cells with Kae, AS-II, Mairin, and Jaranol did not result in any significant changes in the LPS-induced cytotoxicity (Figure 2). Our results demonstrated that AS-I, AS-IV, Iso, and Cal all exerted protective effects against LPS-induced HK-2 cytotoxicity in renal epithelial cells at 50 μM concentration, with AS-IV exhibiting the most pronounced protective effect. Therefore, we further investigated the mechanism through which AS-IV protects against septic acute kidney injury.
AS-IV protects against renal injury in CLP-induced sepsis
The China Pharmacopeia 2015 specifies that the content of AS-IV in Astragalus should not be less than 0.040%. Using HPLC analysis, we determined the AS-IV content of Astragalus to be 0.147%, which met the quality standard requirements stipulated in the China Pharmacopeia 2015 (Supplementary Figure 1). To determine the protective effect of AS-IV on SA-AKI-induced renal damage, we subjected mice to CLP or sham surgery. The treatment group was divided into two groups, low-dose (i.p. 5 mg/kg) and high-dose (i.p. 10 mg/kg). Similarly, the AS-IV 5 mg/kg and 10 gm/kg groups had considerably fewer intracellular vacuoles (Figure 3A).
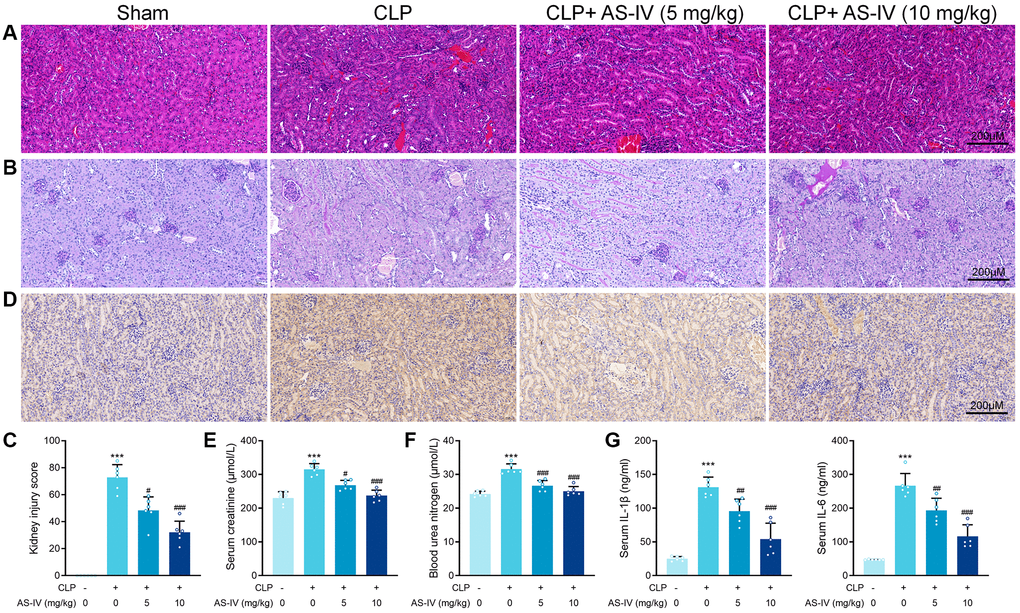
Figure 3. AS-IV protects against renal injury in CLP-induced sepsis. (A) Representative images of H&E staining of tubular epithelial cells of kidney sections obtained at 24 hours after CLP surgery following treatment with low (5 mg/kg) or high dosage (10 mg/kg) AS-IV. (B) Representative images of periodic acid Schiff’s staining reagent depicting basement membrane and brush border in the kidney sections. (C) Semiquantitative analysis of tubular injury. (D) Representative images showing KIM-1 immunohistochemical staining of the apical side of tubular epithelial cells. Serum creatinine (E) and Blood urea nitrogen (F) in mice were measured. (G) Serum IL-6 and IL-1β concentrations were measured by ELISA. Data are presented as the mean ± SEM (n = 6 per group, ***P < 0.001 compared to Sham group, #P < 0.05, ##P < 0.01, ###P < 0.001 compared to CLP group).
Additionally, the treatment groups showed significant loss of brush border in the renal proximal tubular epithelial cells caused by CLP (Figure 3B). After AS-IV treatment, mice showed a markedly decrease kidney injury score (Figure 3C). Immunohistochemical analysis revealed decreased staining of KIM-1 on the apical side of renal tubular epithelial cells in kidney sections from treated septic mice (Figure 3D). Next, BUN and creatinine levels were markedly decreased in mice of the treatment groups, indicating preserved renal function for AS-IV (Figure 3E and 3F). ELISA results confirmed that IL-6 and IL-1β concentrations in serum were deceased after AS-IV treatment (Figure 3G).
Astragalus membranaceus and AS-IV for SA-AKI target prediction
The therapeutic target database (TTD), Genecards, the Comparative Toxicogenomics Database (CTD), and the CTD were searched using the terms “sepsis” and “acute kidney damage (AKI). Genecards and the Comparative Toxicogenomics Database (CTD) were used to search for relevant disease targets, and the CTD database was used to identify experimentally validated disease targets designated as “marker/mechanism” or “therapeutic”. The CTD database selects targets with a high degree of confidence that has been experimentally validated with “marker/mechanism” or “therapeutic” as targets for research. Finally, 2498 sepsis-related genes and 6746 AKI-related genes were retrieved from the GeneCards database, while 48 sepsis-related genes and 120 AKI-related genes were retrieved from the CTD database. A total of 1938 targets were obtained after a comprehensive screening of all targets to eliminate duplicates, and the DAVID database validated a total of 1910 targets related to sepsis and acute kidney injury in humans. All genes were imported into the DAVID database, and the relevant genes were exported by selecting and clicking “show gene lis”. GO enrichment and KEGG pathway enrichment analyses were performed on the disease targets via the DAVID database, using P < 0.01 and False discovery rate (False discovery rate (FDR) <0.01 as the screening thresholds. A total of 686 entries for biological process (BP), 104 for cellular component (CC), and 121 for molecular function (MF) were obtained. The top 20 entries are displayed in Tables 2 and 3 below, are ordered by the magnitude of their P and FDR values. SA-AKI was predicted to be involved in biological processes such as the inflammatory response, immune response, intrinsic immune response, response to LPS, response to hypoxic factors, etc. The cellular components were involved in extracellular gaps, extracellular regions, molecular functions including protein binding, receptor binding, enzyme binding, etc. Astragalus and SA-AKI targets were intersected by Venn to obtain a total of 275 core targets (Figure 4A). AS-IV and SA-AKI targets were intersected by Venn to obtain a total of 167 core targets (Figure 4B). We further examined the relationship between the bio-active ingredients and the core targets, to construct an interaction network between the two (Supplementary Figure 2). A total of 26 active ingredients of Astragalus were identified to be critical to the core target. The high-degree nodes in the network exhibited a greater number of component-target-pathway interactions, which were likely to play a critical role in renal protective AKI. Following that, the Astragalus Target-SA-AKI Target-PPI Network input the compound targets into TCMSP and String databases. There were 275 nodes and 741 edges in the Astragalus Target-SA-AKI Target-PPI Network (Figure 4C). Similarly, the AS-IV Target-SA-AKI Target-PPI Network Input the compound targets into TCMSP and String databases. There were 167 nodes and 2413 edges in the AS-IV Target-SA-AKI Target-PPI Network (Figure 4D). We imported the 254 target genes of Astragalus anti-SA-AKI and the 167 target gene sets of AS-IV anti-SA-AKI into the DAVID version 6.7 database for Gene ontology analysis and KEGG pathway enrichment analysis, using three panels typically included in GO analysis. For Astragalus, a total of 241 MF entries, 3045 BF entries, 123 CC entries, and 169 KEGG entries were obtained. Following that, we screened at P < 0.01 and FDR <0.01 to extract significant card values and discovered 2028 entries under BP, 63 entries under CC, and 132 entries under MF. Figure 4E and Table 4 show the top 10 entries selected for visualization. As shown in Figure 4F and Table 5, 140 KEGG entries were selected for visualization. For AS-IV, 168 entries were obtained under MF, 2661 entries under BP, 64 entries under CC, and 147 entries under KEGG. We used P < 0.01 and FDR <0.01 as significant card values and obtained 1667 entries under BP, 43 entries under CC, and 89 entries under MF. The top 10 entries were selected for visualization, as shown in Figure 4G and Table 6. 114 entries under KEGG were selected for visualization, as shown in Figure 4H and Table 7.
Table 2. GO enrichment analysis of disease mechanisms in SA-AKI.
| Ontology | ID | Description | P value | FDR |
| BP | 0006954 | inflammatory response | 4.41E-98 | 2.71E-94 |
| BP | 0042493 | response to drug | 9.21E-58 | 2.83E-54 |
| BP | 0006955 | immune response | 2.16E-55 | 4.43E-52 |
| BP | 0045087 | innate immune response | 1.08E-48 | 1.66E-45 |
| BP | 0032496 | response to lipopolysaccharide | 2.50E-46 | 3.08E-43 |
| BP | 0001666 | response to hypoxia | 5.70E-43 | 5.19E-40 |
| BP | 0043066 | negative regulation of apoptotic process | 5.90E-43 | 5.19E-40 |
| BP | 0008284 | positive regulation of cell proliferation | 7.95E-41 | 6.11E-38 |
| BP | 0051092 | positive regulation of NF-kappaB transcription factor activity | 3.14E-39 | 2.15E-36 |
| BP | 0050900 | leukocyte migration | 1.88E-37 | 1.16E-34 |
| BP | 0071222 | cellular response to lipopolysaccharide | 4.05E-36 | 2.26E-33 |
| BP | 0045944 | positive regulation of transcription from RNA polymerase II promoter | 1.28E-35 | 6.58E-33 |
| BP | 0007165 | signal transduction | 1.86E-34 | 8.82E-32 |
| BP | 0070374 | positive regulation of ERK1 and ERK2 cascade | 6.05E-33 | 2.66E-30 |
| BP | 0007568 | aging | 1.46E-32 | 6.01E-30 |
| BP | 0045471 | response to ethanol | 2.73E-32 | 1.05E-29 |
| BP | 0010628 | positive regulation of gene expression | 2.58E-30 | 9.35E-28 |
| BP | 0045766 | positive regulation of angiogenesis | 2.86E-30 | 9.79E-28 |
| BP | 0006915 | apoptotic process | 1.04E-29 | 3.36E-27 |
| BP | 0050731 | positive regulation of peptidyl-tyrosine phosphorylation | 3.21E-29 | 9.89E-27 |
| CC | 0005615 | extracellular space | 3.41E-141 | 2.46E-138 |
| CC | 0005576 | extracellular region | 1.01E-97 | 3.64E-95 |
| CC | 0070062 | extracellular exosome | 5.69E-82 | 1.37E-79 |
| CC | 0009986 | cell surface | 1.58E-54 | 2.85E-52 |
| CC | 0005886 | plasma membrane | 3.42E-54 | 4.94E-52 |
| CC | 0009897 | external side of plasma membrane | 1.55E-51 | 1.87E-49 |
| CC | 0005829 | cytosol | 1.28E-44 | 1.32E-42 |
| CC | 0045121 | membrane raft | 2.26E-37 | 2.04E-35 |
| CC | 0005925 | focal adhesion | 4.75E-33 | 3.82E-31 |
| CC | 0005887 | integral component of plasma membrane | 7.79E-32 | 5.63E-30 |
| CC | 0016020 | membrane | 1.32E-28 | 8.68E-27 |
| CC | 0072562 | blood microparticle | 4.56E-26 | 2.75E-24 |
| CC | 0031012 | extracellular matrix | 4.24E-24 | 2.36E-22 |
| CC | 0031093 | platelet alpha granule lumen | 8.12E-21 | 4.20E-19 |
| CC | 0043202 | lysosomal lumen | 2.13E-18 | 1.03E-16 |
| CC | 0043234 | protein complex | 1.55E-17 | 6.82E-16 |
| CC | 0048471 | perinuclear region of cytoplasm | 1.60E-17 | 6.82E-16 |
| CC | 0005764 | lysosome | 2.36E-17 | 9.47E-16 |
| CC | 0005796 | Golgi lumen | 1.88E-14 | 7.16E-13 |
| CC | 0005911 | cell-cell junction | 5.39E-13 | 1.95E-11 |
| MF | 0005515 | protein binding | 7.49E-56 | 1.32E-52 |
| MF | 0005102 | receptor binding | 1.57E-42 | 1.38E-39 |
| MF | 0019899 | enzyme binding | 1.75E-31 | 1.03E-28 |
| MF | 0042803 | protein homodimerization activity | 2.56E-31 | 1.13E-28 |
| MF | 0042802 | identical protein binding | 7.35E-29 | 2.59E-26 |
| MF | 0005125 | cytokine activity | 3.02E-27 | 8.89E-25 |
| MF | 0008083 | growth factor activity | 1.62E-20 | 4.09E-18 |
| MF | 0002020 | protease binding | 5.13E-20 | 1.13E-17 |
| MF | 0004872 | receptor activity | 7.69E-19 | 1.51E-16 |
| MF | 0001618 | virus receptor activity | 2.08E-18 | 3.67E-16 |
| MF | 0019901 | protein kinase binding | 3.64E-18 | 5.83E-16 |
| MF | 0005088 | Ras guanyl-nucleotide exchange factor activity | 1.84E-16 | 2.70E-14 |
| MF | 0005179 | hormone activity | 2.08E-16 | 2.83E-14 |
| MF | 0008201 | heparin binding | 6.82E-16 | 8.60E-14 |
| MF | 0001948 | glycoprotein binding | 7.96E-16 | 9.36E-14 |
| MF | 0004252 | serine-type endopeptidase activity | 3.97E-15 | 4.38E-13 |
| MF | 0046934 | phosphatidylinositol-4,5-bisphosphate 3-kinase activity | 1.00E-14 | 1.04E-12 |
| MF | 0004713 | protein tyrosine kinase activity | 2.83E-14 | 2.77E-12 |
| MF | 0005164 | tumor necrosis factor receptor binding | 1.30E-13 | 1.20E-11 |
| MF | 0005178 | integrin binding | 1.36E-13 | 1.20E-11 |
Table 3. KEGG enrichment analysis of disease mechanisms in SA-AKI.
| Ontology | ID | Description | P value | FDR |
| KEGG_PATHWAY | hsa04668 | TNF signaling pathway | 5.13E-34 | 8.61E-32 |
| KEGG_PATHWAY | hsa05145 | Toxoplasmosis | 1.38E-31 | 1.16E-29 |
| KEGG_PATHWAY | hsa04064 | NF-kappa B signaling pathway | 6.61E-31 | 3.39E-29 |
| KEGG_PATHWAY | hsa04060 | Cytokine-cytokine receptor interaction | 6.61E-31 | 3.39E-29 |
| KEGG_PATHWAY | hsa04610 | Complement and coagulation cascades | 1.01E-30 | 3.39E-29 |
| KEGG_PATHWAY | hsa05200 | Pathways in cancer | 2.62E-30 | 7.33E-29 |
| KEGG_PATHWAY | hsa04380 | Osteoclast differentiation | 6.57E-30 | 1.58E-28 |
| KEGG_PATHWAY | hsa05152 | Tuberculosis | 1.99E-29 | 4.17E-28 |
| KEGG_PATHWAY | hsa04620 | Toll-like receptor signaling pathway | 2.30E-27 | 4.29E-26 |
| KEGG_PATHWAY | hsa05140 | Leishmaniasis | 5.90E-27 | 9.91E-26 |
| KEGG_PATHWAY | hsa04640 | Hematopoietic cell lineage | 1.54E-26 | 2.35E-25 |
| KEGG_PATHWAY | hsa05164 | Influenza A | 4.85E-25 | 6.79E-24 |
| KEGG_PATHWAY | hsa04062 | Chemokine signaling pathway | 6.48E-25 | 8.37E-24 |
| KEGG_PATHWAY | hsa04660 | T cell receptor signaling pathway | 3.49E-23 | 4.19E-22 |
| KEGG_PATHWAY | hsa05133 | Pertussis | 1.03E-22 | 1.15E-21 |
| KEGG_PATHWAY | hsa05142 | Chagas disease (American trypanosomiasis) | 1.17E-22 | 1.22E-21 |
| KEGG_PATHWAY | hsa05161 | Hepatitis B | 2.31E-22 | 2.28E-21 |
| KEGG_PATHWAY | hsa05162 | Measles | 7.21E-22 | 6.73E-21 |
| KEGG_PATHWAY | hsa05321 | Inflammatory bowel disease (IBD) | 1.36E-21 | 1.21E-20 |
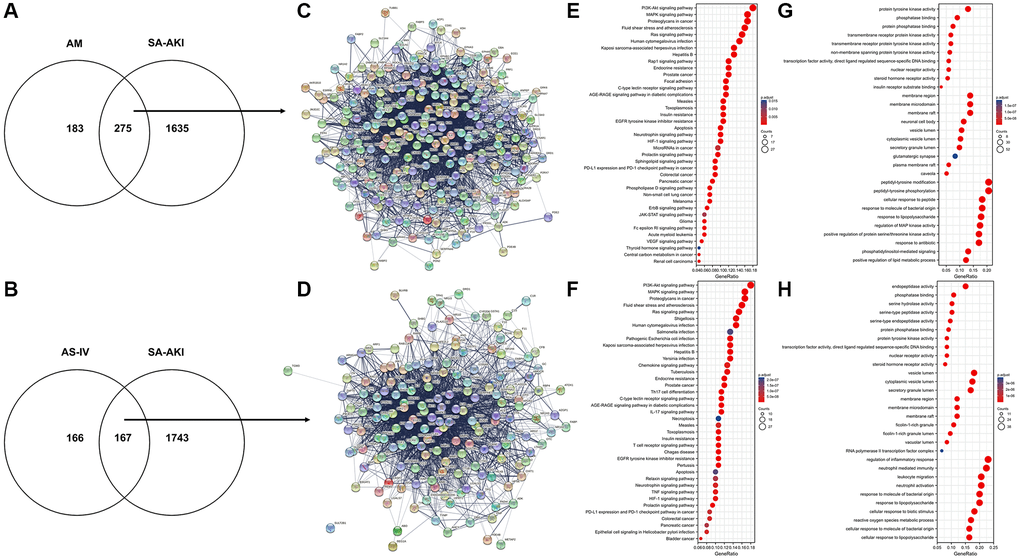
Figure 4. Astragalus membranaceus and AS-IV for SA-AKI target prediction. (A) Venn diagram of Astragalus membranaceus active compounds-related targets and SA-AKI-related targets. (B) Venn diagram of AS-IV active compounds-related targets and SA-AKI-related targets. (C) PPI network of potential targets of Astragalus membranaceus anti-SA-AKI. (D) PPI network of potential targets of AS-IV anti-SA-AKI. (E) KEGG pathway analysis of potential targets of Astragalus membranaceus anti-SA-AKI. (F) KEGG pathway analysis of potential targets of AS-IV anti-SA-AKI. (G) GO analysis of potential targets of Astragalus membranaceus anti-SA-AKI. (H) GO analysis potential targets of AS-IV anti-SA-AKI.
Table 4. GO enrichment analysis of potentially relevant targets of Astragalus anti-SA-AKI.
| Ontology | ID | Description | GeneRatio | BgRatio | p value | FDR | q value |
| BP | GO:0018108 | peptidyl-tyrosine phosphorylation | 52/252 | 363/18670 | 2.78E-38 | 1.01E-34 | 4.03E-35 |
| BP | GO:0018212 | peptidyl-tyrosine modification | 52/252 | 366/18670 | 4.27E-38 | 1.01E-34 | 4.03E-35 |
| BP | GO:0002237 | response to molecule of bacterial origin | 46/252 | 343/18670 | 1.62E-32 | 2.56E-29 | 1.02E-29 |
| BP | GO:0032496 | response to lipopolysaccharide | 45/252 | 330/18670 | 3.94E-32 | 4.66E-29 | 1.86E-29 |
| BP | GO:0043405 | regulation of MAP kinase activity | 44/252 | 337/18670 | 1.35E-30 | 1.27E-27 | 5.09E-28 |
| BP | GO:1901653 | cellular response to peptide | 46/252 | 385/18670 | 2.97E-30 | 2.34E-27 | 9.35E-28 |
| BP | GO:0046677 | response to antibiotic | 43/252 | 327/18670 | 4.90E-30 | 3.32E-27 | 1.32E-27 |
| BP | GO:0071902 | positive regulation of protein serine/threonine kinase activity | 43/252 | 334/18670 | 1.20E-29 | 7.11E-27 | 2.84E-27 |
| BP | GO:0045834 | positive regulation of lipid metabolic process | 31/252 | 146/18670 | 2.11E-28 | 1.11E-25 | 4.42E-26 |
| BP | GO:0048015 | phosphatidylinositol-mediated signaling | 33/252 | 181/18670 | 6.82E-28 | 3.23E-25 | 1.29E-25 |
| CC | GO:0045121 | membrane raft | 35/253 | 315/19717 | 9.60E-23 | 1.99E-20 | 1.34E-20 |
| CC | GO:0098857 | membrane microdomain | 35/253 | 316/19717 | 1.07E-22 | 1.99E-20 | 1.34E-20 |
| CC | GO:0098589 | membrane region | 35/253 | 328/19717 | 3.72E-22 | 4.61E-20 | 3.10E-20 |
| CC | GO:0031983 | vesicle lumen | 27/253 | 339/19717 | 4.05E-14 | 3.76E-12 | 2.53E-12 |
| CC | GO:0060205 | cytoplasmic vesicle lumen | 26/253 | 338/19717 | 2.77E-13 | 2.06E-11 | 1.39E-11 |
| CC | GO:0034774 | secretory granule lumen | 25/253 | 321/19717 | 6.18E-13 | 3.83E-11 | 2.58E-11 |
| CC | GO:0044853 | plasma membrane raft | 15/253 | 109/19717 | 9.65E-12 | 5.13E-10 | 3.45E-10 |
| CC | GO:0043025 | neuronal cell body | 29/253 | 497/19717 | 1.11E-11 | 5.15E-10 | 3.47E-10 |
| CC | GO:0005901 | caveola | 13/253 | 80/19717 | 2.76E-11 | 1.14E-09 | 7.68E-10 |
| CC | GO:0098978 | glutamatergic synapse | 21/253 | 349/19717 | 5.20E-09 | 1.93E-07 | 1.30E-07 |
| MF | GO:0004713 | protein tyrosine kinase activity | 33/253 | 134/17697 | 1.09E-31 | 7.11E-29 | 4.46E-29 |
| MF | GO:0004715 | non-membrane spanning protein tyrosine kinase activity | 16/253 | 46/17697 | 1.28E-18 | 4.16E-16 | 2.61E-16 |
| MF | GO:0004714 | transmembrane receptor protein tyrosine kinase activity | 17/253 | 62/17697 | 1.06E-17 | 2.29E-15 | 1.44E-15 |
| MF | GO:0004879 | nuclear receptor activity | 15/253 | 47/17697 | 7.02E-17 | 9.13E-15 | 5.72E-15 |
| MF | GO:0098531 | transcription factor activity, direct ligand regulated sequence-specific DNA binding | 15/253 | 47/17697 | 7.02E-17 | 9.13E-15 | 5.72E-15 |
| MF | GO:0019199 | transmembrane receptor protein kinase activity | 17/253 | 79/17697 | 9.21E-16 | 9.98E-14 | 6.25E-14 |
| MF | GO:0019902 | phosphatase binding | 23/253 | 185/17697 | 2.27E-15 | 2.11E-13 | 1.32E-13 |
| MF | GO:0003707 | steroid hormone receptor activity | 14/253 | 56/17697 | 3.54E-14 | 2.87E-12 | 1.80E-12 |
| MF | GO:0019903 | protein phosphatase binding | 19/253 | 140/17697 | 1.33E-13 | 9.57E-12 | 6.00E-12 |
| MF | GO:0043560 | insulin receptor substrate binding | 8/253 | 11/17697 | 2.48E-13 | 1.61E-11 | 1.01E-11 |
Table 5. KEGG enrichment analysis of potentially relevant targets of Astragalus anti-SA-AKI.
| Ontology | ID | Description | GeneRatio | BgRatio | p value | FDR | q value |
| KEGG | hsa05418 | Fluid shear stress and atherosclerosis | 24/150 | 139/8076 | 3.87E-17 | 9.95E-15 | 4.57E-15 |
| KEGG | hsa05205 | Proteoglycans in cancer | 25/150 | 205/8076 | 3.74E-14 | 4.81E-12 | 2.21E-12 |
| KEGG | hsa05215 | Prostate cancer | 18/150 | 97/8076 | 1.19E-13 | 9.19E-12 | 4.22E-12 |
| KEGG | hsa01522 | Endocrine resistance | 18/150 | 98/8076 | 1.43E-13 | 9.19E-12 | 4.22E-12 |
| KEGG | hsa01521 | EGFR tyrosine kinase inhibitor resistance | 16/150 | 79/8076 | 7.08E-13 | 2.60E-11 | 1.19E-11 |
| KEGG | hsa04933 | AGE-RAGE signaling pathway in diabetic complications | 17/150 | 100/8076 | 2.66E-12 | 7.59E-11 | 3.48E-11 |
| KEGG | hsa04625 | C-type lectin receptor signaling pathway | 17/150 | 104/8076 | 5.15E-12 | 1.32E-10 | 6.08E-11 |
| KEGG | hsa05161 | Hepatitis B | 20/150 | 162/8076 | 1.33E-11 | 2.86E-10 | 1.31E-10 |
| KEGG | hsa04917 | Prolactin signaling pathway | 14/150 | 70/8076 | 2.50E-11 | 4.94E-10 | 2.26E-10 |
| KEGG | hsa04014 | Ras signaling pathway | 23/150 | 232/8076 | 3.42E-11 | 6.29E-10 | 2.88E-10 |
| KEGG | hsa04931 | Insulin resistance | 16/150 | 108/8076 | 1.06E-10 | 1.60E-09 | 7.32E-10 |
| KEGG | hsa05163 | Human cytomegalovirus infection | 22/150 | 225/8076 | 1.26E-10 | 1.70E-09 | 7.82E-10 |
| KEGG | hsa04010 | MAPK signaling pathway | 25/150 | 294/8076 | 1.26E-10 | 1.70E-09 | 7.82E-10 |
| KEGG | hsa05145 | Toxoplasmosis | 16/150 | 112/8076 | 1.85E-10 | 2.38E-09 | 1.09E-09 |
| KEGG | hsa04151 | PI3K-Akt signaling pathway | 27/150 | 354/8076 | 2.50E-10 | 3.06E-09 | 1.41E-09 |
| KEGG | hsa05167 | Kaposi sarcoma-associated herpesvirus infection | 20/150 | 193/8076 | 3.33E-10 | 3.90E-09 | 1.79E-09 |
| KEGG | hsa04066 | HIF-1 signaling pathway | 15/150 | 109/8076 | 1.21E-09 | 1.20E-08 | 5.50E-09 |
| KEGG | hsa04722 | Neurotrophin signaling pathway | 15/150 | 119/8076 | 4.22E-09 | 3.61E-08 | 1.66E-08 |
| KEGG | hsa05162 | Measles | 16/150 | 139/8076 | 4.80E-09 | 3.85E-08 | 1.77E-08 |
| KEGG | hsa05210 | Colorectal cancer | 13/150 | 86/8076 | 5.04E-09 | 3.93E-08 | 1.80E-08 |
| KEGG | hsa05235 | PD-L1 expression and PD-1 checkpoint pathway in cancer | 13/150 | 89/8076 | 7.76E-09 | 5.87E-08 | 2.69E-08 |
| KEGG | hsa05212 | Pancreatic cancer | 12/150 | 76/8076 | 1.21E-08 | 8.89E-08 | 4.08E-08 |
| KEGG | hsa04210 | Apoptosis | 15/150 | 136/8076 | 2.69E-08 | 1.87E-07 | 8.56E-08 |
| KEGG | hsa04015 | Rap1 signaling pathway | 18/150 | 210/8076 | 5.52E-08 | 3.38E-07 | 1.55E-07 |
| KEGG | hsa05218 | Melanoma | 11/150 | 72/8076 | 7.18E-08 | 4.19E-07 | 1.92E-07 |
| KEGG | hsa05223 | Non-small cell lung cancer | 11/150 | 72/8076 | 7.18E-08 | 4.19E-07 | 1.92E-07 |
| KEGG | hsa04510 | Focal adhesion | 17/150 | 201/8076 | 1.61E-07 | 8.47E-07 | 3.88E-07 |
| KEGG | hsa04071 | Sphingolipid signaling pathway | 13/150 | 119/8076 | 2.67E-07 | 1.32E-06 | 6.06E-07 |
| KEGG | hsa04012 | ErbB signaling pathway | 10/150 | 85/8076 | 3.44E-06 | 1.42E-05 | 6.53E-06 |
| KEGG | hsa05221 | Acute myeloid leukemia | 9/150 | 67/8076 | 3.53E-06 | 1.42E-05 | 6.53E-06 |
| KEGG | hsa04664 | Fc epsilon RI signaling pathway | 9/150 | 68/8076 | 4.01E-06 | 1.56E-05 | 7.16E-06 |
| KEGG | hsa05214 | Glioma | 9/150 | 75/8076 | 9.14E-06 | 3.41E-05 | 1.56E-05 |
| KEGG | hsa04370 | VEGF signaling pathway | 8/150 | 59/8076 | 1.17E-05 | 4.23E-05 | 1.94E-05 |
| KEGG | hsa04072 | Phospholipase D signaling pathway | 11/150 | 148/8076 | 9.06E-05 | 2.88E-04 | 1.32E-04 |
| KEGG | hsa05211 | Renal cell carcinoma | 7/150 | 69/8076 | 2.73E-04 | 7.88E-04 | 3.61E-04 |
| KEGG | hsa05230 | Central carbon metabolism in cancer | 7/150 | 70/8076 | 2.98E-04 | 8.50E-04 | 3.90E-04 |
| KEGG | hsa05206 | MicroRNAs in cancer | 14/150 | 310/8076 | 0.002 | 0.004 | 0.002 |
| KEGG | hsa04630 | JAK-STAT signaling pathway | 9/150 | 162/8076 | 0.003 | 0.007 | 0.003 |
| KEGG | hsa04919 | Thyroid hormone signaling pathway | 7/150 | 121/8076 | 0.007 | 0.015 | 0.007 |
Table 6. GO enrichment analysis of potentially relevant targets of AS-IV anti-SA-AKI.
| Ontology | ID | Description | GeneRatio | BgRatio | p value | FDR | q value |
| BP | GO:0071216 | cellular response to biotic stimulus | 30/165 | 23 6/18670 | 2.99E-26 | 1.27E-22 | 5.26E-23 |
| BP | GO:0032496 | response to lipopolysaccharide | 33/165 | 330/18670 | 2.03E-25 | 3.21E-22 | 1.32E-22 |
| BP | GO:0050727 | regulation of inflammatory response | 38/165 | 485/18670 | 2.26E-25 | 3.21E-22 | 1.32E-22 |
| BP | GO:0002237 | response to molecule of bacterial origin | 33/165 | 343/18670 | 7.08E-25 | 7.55E-22 | 3.11E-22 |
| BP | GO:0071222 | cellular response to lipopolysaccharide | 27/165 | 205/18670 | 4.25E-24 | 3.62E-21 | 1.49E-21 |
| BP | GO:0002446 | neutrophil mediated immunity | 37/165 | 499/18670 | 7.35E-24 | 5.22E-21 | 2.16E-21 |
| BP | GO:0071219 | cellular response to molecule of bacterial origin | 27/165 | 212/18670 | 1.06E-23 | 6.48E-21 | 2.67E-21 |
| BP | GO:0072593 | reactive oxygen species metabolic process | 28/165 | 284/18670 | 1.85E-21 | 9.83E-19 | 4.06E-19 |
| BP | GO:0042119 | neutrophil activation | 34/165 | 498/18670 | 8.71E-21 | 3.84E-18 | 1.59E-18 |
| BP | GO:0050900 | leukocyte migration | 34/165 | 499/18670 | 9.28E-21 | 3.84E-18 | 1.59E-18 |
| CC | GO:0031983 | vesicle lumen | 30/166 | 339/19717 | 3.61E-22 | 1.03E-19 | 7.75E-20 |
| CC | GO:0060205 | cytoplasmic vesicle lumen | 29/166 | 338/19717 | 4.53E-21 | 6.44E-19 | 4.87E-19 |
| CC | GO:0034774 | secretory granule lumen | 28/166 | 321/19717 | 1.48E-20 | 1.40E-18 | 1.06E-18 |
| CC | GO:1904813 | ficolin-1-rich granule lumen | 16/166 | 124/19717 | 7.59E-15 | 5.39E-13 | 4.07E-13 |
| CC | GO:0101002 | ficolin-1-rich granule | 18/166 | 185/19717 | 2.27E-14 | 1.29E-12 | 9.74E-13 |
| CC | GO:0045121 | membrane raft | 20/166 | 315/19717 | 2.47E-12 | 1.06E-10 | 8.03E-11 |
| CC | GO:0098857 | membrane microdomain | 20/166 | 316/19717 | 2.62E-12 | 1.06E-10 | 8.03E-11 |
| CC | GO:0098589 | membrane region | 20/166 | 328/19717 | 5.18E-12 | 1.84E-10 | 1.39E-10 |
| CC | GO:0005775 | vacuolar lumen | 14/166 | 172/19717 | 2.17E-10 | 6.83E-09 | 5.17E-09 |
| CC | GO:0090575 | RNA polymerase II transcription factor complex | 11/166 | 163/19717 | 1.37E-07 | 3.90E-06 | 2.95E-06 |
| MF | GO:0004879 | nuclear receptor activity | 14/166 | 47/17697 | 6.12E-18 | 1.53E-15 | 1.02E-15 |
| MF | GO:0098531 | transcription factor activity, direct ligand regulated sequence-specific DNA binding | 14/166 | 47/17697 | 6.12E-18 | 1.53E-15 | 1.02E-15 |
| MF | GO:0003707 | steroid hormone receptor activity | 13/166 | 56/17697 | 3.61E-15 | 6.02E-13 | 4.02E-13 |
| MF | GO:0019902 | phosphatase binding | 18/166 | 185/17697 | 1.38E-13 | 1.73E-11 | 1.16E-11 |
| MF | GO:0004175 | endopeptidase activity | 25/166 | 427/17697 | 2.45E-13 | 2.45E-11 | 1.64E-11 |
| MF | GO:0008236 | serine-type peptidase activity | 17/166 | 182/17697 | 1.33E-12 | 1.11E-10 | 7.42E-11 |
| MF | GO:0017171 | serine hydrolase activity | 17/166 | 186/17697 | 1.90E-12 | 1.36E-10 | 9.07E-11 |
| MF | GO:0004252 | serine-type endopeptidase activity | 16/166 | 160/17697 | 2.19E-12 | 1.37E-10 | 9.16E-11 |
| MF | GO:0019903 | protein phosphatase binding | 15/166 | 140/17697 | 4.01E-12 | 2.23E-10 | 1.49E-10 |
| MF | GO:0004713 | protein tyrosine kinase activity | 14/166 | 134/17697 | 3.03E-11 | 1.52E-09 | 1.02E-09 |
Table 7. KEGG enrichment analysis of potentially relevant targets of AS-IV anti-SA-AKI.
| Ontology | ID | Description | GeneRatio | BgRatio | p value | FDR | q value |
| KEGG | hsa05418 | Fluid shear stress and atherosclerosis | 24/150 | 139/8076 | 3.87E-17 | 9.95E-15 | 4.57E-15 |
| KEGG | hsa05205 | Proteoglycans in cancer | 25/150 | 205/8076 | 3.74E-14 | 4.81E-12 | 2.21E-12 |
| KEGG | hsa05215 | Prostate cancer | 18/150 | 97/8076 | 1.19E-13 | 9.19E-12 | 4.22E-12 |
| KEGG | hsa01522 | Endocrine resistance | 18/150 | 98/8076 | 1.43E-13 | 9.19E-12 | 4.22E-12 |
| KEGG | hsa05133 | Pertussis | 16/150 | 76/8076 | 3.73E-13 | 1.92E-11 | 8.79E-12 |
| KEGG | hsa05135 | Yersinia infection | 20/150 | 137/8076 | 5.46E-13 | 2.34E-11 | 1.07E-11 |
| KEGG | hsa01521 | EGFR tyrosine kinase inhibitor resistance | 16/150 | 79/8076 | 7.08E-13 | 2.60E-11 | 1.19E-11 |
| KEGG | hsa04657 | IL-17 signaling pathway | 17/150 | 94/8076 | 9.25E-13 | 2.97E-11 | 1.36E-11 |
| KEGG | hsa04933 | AGE-RAGE signaling pathway in diabetic complications | 17/150 | 100/8076 | 2.66E-12 | 7.59E-11 | 3.48E-11 |
| KEGG | hsa04625 | C-type lectin receptor signaling pathway | 17/150 | 104/8076 | 5.15E-12 | 1.32E-10 | 6.08E-11 |
| KEGG | hsa04659 | Th17 cell differentiation | 17/150 | 107/8076 | 8.30E-12 | 1.94E-10 | 8.90E-11 |
| KEGG | hsa05161 | Hepatitis B | 20/150 | 162/8076 | 1.33E-11 | 2.86E-10 | 1.31E-10 |
| KEGG | hsa04917 | Prolactin signaling pathway | 14/150 | 70/8076 | 2.50E-11 | 4.94E-10 | 2.26E-10 |
| KEGG | hsa04014 | Ras signaling pathway | 23/150 | 232/8076 | 3.42E-11 | 6.29E-10 | 2.88E-10 |
| KEGG | hsa05142 | Chagas disease | 16/150 | 102/8076 | 4.33E-11 | 7.41E-10 | 3.40E-10 |
| KEGG | hsa04660 | T cell receptor signaling pathway | 16/150 | 104/8076 | 5.87E-11 | 9.42E-10 | 4.32E-10 |
| KEGG | hsa04931 | Insulin resistance | 16/150 | 108/8076 | 1.06E-10 | 1.60E-09 | 7.32E-10 |
| KEGG | hsa05163 | Human cytomegalovirus infection | 22/150 | 225/8076 | 1.26E-10 | 1.70E-09 | 7.82E-10 |
| KEGG | hsa04010 | MAPK signaling pathway | 25/150 | 294/8076 | 1.26E-10 | 1.70E-09 | 7.82E-10 |
| KEGG | hsa05145 | Toxoplasmosis | 16/150 | 112/8076 | 1.85E-10 | 2.38E-09 | 1.09E-09 |
| KEGG | hsa04151 | PI3K-Akt signaling pathway | 27/150 | 354/8076 | 2.50E-10 | 3.06E-09 | 1.41E-09 |
| KEGG | hsa05167 | Kaposi sarcoma-associated herpesvirus infection | 20/150 | 193/8076 | 3.33E-10 | 3.90E-09 | 1.79E-09 |
| KEGG | hsa05130 | Pathogenic Escherichia coli infection | 20/150 | 197/8076 | 4.82E-10 | 5.39E-09 | 2.47E-09 |
| KEGG | hsa05152 | Tuberculosis | 19/150 | 180/8076 | 6.98E-10 | 7.27E-09 | 3.34E-09 |
| KEGG | hsa05131 | Shigellosis | 22/150 | 246/8076 | 7.07E-10 | 7.27E-09 | 3.34E-09 |
| KEGG | hsa04066 | HIF-1 signaling pathway | 15/150 | 109/8076 | 1.21E-09 | 1.20E-08 | 5.50E-09 |
| KEGG | hsa04668 | TNF signaling pathway | 15/150 | 112/8076 | 1.79E-09 | 1.70E-08 | 7.81E-09 |
| KEGG | hsa04062 | Chemokine signaling pathway | 19/150 | 192/8076 | 2.10E-09 | 1.93E-08 | 8.84E-09 |
| KEGG | hsa05219 | Bladder cancer | 10/150 | 41/8076 | 2.48E-09 | 2.19E-08 | 1.01E-08 |
| KEGG | hsa04722 | Neurotrophin signaling pathway | 15/150 | 119/8076 | 4.22E-09 | 3.61E-08 | 1.66E-08 |
| KEGG | hsa05120 | Epithelial cell signaling in Helicobacter pylori infection | 12/150 | 70/8076 | 4.57E-09 | 3.79E-08 | 1.74E-08 |
| KEGG | hsa05162 | Measles | 16/150 | 139/8076 | 4.80E-09 | 3.85E-08 | 1.77E-08 |
| KEGG | hsa05210 | Colorectal cancer | 13/150 | 86/8076 | 5.04E-09 | 3.93E-08 | 1.80E-08 |
| KEGG | hsa05235 | PD-L1 expression and PD-1 checkpoint pathway in cancer | 13/150 | 89/8076 | 7.76E-09 | 5.87E-08 | 2.69E-08 |
| KEGG | hsa05212 | Pancreatic cancer | 12/150 | 76/8076 | 1.21E-08 | 8.89E-08 | 4.08E-08 |
| KEGG | hsa04926 | Relaxin signaling pathway | 15/150 | 129/8076 | 1.30E-08 | 9.28E-08 | 4.26E-08 |
| KEGG | hsa04210 | Apoptosis | 15/150 | 136/8076 | 2.69E-08 | 1.87E-07 | 8.56E-08 |
| KEGG | hsa05132 | Salmonella infection | 20/150 | 249/8076 | 2.86E-08 | 1.93E-07 | 8.86E-08 |
| KEGG | hsa04217 | Necroptosis | 16/150 | 159/8076 | 3.37E-08 | 2.22E-07 | 1.02E-07 |
Astragalus and AS-IV enhance the PI3K/AKT pathway in both in vivo and in vitro
Gene enrichment analysis revealed that the PI3K/AKT pathway was enriched for the most corresponding target genes, indicating that PI3K/AKT is an important intracellular pathway, and a previous study confirmed that PI3K/AKT plays an important role in SA-AKI [15, 16]. Western Blot analysis revealed that PI3K and AKT phosphorylation was inhibited in the apical kidney tissues of the mice model when compared to the sham-operated group and that PI3K and AKT phosphorylation was significantly increased in the astragalus and AS-IV administration group when compared to the CLP group in a dose-dependent manner (Figure 5A, 5B and Supplementary Figure 3A, 3B). This confirms the hypothesis in the preceding section on network pharmacology that Astragalus and AS-IV are involved in the regulation of the PI3K/AKT pathway. Additionally, immunohistochemistry was used to determine the phosphorylation levels of AKT in the kidney tissues. The results indicated that AKT phosphorylation was inhibited in the apoptotic kidney tissues of the mice model when compared to the sham-operated group, but was significantly elevated in the Astragalus and AS-IV group compared to the CLP group (Figure 5C and 5D). Furthermore, Western Blot analysis revealed that LPS suppressed PI3K and AKT phosphorylation relative to the normal group, but AS-IV significantly elevated PI3K and AKT phosphorylation compared to the LPS group in a dose-dependent manner (Figure 5E and Supplementary Figure 3C). To further establish the involvement of PI3K/AKT, we used the PI3K inhibitor IC-87114 (5 μM) in HK-2 cells followed by incubation with LPS and AS-IV (20 μM) and found that phosphorylation was inhibited (Figure 5F and Supplementary Figure 3D). Additionally, the MTT assay confirmed that inhibiting the PI3K/AKT pathway decreased the cell survival rate induced by AS-IV (Figure 5G), corroborating our network pharmacology prediction that AS-IV was involved in regulating the PI3K/AKT pathway.
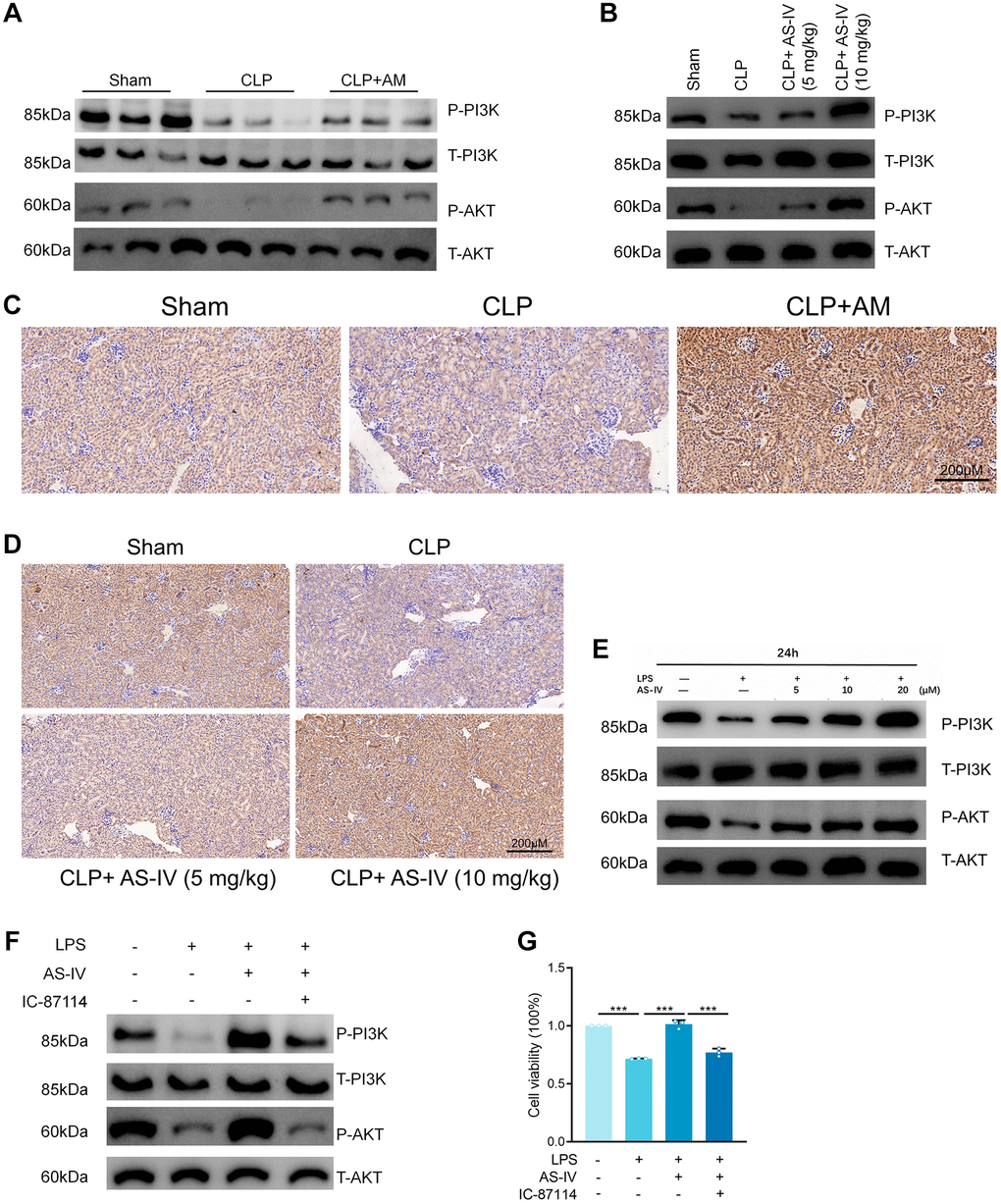
Figure 5. Astragalus and AS-IV enhanced PI3K/AKT pathway in vivo and vitro. (A and B) Expression level of Phospho-PI3K, PI3K, Phospho-AKT and AKT in kidney sections collected at 24 hours after CLP surgery following treatment with (250 mg/kg) Astragalus membranaceus or low (5 mg/kg) or high dosage (10 mg/kg) AS-IV. (C and D) Representative images showing Phospho-AKT immunohistochemical staining of mice kidney tissues. (E) Expression level of Phospho-PI3K, PI3K, Phospho-AKT and AKT in HK-2 cells incubated with LPS (1 μg/ml) plus AS-IV at the concentrations of 5, 10, 20 μM for 6 hours. (F) Protein expression of Phospho-PI3K, PI3K, Phospho-AKT and AKT in HK-2 cells incubated with LPS (1 μg/ml) plus AS-IV (20 μM) or PI3K inhibitor IC-87114 (5 μM) for 6 hours. (G) MTT results for AS-IV (20 μM) or PI3K inhibitor IC-87114 (5 μM) treatment on HK-2 cells incubated with LPS (1 μg/ml) for 6 hours. Data are presented as the mean ± SEM (n = 3 per group, *P < 0.05, **P < 0.01, ***P < 0.001).
Discussion
Chinese medicine is an important treasure of traditional Chinese medicine and is the material basis for the treatment of diseases in Chinese medicine. However, because of its complex composition and diverse effects, it cannot be precisely located in the body. Modern medical research has discovered that new drug development, is not only time-consuming and costly but also requires the application of network pharmacology and bioinformatics to significantly reduce the difficulties in new drug development. New drugs with significant efficacy against septic acute kidney injury and minimal adverse effects and cytotoxicity are urgently needed, and some monomers derived from Chinese herbal medicines are better choices than the herbs themselves because they can be administered in smaller doses, and have better-expected effects [12, 17].
Astragalus membranaceus is a widely used Chinese herb known for its ability to tonify Qi and invigorate blood. In this web-based pharmacology study, a total of 22 preferred compounds with good oral bioavailability and drug-like coefficients were identified using the ADME parameters OB (>30%) and DL (>0.18) reference values. Additionally, a review of the literature revealed that astragaloside analogues, the active components of Astragalus membranaceus, protect against sepsis-induced by cecum ligation perforation by inhibiting the inflammatory response and upregulation of the protein C pathway and that astragaloside has a protective effect on the kidney. Therefore, the present network pharmacology investigation included saponin components such as AS-I, AS-II, and AS-IV [18]. By establishing a compound-target network, 17 preferred Astragalus compounds against SA-AKI were identified based on network topology parameters. These compounds were then combined with a literature search, and we found that isorhamnetin [19], kaempferol [20], formononetin [21], Mairin [22], Jaranol [23], and Calycosin [24] had been reported in the literature in sepsis or acute kidney injury. Calycosins are flavonoids that have been shown to improve the pathological parameters associated with LPS-induced acute kidney injury, including lowering blood creatinine and urea nitrogen levels and reducing the expression of inflammatory factors, possibly by acting on the TLR4/NF-κB pathway to protect renal function and maintain normal kidney status [25]. Isorhamnetin, another flavonoid, has been investigated as a prospective option for bacterial infectious sepsis [19]. Kaempferol has been shown to protect septic mice against CLP-induced acute lung injury by inhibiting oxidative stress, iNOS, and ICAM-1 pathways [26]. Formononetin protects against cisplatin-induced AKI by activating the PPARα/Nrf2/HO 1/NQO1 pathway, promoting renal tubular cell proliferation, and inhibiting apoptosis following cisplatin-induced AKI [27]. Therefore, nine components were evaluated for their ability to induce septic acute kidney injury in HK-2 cells using LPS in vitro. Our findings indicated that that formononetin, AS-I and AS-IV, isorhamnetin, and Calycosin all exerted protective effects against LPS-induced cytotoxicity in HK-2 cells at 50 μM dosage, with AS-IV exerting the most prominent protective effect. Therefore, we investigated the mechanism of action of AS-IV against septic acute kidney injury both in vitro and in vivo.
AS-IV is the main active component of Astragalus and is frequently used in Chinese medicine as a quantitative and qualitative indicator of Astragalus. In recent years, various studies and reports have been published on the pharmacological effects of AS-IV in vitro and in vivo, including anti-oxidative stress, anti-inflammation, anti-apoptosis, and anti-fibrosis [18]. Numerous in vivo and in vitro studies have demonstrated that AS-IV has a high level of biological activity and can protect cells by inhibiting the production of epithelial-mesenchymal transition-related proteins, attenuating oxidative stress damage, inhibiting the release of inflammatory factors and other mechanisms [28]. AS-IV has been shown in previous studies to protect the nervous system and inhibit tumor disease progression by inhibiting endoplasmic reticulum stress [29] and to effectively reduce endoplasmic reticulum stress-induced renal tubular epithelial cell injury, podocyte autophagy, and endothelial cell inflammatory response, as well as to improve diabetic nephropathy injury [30]. In the present study, we found that LPS exposure significantly decreased HK-2 cell viability. AS-IV therapies, on the other hand, dose-dependently improved the viability of LPS-induced HK-2 cells in the model group. Similar to in vitro studies, in septic rats, AS-IV therapies markedly decreased CLP-induced tubular damage and improved tubular pathology. PI3K is an important inositol and phosphatidylinositol kinase. Three subtypes of PI3K have been identified: class I, class II, and class III, with class I being the most extensively investigated. PI3K-IA is a receptor-mediated dimeric protein with dual protein kinase and lipid-like kinase activity [31], while PI3K-IB is a receptor-mediated and delivered G protein-coupled receptor [32]. Protein kinase B (protein kinase B, PKB/AKT) is a serine/threonine kinase that regulates a variety of cellular biological activities, including cell cycle, proliferation, and apoptosis [33]. It also plays a key role in the PI3K/AKT pathway [34]. The PI3K/AKT signaling pathway is closely associated with the inflammatory response. Inhibiting this pathway significantly decreases the expression of inflammatory vesicles containing the Nod-like receptor protein 3 (NLRP3), which in turn significantly decreases the release of the downstream inflammatory factor IL-1β [35]. Lysosomes respond to their damaged organelles and macromolecules in response to hypoxia, infection, or starvation. The PI3K/AKT pathway is an important signaling pathway involved in this process [36]. Wullschleger et al. discovered a strong correlation between the PI3K/AKT pathway and inflammation [37]. Inhibition of the PI3K/AKT pathway resulted in a significant decrease in the release of the inflammatory factor NLRP3. Inhibition of the PI3K/AKT pathway decreased NLRP3 expression, which resulted in a significant decrease in IL-1β release. While the current study did not establish a direct and irreversible relationship between PI3K/AKT and sepsis-associated AKI, further exploration of its pathogenesis and prevention offers new possibilities for early diagnosis and treatment of the disease. Evidence suggests that AS-IV may have cardioprotective effects via altering the PI3K/AKT pathway and activating autophagic flow [38]. AS-IV has been demonstrated to improve cardiac function and decrease myocardial hypertrophy in several in vivo studies by decreasing oxidative stress and activating calpain-1, inhibiting the TBK1/PI3K/AKT signaling pathway, and several other additional mechanisms. Additionally, AS-IV can exert angiogenic and cardioprotective effects on the myocardium following myocardial infarction by activating the PTEN/PI3K/AKT signaling pathway and modulating the PI3K/AKT/GSK-3β pathway to alleviate the I/R effects caused by ligation of the left anterior descending coronary artery in rat hearts [39]. This study demonstrated that Astragalus and AS-IV treatment attenuated renal tubular epithelial cell injury in CLP mice and LPS-induced HK-2 cells, perhaps via a mechanism involving PI3K/AKT activation. However, this study only demonstrated the protective effect of LPS-induced tubular injury in HK-2 cells and CLP mice, although the function and mechanism of other pathogenic factors such as hyperglycemia and hyperlipidemia, on renal tubular epithelial cells need to be investigated further.
In summary, our findings demonstrate for the first time that Astragalus membranaceus and AS- IV protect against sepsis-induced AKI in renal tubular epithelial cells by enhancing the PI3K/AKT pathway. This study provides evidence that Astragalus membranaceus and AS- IV may be a significant therapeutic approach in the treatment of sepsis-induced AKI.
Author Contributions
Jia-Long Tang: performed experiments. Li-Chao Zhang: designed the study. Jia-Long Tang and Meng Xin: collected data. Jia-Long Tang: analyzed data. Jia-Long Tang and Meng Xin: wrote the paper.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
This study was supported by grants from the National Natural Science Foundation of China (No. 81973551), Scientific and Technologic Innovation Action of Shanghai Municipal Science and Technology Commission (No. 18401932700), the Health Commission of Shanghai Municipality (ZY(2021-2023)-0203-04) and the Chinese Major Scientific and Technological Special Project for Major New Drug Creation (2018X09201008-002-071).
Editorial Note
This corresponding author has a verified history of publications using a personal email address for correspondence
References
- 1. Singer M, Deutschman CS, Seymour CW, Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche JD, Coopersmith CM, Hotchkiss RS, Levy MM, Marshall JC, et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA. 2016; 315:801–10. https://doi.org/10.1001/jama.2016.0287 [PubMed]
- 2. Cohen J, Vincent JL, Adhikari NK, Machado FR, Angus DC, Calandra T, Jaton K, Giulieri S, Delaloye J, Opal S, Tracey K, van der Poll T, Pelfrene E. Sepsis: a roadmap for future research. Lancet Infect Dis. 2015; 15:581–614. https://doi.org/10.1016/S1473-3099(15)70112-X [PubMed]
- 3. Bellomo R, Kellum JA, Ronco C, Wald R, Martensson J, Maiden M, Bagshaw SM, Glassford NJ, Lankadeva Y, Vaara ST, Schneider A. Acute kidney injury in sepsis. Intensive Care Med. 2017; 43:816–28. https://doi.org/10.1007/s00134-017-4755-7 [PubMed]
- 4. Sun J, Zhang J, Tian J, Virzì GM, Digvijay K, Cueto L, Yin Y, Rosner MH, Ronco C. Mitochondria in Sepsis-Induced AKI. J Am Soc Nephrol. 2019; 30:1151–61. https://doi.org/10.1681/ASN.2018111126 [PubMed]
- 5. Gomez H, Ince C, De Backer D, Pickkers P, Payen D, Hotchkiss J, Kellum JA. A unified theory of sepsis-induced acute kidney injury: inflammation, microcirculatory dysfunction, bioenergetics, and the tubular cell adaptation to injury. Shock. 2014; 41:3–11. https://doi.org/10.1097/SHK.0000000000000052 [PubMed]
- 6. Li X, Qu L, Dong Y, Han L, Liu E, Fang S, Zhang Y, Wang T. A review of recent research progress on the astragalus genus. Molecules. 2014; 19:18850–80. https://doi.org/10.3390/molecules191118850 [PubMed]
- 7. Xia YG, Yu SM, Liang J, Yang BY, Kuang HX. Chemical fingerprinting techniques for the differentiation of polysaccharides from genus Astragalus. J Pharm Biomed Anal. 2020; 178:112898. https://doi.org/10.1016/j.jpba.2019.112898 [PubMed]
- 8. Liu F, Sun L, You G, Liu H, Ren X, Wang M. Effects of Astragalus polysaccharide on the solubility and stability of 15 flavonoids. Int J Biol Macromol. 2020; 143:873–80. https://doi.org/10.1016/j.ijbiomac.2019.09.148 [PubMed]
- 9. Shkondrov A, Krasteva I, Bucar F, Kunert O, Kondeva-Burdina M, Ionkova I. A new tetracyclic saponin from Astragalus glycyphyllos L. and its neuroprotective and hMAO-B inhibiting activity. Nat Prod Res. 2020; 34:511–7. https://doi.org/10.1080/14786419.2018.1491040 [PubMed]
- 10. Ren JH, Xue HQ, Liu Y, Wei YM, Adelson D, Chen XJ, Zhou JH, Yu JD. [Purification and biochemical function of Astragalus membtanaceus pathogenesis-related protein 10]. Zhongguo Zhong Yao Za Zhi. 2018; 43:3662–7. https://doi.org/10.19540/j.cnki.cjcmm.20180702.003 [PubMed]
- 11. Wang J, Wong YK, Liao F. What has traditional Chinese medicine delivered for modern medicine? Expert Rev Mol Med. 2018; 20:e4. https://doi.org/10.1017/erm.2018.3 [PubMed]
- 12. Hopkins AL. Network pharmacology: the next paradigm in drug discovery. Nat Chem Biol. 2008; 4:682–90. https://doi.org/10.1038/nchembio.118 [PubMed]
- 13. Zhang R, Zhu X, Bai H, Ning K. Network Pharmacology Databases for Traditional Chinese Medicine: Review and Assessment. Front Pharmacol. 2019; 10:123. https://doi.org/10.3389/fphar.2019.00123 [PubMed]
- 14. Rittirsch D, Huber-Lang MS, Flierl MA, Ward PA. Immunodesign of experimental sepsis by cecal ligation and puncture. Nat Protoc. 2009; 4:31–6. https://doi.org/10.1038/nprot.2008.214 [PubMed]
- 15. Gómez H, Kellum JA. Sepsis-induced acute kidney injury. Curr Opin Crit Care. 2016; 22:546–53. https://doi.org/10.1097/MCC.0000000000000356 [PubMed]
- 16. Peerapornratana S, Manrique-Caballero CL, Gómez H, Kellum JA. Acute kidney injury from sepsis: current concepts, epidemiology, pathophysiology, prevention and treatment. Kidney Int. 2019; 96:1083–99. https://doi.org/10.1016/j.kint.2019.05.026 [PubMed]
- 17. Luo TT, Lu Y, Yan SK, Xiao X, Rong XL, Guo J. Network Pharmacology in Research of Chinese Medicine Formula: Methodology, Application and Prospective. Chin J Integr Med. 2020; 26:72–80. https://doi.org/10.1007/s11655-019-3064-0 [PubMed]
- 18. Zhang J, Wu C, Gao L, Du G, Qin X. Astragaloside IV derived from Astragalus membranaceus: A research review on the pharmacological effects. Adv Pharmacol. 2020; 87:89–112. https://doi.org/10.1016/bs.apha.2019.08.002 [PubMed]
- 19. Chauhan AK, Kim J, Lee Y, Balasubramanian PK, Kim Y. Isorhamnetin Has Potential for the Treatment of Escherichia coli-Induced Sepsis. Molecules. 2019; 24:3984. https://doi.org/10.3390/molecules24213984 [PubMed]
- 20. Fernández-Del-Río L, Soubeyrand E, Basset GJ, Clarke CF. Metabolism of the Flavonol Kaempferol in Kidney Cells Liberates the B-ring to Enter Coenzyme Q Biosynthesis. Molecules. 2020; 25:2955. https://doi.org/10.3390/molecules25132955 [PubMed]
- 21. Aladaileh SH, Hussein OE, Abukhalil MH, Saghir SAM, Bin-Jumah M, Alfwuaires MA, Germoush MO, Almaiman AA, Mahmoud AM. Formononetin Upregulates Nrf2/HO-1 Signaling and Prevents Oxidative Stress, Inflammation, and Kidney Injury in Methotrexate-Induced Rats. Antioxidants (Basel). 2019; 8:430. https://doi.org/10.3390/antiox8100430 [PubMed]
- 22. Fan R, Hu PC, Wang Y, Lin HY, Su K, Feng XS, Wei L, Yang F. Betulinic acid protects mice from cadmium chloride-induced toxicity by inhibiting cadmium-induced apoptosis in kidney and liver. Toxicol Lett. 2018; 299:56–66. https://doi.org/10.1016/j.toxlet.2018.09.003 [PubMed]
- 23. Kim TW, Kim YJ, Seo CS, Kim HT, Park SR, Lee MY, Jung JY. Elsholtzia ciliata (Thunb.) Hylander attenuates renal inflammation and interstitial fibrosis via regulation of TGF-ß and Smad3 expression on unilateral ureteral obstruction rat model. Phytomedicine. 2016; 23:331–9. https://doi.org/10.1016/j.phymed.2016.01.013 [PubMed]
- 24. Ding X, Lv J, Luan J, Zhang J. Calycosin may Alleviate Ang II-Induced Pro-proliferative Effects on Glomerular Mesangial Cells via Partially Inhibiting Autophagy and ERK Signaling Pathway. Biol Pharm Bull. 2020; 43:1893–8. https://doi.org/10.1248/bpb.b20-00520 [PubMed]
- 25. Zhang YY, Tan RZ, Zhang XQ, Yu Y, Yu C. Calycosin Ameliorates Diabetes-Induced Renal Inflammation via the NF-κB Pathway In Vitro and In Vivo. Med Sci Monit. 2019; 25:1671–8. https://doi.org/10.12659/MSM.915242 [PubMed]
- 26. Harasstani OA, Tham CL, Israf DA. Kaempferol and Chrysin Synergies to Improve Septic Mice Survival. Molecules. 2017; 22:92. https://doi.org/10.3390/molecules22010092 [PubMed]
- 27. Hao Y, Miao J, Liu W, Peng L, Chen Y, Zhong Q. Formononetin protects against cisplatin-induced acute kidney injury through activation of the PPARα/Nrf2/HO-1/NQO1 pathway. Int J Mol Med. 2021; 47:511–22. https://doi.org/10.3892/ijmm.2020.4805 [PubMed]
- 28. Wang E, Wang L, Ding R, Zhai M, Ge R, Zhou P, Wang T, Fang H, Wang J, Huang J. Astragaloside IV acts through multi-scale mechanisms to effectively reduce diabetic nephropathy. Pharmacol Res. 2020; 157:104831. https://doi.org/10.1016/j.phrs.2020.104831 [PubMed]
- 29. Ge B, Li SL, Li FR. Astragaloside-IV regulates endoplasmic reticulum stress-mediated neuronal apoptosis in a murine model of Parkinson's disease via the lincRNA-p21/CHOP pathway. Exp Mol Pathol. 2020; 115:104478. https://doi.org/10.1016/j.yexmp.2020.104478 [PubMed]
- 30. Lai ST, Wang Y, Peng F. Astragaloside IV sensitizes non-small cell lung cancer cells to cisplatin by suppressing endoplasmic reticulum stress and autophagy. J Thorac Dis. 2020; 12:3715–24. https://doi.org/10.21037/jtd-20-2098 [PubMed]
- 31. Xu YP, Han F, Tan J. Edaravone protects the retina against ischemia/reperfusion-induced oxidative injury through the PI3K/Akt/Nrf2 pathway. Mol Med Rep. 2017; 16:9210–6. https://doi.org/10.3892/mmr.2017.7739 [PubMed]
- 32. Ma XM, Blenis J. Molecular mechanisms of mTOR-mediated translational control. Nat Rev Mol Cell Biol. 2009; 10:307–18. https://doi.org/10.1038/nrm2672 [PubMed]
- 33. Wang H, Wang Y, Gao H, Wang B, Dou L, Li Y. Piperlongumine induces apoptosis and autophagy in leukemic cells through targeting the PI3K/Akt/mTOR and p38 signaling pathways. Oncol Lett. 2018; 15:1423–8. https://doi.org/10.3892/ol.2017.7498 [PubMed]
- 34. Yang S, Wang Y, Gao H, Wang B. MicroRNA-30a-3p overexpression improves sepsis-induced cell apoptosis in vitro and in vivo via the PTEN/PI3K/AKT signaling pathway. Exp Ther Med. 2018; 15:2081–7. https://doi.org/10.3892/etm.2017.5644 [PubMed]
- 35. Qiu Y, Huang X, Huang L, Tang L, Jiang J, Chen L, Li S. 5-HT(1A) receptor antagonist improves behavior performance of delirium rats through inhibiting PI3K/Akt/mTOR activation-induced NLRP3 activity. IUBMB Life. 2016; 68:311–9. https://doi.org/10.1002/iub.1491 [PubMed]
- 36. Margaria JP, Campa CC, De Santis MC, Hirsch E, Franco I. The PI3K/Akt/mTOR pathway in polycystic kidney disease: A complex interaction with polycystins and primary cilium. Cell Signal. 2020; 66:109468. https://doi.org/10.1016/j.cellsig.2019.109468 [PubMed]
- 37. Edwards LA, Thiessen B, Dragowska WH, Daynard T, Bally MB, Dedhar S. Inhibition of ILK in PTEN-mutant human glioblastomas inhibits PKB/Akt activation, induces apoptosis, and delays tumor growth. Oncogene. 2005; 24:3596–605. https://doi.org/10.1038/sj.onc.1208427 [PubMed]
- 38. Cheng S, Zhang X, Feng Q, Chen J, Shen L, Yu P, Yang L, Chen D, Zhang H, Sun W, Chen X. Astragaloside IV exerts angiogenesis and cardioprotection after myocardial infarction via regulating PTEN/PI3K/Akt signaling pathway. Life Sci. 2019; 227:82–93. https://doi.org/10.1016/j.lfs.2019.04.040 [PubMed]
- 39. Xia ML, Xie XH, Ding JH, Du RH, Hu G. Astragaloside IV inhibits astrocyte senescence: implication in Parkinson's disease. J Neuroinflammation. 2020; 17:105. https://doi.org/10.1186/s12974-020-01791-8 [PubMed]