



Introduction
In the U.S. ~1.2 million people are living with human immunodeficiency virus type-1 (HIV-1) with men accounting for the majority of cases (~75%; [1]). Although combined antiretroviral therapy (cART) has reduced HIV-associated morbidity and mortality [2–4], patients continue to experience neurological and neuropsychiatric symptoms, collectively called “neuroHIV” [5–8]. This is likely due in part to efflux of some cART regimen from the central nervous system (CNS), high variance in brain accumulation of cART, and non-uniform distribution throughout the brain, all resulting in a lack of linear HIV RNA reduction in the CNS [9–11]. HIV is widely recognized as a disease-driver of aging as age-related disorders present prematurely and/or with greater severity in HIV-infected populations [12–15]. Actions of HIV in the CNS may contribute to premature aging partly by promoting secondary hypogonadism (i.e., hypothalamic and/or pituitary hormonal dysregulation) which is observed in 16% to 25% of young adult or middle-aged HIV-infected men [15–19]. In support, HIV+ men transition sooner to andropause [18, 19], characterized by lower total and/or free testosterone (T) seen in patients aged 20–39 years old [16–18, 20], greater circulating sex hormone-binding globulin [16, 17], and greater estradiol (E2)-to-T ratios [18] versus healthy age-matched men [21–23]. In the post-cART era, neurotoxic viral proteins persist within the CNS and contribute to mild and moderate forms of neuroHIV [24, 25], yet their contributions to neuroendocrine dysfunction and/or age-related neuroHIV remain unclear and understudied.
One HIV protein secreted from CNS reservoirs (predominantly microglia) that has been well-characterized is the trans-activator of transcription (Tat) [26–28]. Tat is present in the cerebrospinal fluid of cART-treated patients and even when patients achieve complete viral suppression peripherally, reservoirs in the brain continue to secrete Tat [24, 25, 29, 30]. Furthermore, anti-Tat antibodies are associated with reduced cognitive deficits among people living with HIV (PLWH) [31]. These data support the potential role of Tat in the pathophysiology of HIV-associated neurological disorders [24, 25, 29, 30]. Given its persistence in the CNS, Tat may contribute to neuroendocrine dysfunction observed in PLWH, partly due to its well-characterized mitotoxic effects. Tat promotes mitochondrial bioenergetic dysregulation, depolarization, and mitophagy, all of which are known to instigate and exacerbate cellular dysfunction in advanced age [32–37]. Tat-induced mitochondrial dysfunction could also negatively impact neuroendocrine function as mitochondria are the rate-limiting organelle for all steroidogenesis. Indeed, Tat alters lipid bioavailability, limits necessary steroid substrates including cholesterol [38, 39], and promotes the accumulation of ceramides which are inhibitors of steroid-synthesizing enzymes [40]. Using transgenic mice that conditionally express Tat protein, we have observed disruption of the neuroendocrine system, including the hypothalamic-pituitary-adrenal (HPA) and -gonadal (HPG) axes in young adult mice as well as HPG dysregulation in middle-aged female mice [36, 41, 42]. Tat-mediated alterations in steroid hormones may contribute to neurotoxicity. In support, exogenous treatment with T, E2, progesterone (P4), and 5α-pregnan-3α-ol-20-one (a.k.a., allopregnanolone or alloP) ameliorate Tat-mediated neurotoxicity in cell culture [42–45] and exogenous administration of P4 or alloP to Tat-transgenic mice attenuates some neuroHIV-like symptomatology [36, 45, 46]. These steroids, particularly alloP, are found to reduce neurodegeneration in several neurological disease states including Alzheimer’s disease [47, 48]. As such, gonadal hormones may be neuroprotective in middle-aged PLWH. However, the functional effects of Tat on the neuroendocrine milieu of aging males is unknown.
Herein, we investigate the combined effects of aging and HIV-1 Tat expression on the development of neuroHIV-like sequelae in young adult (6–8 months) and middle-aged (11–13 months) male mice to determine whether Tat precipitates age-related dysfunction. We hypothesized that conditional Tat expression in transgenic male mice [Tat(+)] would accelerate the development of (i.e., catalyze earlier presentation) or accentuate (i.e., increase magnitude) age-related affective, cognitive, neuromuscular, and neuropathic pain symptomatology compared to age-matched controls [Tat(–)] (Figure 1). We further expected that Tat would alter the production of circulating and central steroids (T, E2, P4, corticosterone, or alloP) concurrent with behavioral impairment.
Figure 1. Time-course of behavioral testing. In week 1, Tat protein expression was induced in young adult (6–8 mos. old) and middle-aged (11–13 mos. old) male mice via doxycycline administration (30 mg/kg, i.p. for 5 d) followed by two days of doxycycline washout. In week 2, mice were assessed for affective-like behavior in the open field (OF) and elevated plus-maze (EPM). In weeks 3 and 4, spatial memory performance was assessed in a radial arm water maze (RAWM). In week 5, neuromuscular function was assessed by grip strength and nociceptive/analgesic responding was assessed via the electronic-Von Frey and thermal probe tests.
Results
Aging and Tat expression altered steroidogenesis in male mice
Aging and Tat expression altered circulating gonadal and adrenal steroid hormones. Older age and Tat expression interacted to influence circulating T [F(1,24) = 5.46, p < 0.05; Figure 2A]. Middle-aged Tat(−) mice exhibited greater T than did their respective young adult Tat(−) controls (p = 0.05; Figure 2A, see §) or their respective middle-aged Tat(+) counterparts (p = 0.007; Figure 2A, see ^). Among young adult mice, circulating E2 was greater in Tat(+) males compared to all other groups [F(1,23) = 34.98, p < 0.05; Figure 2B; see †]. Moreover, the calculated E2:T ratio (a measure of aromatization) was greatest in young adult Tat(+) mice compared to any other group [F(1,20) = 10.39, p < 0.05], [E2:T ratio: Tat(−)Young Adult = 0 ± 0 ng/mL, Tat(+)Young Adult = 0.06 ± 0.01 ng/mL, Tat(−)Middle-aged = 0.02 ± 0.02 ng/mL, Tat(+)Middle-aged = 0.01 ± 0.01 ng/mL]. E2 and the E2:T ratio were notably increased with middle-age; however, this did not reach statistical significance (p = 0.09, n.s.). Circulating P4 also interacted with age and Tat expression [F(1,27) = 13.16, p < 0.05; Figure 2C]. Middle-aged Tat(−) mice demonstrated the greatest circulating P4 concentrations of all groups (p < 0.0001 – 0.04; Figure 2C, see ^) and young adult Tat(+) mice demonstrated greater P4 than their age-matched Tat(−) controls (p = 0.02; Figure 2C, see *. Corticosterone was highest among middle-aged Tat(+) mice versus all other groups [F(1,23) = 4.69, p < 0.05; Figure 2D, see †].
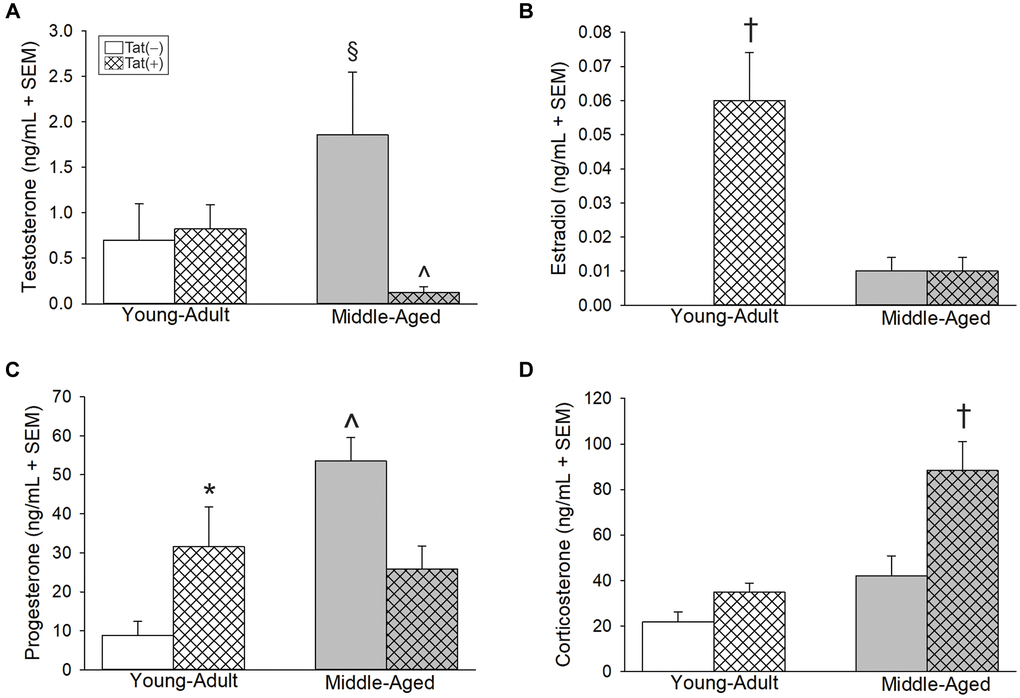
Figure 2. Circulating steroids (ng/mL) among young adult and middle-aged HIV-1 Tat-transgenic male mice [Tat(+)] or their non-Tat-expressing age-matched counterparts [Tat(−)]. (A) Testosterone, (B) estradiol, (C) progesterone, and (D) corticosterone detected in serum. §indicated group differs from young adult Tat(−) controls. ^significant interaction wherein indicated group differs from respective Tat(−) controls. †indicated group differs from all other groups. *main effect for Tat(+) mice to differ from Tat(−) controls, (two-way ANOVA, p < 0.05).
Older age and Tat expression also interacted to influence brain alloP content. We previously observed that Tat expression increased alloP protein content in the whole brain of young male mice, without changes in serum [36]. However, regional analysis of Tat-provoked alloP in the brain has not been conducted heretofore. In hippocampus, older age [F(1,22) = 6.21, p < 0.05; Figure 3A, see †] or Tat expression [F(1,22) = 14.12, p < 0.05; Figure 3A, see *] increased alloP content. In midbrain, age and Tat expression interacted to increase alloP content in middle-aged Tat(+) mice to a greater extent than that seen in all other groups [F(1,24) = 13.71, p < 0.05, Figure 3B, see ‡]. No significant differences were observed in alloP content within the frontal cortex (Figure 3C).
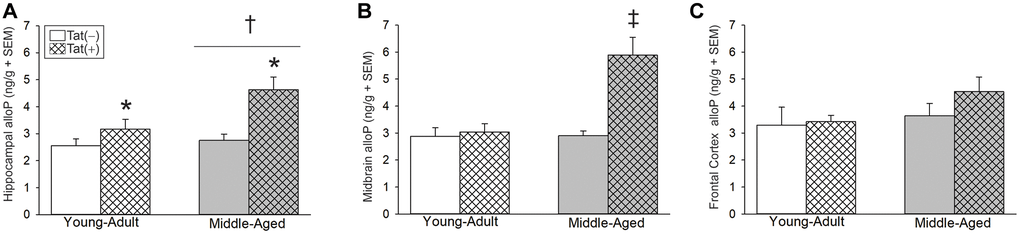
Figure 3. Central steroid allopregnanolone (ng/g) among young adult and middle-aged HIV-1 Tat-transgenic male mice [Tat(+)] or their non-Tat-expressing age-matched counterparts [Tat(−)]. (A) Hippocampal, (B) midbrain, and (C) frontal cortex allopregnanolone (alloP). *main effect for Tat(−) mice to differ from Tat(−) controls. †main effect for middle-aged mice to differ from young adults. ‡indicated group differs from all other groups, (two-way ANOVA, p < 0.05).
Aging or Tat increased anxiety-like behavior among male mice
Aging and Tat expression promoted anxiety-like behavior. In the open field, there were main effects for age [F(1,35) = 9.14, p < 0.05; Figure 4A, see †] and Tat expression [F(1,35) = 5.29, p < 0.05; Figure 4A, see *], both of which increased anxiety-like behavior and reduced time spent in the center of the open field compared to young adult or Tat(−) counterparts, respectively. As well, there was an interaction wherein age or Tat genotype increased anxiety-like behavior [F(1,36) = 5.86, p < 0.05; Figure 4B]. Both middle-aged mice and Tat(+) young adult mice made fewer entries into the brightly-lit center of the open field compared to young adult Tat(−) controls (p = 0.001–0.027; Figure 4B, see ^). In the elevated plus-maze, we observed a main effect for Tat [F(1,38) = 13.13, p < 0.05; Figure 4C, see *] to increase the latency to enter the open arms compared to Tat(−) controls. As well, Tat(+) mice demonstrated a notable decrease in the proportion of open arm time compared to Tat(−) mice; albeit, this did not reach statistical significance (p = 0.09; Figure 4D). Notably, motor differences were detected for the distance (cm) [F(1,36) = 16.82, p < 0.05; Table 1] and velocity (cm/s) [F(1,36) = 16.85, p < 0.05; Table 1] traveled in the open field. Middle-aged or Tat(+) mice demonstrated motor deficits and travelled shorter distances (p = 0.0001–0.042), and at lower velocities (p < 0.0001–0.041), than did young adult Tat(−) mice. Within the middle-aged group, Tat(−) males travelled shorter distances (p = 0.03) and at slower velocities than did Tat(+) mice (p = 0.03). No differences were observed in the total arm entries made in the elevated plus-maze (Table 1).
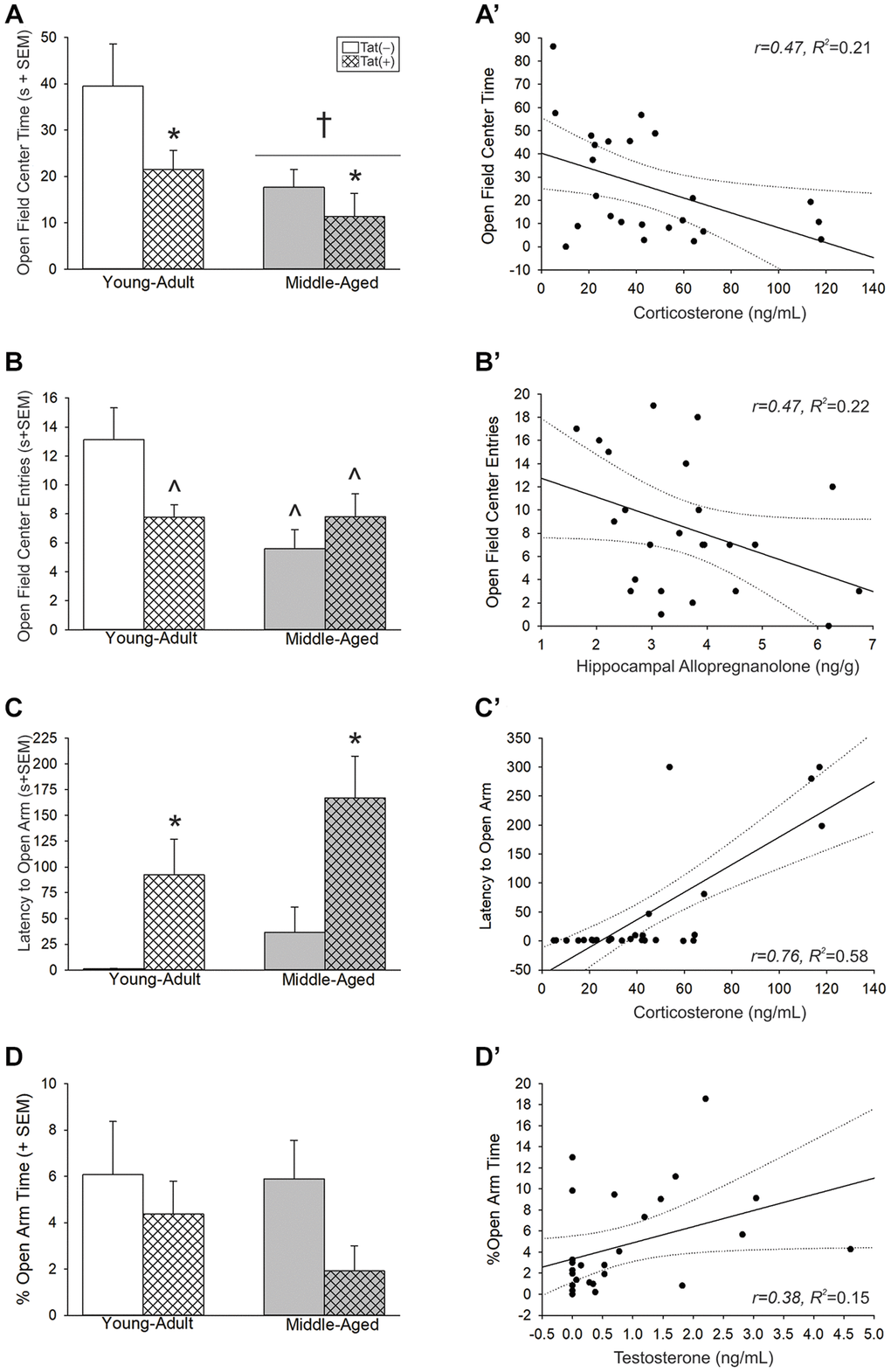
Figure 4. (A–D) Anxiety-like behavior in the open field and elevated plus-maze and (A’–D’) simple linear regressions between circulating and central steroid hormones among young adult and middle-aged HIV-1 Tat-transgenic male mice [Tat(+)] or their non-Tat-expressing age-matched counterparts [Tat(−)]. (A) Time (s) spent in the brightly-lit center of an open field. (B) Numbers of entries made into the center of an open field. (C) Latency (s) to enter the open arms of an elevated plus-maze. (D) The proportional time spent in the open arms of an elevated plus-maze. Regressions between (A’) circulating corticosterone and center time, (B’) hippocampal allopregnanolone and center entries, (C’) circulating corticosterone and latency to enter open arms, and (D’) circulating T and proportional open arm time. *main effect for Tat genotype wherein Tat(+) mice to differ from Tat(−) mice. †main effect for middle-aged mice to differ from young adult mice. ^significant interaction wherein indicated group differs from young adult Tat(−) controls. Regression lines (solid) are depicted with 95% confidence intervals (dotted), (two-way ANOVA, p < 0.05).
Table 1. Motor behavior (open field and elevated plus maze), raw open arm time (elevated plus maze), grip strength, and proportional body weight and peripheral organ wet weights (mean ± SEM) among HIV-1 Tat transgenic [Tat(+)] young adult and middle-aged male mice and age-matched non-Tat-expressing controls [Tat(−)].
| Young Adult | Middle-Aged | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tat(−) n = 9 | Tat(+) n = 9–12 | Tat(−) n = 12 | Tat(+) n = 10 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Open Field | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Distance (cm) | 1580.40 ± 164.05 | 1127.45 ± 147.46^ | 907.72 ± 99^ | 1242.34 ± 79.64^* | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Velocity (cm/s) | 5.28 ± 0.55 | 3.76 ± 0.49^ | 3.03 ± 0.33^ | 4.14 ± 0.27^ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Elevated Plus Maze | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Open Arm Time (s) | 13.10 ± 4.96 | 11.72 ± 3.75 | 15.34 ± 4.41 | 3.86 ± 2.10 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Total Arm Entries | 29.78 ± 3.15 | 25.58 ± 3.53 | 25.00 ± 2.66 | 31.23 ± 4.80 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Grip Strength | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Forelimbs alone | 0.50 ± 0.02 | 0.35 ± 0.01^ | 0.35 ± 0.02^ | 0.28 ± 0.01^§ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Forelimbs and hindlimbs | 0.62 ± 0.04 | 0.50 ± 0.01* | 0.54 ± 0.01*# | 0.41 ± 0.02*# | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Body and Organ Weight | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Body Weight (g) | 31.76 ± 0.96 | 32.20 ± 1.29 | 41.92 ± 1.98‡ | 30.50 ± 0.90 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Brain weight (%) | 1.40 ± 0.06 | 1.40 ± 0.08 | 1.14 ± 0.12 | 1.49 ± 0.10 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Heart weight (%) | 0.50 ± 0.02 | 0.71 ± 0.07^ | 0.64 ± 0.07 | 1.49 ± 0.10† | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liver weight (%) | 4.48 ± 0.17 | 4.32 ± 0.33 | 5.14 ± 0.62 | 4.92 ± 0.04 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Spleen weight (%) | 0.28 ± 0.02 | 0.22 ± 0.04 | 0.26 ± 0.03 | 0.62 ± 0.12‡ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| *indicates a main effect for Tat(+) mice to differ from Tat(−) mice, irrespective of age. #indicates a main effect for middle-aged mice to differ from young adults, irrespective of genotype. ^indicates a difference from young adult Tat(−) controls. †indicates a difference from young adult Tat(+) mice. §indicates a difference from young adult Tat(+) mice and middle-aged Tat(−) controls.‡indicates a difference from all other groups, p < 0.05. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Affective-like behavior correlated with steroid fluctuations. In the open field, circulating corticosterone (Figure 4A’; Table 2) and frontal cortex alloP (Table 2) were negatively correlated with the time spent in the center of the open field. Similarly, circulating P4 (Table 2) and hippocampal alloP (Figure 4B’; Table 2) were negatively correlated with open field central entries. P4 also negatively correlated with the velocity travelled (Table 2). In the elevated plus-maze, circulating corticosterone (Figure 4C’; Table 2) and alloP, within all brain regions examined (i.e., hippocampus, midbrain, frontal cortex; Table 2), positively correlated with the latency to enter the open arms. T levels were also positively correlated with the proportion of time spent on the open arms (Figure 4D’; Table 2).
Table 2. Simple linear regressions between circulating or central steroid hormones and behavioral assessments among HIV-1 Tat transgenic [Tat(+)] young adult and middle-aged male mice and age-matched non-Tat-expressing controls [Tat(−)].
| Dependent Variable | Independent Variable | β | t-value (df) | r | R2 | F-value (df) | p-value |
| Open field | |||||||
| Center time | Corticosterone | 40.34 | 2.41 (23) | −0.46 | 0.21 | 5.82 (1,23) | 0.025 |
| Frontal cortex alloP | 57.21 | 3.29 (23) | −0.57 | 0.33 | 10.80 (1,23) | 0.003 | |
| Center entries | Progesterone | 11.31 | 2.65 (26) | 0.47 | 0.22 | 7.00 (1,26) | 0.01 |
| Hippocampal alloP | 15.64 | 2.34 (20) | −0.47 | 0.22 | 5.50 (1,20) | 0.031 | |
| Frontal cortex alloP | 13.84 | 2.06 (22) | −0.41 | 0.17 | 4.23 (1,22) | 0.05 | |
| Velocity | Progesterone | 4.54 | 2.24 (27) | −0.40 | 0.16 | 5.01 (1,27) | 0.03 |
| Distance | Progesterone | 1360.40 | 2.25 (27) | −0.40 | 0.16 | 5.05 (1,27) | 0.03 |
| Elevated plus maze | |||||||
| Latency to open arm | Corticosterone | −58.35 | 5.72 (24) | −0.76 | 0.58 | 32.74 (1,25) | <0.0001 |
| Hippocampal alloP | −126.87 | 3.89 (24) | 0.62 | 0.39 | 15.12 (1,25) | 0.001 | |
| Midbrain alloP | −63.86 | 2.87 (26) | 0.49 | 0.24 | 8.24 (1,27) | 0.01 | |
| Frontal cortex alloP | −66.32 | 2.47 (26) | 0.44 | 0.20 | 6.12 (1,27) | 0.02 | |
| % Open arm time | Testosterone | 3.35 | 2.10 (25) | 0.38 | 0.15 | 4.29 (1,26) | 0.05 |
| RAWM | |||||||
| Reference memory error | Estradiol | 5.10 | 2.56 (24) | 0.29 | 0.08 | 6.53 (1,74) | 0.013 |
| Velocity | Progesterone | 7.52 | 4.72 (28) | 0.46 | 0.21 | 22.23 (1,86) | <0.0001 |
| Estradiol | 8.12 | 3.74 (24) | 0.40 | 0.16 | 14.00 (1,74) | 0.0004 | |
| Hippocampal alloP | 6.30 | 3.26 (23) | 0.35 | 0.12 | 10.65 (1,77) | 0.002 | |
| Testosterone | 8.26 | 3.10 (25) | 0.33 | 0.11 | 9.61 (1,80) | 0.003 | |
| Corticosterone | 8.01 | 2.47 (24) | 0.28 | 0.08 | 6.12 (1,74) | 0.02 | |
| Distance | Testosterone | 365.30 | 2.11 (24) | 0.23 | 0.05 | 4.44 (1,80) | 0.04 |
| Grip strength | |||||||
| % forelimbs alone | Progesterone | 1.33 | 2.04 (26) | −0.37 | 0.14 | 4.17 (1,27) | 0.05 |
| % Forelimbs and hindlimbs together | Corticosterone | 1.82 | −2.03 (21) | −0.41 | 0.17 | 4.14 (1,22) | 0.05 |
| Nociceptive-like behavior | |||||||
| Thermal hyperalgesia | Hippocampal alloP | 42.89 | 2.54 (23) | 0.47 | 0.22 | 6.45 (1,24) | 0.02 |
| Body and Organs Weight | |||||||
| Brain weight (%) | Testosterone | 1.43 | 2.23 (24) | 0.41 | 0.17 | 5.00 (1,25) | 0.04 |
Aging or Tat reduced cognitive performance in male mice
Older age and Tat expression interacted to influence latencies to escape a radial arm water maze [F(2,76) = 3.42, p < 0.05; Table 3]. Young adult, Tat(−) mice demonstrated longer latencies to escape than young adult Tat(+) mice (day 2; p = 0.004) and middle-aged groups (day 3; p < 0.001–0.005). Age and Tat expression also interacted to alter the proportional performance improvements from day 1 [F(1,35) = 9.46, p < 0.05; Figure 5A]. Unexpectedly, young adult Tat(−) mice demonstrated less improvement than did young adult Tat(+) (p = 0.022; Figure 5A; see ^) and middle-aged Tat(−) mice (p = 0.018; Figure 5A; see ^). Middle-aged Tat(+) mice demonstrated less improvement than did their age-matched, control counterparts (p = 0.004; Figure 5A; see ‡) or young adult Tat(+) mice (p = 0.006; Figure 5A; see ‡). When assessing the total errors made, age and Tat expression interacted [F(1,76) = 17.95, p < 0.05; Figure 5B]. Among young adult mice, Tat expression increased the number of errors made (p < 0.0001; Figure 5B, see ^). Older age also increased the number of errors made among Tat(−) controls, compared to their young adult counterparts (p < 0.0001; Figure 5B, see ^). Contrary to expectation, middle-aged Tat(+) mice made fewer errors than did their respective Tat(−) controls or their young adult Tat(+) counterparts (p = 0.001–0.006; Figure 5B; see ‡). This may have been related to differences in swim speed. Among the middle-aged group, Tat(+) mice swam faster than their age-matched, or young-adult, Tat(−) counterparts only on day 1 (p < 0.0001–0.002; Figure 5C, see *). Middle-aged mice were faster than their young adult controls on days 2 (p < 0.0001–0.03; Figure 5C, see ||) and 3 (p < 0.0001; Figure 5C, see ^). Fewer errors made by middle-aged Tat(+) mice may be a consequence of increased anxiety and faster latencies to escape (Table 3). In support, despite observing no difference in the distance travelled (Table 3), a significant interaction between age, Tat expression, and day of testing was observed for swim speed [F(2,78) = 5.82, p < 0.05; Figure 5C]. Among young adults, Tat(+) mice swam faster than their Tat(−) counterparts over all three days (p < 0.0001–0.0003; Figure 5C, see ^).
Table 3. Latency to escape (s) and distance travelled (cm) in a radial arm water maze among HIV-1 Tat transgenic [Tat(+)] young adult and middle-aged male mice and age-matched non-Tat-expressing controls [Tat(−)].
| Latency to Escape (s) | Young adult | Middle-aged | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tat(−) n = 8–9 | Tat(+) n = 12 | Tat(−) n = 12 | Tat(+) n = 10 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Day 1 | 47 ± 6 | 36 ± 3 | 49 ± 6 | 37 ± 4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Day 2 | 49 ± 8 | 27 ± 2* | 35 ± 6 | 37 ± 4 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Day 3 | 50 ± 5 | 25 ± 3^ | 31 ± 6^ | 33 ± 4^ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Distance (cm) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Day 1 | 372 ± 31 | 386 ± 29 | 410 ± 30 | 387 ± 30 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Day 2 | 318 ± 32 | 320 ± 27 | 334 ± 37 | 380 ± 31 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Day 3 | 349 ± 48 | 271 ± 23 | 307 ± 29 | 298 ± 31 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| *indicates an effect for young adult Tat(+) mice to differ from young adult Tat(−) mice. ^indicates a difference from young adult Tat(−) controls. p < 0.05. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
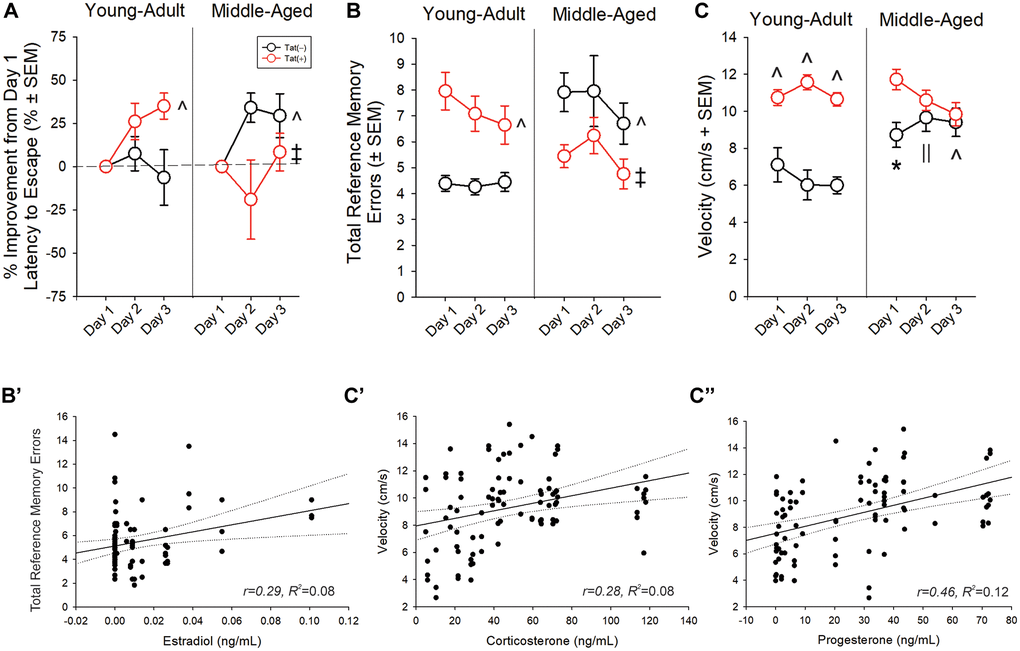
Figure 5. (A, B) Spatial memory performance and (C) swim speed in a radial arm water maze and (B’, C’’) simple linear regressions for circulating and central steroid hormones among young adult and middle-aged HIV-1 Tat-transgenic male mice [Tat(+)] or their non-Tat-expressing age-matched counterparts [Tat(−)]. (A) Proportion of mice that exhibited improvement from day one performance (latency to escape). (B) Total frequency of errors. (C) Swim speed (cm/s). Simple linear regressions between (B’) circulating estradiol and frequency of errors, (C’) circulating corticosterone and swim speed, and (C’’) progesterone and swim speed. *main effect for Tat(+) mice to differ from Tat(−) mice. ^interaction effect wherein indicated group differs from young adult Tat(−) controls. ||indicates middle-aged differs from young-adult groups. ‡indicated middle-aged Tat(+) groups differs from middle-aged Tat(−) and young adult Tat(+) mice. Regression lines (solid) are depicted with 95% confidence intervals (dotted), (repeated measure ANOVA, p < 0.05).
Radial arm water maze performance correlated with circulating and central steroid levels. Circulating E2 was positively correlated with the total reference memory errors (Figure 5B’; Table 2). Corticosterone (Figure 5C’; Table 2), P4 (Figure 5C’’; Table 2), E2 (Table 2), hippocampal alloP (Table 2), and T (Table 2) were positively correlated with swim speed. T was negatively correlated with the total distance travelled (Table 2).
Aging and Tat reduced neuromuscular strength among male mice
Advanced age and Tat interacted to reduce grip strength when assessed either by forelimbs alone (normalized to body weight) [F(1,37) = 13.68, p < 0.05; Figure 6A] or by forelimbs and hindlimbs together (normalized to body weight) [F(1,36) = 7.00, p < 0.05; Figure 6B]. In either measure, middle-aged or Tat(+) mice were weaker than young Tat(−) controls (p < 0.0001–0.004, Figure 6A, 6B, see ^). As well, middle-aged Tat(−) mice were weaker than young adult Tat(+) mice (p = 0.02–0.03, Figure 6A, 6B, see †). Grip strength assessment without body weight normalization yielded comparable effects (Table 1). There was an interaction between age and Tat exposure when assessing forelimbs alone [F(1,37) = 9.24, p < 0.05; Table 1] such that all groups were weaker than young adult Tat(−) controls (p < 0.0001). As well, there was a “stair-step” effect wherein middle-aged Tat(+) mice were weaker than their own middle-aged Tat(−) controls (p = 0.002) or young adult Tat(+) mice (p = 0.002). There were also main effects for age [F(1,37) = 12.14, p < 0.05; Table 1] or Tat exposure [F(1,37) = 25.64, p < 0.05; Table 1] to reduce combined forelimb and hindlimb grip strength.
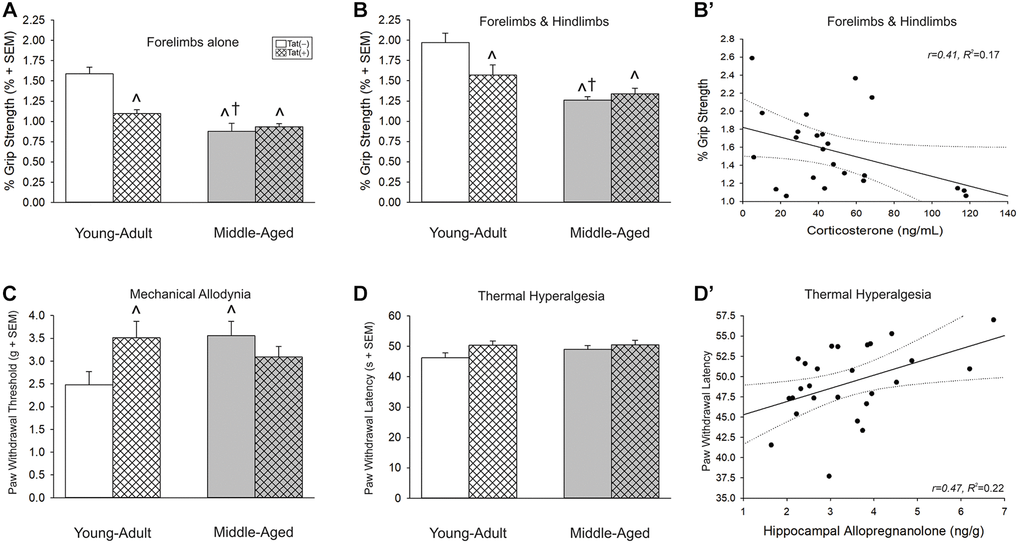
Figure 6. (A, B) Grip strength, (C) mechanical allodynia, and (D) thermal hyperalgesia and (B’ and D’) simple linear regression for circulating and central steroid hormones among young adult and middle-aged HIV-1 Tat-transgenic male mice [Tat(+)] or their non-Tat-expressing age-matched counterparts [Tat(−)]. Grip strength threshold for (A) forelimbs alone or (B) forelimbs and hindlimbs together. (C) Paw-withdrawal threshold (g) in an electronic Von Frey test. (D) Paw-withdrawal latency (s) in a thermal probe test. Simple linear regressions between (B’) circulating corticosterone and forelimbs and hindlimbs together or (D’) hippocampal allopregnanolone and paw-withdrawal latency. *main effect for Tat(+) mice to differ from Tat(−) mice. †main effect for middle-aged mice to differ from young adults. ^significant interaction wherein indicated group differs from young adult Tat(−) controls. Regression lines (solid) are depicted with 95% confidence intervals (dotted), (two-way ANOVA, p ≤ 0.05).
Grip strength was also correlated with circulating steroids. Corticosterone was negatively correlated with neuromuscular function as assessed by the forelimbs and hindlimbs together (Figure 6B’; Table 2) and P4 negatively correlated with neuromuscular function as assessed by the forelimbs alone (Table 2).
Aging and Tat altered nociceptive tolerance in male mice
Older age and Tat expression interacted to influence the threshold for paw withdrawal in response to a mechanical stimulus [F(1,37) = 5.87, p < 0.05; Figure 6C]. Unexpectedly, middle-aged Tat(−) and young adult Tat(+) mice exhibited a greater mechanical threshold than did young adult Tat(−) controls (p = 0.02–0.03; Figure 6C, see ^). No differences were observed in thermal hyperalgesia among middle-aged or Tat-exposed mice (Figure 6D); however, hippocampal alloP was positively correlated with the latency to paw withdrawal from a thermal stimulus (Figure 6D’; Table 2).
Aging and Tat influenced body and peripheral organ weight
Aging and Tat expression interacted to influence body weight [F(1,37) = 17.00, p < 0.05; Table 1] such that middle-aged Tat(−) mice were heavier than all other groups (p = 0.0001–0.004). There was also an interaction effect between age and Tat to increase proportional heart wet weights [F(1,25) = 8.41, p < 0.05; Table 1] wherein young adult Tat(+) mice had greater proportional mass than their age-matched Tat(−) counterparts (p = 0.011) or middle-aged Tat(+) mice (p = 0.04). The proportional spleen wet weight [F(1,24) = 12.33, p < 0.05; Table 1] was greatest in middle-aged Tat(+) mice compared to all other groups (p = 0.0001–0.0003). We did not observe differences in the proportional weights of brain or liver (Table 1). However, circulating T was positively correlated with proportional brain wet weight (Table 2).
Discussion
HIV-infected individuals contend with an accelerated onset of age-related diseases and disorders [49–51]; however, the pathophysiology underlying accelerated aging is poorly understood. Findings from this study support the hypothesis that aging and HIV-1 Tat protein exert independent and interactive effects to induce neuroendocrine dysfunction which coincide with neuroHIV-like symptomatology in male mice. Consistent with observations in HIV+ patients [16–17, 20], Tat expression in mice reduced circulating T among middle-aged males. Moreover, HIV+ men (ages 18–70 y/o) experience increased E2: T ratios [18], and we find that Tat expression recapitulates this endophenotype in male mice. Tat induction also elevated circulating P4 in young adult mice, an endocrine profile observed in our middle-aged controls suggesting that Tat expression may accelerate the onset of endocrine dysregulation. These preclinical data support the notion that exposure to Tat is sufficient to induce changes in the HPG axis that are consistent with clinical phenotypes. We observed further changes in the HPA axis as the animals aged. Tat expression elevated circulating corticosterone and central alloP in the hippocampus and midbrain of middle-aged mice. These neuroendocrine responses correlated with Tat-induced behavioral deficits, confirming our prior findings of Tat-disrupted alloP neurosteroidogenesis [36] and HPA axis dysregulation [52] and revealing their long-term consequences in the development of age-related neurological comorbidities. In addition, Tat altered peripheral organs, inducing an increase in the proportion of heart wet-weight. In particular, middle-aged Tat(+) mice had the greatest heart wet-weight of any group. These data are consistent with premature heart remodeling to promote hypertrophy; heart disease is notably a leading cause of mortality among PLWH [53] and its incidence increases with age [54, 55]. Middle-aged Tat(+) mice also had greater spleen weights than other groups which could be associated with several age-related comorbidities including lymphoma. Future pathological studies may expand on these findings.
Mood and cognitive disorders are common among HIV+ patients [49, 56–59] and their prevalence increases with age [60] and with endocrine dysfunction [61, 62]. Irrespective of HIV status, age-related androgen decline is associated with a greater risk of developing dementia, Alzheimer’s disease [63, 64], and cognitive impairment in men [65]. In the pre-cART era, hypogonadal men had higher depression scores than eugonadal men [61]. Among individuals having progressed to AIDS, risk for depression was greater than in eugonadal HIV+ men [66]. While, endocrine disorders have declined in the post-cART era, secondary hypogonadism is still prevalent among HIV+ patients (16–25%) [18, 21, 67, 68]. Testosterone insufficiency is associated with depression/apathy, cognitive impairment, sexual dysfunction, fatigue, reduced muscle strength, and increased risk for HIV-associated lipodystrophy [19, 20]. Herein, we find that older age and/or Tat expression increase anxiety-like behavior in an open field and in an elevated plus-maze. In both tasks, Tat expression engendered an anxiety-like phenotype in young adults comparable to that observed in middle-aged mice, consistent with accelerated aging. Testosterone positively correlated with anti-anxiety-like behavior on the elevated plus maze supporting a role for androgens to improve mood. On the RAWM, Tat expression increased the frequency of errors made throughout the learning phase of the task, consistent with prior demonstrations of Tat-impaired spatial memory [41, 69–74]. These data extend prior findings to demonstrate that older age also increases errors, an effect that was positively correlated with greater circulating E2. The distance travelled to escape the RAWM was negatively correlated with circulating T, further supporting the benefits of a eugonadal state. Contrary to expectation, Tat-exposed middle-aged mice made fewer errors than their age-matched Tat(−) counterparts. This may reflect greater activation of the HPA axis in older Tat(+) mice. In support, middle-aged Tat(+) mice were faster than controls on day 1 of the RAWM and had greater corticosterone levels that also positively correlated with swim speed. This group also had the lowest circulating T and lower body weight than their non-Tat expressing middle-aged counterparts supportive of a reduced anabolic state. Consistent with the possibility of a generalized HPG/HPA activation in response to stress, circulating steroids and hippocampal alloP levels were positively correlated with RAWM swim speed. Thus, age and Tat effects on HPG/HPA axis function influenced ‘performative measures’ (i.e., open field and swim velocity) and these were associated with affective and cognitive dependent measures.
In the present work, an attenuated HPA axis was associated with improved neuromuscular function and hyperalgesia. Moreover, young adult Tat(+) mice demonstrated grip strength that was commensurate to that of middle-aged Tat(−) or Tat(+) mice suggestive of Tat’s capacity to accelerate age-related neuromuscular dysfunction. These observations correlated negatively with circulating corticosterone and P4. In HIV+ patients, neuromuscular deficits occur concurrent with disease progression and the onset of painful neuropathies [75]. As such, we also assessed mechanical antinociception and thermal hyperalgesia. We observed that aging or Tat expression increased pain-like thresholds among mice. While, this is contrary to the expectation of peripheral neuropathy, we previously observed similar effects in young adult male Tat(+) mice, which had greater antinociceptive thresholds compared to their Tat(−) counterparts [76]. This effect may involve peripheral sensory damage given evidence for intra-epidermal nerve fiber regression [77], which may impact sensation. In support, ~50% of HIV+ patients experience neuropathic pain and/or paresthesias, the latter of which are difficult to assess in mouse models [78]. We did not observe any thermal hyperalgesia, consistent with prior reports [41, 77], but hippocampal alloP was positively correlated with greater thermal thresholds. Several factors may contribute to neuropathic pain in the HIV+ population, including neurotoxic antiretroviral treatment (nucleoside reverse transcriptase inhibitors; albeit, used more sparingly in developed nations [79]), or metabolic syndrome [80], and age [81, 82]. As well, HIV-1 proteins beyond Tat, including the envelope glycoprotein 120 (gp120), likely contribute to neuropathic pain. For example, in cultured dorsal root ganglion neurons, gp120 exerts chemokine-like effects via increased neural excitation and substance P release [83]. Intradermal injection of gp120 induces allodynia [83] and reduces epidermal nerve fiber density [84]. Discrepancies in findings for Tat-induced mechanical allodynia may also involve sex differences. We previously found that middle-aged Tat(+) females have reduced mechanical pain thresholds compared to controls [41], in contrast to current findings in middle-aged Tat(+) males. Bagdas et al. (2020) observed similar effects in a cohort of age-matched young adult Tat-transgenic mice.
Beyond typical hormone replacement therapies, neurosteroid-based therapeutics may hold promise for numerous age-related diseases and disorders. In rodents, androgen replacement therapy including testosterone and its neuroactive metabolite, 5α-androstan-3α, 17β-diol (a.k.a. 3α−androstanediol), reduces anxiety- and depression-like behavior and improves neurocognitive performance [85–87]. Similarly, exogenous alloP improved hippocampal-dependent learning and memory among aged male mice including the triple-transgenic Alzheimer’s model [87, 88]. We have found that exogenous alloP attenuated affective dysfunction, mitotoxicity, and neurotoxicity associated with HIV-1 Tat [36, 45]. Together, these data support the notion that maintaining the endocrine milieu of the CNS promotes resilience to neurodegenerative insults, including those associated with older age. In support, we recently found that T declines with age in the medial prefrontal cortex and striatum of Tat-expressing transgenic mice, concurrent with decrements in excitatory and inhibitory neurotransmitters as well as endogenous antioxidants detected with magnetic resonance spectroscopy [89]. These findings are consistent with clinical observations of cognitive impairment [90] that occurred in parallel with cerebral metabolic disturbances including a reduction of N-acetyl aspartate and an increase of choline and myoinositol, markers of inflammation, in HIV+ patients [29, 90, 91]. Further, older age and HIV tended to exert additive effects at reducing subcortical gray matter in regions that included amygdala, caudate, and corpus callosum in one study [92] and altered β-amyloid deposition in another [93]. These findings are consistent with observations of accumulated hyperphosphorylated Tau in the hippocampus of HIV+ patients [94]. Others have found synergistic effects of older age and HIV on the exacerbation of verbal memory deficits [95]. Given the potential protective benefits of neurosteroids for neurodegenerative disease states ranging from Alzheimer’s [96] to neuroHIV, neurosteroids may serve as scaffolds for a new generation of age-based therapeutics.
The present study had some limitations. We measured total circulating steroid content but did not assess the free, bioavailable fraction. Given increases in sex hormone-binding globulin among HIV+ patients [16, 17, 67, 97, 98], future studies will discern the amount of bioavailable androgen. As well, we have recently found that middle-aged female mice also display HPG dysregulation that correlated with some behavioral deficits [41]. Together, these data support the assessment of exogenous steroid hormone treatment for potential benefits using aged models. While the present study did not include mice greater than 13 months of age, the current work highlights Tat’s capacity to precipitate several characteristic phenotypes of advanced age on its own; thus, suggesting that Tat accumulation can decrease health span within mice.
The current work provides potentially actionable information that could help to limit premature or accentuated aging in the context of HIV. The hypogonadism observed in clinical populations may reflect substantial neuroendocrine dysfunction that can be readily assessed in preclinical models. In support, the formation of neurosteroids are dysregulated in post-mortem HIV+ brains [99] and in the circulation of HIV+ patients [100]. The latter included the dehydroepiandrosterone sulfate-to-cortisol ratio, a biomarker for HPA dysregulation, which was associated with depressive symptomatology [100]. As well, T replacement improves mood disorders and lean body weight in HIV-infected men [66, 101, 102]. In conclusion, our data suggest that older age and Tat expression exert independent and interactive effects to worsen neuroendocrine, affective, cognitive, and neuromuscular comorbidities. Novel steroid replacement therapies may be useful adjunctive therapeutics to cART in the aging HIV+ population.
Materials and Methods
All experimental procedures were approved by the Institutional Animal Care and Use Committee at the University of Mississippi and conduced in accordance with the National Institutes of Health Guide for Care and Use of Laboratory Animals (NIH Publication No. 85–23) ethical guidelines.
Animals and housing
Young adult (6–8 months old; nTat(−) = 9, nTat(+) = 12) or middle-aged (11–13 months old; nTat(−) = 12, nTat(+) = 10) HIV-1 Tat-transgenic male mice were generated in the vivarium at the University of Mississippi (University, MS). These mice were founded on a C3H × C57BL/6J background and have been repeatedly back-crossed to C57BL/6J [103, 104]. In Tat(+) mice, the Tat1-86 protein is conditionally-expressed via administration of doxycycline, as described [103]. Tat(−) mice express the rtTA transcription factor necessary for tat expression, but lack the tat transgene [103]. Mice were group-housed (3–4 per cage) in a temperature- and humidity-controlled facility on a reversed 12:12-h light:dark cycle (lights off at 09:00 h). Food and water were available ad libitum. Animal housing rooms were specific pathogen free (parvovirus, helicobacter, etc.). Mouse ages were selected from prior work that associates human age with murine development, finding one year of human life equivalent to nine days in the moues during adulthood [105]. These ages are consistent with prior work comparing young adult and middle-aged mice [106–108].
HIV-1 Tat induction
To conditionally express the HIV-1 tat transgene (or not), Tat(+) mice and their Tat(−) counterparts received doxycycline (Dox) hyclate (30 mg/kg; Cayman Chemical, Ann Arbor, MI) intraperitoneally (i.p.) once daily for five consecutive days, followed by a 2-day wash-out period to rule out non-specific effects of Dox (Dox t1/2z = 5–6 h; [109]).
Behavioral assessment
Mice were tested over the course of four-weeks using several behavioral assessments in the following order: open field, elevated plus maze, radial arm water maze, grip strength, electronic Von Frey (eVF), and thermal probe. A 48-h break occurred between each behavioral test except for neuropathic-like pain assessments, eVF and thermal probe, which were conducted on the same day (Figure 1). Mice were handled daily before behavioral assessments, which were conducted ~1 h into their dark phase. Thirty min prior to testing, mice were transferred to a testing room and habituated to 70dB of white noise. Behavioral apparatus was cleaned with 70% ethanol between trials to avoid olfactory bias. All behavioral assessments were tracked and digitally encoded by EthoVision animal tracking software (Noldus, Leesburg, VA, USA).
Open field
The open field test was used to assess anxiety-like behavior and ataxia in rodents [46, 110]. Mice were placed in the center of the brightly-lit area of the open field apparatus (40 × 40 × 35 cm) and allowed to freely explore the apparatus for 5 min. Greater time spent, and more entries made into, the brightly-lit center area were used as indices of anti-anxiety-like behavior. Total distance moved (cm) and velocity (cm/s) were considered indices of locomotor activity [46, 110].
Elevated plus maze
The elevated plus maze was used to assess anxiety-like behavior [41, 111]. Briefly, the maze consisted of two cross-arms (two open and two enclosed arms; 61 × 5 cm ea.) that were elevated from the floor (37.5 cm). Mice were placed in the plus-maze center area facing the open arm and allowed to freely explore for 5 min. A shorter latency to enter the open arm and a greater proportional time spent on the open arms were considered indices of greater anti-anxiety-like behavior. The total number of entries made into arms was used as an index of locomotor activity [41].
Radial arm water maze
The radial arm water maze (RAWM) was used to assess learning and spatial memory [112, 113]. The maze consisted of eight arms (24 × 8 cm) attached to a central area (18 cm). Briefly, the maze was filled with room-temperature tap water (equilibrated overnight) and a hidden platform (6 × 8 cm) was located at the end of a predetermined arm (goal arm). Mice completed 6 trials daily for 3 days. In each trial, mice were placed in a randomized, predetermined (starting arm) and allowed to swim freely for 90 s to reach a hidden platform. When mice failed to locate the hidden platform, they were gently guided by the investigator to the platform and allowed to remain for 15 s. The starting arm was randomized for each trial. A shorter time to locate the platform and fewer total errors made were considered indices of greater spatial cognitive performance [112, 113]. To assess the proportional improvement from day 1, the following calculation was performed: 1 – (latency to escape on days 2 or 3/latency to escape on day 1) × 100 [41]. Entries into an incorrect arm were counted as errors. Total distance (cm) and velocity (cm/s) were used as indices of locomotor activity.
Grip strength
Grip Strength was used to assess neuromuscular function in rodents as previously described [41, 114]. Briefly, mice were suspended by the tail and allowed to grab a metal grid bar connected to a transducer that measured peak force (g). Mice were gently pulled by the base of the tail horizontally away from the bar until grip was lost. The strength of forelimbs, alone or in combination with hindlimbs, was measured. Each mouse was tested in five trials with a 1 min break between trials to prevent fatigue. The mean of five trials was used as the grip strength score. Grip strength values were normalized to body weight and considered as an index of neuromuscular function [41, 114].
Electronic Von Frey
The electronic Von Frey (eVF; Top Cat Metrology) test was used to measure mechanical allodynia as previously described [41, 115]. Mice were allowed to habituate for 15 min on an elevated wire mesh. A series of eVF probes was applied to the hind paw (at the middle plantar surface) with force gradually increasing until paw withdrawal occurred. Each mouse was assessed in eight trials alternating between right and left paws (4 for each side) with a 3-min inter-trial interval. The mean of 4 trials for each paw was calculated. A lower mechanical threshold indicated an increase in the allodynic response [41, 115, 116].
Thermal probe test
A thermal probe test was used to evaluate thermal hyperalgesia in rodents as previously described [41, 117]. A radiant heat stimulus was applied to the mid-plantar surface of the hind paw. The heat source was increased from room temperature to 60°C at a rate of 2.5°C/s. The test was stopped once a paw withdrawal occurred. Each mouse was assessed in 4 trials (2 for the right and 2 for the left paw) by alternating between right and left paws. Mice had 3–5 min breaks between trials. Paw withdrawal latencies were averaged and used as an index of hyperalgesia [41, 117].
Central and circulating steroid assay
Tissue collection
Two days following behavioral assessments, a subset of animals (young adult; nTat(−) = 9, nTat(+) = 8) or middle-aged; nTat(−) = 6, nTat(+) = 8) was sacrificed via cervical dislocation followed by rapid decapitation. Brain, heart, liver, and spleen were collected, weighed and stored at −80°C for later analysis. Blood was collected and centrifuged at 13,500 × g (4°C) for 20 min. Serum and all tissues were stored at −80°C. Remaining mice underwent perfusion for imaging to be reported elsewhere.
Enzyme-linked immunosorbent assay (ELISA)
Steroid extraction from serum was achieved via diethyl ether snap-freezing as previously described [42, 45]. Briefly, serum was transferred to glass borosilicate culture tubes and the total volume was measured. One mL of ice-cold anhydrous diethyl ether was added to all samples which then underwent snap-freeze for 30 s. Supernatant was collected and allowed to evaporate to dryness overnight in a fume hood. Using ELISA extraction buffer (Neogen Life Sciences, Lexington, KY, USA), samples were reconstituted to 5× their original volume. Circulating steroid hormones (E2, P4, T, and corticosterone) were analyzed using ELISA kits according to manufacture instructions (Neogen Life Sciences, Lexington, KY, USA) and previous methods [42, 45]. Steroids were reconstituted with the extraction buffer at 1:5, 1:25, 1:100, and 1:50 dilutions for E2, P4, T, and corticosterone, respectively. Hydrolysis of substrate was measured at 560 nm (optic density) using a CLARIOstar microplate reader (BMG Labtech Inc., Cary, NC, USA). Steroid hormone concentrations were calculated using a curve-fitting routine (4-parameter). The antibodies supplied reported a 100% cross-reactivity with the steroids of interest. The E2 antibody exerts low cross-reactivities (<1%) with other steroids. The P4 antibody has 2.5% and <2% cross-reactivities with deoxycorticosterone and other steroids, respectively. The T antibody exhibits 100% cross-reactivity with dihydrotestosterone, 16.12% cross-reactivity with T-glucuronide, and <1% cross-reactivity with other steroids. The corticosterone antibody exhibits cross-reactivities with deoxycorticosterone (38%), 6-hydrocortisone (19%), P4 (5.1%), and tetrahydrocortisone (2.7%), as reported by the manufacture (Neogen Life Sciences, Lexington, KY, USA). Standards ranged from (0.02–2 ng/ml), (0.4–10 ng/ml), (0.002–0.2 ng/ml), and (0.04–10 ng/ml) for E2, P4, T, and corticosterone, respectively.
Ultra performance liquid chromatography (UPLC) - mass spectrometry (MS)
Brain regions (hippocampus, midbrain, and frontal cortex) were grossly-dissected, wet weighed, and stored at −80°C until UPLC-MS/MS was conducted. Charcoal-stripped brain tissues derived from Tat(−) and Tat(+) mice were used for calibration and quality control. Samples were homogenized in 100 μl of PBS (pH 7.4) and steroid extraction was achieved via protein precipitation. Samples were precipitated with 100 μl of acetonitrile followed by vortexing for two min and centrifugation for ten min (14,000 rpm). Following centrifugation, supernatants were mixed with 50 μl of derivatizing solution (20 mg/mL 2-hydrazinopyridine prepared in 0.5% trifluoroacetic acid ethanol solution) and incubated for 1 h at 60°C. 20 μl of the internal standard solution (LLOQ: 0.25 ng/ml) was added and vortexed. For analysis, 2 μl of sample was injected onto the UPLC-MS/MS instrument [118].
Statistical analyses
Behavioral endpoints (open field, elevated plus-maze, grip strength, eVF and thermal probe) were analyzed via two-way analyses of variance (ANOVA) using age-group and Tat genotype as factors. Data obtained from radial arm water maze were analyzed via repeated measures ANOVA with testing day as the within-subjects factor and age-group and Tat genotype as the between-subjects factors. Main effects were delineated using Fisher’s Protected Least Significant Difference post hoc tests to determine group differences. Interactions were delineated via the assessment of simple main effects and main effect contrasts with alpha (0.05) controlled for family-wise error. Outliers were determined by Dixon’s test and were excluded. All data were considered significant when p ≤ 0.05.
Data and materials availability
Data are available upon request.
Author Contributions
JJP, NMA, and MJK conceptualized the project. ANQ, JJP, NMA, and MJK garnered funding for the project. NMA oversaw behavioral work conducted in the UM Neuropharmacology Core. ANQ, JJP, and NMA analyzed the data. ANQ and FM collected data. ANQ first drafted and revised the manuscript. All authors revised the manuscript and approved the final version.
Acknowledgments
We appreciate the technical support from the University of Mississippi Neuropharmacology and DMPK Cores.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Funding
This work was supported by funds from the University of Mississippi (Graduate Student Council award to ANQ) and the National Institutes of Health: R00 DA039791 (JJP), R01 DA052851 (JJP and NMA), R01 DA039044 (MJK), and an administrative supplement from award P30 GM122733 (pilot project to JJP and MJK).
References
- 1. Centers for Disease Control and Prevention. HIV Surveillance Report. 2018. [Updated 2020 May; accessed 2021 Mar 15]. http://www.cdc.gov/hiv/library/reports/hiv-surveillance.html.
- 2. Saylor D, Dickens AM, Sacktor N, Haughey N, Slusher B, Pletnikov M, Mankowski JL, Brown A, Volsky DJ, McArthur JC. HIV-associated neurocognitive disorder--pathogenesis and prospects for treatment. Nat Rev Neurol. 2016; 12:234–48. https://doi.org/10.1038/nrneurol.2016.27 [PubMed]
- 3. Thakur KT, Boubour A, Saylor D, Das M, Bearden DR, Birbeck GL. Global HIV neurology: a comprehensive review. AIDS. 2019; 33:163–84. https://doi.org/10.1097/QAD.0000000000001796 [PubMed]
- 4. Veenhuis RT, Clements JE, Gama L. HIV Eradication Strategies: Implications for the Central Nervous System. Curr HIV/AIDS Rep. 2019; 16:96–104. https://doi.org/10.1007/s11904-019-00428-7 [PubMed]
- 5. Bing EG, Burnam MA, Longshore D, Fleishman JA, Sherbourne CD, London AS, Turner BJ, Eggan F, Beckman R, Vitiello B, Morton SC, Orlando M, Bozzette SA, et al. Psychiatric disorders and drug use among human immunodeficiency virus-infected adults in the United States. Arch Gen Psychiatry. 2001; 58:721–8. https://doi.org/10.1001/archpsyc.58.8.721 [PubMed]
- 6. Clifford DB, Ances BM. HIV-associated neurocognitive disorder. Lancet Infect Dis. 2013; 13:976–86. https://doi.org/10.1016/S1473-3099(13)70269-X [PubMed]
- 7. Mann R, Sadosky A, Schaefer C, Baik R, Parsons B, Nieshoff E, Stacey BR, Tuchman M, Nalamachu S. Burden of HIV-Related Neuropathic Pain in the United States. J Int Assoc Provid AIDS Care. 2016; 15:114–25. https://doi.org/10.1177/2325957415592474 [PubMed]
- 8. Woods SP, Moore DJ, Weber E, Grant I. Cognitive neuropsychology of HIV-associated neurocognitive disorders. Neuropsychol Rev. 2009; 19:152–68. https://doi.org/10.1007/s11065-009-9102-5 [PubMed]
- 9. Ferrara M, Bumpus NN, Ma Q, Ellis RJ, Soontornniyomkij V, Fields JA, Bharti A, Achim CL, Moore DJ, Letendre SL. Antiretroviral drug concentrations in brain tissue of adult decedents. AIDS. 2020; 34:1907–14. https://doi.org/10.1097/QAD.0000000000002628 [PubMed]
- 10. Nath A. Eradication of human immunodeficiency virus from brain reservoirs. J Neurovirol. 2015; 21:227–34. https://doi.org/10.1007/s13365-014-0291-1 [PubMed]
- 11. Shaik N, Giri N, Pan G, Elmquist WF. P-glycoprotein-mediated active efflux of the anti-HIV1 nucleoside abacavir limits cellular accumulation and brain distribution. Drug Metab Dispos. 2007; 35:2076–85. https://doi.org/10.1124/dmd.107.017723 [PubMed]
- 12. Bhasin S, Singh AB, Javanbakht M. Neuroendocrine abnormalities associated with HIV infection. Endocrinol Metab Clin North Am. 2001; 30:749–64. https://doi.org/10.1016/s0889-8529(05)70210-3 [PubMed]
- 13. Hodes RJ, Sierra F, Austad SN, Epel E, Neigh GN, Erlandson KM, Schafer MJ, LeBrasseur NK, Wiley C, Campisi J, Sehl ME, Scalia R, Eguchi S, et al. Disease drivers of aging. Ann N Y Acad Sci. 2016; 1386:45–68. https://doi.org/10.1111/nyas.13299 [PubMed]
- 14. Pathai S, Bajillan H, Landay AL, High KP. Is HIV a model of accelerated or accentuated aging? J Gerontol A Biol Sci Med Sci. 2014; 69:833–42. https://doi.org/10.1093/gerona/glt168 [PubMed]
- 15. Tripathy SK, Agrawala RK, Baliarsinha AK. Endocrine alterations in HIV-infected patients. Indian J Endocrinol Metab. 2015; 19:143–7. https://doi.org/10.4103/2230-8210.146870 [PubMed]
- 16. Gomes AR, Souteiro P, Silva CG, Sousa-Pinto B, Almeida F, Sarmento A, Carvalho D, Freitas P. Prevalence of testosterone deficiency in HIV-infected men under antiretroviral therapy. BMC Infect Dis. 2016; 16:628. https://doi.org/10.1186/s12879-016-1892-5 [PubMed]
- 17. Lachâtre M, Pasquet A, Ajana F, Soudan B, Lion G, Bocket L, Cornavin P, Senneville E, Boufassa F, Chéret A. HIV and hypogonadism: a new challenge for young-aged and middle-aged men on effective antiretroviral therapy. AIDS. 2017; 31:451–3. https://doi.org/10.1097/QAD.0000000000001348 [PubMed]
- 18. Rochira V, Zirilli L, Orlando G, Santi D, Brigante G, Diazzi C, Carli F, Carani C, Guaraldi G. Premature decline of serum total testosterone in HIV-infected men in the HAART-era. PLoS One. 2011; 6:e28512. https://doi.org/10.1371/journal.pone.0028512 [PubMed]
- 19. Slama L, Jacobson LP, Li X, Palella FJ
Jr , Margolick JB, Kingsley LA, Wiley DJ, Pialoux G, Dobs AS, Brown TT, and Multicenter AIDS Cohort Study. Longitudinal Changes Over 10 Years in Free Testosterone Among HIV-Infected and HIV-Uninfected Men. J Acquir Immune Defic Syndr. 2016; 71:57–64. https://doi.org/10.1097/QAI.0000000000000821 [PubMed] - 20. Ashby J, Goldmeier D, Sadeghi-Nejad H. Hypogonadism in human immunodeficiency virus-positive men. Korean J Urol. 2014; 55:9–16. https://doi.org/10.4111/kju.2014.55.1.9 [PubMed]
- 21. Araujo AB, O'Donnell AB, Brambilla DJ, Simpson WB, Longcope C, Matsumoto AM, McKinlay JB. Prevalence and incidence of androgen deficiency in middle-aged and older men: estimates from the Massachusetts Male Aging Study. J Clin Endocrinol Metab. 2004; 89:5920–6. https://doi.org/10.1210/jc.2003-031719 [PubMed]
- 22. Araujo AB, Esche GR, Kupelian V, O'Donnell AB, Travison TG, Williams RE, Clark RV, McKinlay JB. Prevalence of symptomatic androgen deficiency in men. J Clin Endocrinol Metab. 2007; 92:4241–7. https://doi.org/10.1210/jc.2007-1245 [PubMed]
- 23. Feldman HA, Longcope C, Derby CA, Johannes CB, Araujo AB, Coviello AD, Bremner WJ, McKinlay JB. Age trends in the level of serum testosterone and other hormones in middle-aged men: longitudinal results from the Massachusetts male aging study. J Clin Endocrinol Metab. 2002; 87:589–98. https://doi.org/10.1210/jcem.87.2.8201 [PubMed]
- 24. Mediouni S, Darque A, Baillat G, Ravaux I, Dhiver C, Tissot-Dupont H, Mokhtari M, Moreau H, Tamalet C, Brunet C, Paul P, Dignat-George F, Stein A, et al. Antiretroviral therapy does not block the secretion of the human immunodeficiency virus tat protein. Infect Disord Drug Targets. 2012; 12:81–6. https://doi.org/10.2174/187152612798994939 [PubMed]
- 25. Henderson LJ, Johnson TP, Smith BR, Reoma LB, Santamaria UA, Bachani M, Demarino C, Barclay RA, Snow J, Sacktor N, Mcarthur J, Letendre S, Steiner J, et al. Presence of Tat and transactivation response element in spinal fluid despite antiretroviral therapy. AIDS. 2019 (Suppl 2); 33:S145–57. https://doi.org/10.1097/QAD.0000000000002268 [PubMed]
- 26. Ajasin D, Eugenin EA. HIV-1 Tat: Role in Bystander Toxicity. Front Cell Infect Microbiol. 2020; 10:61. https://doi.org/10.3389/fcimb.2020.00061 [PubMed]
- 27. King JE, Eugenin EA, Buckner CM, Berman JW. HIV tat and neurotoxicity. Microbes Infect. 2006; 8:1347–57. https://doi.org/10.1016/j.micinf.2005.11.014 [PubMed]
- 28. Yao H, Buch S. Rodent models of HAND and drug abuse: exogenous administration of viral protein(s) and cocaine. J Neuroimmune Pharmacol. 2012; 7:341–51. https://doi.org/10.1007/s11481-012-9355-2 [PubMed]
- 29. Cysique LA, Jugé L, Lennon MJ, Gates TM, Jones SP, Lovelace MD, Rae CD, Johnson TP, Nath A, Brew BJ. HIV brain latency as measured by CSF BcL11b relates to disrupted brain cellular energy in virally suppressed HIV infection. AIDS. 2019; 33:433–41. https://doi.org/10.1097/QAD.0000000000002076 [PubMed]
- 30. Johnson TP, Patel K, Johnson KR, Maric D, Calabresi PA, Hasbun R, Nath A. Induction of IL-17 and nonclassical T-cell activation by HIV-Tat protein. Proc Natl Acad Sci U S A. 2013; 110:13588–93. https://doi.org/10.1073/pnas.1308673110 [PubMed]
- 31. Bachani M, Sacktor N, McArthur JC, Nath A, Rumbaugh J. Detection of anti-tat antibodies in CSF of individuals with HIV-associated neurocognitive disorders. J Neurovirol. 2013; 19:82–8. https://doi.org/10.1007/s13365-012-0144-8 [PubMed]
- 32. Ferri KF, Jacotot E, Blanco J, Esté JA, Kroemer G. Mitochondrial control of cell death induced by HIV-1-encoded proteins. Ann N Y Acad Sci. 2000; 926:149–64. https://doi.org/10.1111/j.1749-6632.2000.tb05609.x [PubMed]
- 33. Fields JA, Ellis RJ. HIV in the cART era and the mitochondrial: immune interface in the CNS. Int Rev Neurobiol. 2019; 145:29–65. https://doi.org/10.1016/bs.irn.2019.04.003 [PubMed]
- 34. Fitting S, Knapp PE, Zou S, Marks WD, Bowers MS, Akbarali HI, Hauser KF. Interactive HIV-1 Tat and morphine-induced synaptodendritic injury is triggered through focal disruptions in Na+ influx, mitochondrial instability, and Ca2+ overload. J Neurosci. 2014; 34:12850–64. https://doi.org/10.1523/JNEUROSCI.5351-13.2014 [PubMed]
- 35. Norman JP, Perry SW, Kasischke KA, Volsky DJ, Gelbard HA. HIV-1 trans activator of transcription protein elicits mitochondrial hyperpolarization and respiratory deficit, with dysregulation of complex IV and nicotinamide adenine dinucleotide homeostasis in cortical neurons. J Immunol. 2007; 178:869–76. https://doi.org/10.4049/jimmunol.178.2.869 [PubMed]
- 36. Paris JJ, Liere P, Kim S, Mahdi F, Buchanan ME, Nass SR, Qrareya AN, Salahuddin MF, Pianos A, Fernandez N, Shariat-Madar Z, Knapp PE, Schumacher M, Hauser KF. Pregnane steroidogenesis is altered by HIV-1 Tat and morphine: Physiological allopregnanolone is protective against neurotoxic and psychomotor effects. Neurobiol Stress. 2020; 12:100211. https://doi.org/10.1016/j.ynstr.2020.100211 [PubMed]
- 37. Rozzi SJ, Avdoshina V, Fields JA, Trejo M, Ton HT, Ahern GP, Mocchetti I. Human Immunodeficiency Virus Promotes Mitochondrial Toxicity. Neurotox Res. 2017; 32:723–33. https://doi.org/10.1007/s12640-017-9776-z [PubMed]
- 38. Cotto B, Natarajaseenivasan K, Ferrero K, Wesley L, Sayre M, Langford D. Cocaine and HIV-1 Tat disrupt cholesterol homeostasis in astrocytes: Implications for HIV-associated neurocognitive disorders in cocaine user patients. Glia. 2018; 66:889–902. https://doi.org/10.1002/glia.23291 [PubMed]
- 39. Mohseni Ahooyi T, Shekarabi M, Torkzaban B, Langford TD, Burdo TH, Gordon J, Datta PK, Amini S, Khalili K. Dysregulation of Neuronal Cholesterol Homeostasis upon Exposure to HIV-1 Tat and Cocaine Revealed by RNA-Sequencing. Sci Rep. 2018; 8:16300. https://doi.org/10.1038/s41598-018-34539-9 [PubMed]
- 40. Dickens AM, Yoo SW, Chin AC, Xu J, Johnson TP, Trout AL, Hauser KF, Haughey NJ. Chronic low-level expression of HIV-1 Tat promotes a neurodegenerative phenotype with aging. Sci Rep. 2017; 7:7748. https://doi.org/10.1038/s41598-017-07570-5 [PubMed]
- 41. Qrareya AN, Mahdi F, Kaufman MJ, Ashpole NM, Paris JJ. HIV-1 Tat promotes age-related cognitive, anxiety-like, and antinociceptive impairments in female mice that are moderated by aging and endocrine status. Geroscience. 2021; 43:309–27. https://doi.org/10.1007/s11357-020-00268-z [PubMed]
- 42. Salahuddin MF, Qrareya AN, Mahdi F, Jackson D, Foster M, Vujanovic T, Box JG, Paris JJ. Combined HIV-1 Tat and oxycodone activate the hypothalamic-pituitary-adrenal and -gonadal axes and promote psychomotor, affective, and cognitive dysfunction in female mice. Horm Behav. 2020; 119:104649. https://doi.org/10.1016/j.yhbeh.2019.104649 [PubMed]
- 43. Kendall SL, Anderson CF, Nath A, Turchan-Cholewo J, Land CL, Mactutus CF, Booze RM. Gonadal steroids differentially modulate neurotoxicity of HIV and cocaine: testosterone and ICI 182,780 sensitive mechanism. BMC Neurosci. 2005; 6:40. https://doi.org/10.1186/1471-2202-6-40 [PubMed]
- 44. Wilson ME, Allred KF, Bisotti AJ, Bruce-Keller A, Chuahan A, Nath A. Estradiol negatively regulates HIV-LTR promoter activity in glial cells. AIDS Res Hum Retroviruses. 2006; 22:350–6. https://doi.org/10.1089/aid.2006.22.350 [PubMed]
- 45. Paris JJ, Zou S, Hahn YK, Knapp PE, Hauser KF. 5α-reduced progestogens ameliorate mood-related behavioral pathology, neurotoxicity, and microgliosis associated with exposure to HIV-1 Tat. Brain Behav Immun. 2016; 55:202–14. https://doi.org/10.1016/j.bbi.2016.01.007 [PubMed]
- 46. Paris JJ, Fenwick J, McLaughlin JP. Progesterone protects normative anxiety-like responding among ovariectomized female mice that conditionally express the HIV-1 regulatory protein, Tat, in the CNS. Horm Behav. 2014; 65:445–53. https://doi.org/10.1016/j.yhbeh.2014.04.001 [PubMed]
- 47. Wang JM, Liu L, Irwin RW, Chen S, Brinton RD. Regenerative potential of allopregnanolone. Brain Res Rev. 2008; 57:398–409. https://doi.org/10.1016/j.brainresrev.2007.08.010 [PubMed]
- 48. Wang JM, Singh C, Liu L, Irwin RW, Chen S, Chung EJ, Thompson RF, Brinton RD. Allopregnanolone reverses neurogenic and cognitive deficits in mouse model of Alzheimer's disease. Proc Natl Acad Sci U S A. 2010; 107:6498–503. https://doi.org/10.1073/pnas.1001422107 [PubMed]
- 49. Greene M, Covinsky KE, Valcour V, Miao Y, Madamba J, Lampiris H, Cenzer IS, Martin J, Deeks SG. Geriatric Syndromes in Older HIV-Infected Adults. J Acquir Immune Defic Syndr. 2015; 69:161–7. https://doi.org/10.1097/QAI.0000000000000556 [PubMed]
- 50. Valcour V, Shikuma C, Shiramizu B, Watters M, Poff P, Selnes O, Holck P, Grove J, Sacktor N. Higher frequency of dementia in older HIV-1 individuals: the Hawaii Aging with HIV-1 Cohort. Neurology. 2004; 63:822–7. https://doi.org/10.1212/01.wnl.0000134665.58343.8d [PubMed]
- 51. Wendelken LA, Valcour V. Impact of HIV and aging on neuropsychological function. J Neurovirol. 2012; 18:256–63. https://doi.org/10.1007/s13365-012-0094-1 [PubMed]
- 52. Salahuddin MF, Mahdi F, Paris JJ. HIV-1 Tat Dysregulates the Hypothalamic-Pituitary-Adrenal Stress Axis and Potentiates Oxycodone-Mediated Psychomotor and Anxiety-Like Behavior of Male Mice. Int J Mol Sci. 2020; 21:8212. https://doi.org/10.3390/ijms21218212 [PubMed]
- 53. Feinstein MJ, Bogorodskaya M, Bloomfield GS, Vedanthan R, Siedner MJ, Kwan GF, Longenecker CT. Cardiovascular Complications of HIV in Endemic Countries. Curr Cardiol Rep. 2016; 18:113. https://doi.org/10.1007/s11886-016-0794-x [PubMed]
- 54. Esser S, Gelbrich G, Brockmeyer N, Goehler A, Schadendorf D, Erbel R, Neumann T, Reinsch N. Prevalence of cardiovascular diseases in HIV-infected outpatients: results from a prospective, multicenter cohort study. Clin Res Cardiol. 2013; 102:203–13. https://doi.org/10.1007/s00392-012-0519-0 [PubMed]
- 55. Smit M, Brinkman K, Geerlings S, Smit C, Thyagarajan K, van Sighem A, de Wolf F, Hallett TB, and ATHENA observational cohort. Future challenges for clinical care of an ageing population infected with HIV: a modelling study. Lancet Infect Dis. 2015; 15:810–8. https://doi.org/10.1016/S1473-3099(15)00056-0 [PubMed]
- 56. Ciesla JA, Roberts JE. Meta-analysis of the relationship between HIV infection and risk for depressive disorders. Am J Psychiatry. 2001; 158:725–30. https://doi.org/10.1176/appi.ajp.158.5.725 [PubMed]
- 57. Do AN, Rosenberg ES, Sullivan PS, Beer L, Strine TW, Schulden JD, Fagan JL, Freedman MS, Skarbinski J. Excess burden of depression among HIV-infected persons receiving medical care in the united states: data from the medical monitoring project and the behavioral risk factor surveillance system. PLoS One. 2014; 9:e92842. https://doi.org/10.1371/journal.pone.0092842 [PubMed]
- 58. Kalichman SC, Heckman T, Kochman A, Sikkema K, Bergholte J. Depression and thoughts of suicide among middle-aged and older persons living with HIV-AIDS. Psychiatr Serv. 2000; 51:903–7. https://doi.org/10.1176/appi.ps.51.7.903 [PubMed]
- 59. Penzak SR, Reddy YS, Grimsley SR. Depression in patients with HIV infection. Am J Health Syst Pharm. 2000; 57:376–86. https://doi.org/10.1093/ajhp/57.4.376 [PubMed]
- 60. Groeneweg-Koolhoven I, Comijs HC, Naarding P, de Waal MW, van der Mast RC. Apathy in Older Persons With Depression: Course and Predictors: The NESDO Study. J Geriatr Psychiatry Neurol. 2016; 29:178–86. https://doi.org/10.1177/0891988716632914 [PubMed]
- 61. Burris AS, Banks SM, Carter CS, Davidson JM, Sherins RJ. A long-term, prospective study of the physiologic and behavioral effects of hormone replacement in untreated hypogonadal men. J Androl. 1992; 13:297–304. [PubMed]
- 62. Wang C, Alexander G, Berman N, Salehian B, Davidson T, McDonald V, Steiner B, Hull L, Callegari C, Swerdloff RS. Testosterone replacement therapy improves mood in hypogonadal men--a clinical research center study. J Clin Endocrinol Metab. 1996; 81:3578–83. https://doi.org/10.1210/jcem.81.10.8855804 [PubMed]
- 63. Hogervorst E, Combrinck M, Smith AD. Testosterone and gonadotropin levels in men with dementia. Neuro Endocrinol Lett. 2003; 24:203–8. [PubMed]
- 64. Hogervorst E, Bandelow S, Combrinck M, Smith AD. Low free testosterone is an independent risk factor for Alzheimer's disease. Exp Gerontol. 2004; 39:1633–9. https://doi.org/10.1016/j.exger.2004.06.019 [PubMed]
- 65. Hsu B, Cumming RG, Waite LM, Blyth FM, Naganathan V, Le Couteur DG, Seibel MJ, Handelsman DJ. Longitudinal Relationships between Reproductive Hormones and Cognitive Decline in Older Men: The Concord Health and Ageing in Men Project. J Clin Endocrinol Metab. 2015; 100:2223–30. https://doi.org/10.1210/jc.2015-1016 [PubMed]
- 66. Grinspoon S, Corcoran C, Stanley T, Baaj A, Basgoz N, Klibanski A. Effects of hypogonadism and testosterone administration on depression indices in HIV-infected men. J Clin Endocrinol Metab. 2000; 85:60–5. https://doi.org/10.1210/jcem.85.1.6224 [PubMed]
- 67. Monroe AK, Dobs AS, Palella FJ, Kingsley LA, Witt MD, Brown TT. Morning free and total testosterone in HIV-infected men: implications for the assessment of hypogonadism. AIDS Res Ther. 2014; 11:6. https://doi.org/10.1186/1742-6405-11-6 [PubMed]
- 68. Schneider HJ, Sievers C, Klotsche J, Böhler S, Pittrow D, Lehnert H, Wittchen HU, Stalla GK. Prevalence of low male testosterone levels in primary care in Germany: cross-sectional results from the DETECT study. Clin Endocrinol (Oxf). 2009; 70:446–54. https://doi.org/10.1111/j.1365-2265.2008.03370.x [PubMed]
- 69. Carey AN, Sypek EI, Singh HD, Kaufman MJ, McLaughlin JP. Expression of HIV-Tat protein is associated with learning and memory deficits in the mouse. Behav Brain Res. 2012; 229:48–56. https://doi.org/10.1016/j.bbr.2011.12.019 [PubMed]
- 70. Joshi CR, Stacy S, Sumien N, Ghorpade A, Borgmann K. Astrocyte HIV-1 Tat Differentially Modulates Behavior and Brain MMP/TIMP Balance During Short and Prolonged Induction in Transgenic Mice. Front Neurol. 2020; 11:593188. https://doi.org/10.3389/fneur.2020.593188 [PubMed]
- 71. Marks WD, Paris JJ, Schier CJ, Denton MD, Fitting S, McQuiston AR, Knapp PE, Hauser KF. HIV-1 Tat causes cognitive deficits and selective loss of parvalbumin, somatostatin, and neuronal nitric oxide synthase expressing hippocampal CA1 interneuron subpopulations. J Neurovirol. 2016; 22:747–62. https://doi.org/10.1007/s13365-016-0447-2 [PubMed]
- 72. Marks WD, Paris JJ, Barbour AJ, Moon J, Carpenter VJ, McLane VD, Lark ARS, Nass SR, Zhang J, Yarotskyy V, McQuiston AR, Knapp PE, Hauser KF. HIV-1 Tat and Morphine Differentially Disrupt Pyramidal Cell Structure and Function and Spatial Learning in Hippocampal Area CA1: Continuous versus Interrupted Morphine Exposure. eNeuro. 2021; 8:ENEURO.0547-20.2021. https://doi.org/10.1523/ENEURO.0547-20.2021 [PubMed]
- 73. Nookala AR, Schwartz DC, Chaudhari NS, Glazyrin A, Stephens EB, Berman NEJ, Kumar A. Methamphetamine augment HIV-1 Tat mediated memory deficits by altering the expression of synaptic proteins and neurotrophic factors. Brain Behav Immun. 2018; 71:37–51. https://doi.org/10.1016/j.bbi.2018.04.018 [PubMed]
- 74. Zhao X, Fan Y, Vann PH, Wong JM, Sumien N, He JJ. Long-term HIV-1 Tat Expression in the Brain Led to Neurobehavioral, Pathological, and Epigenetic Changes Reminiscent of Accelerated Aging. Aging Dis. 2020; 11:93–107. https://doi.org/10.14336/AD.2019.0323 [PubMed]
- 75. Martin C, Solders G, Sönnerborg A, Hansson P. Painful and non-painful neuropathy in HIV-infected patients: an analysis of somatosensory nerve function. Eur J Pain. 2003; 7:23–31. https://doi.org/10.1016/s1090-3801(02)00053-8 [PubMed]
- 76. Bagdas D, Paris JJ, Carper M, Wodarski R, Rice ASC, Knapp PE, Hauser KF, Damaj MI. Conditional expression of HIV-1 tat in the mouse alters the onset and progression of tonic, inflammatory and neuropathic hypersensitivity in a sex-dependent manner. Eur J Pain. 2020; 24:1609–23. https://doi.org/10.1002/ejp.1618 [PubMed]
- 77. Wodarski R, Bagdas D, Paris JJ, Pheby T, Toma W, Xu R, Damaj MI, Knapp PE, Rice ASC, Hauser KF. Reduced intraepidermal nerve fibre density, glial activation, and sensory changes in HIV type-1 Tat-expressing female mice: involvement of Tat during early stages of HIV-associated painful sensory neuropathy. Pain Rep. 2018; 3:e654. https://doi.org/10.1097/PR9.0000000000000654 [PubMed]
- 78. Schütz SG, Robinson-Papp J. HIV-related neuropathy: current perspectives. HIV AIDS (Auckl). 2013; 5:243–51. https://doi.org/10.2147/HIV.S36674 [PubMed]
- 79. Huang W, Calvo M, Karu K, Olausen HR, Bathgate G, Okuse K, Bennett DLH, Rice ASC. A clinically relevant rodent model of the HIV antiretroviral drug stavudine induced painful peripheral neuropathy. Pain. 2013; 154:560–75. https://doi.org/10.1016/j.pain.2012.12.023 [PubMed]
- 80. Ances BM, Vaida F, Rosario D, Marquie-Beck J, Ellis RJ, Simpson DM, Clifford DB, McArthur JC, Grant I, McCutchan JA, and CNS HIV Antiretroviral Therapy Effects Research (CHARTER) Metabolic Study Group. Role of metabolic syndrome components in HIV-associated sensory neuropathy. AIDS. 2009; 23:2317–22. https://doi.org/10.1097/QAD.0b013e328332204e [PubMed]
- 81. Cherry CL, Affandi JS, Imran D, Yunihastuti E, Smyth K, Vanar S, Kamarulzaman A, Price P. Age and height predict neuropathy risk in patients with HIV prescribed stavudine. Neurology. 2009; 73:315–20. https://doi.org/10.1212/WNL.0b013e3181af7a22 [PubMed]
- 82. Ellis RJ, Rosario D, Clifford DB, McArthur JC, Simpson D, Alexander T, Gelman BB, Vaida F, Collier A, Marra CM, Ances B, Atkinson JH, Dworkin RH, et al, and CHARTER Study Group. Continued high prevalence and adverse clinical impact of human immunodeficiency virus-associated sensory neuropathy in the era of combination antiretroviral therapy: the CHARTER Study. Arch Neurol. 2010; 67:552–8. https://doi.org/10.1001/archneurol.2010.76 [PubMed]
- 83. Oh SB, Tran PB, Gillard SE, Hurley RW, Hammond DL, Miller RJ. Chemokines and glycoprotein120 produce pain hypersensitivity by directly exciting primary nociceptive neurons. J Neurosci. 2001; 21:5027–35. https://doi.org/10.1523/jneurosci.21-14-05027.2001 [PubMed]
- 84. Wallace VC, Blackbeard J, Pheby T, Segerdahl AR, Davies M, Hasnie F, Hall S, McMahon SB, Rice AS. Pharmacological, behavioural and mechanistic analysis of HIV-1 gp120 induced painful neuropathy. Pain. 2007; 133:47–63. https://doi.org/10.1016/j.pain.2007.02.015 [PubMed]
- 85. Frye CA, Edinger K, Sumida K. Androgen administration to aged male mice increases anti-anxiety behavior and enhances cognitive performance. Neuropsychopharmacology. 2008; 33:1049–61. https://doi.org/10.1038/sj.npp.1301498 [PubMed]
- 86. Frye CA, Walf AA. Depression-like behavior of aged male and female mice is ameliorated with administration of testosterone or its metabolites. Physiol Behav. 2009; 97:266–9. https://doi.org/10.1016/j.physbeh.2009.02.022 [PubMed]
- 87. Parks EE, Logan S, Yeganeh A, Farley JA, Owen DB, Sonntag WE. Interleukin 6 reduces allopregnanolone synthesis in the brain and contributes to age-related cognitive decline in mice. J Lipid Res. 2020; 61:1308–19. https://doi.org/10.1194/jlr.RA119000479 [PubMed]
- 88. Singh C, Liu L, Wang JM, Irwin RW, Yao J, Chen S, Henry S, Thompson RF, Brinton RD. Allopregnanolone restores hippocampal-dependent learning and memory and neural progenitor survival in aging 3xTgAD and nonTg mice. Neurobiol Aging. 2012; 33:1493–506. https://doi.org/10.1016/j.neurobiolaging.2011.06.008 [PubMed]
- 89. Paris JJ, Chen X, Anderson J, Qrareya AN, Mahdi F, Du F, McLaughlin JP, Kaufman MJ. In vivo proton magnetic resonance spectroscopy detection of metabolite abnormalities in aged Tat-transgenic mouse brain. Geroscience. 2021; 43:1851–62. https://doi.org/10.1007/s11357-021-00354-w [PubMed]
- 90. Harezlak J, Buchthal S, Taylor M, Schifitto G, Zhong J, Daar E, Alger J, Singer E, Campbell T, Yiannoutsos C, Cohen R, Navia B, and HIV Neuroimaging Consortium. Persistence of HIV-associated cognitive impairment, inflammation, and neuronal injury in era of highly active antiretroviral treatment. AIDS. 2011; 25:625–33. https://doi.org/10.1097/QAD.0b013e3283427da7 [PubMed]
- 91. Cohen RA, Harezlak J, Gongvatana A, Buchthal S, Schifitto G, Clark U, Paul R, Taylor M, Thompson P, Tate D, Alger J, Brown M, Zhong J, et al, and HIV Neuroimaging Consortium. Cerebral metabolite abnormalities in human immunodeficiency virus are associated with cortical and subcortical volumes. J Neurovirol. 2010; 16:435–44. https://doi.org/10.3109/13550284.2010.520817 [PubMed]
- 92. Ances BM, Ortega M, Vaida F, Heaps J, Paul R. Independent effects of HIV, aging, and HAART on brain volumetric measures. J Acquir Immune Defic Syndr. 2012; 59:469–77. https://doi.org/10.1097/QAI.0b013e318249db17 [PubMed]
- 93. Ortega M, Ances BM. Role of HIV in amyloid metabolism. J Neuroimmune Pharmacol. 2014; 9:483–91. https://doi.org/10.1007/s11481-014-9546-0 [PubMed]
- 94. Anthony IC, Ramage SN, Carnie FW, Simmonds P, Bell JE. Accelerated Tau deposition in the brains of individuals infected with human immunodeficiency virus-1 before and after the advent of highly active anti-retroviral therapy. Acta Neuropathol. 2006; 111:529–38. https://doi.org/10.1007/s00401-006-0037-0 [PubMed]
- 95. Seider TR, Luo X, Gongvatana A, Devlin KN, de la Monte SM, Chasman JD, Yan P, Tashima KT, Navia B, Cohen RA. Verbal memory declines more rapidly with age in HIV infected versus uninfected adults. J Clin Exp Neuropsychol. 2014; 36:356–67. https://doi.org/10.1080/13803395.2014.892061 [PubMed]
- 96. Irwin RW, Solinsky CM, Brinton RD. Frontiers in therapeutic development of allopregnanolone for Alzheimer's disease and other neurological disorders. Front Cell Neurosci. 2014; 8:203. https://doi.org/10.3389/fncel.2014.00203 [PubMed]
- 97. Pezzaioli LC, Quiros-Roldan E, Paghera S, Porcelli T, Maffezzoni F, Delbarba A, Degli Antoni M, Cappelli C, Castelli F, Ferlin A. The importance of SHBG and calculated free testosterone for the diagnosis of symptomatic hypogonadism in HIV-infected men: a single-centre real-life experience. Infection. 2021; 49:295–303. https://doi.org/10.1007/s15010-020-01558-6 [PubMed]
- 98. Wong N, Levy M, Stephenson I. Hypogonadism in the HIV-Infected Man. Curr Treat Options Infect Dis. 2017; 9:104–16. https://doi.org/10.1007/s40506-017-0110-3 [PubMed]
- 99. Maingat FG, Polyak MJ, Paul AM, Vivithanaporn P, Noorbakhsh F, Ahboucha S, Baker GB, Pearson K, Power C. Neurosteroid-mediated regulation of brain innate immunity in HIV/AIDS: DHEA-S suppresses neurovirulence. FASEB J. 2013; 27:725–37. https://doi.org/10.1096/fj.12-215079 [PubMed]
- 100. Mukerji SS, Misra V, Lorenz DR, Chettimada S, Keller K, Letendre S, Ellis RJ, Morgello S, Parker RA, Gabuzda D. Low Neuroactive Steroids Identifies a Biological Subtype of Depression in Adults with Human Immunodeficiency Virus on Suppressive Antiretroviral Therapy. J Infect Dis. 2021; 223:1601–11. https://doi.org/10.1093/infdis/jiaa104 [PubMed]
- 101. Amini Lari M, Parsa N, Marzban M, Shams M, Faramarzi H. Depression, Testosterone concentration, sexual dysfunction and methadone use among men with hypogonadism and HIV Infection. AIDS Behav. 2012; 16:2236–43. https://doi.org/10.1007/s10461-012-0234-x [PubMed]
- 102. Rabkin JG, Wagner GJ, Rabkin R. A double-blind, placebo-controlled trial of testosterone therapy for HIV-positive men with hypogonadal symptoms. Arch Gen Psychiatry. 2000; 57:141–7. https://doi.org/10.1001/archpsyc.57.2.141 [PubMed]
- 103. Bruce-Keller AJ, Turchan-Cholewo J, Smart EJ, Geurin T, Chauhan A, Reid R, Xu R, Nath A, Knapp PE, Hauser KF. Morphine causes rapid increases in glial activation and neuronal injury in the striatum of inducible HIV-1 Tat transgenic mice. Glia. 2008; 56:1414–27. https://doi.org/10.1002/glia.20708 [PubMed]
- 104. Zou S, Balinang JM, Paris JJ, Hauser KF, Fuss B, Knapp PE. Effects of HIV-1 Tat on oligodendrocyte viability are mediated by CaMKIIβ-GSK3β interactions. J Neurochem. 2019; 149:98–110. https://doi.org/10.1111/jnc.14668 [PubMed]
- 105. Dutta S, Sengupta P. Men and mice: Relating their ages. Life Sci. 2016; 152:244–8. https://doi.org/10.1016/j.lfs.2015.10.025 [PubMed]
- 106. Xie Y, Seawell J, Boesch E, Allen L, Suchy A, Longo FM, Meeker RB. Small molecule modulation of the p75 neurotrophin receptor suppresses age- and genotype-associated neurodegeneration in HIV gp120 transgenic mice. Exp Neurol. 2021; 335:113489. https://doi.org/10.1016/j.expneurol.2020.113489 [PubMed]
- 107. Ohene-Nyako M, Nass SR, Hahn YK, Knapp PE, Hauser KF. Morphine and HIV-1 Tat interact to cause region-specific hyperphosphorylation of tau in transgenic mice. Neurosci Lett. 2021; 741:135502. https://doi.org/10.1016/j.neulet.2020.135502 [PubMed]
- 108. Nass SR, Hahn YK, McLane VD, Varshneya NB, Damaj MI, Knapp PE, Hauser KF. Chronic HIV-1 Tat exposure alters anterior cingulate cortico-basal ganglia-thalamocortical synaptic circuitry, associated behavioral control, and immune regulation in male mice. Brain Behav Immun Health. 2020; 5:100077. https://doi.org/10.1016/j.bbih.2020.100077 [PubMed]
- 109. Lucchetti J, Fracasso C, Balducci C, Passoni A, Forloni G, Salmona M, Gobbi M. Plasma and Brain Concentrations of Doxycycline after Single and Repeated Doses in Wild-Type and APP23 Mice. J Pharmacol Exp Ther. 2019; 368:32–40. https://doi.org/10.1124/jpet.118.252064 [PubMed]
- 110. Hall C, Ballachey EL. A study of the rat’s behavior in a field. A contribution to method in comparative psychology. University of California Publications in Psychology. 1932; 6:1–12.
- 111. Lister RG. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology (Berl). 1987; 92:180–5. https://doi.org/10.1007/BF00177912 [PubMed]
- 112. Buresová O, Bures J, Oitzl MS, Zahálka A. Radial maze in the water tank: an aversively motivated spatial working memory task. Physiol Behav. 1985; 34:1003–5. https://doi.org/10.1016/0031-9384(85)90028-9 [PubMed]
- 113. Sparkman NL, Buchanan JB, Dos Santos NL, Johnson RW, Burton MD. Aging sensitizes male mice to cognitive dysfunction induced by central HIV-1 gp120. Exp Gerontol. 2019; 126:110694. https://doi.org/10.1016/j.exger.2019.110694 [PubMed]
- 114. Crabbe JC, Cotnam CJ, Cameron AJ, Schlumbohm JP, Rhodes JS, Metten P, Wahlsten D. Strain differences in three measures of ethanol intoxication in mice: the screen, dowel and grip strength tests. Genes Brain Behav. 2003; 2:201–13. https://doi.org/10.1034/j.1601-183x.2003.00023.x [PubMed]
- 115. Kim SH, Chung JM. Sympathectomy alleviates mechanical allodynia in an experimental animal model for neuropathy in the rat. Neurosci Lett. 1991; 134:131–4. https://doi.org/10.1016/0304-3940(91)90524-w [PubMed]
- 116. Deuis JR, Dvorakova LS, Vetter I. Methods Used to Evaluate Pain Behaviors in Rodents. Front Mol Neurosci. 2017; 10:284. https://doi.org/10.3389/fnmol.2017.00284 [PubMed]
- 117. Deuis JR, Vetter I. The thermal probe test: A novel behavioral assay to quantify thermal paw withdrawal thresholds in mice. Temperature (Austin). 2016; 3:199–207. https://doi.org/10.1080/23328940.2016.1157668 [PubMed]
- 118. Salahuddin MF, Mahdi F, Sulochana SP, Paris JJ. HIV-1 Tat Protein Promotes Neuroendocrine Dysfunction Concurrent with the Potentiation of Oxycodone's Psychomotor Effects in Female Mice. Viruses. 2021; 13:813. https://doi.org/10.3390/v13050813 [PubMed]