



Introduction
Ulcerative colitis (UC) is the inflammation of the rectal mucosa that usually extends proximally to other areas of the colon [1]. UC is characterized by cytokine imbalance, immune dysfunction and intestinal mucosa-associated inflammation [1]. UC patients generally present with bloody diarrhea, abdominal pain and tenesmus, with increasing global incidence [2]. There are approximately 286 cases per 100,000 people in the United States and 505 cases per 100,000 people in Europe [3]. There are limited data in developing countries. However, the incidence of UC is also increasing year by year in Asia, Middle East and South America [4].
The pathogenesis of UC remains unclear at present, which is considered to be associated with the interaction of multiple environmental, genetic and immune factors [5]. The common pathogenic feature is the destruction of the integrity of the intestinal epithelial barrier. Under normal circumstances, several types of cellular proteins secreted by epithelial cells can support the intestinal mucosal barrier. The barrier is mainly composed of intercellular junction complexes, consisting of tight junction (TJ) protein (occlusion proteins and claudin-1) and zona [6]. The destruction of TJ leads to the disorder of the paracellular barrier and the increasing permeability of the intestinal epithelial cells [7]. This change in permeability can cause the penetration of harmful antigens and intraluminal bacteria into the intestine, leading to the initiation and acceleration of mucosal inflammation of UC [8]. Therefore, therapies to attenuate intestinal barrier dysfunction can effectively treat UC.
AMP-activated protein kinase (AMPK) is a highly conserved cellular energy sensor. Under inflammatory pathological conditions, the phosphorylation level of AMPK is decreased, which subsequently affects the expression of inflammatory mediators and downstream biological functions [9, 10]. AMPK activators can exert anti-inflammatory effects [11, 12].
AMPK may inhibit the pathogenesis of inflammatory bowel disease (IBD) by inhibiting the activity of nuclear factor-kappaB (NF-kappaB) [13]. The expression of NF-κB is increased in the colon lesion of IBD patients [14]. In the dextran sulfate sodium (DSS)-induced mouse colitis model, metformin, an AMPK activator, could significantly inhibit the activation of IKK induced by DSS, thereby down-regulating the expression of the inflammatory factor IL-18 [15]. Additionally, AMPK can suppress the activation of NLRP3 inflammasome by decreasing NF-κB activation, thereby attenuating colitis [16].
Pyroptosis is a form of intestinal cell death, which is characterized by swollen cells and the appearance of large bubbles on the cell membrane [17]. Studies have reported that attenuating pyroptosis can relieve experimental colitis [17, 18]. Studies have also shown that the nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin domain-containing 3 (NLRP3) inflammasome is an important signal transduction pathway of colitis injury, playing an important role in regulating cell proliferation, differentiation and apoptosis [18, 19]. Caspase-1 and NLRP3 are key components of inflammation, and their dysregulation promotes the production of UC-associated pro-inflammatory cytokines (interleukin-1beta (IL-1β) and IL-18) [20].
Inflammasome proteins include NLRP1, NLRP3, NLRC4, and NLRP6, etc., which are assembled with apoptosis-associated speck-like protein (ASC) and pro-inflammatory caspases (Caspase-1 and Caspase-11) into the inflammasome complex [21]. It can not only lead to the automatic activation of caspases and control the cleavage of IL-1β and IL-18 precursors into their mature forms, but also induce pyroptosis [22]. The IL-1 cytokine family can regulate the intestinal homeostasis, inflammation and healthy microbiota [23]. To be specific, NLRP3 inflammasome induces IL-1β in myeloid cells and promotes intestinal inflammation [23]. Therefore, the function of NLRP3 in mucosal immunity and colitis seems to be complex, with unclear underlying mechanism [24].
Natural products commonly used in traditional Chinese medicine (TCM) have attracted accumulative medical interest globally due to their powerful anti-inflammatory effects [25]. Schisandra chinensis, a type of TCM, is originally produced in northeastern China, South Korea, eastern Russia and Japan, which is used to treat various diseases, including autoimmune diseases, cardiovascular diseases, acne and neurological diseases [26]. At present, the chemical composition of Schisandra chinensis is not completely understood. Schisandrin B is the most important ingredient in Schisandra chinensis, which has been revealed to have anti-cancer, anti-inflammation, hepatoprotection, anti-bacteria and many other activities by accumulative evidence [27]. Schisandrin B and Schisandrol are representative lignans of Schisandra chinensis [27]. Multiple studies have demonstrated their anti-inflammatory effects. According to relevant reports, Schisandrin B can significantly inhibit the level of inflammation by blocking NLRP3 activation and of IL-1β secretion [28]. However, there is no study concerning the specific mechanism of Schisandrin B in attenuating pyroptosis. Meanwhile, there is also no study on Schisandrin B in relieving sepsis-induced colitis by regulating pyroptosis [29, 30]. Additionally, Schisandrin B has also been revealed to play a protective role in arthritis [31]. Therefore, in this study, we mainly investigate the molecular mechanism of Schisandrin B in relieving epithelial cell injury and regulating pyroptosis in colitis.
Materials and Methods
Animal care model of colitis and treatment
All experiments related to animal were approved by the Animal Care and Use Committee of Wannan Medical College. All C57BL/6 mice (5-6 weeks, 18-20 g) were obtained from Animal testing center of Qinglongshan (Nanjing, Suzhou, China). C57BL/6 mice were randomly assigned to three groups: sham group (n = 6), model group (n = 8), 10 mg/kg Schisandrin B (Sigma-Aldrich LLC.) treatment group (n = 8). All mice of model group were induced by giving 2.0% dextran sulfate sodium (DSS, MP) in the drinking water for 7 days. All mice of Schisandrin B were given with 10 mg/kg/ per day Schisandrin B body weight by intraperitoneal injection 1 h, and then induced by giving 2.0% dextran sulfate sodium (DSS, MP) in the drinking water for 7 days. Weight of every mice were measured at every day.
AMPK i group (AMPK inhibitor, n = 8), all mice of Schisandrin B were given with 10 mg/kg/day Schisandrin B (7 day) and 10 mg/kg of Dorsomorphin (7 day, MedChemExpress, i.p.) body weight by intraperitoneal injection 1 h, and then induced by giving 2.0% dextran sulfate sodium (DSS, MP) in the drinking water for 7 days.
After induction model, mice were anaesthetized with isoflurane (induction: 3-5%, maintenance: 1.5-3%), and peripheral blood were collected form submaxillary vein. Then, mice with sacrificed using cervical dislocation under anaesthesia (1.5-3% of isoflurane). Colon (1 cm) was collected and fixed in 4% paraformaldehyde for histological examination. colon was collected and saved at -80° C for other experiment.
Assessment of disease activity index (DAI): body weight loss <2%, 0 Score; body weight loss≥2%-<5%, 1 Score; body weight loss≥5%-<10%, 2 Score; body weight loss≥10%-<15%, 3 Score; body weight loss≥15%, 4 Score.
Bioinformatics analysis
GEO DataSets and TCMSP server (http://ibts.hkbu.edu.hk/LSP/tcmsp.php) were a systems-level pharmacology database and a flexible, user-friendly web interface. Targets gene and diseases, networks were constructed and analyzed using Cytoscape 3.0 for a deeper understanding of the complex.
Histological examination
Colon tissue samples were fixed in 4% paraformaldehyde, paraffin-embedded and then sectioned into 5 μM slices for H&E staining. Colon tissue samples were observed using fluorescence microscope (Zeiss Axio Observer A1, Germany). Colon tissue scored according to the following criteria: 1, normal; 2, minimal (little) damage; 3, mild damage; 4, moderate damage; 5, severe damage; and 6, maximal damage as document [13].
Cell culture and treatment
HCT-116 cells were seeded in culture dish with RPMI 1640 (Gibco) supplemented with 10% FBS (Gibco) under a humidified 5% (v/v) CO2 atmosphere at 37° C. HCT-116 cells of vitro model group were stimulated with 500 ng/ml of LPS (Sigma-Aldrich, MO, USA) for 4 h, and then pulsed with 1 mM of ATP (Sigma-Aldrich, MO, USA) for 30 min as references [32].
HCT-116 cells of Schisandrin B group were stimulated with 500 ng/ml of LPS (Sigma-Aldrich, MO, USA) or 40 μM of Schisandrin B for 4 h without FBS, and then pulsed with 1 mM of ATP (Sigma-Aldrich, MO, USA) for 30 min.
AMPK i group of HCT-116 cells were stimulated with (500 ng/ml) LPS+40 μM of Schisandrin B + epiberberine (25 μM, MedChemExpress) for 4 h, and then pulsed with 1 mM of ATP (Sigma-Aldrich, MO, USA) for 30 min as document [33].
ELISA assay, ROS production and JC-1 assay
After induction model, mice were anaesthetized with Isoflurane (Induction: 3-5%, Maintenance: 1.5-3%), and peripheral blood were collected form submaxillary vein. Blood was centrifugated at1000 g at 4° C for 10 min and serum was collected and saved at -80° C for other experiment. Blood or cell samples were collected and used to measure ROS (E004-1-1), TNF-α (H052-1), IL-6 (H007-1-1), IL-18 (H015) and IL-1β (H002) levels using ROS, TNF-α, IL-6, IL-18 and IL-1β ELISA kits (Nanjing Jiancheng Biological Engineering Institute, Nanjing, China) following the manufacturer’s instructions.
1x 105/well cells were seeded into a 96-well plate and 100 μL JC-1 probe solution (C2006, Beyotime Biotechnology) was added into every well as previously described [34]. Absorbance was measured using a fluorescent reader (Synergy H1 Microplate Reader, Bio Tek, VT, USA).
Western blotting analysis
Colon tissue samples or cell samples or supernatant samples were split using RIPA assay (Beyotime) in ice. Total proteins were quantified using BCA assay (Beyotime) and were electrophoresed on 10% SDS-acrylamide gels. Total proteins were transferred to nitrocellulose membranes and membranes were blocked with 5% non-fat milk in TBS for 1 h at 37° C. Membranes were incubated with p-AMPK (ab23875, abcam), AMPK (ab32047, abcam), Nrf2 (ab62352, 1:1000, abcam), GSDMD (ab219800, 1:1000, abcam), NLRP3 (sc-66846, 1:500, Santa Cruz, USA), caspase-1 (sc-1780, 1:500, Santa Cruz, USA), and β-Actin (BS6007MH, 1:5000, Bioworld Technology, Inc.) at 4° C overnight. The membranes were incubated with horseradish peroxidase-conjugated secondary antibodies (sc-2004 or sc-2005, 1:5000, Santa Cruz, USA) for 1 h at 37° C after washing with TBST for 15 min. Protein was measured using an enhanced chemiluminescence system (ECL, Beyotime) and analyzed using an Image Lab 3.0 (Bio-Rad Laboratories, Inc.).
Immunofluorescence
Cell were washed with PBS and fixed with 4% paraformaldehyde supplemented with 0.25% Tris-X100 at room temperature for 10 min. After blocking with PBS supplemented with 5% BSA for 2 h at room temperature, cells were incubated with Mito-Tracker Red CMXRos (C1035, Beyotime), NLRP3 (sc-66846, 1:500, Santa Cruz, USA), at 4° C overnight. Cells were incubated with secondary peroxidase conjugated goat anti-rabbit IgG (1:100, Santa Cruz Biotechnology) antibody for 2 h at room temperature, after washing with PBST for 15 min. Cells were stained with DAPI for 15 min at darkness, after washing with PBST for 15 min. Cell samples were observed using fluorescence microscope (Zeiss Axio Observer A1, Germany).
Next, cells were washed with PBS and stained with Mito-Tracker Red CMXRos for 30 min. Cells washed with PBST for 15 min and stained with DCFH-DA for 30 min. Cell samples were observed using fluorescence microscope (Zeiss Axio Observer A1, Germany) after washing with PBST for 15 min.
Cell were washed with PBS and fixed with 4% paraformaldehyde for 15 min. Cells were stained with PI for 15 min at darkness, after washing with PBST for 15 min. Cell samples were observed using fluorescence microscope (Zeiss Axio Observer A1, Germany).
Electron microscopy
Cells were prepared as previously document [35]. 1 x 106 HCT-116 cells were fixed in 0.2 M phosphate buffer supplemented with 2.5% glutaraldehyde (G5882, Sigma-Aldrich) for 4 h. Cells were post-fixed for 60 min in 1% osmium tetroxide (75632, Sigma-Aldrich) and dehydrated in gradient ethanol solutions (15 min each step). Cells were infiltrated sequentially in ethanol and 1:3 of ethanol for 30 min, and finally 100% spurr resin for 24 h at 60° C for polymerization. Cells were examined at 80 kV using a Hitachi H7650 transmission electron microscope (Tokyo, Japan).
Structure the protein of AMPK and molecular docking verification
Molecular docking was performed between the screened representative components and the target gene. The “PDB” file of the protein skeleton of the target gene was downloaded using RCSB PDB database, and the “mol2” file of the molecular structure diagram of the representative components was downloaded using tcmsp database. Import the target gene protein skeleton file into Python PyMOL 2.5.0 software, and use the “remove solvent” and “remove organic” commands to remove water and ligands. Import the processed two types of files into autodock tools software to create a “pdbqt” file, then use autodock Vina software for molecular docking and calculate the binding energy of each conformation, select the conformation with the lowest binding energy to dock with the target protein, save the docked complex file, and import the complex file into Python PyMOL 2.5.0 software for final visualization and local optimization.
This experiment used molecular biology to reform the protein of AMPK, 316-Serine changed 316-Glu.
WT (PRKAA1, 5’-CGGATCCTGCGCAGACTCAGTTCCTGGAGGAA-3’ and 5’-AATCTAATTAAAATTCTTGCACAATAAACTATCGATG-3’, Sangon Biotech Co., Ltd.) and Mutant (5’-CAAGCTTAGTTTGAGTGCGAAGAAGAGGAAGTTA-3’ and 5’-CTCAAAGCACTTCTTCTCCTTCAAGGCGGCGGCAA-3’, Sangon Biotech Co., Ltd.) plasmids were transfected into HCT-116 cells using Lipofectamine 2000 for 48 h. Then, HCT-116 cells were stimulated with 500 ng/ml of LPS (Sigma-Aldrich, MO, USA) and 40 μM of Schisandrin B for 4 h, and then pulsed with 1 mM of ATP (Sigma-Aldrich, MO, USA) for 30 min.
Cell proliferation and LDH activity
HCT-116 cells were induced for vitro model. Then, 100 μl of cell suspension (1×103/ml) was inoculated into a 96-well plate at 37° C. 10 μl CCK-8 was added followed by cultivation in an incubator at 37° C for 1-2 h. The cell proliferation curve was plotted based on optical density (OD) values. OD values were performed on a microplate reader with a detection wavelength of 450 nm.
HCT-116 cells were induced for vitro model. LDH activity levels were measured using LDH activity kit ELISA kits (C0016, Beyotime, Nanjing, China) following the manufacturer’s instructions.
Flow cytometry
HCT-116 cells were induced for vitro model. Then, cells were collected to measure apoptosis rate using Annexin V-FITC Apoptosis Detection Kit (C1062M Beyotime, Nanjing, China) following the manufacturer’s instructions. Cells were fixed with 4% paraformaldehyde for 15 min and stained with PI and Annexin V-FITC for 15 min at darkness at room temperature. HCT-116 cells were checked using Flow cytometry (C6, BD Biosciences).
Statistical analysis
Data were expressed as mean ± SEM. Multiple comparisons were used GraphPad Prism 8 to perform by one-way ANOVA followed by Tukey’s post-test or Kruskal-Wallis test followed by Dunn’s post hoc test. P values < 0.05 were considered statistically significant.
Data availability
If the data are all contained within the manuscript and/or Supporting Information files, enter the following: All relevant data are within the manuscript and its Supporting Information files.
Results
Schisandrin B presented colitis in mice model
In this experiment, the effect of Schisandrin B in mice model of colitis was explored. The ulcer area, ulcer score, body weight and DAI score in DSS induced colitis mice model group were significantly increased than those in the control normal group (Figure 1). Schisandrin B could not only reduce the ulcer area and ulcer score, but also recover the weight and DAI score in DSS induced colitis mice. Meanwhile, there were significant differences in comparison to the DSS induced colitis model group (Figure 1).
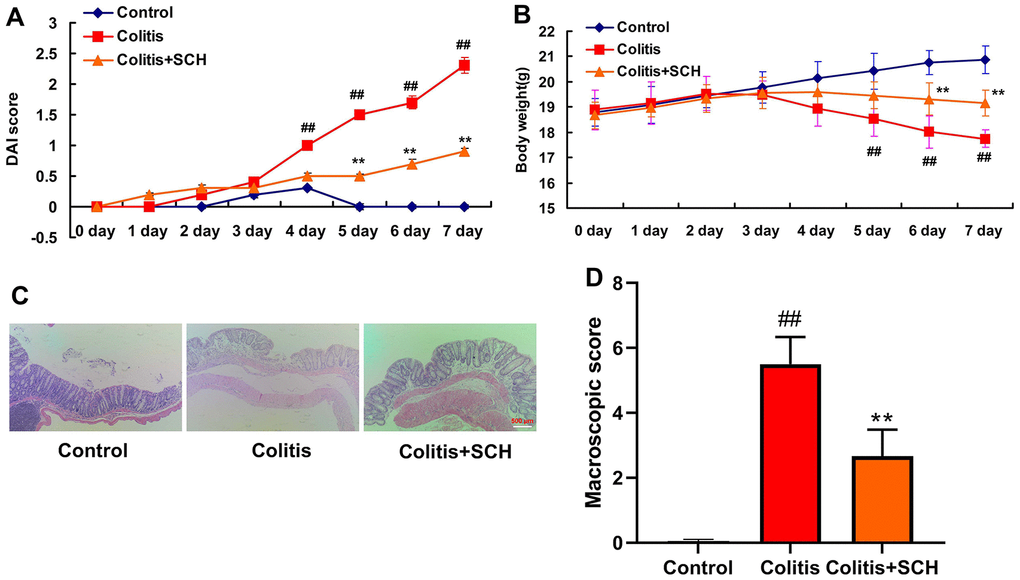
Figure 1. Schisandrin B presented colitis in mice model. (A) DAI score; (B) weight of mice; (C) HE staining results of colon (×200); (D) Colon macroscopic score. ##P<0.01 vs control group; **P<0.01 vs DSS- induced colitis group. Control: blank control group; Colitis: DSS- induced colitis group; Colitis+SCH: Schisandrin was used to treat DSS- induced colitis. SCH, Schisandrin B. Data were expressed as mean ± SEM.
Schisandrin B reduced inflammation in vivo and vitro model of colitis
This study determined whether Schisandrin B reduced inflammation in vivo and vitro model of colitis. It was found that serum TNF-α, IL-6, IL-18 and IL-1β levels in DSS induced colitis mice model was significantly higher than those in the control normal group (Figure 2A–2D). Compared with the DSS induced colitis mice model group (Figure 2A–2D), administration of Schisandrin B more significantly reduced the serum TNF-α, IL-6, IL-18 and IL-1β levels in DSS induced colitis mice model. Apart from that, LPS+ATP induced the supernatant TNF-α, IL-6, IL-18 and IL-1β levels in intestinal epithelial cells (Figure 2E–2H). Schisandrin B reduced the supernatant TNF-α, IL-6, IL-18 and IL-1β levels in intestinal epithelial cells by LPS+ATP (Figure 2E–2H).
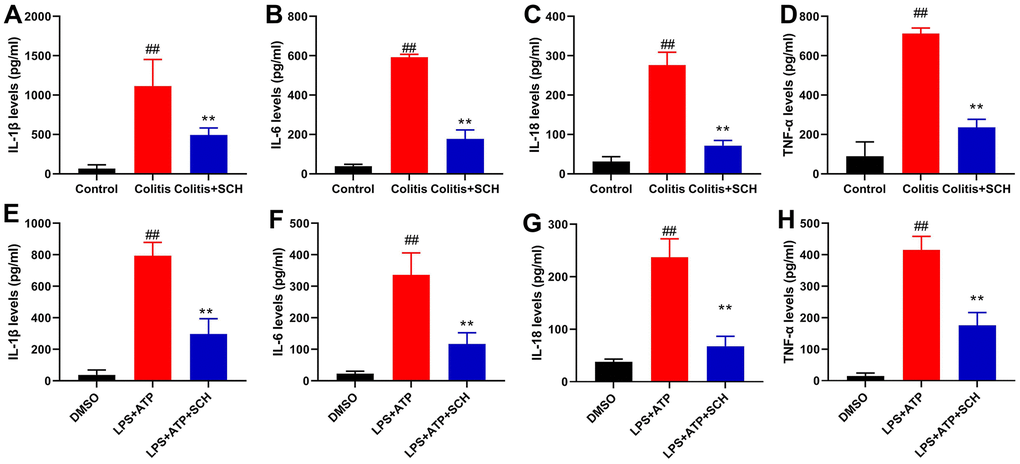
Figure 2. Schisandrin B reduced inflammation in vivo and vitro model of colitis. (A–D) The levels of TNF-α, IL-6, IL-1β and IL-18 in colon tissue; (E–H) the levels of TNF-α, IL-6, IL-1β and IL-18 in intestinal epithelial cells. ##P<0.01 vs control group; **P<0.01 vs DSS- induced colitis group. Control: blank control group; Colitis: DSS- induced colitis group; Colitis+SCH: DSS- induced colitis mice with Schisandrin. ##P<0.01 vs MDSO group; **P<0.01 vs LPS+ATP induced intestinal epithelial cells group. MDSO: blank control group; LPS+ATP: intestinal epithelial cells with LPS+ATP group; LPS+ATP +SCH: intestinal epithelial cells induced by LPS+ATP with Schisandrin. SCH, Schisandrin B. Data were expressed as mean ± SEM.
Schisandrin B suppressed NLRP3 inflammasome in vivo and vitro model of colitis
In order to determine the potential mechanisms by Schisandrin B in model of colitis, NLRP3 inflammasome had been previously defined as the key mediators in inflammation reactions in colitis. Furthermore, NLRP3 protein expression in colon tissue of DSS induced colitis mice model was induced (Figure 3A, 3B). On the one hand, administration of Schisandrin B suppressed NLRP3 protein expression in colon tissue of DSS induced colitis mice model (Figure 3A, 3B). On the other hand, LPS+ATP induced the NLRP3 protein expression in intestinal epithelial cells (Figure 3C, 3D). Schisandrin B reduced NLRP3 protein expression in intestinal epithelial cells by LPS+ATP (Figure 3C, 3D). According to immunofluorescence, LPS+ATP induced NLRP3 expression (green) and activated mitochondrial damage (red) in intestinal epithelial cells. Meanwhile, NLRP3 protein coincided with mitochondrial damage in intestinal epithelial cells (Figure 3E). Schisandrin B suppressed NLRP3 expression (green) and reduced mitochondrial damage (red) in intestinal epithelial cells by LPS+ATP (Figure 3E). These results suggest that Schisandrin B suppressed NLRP3 inflammasome in vivo and vitro model of colitis, which may be attributed to mitochondrial damage.
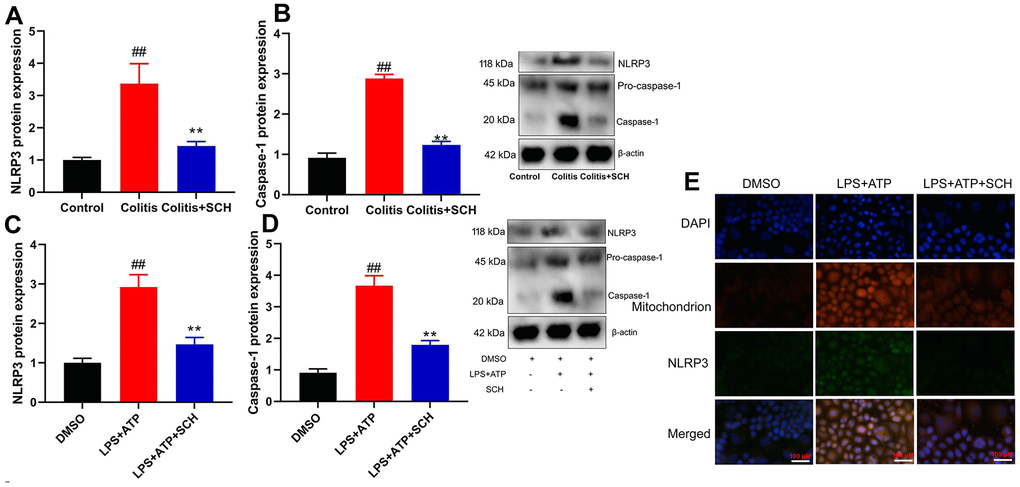
Figure 3. Schisandrin B suppressed NLRP3 inflammasone in vivo and vitro model of colitis. (A, B) the protein expression of NLRP3 and caspase-1 in mouse colon tissue; (C, D) the protein expression of NLRP3 and caspase-1 in intestinal epithelial cells induced by LPS + ATP; (E) the protein expression of NLRP3 and caspase-1 in intestinal epithelial cells detected by cell immunofluorescence. ##P<0.01 vs control group; **P<0.01 vs DSS- induced colitis group. Control: blank control group; Colitis: DSS- induced colitis group; Colitis+SCH: DSS- induced colitis mice with Schisandrin. ##P<0.01 vs MDSO group; **P<0.01 vs LPS+ATP induced intestinal epithelial cells group. MDSO: blank control group; LPS+ATP: intestinal epithelial cells with LPS+ATP group; LPS+ATP +SCH: intestinal epithelial cells induced by LPS+ATP with Schisandrin. SCH, Schisandrin B. Data were expressed as mean ± SEM.
Systematic understanding of how Schisandrin B induced AMPK/Nrf2 signaling pathway in colitis model
TCMSP server (http://ibts.hkbu.edu.hk/LSP/tcmsp.php) was used to further investigate the anti-inflammatory mechanism of Schisandrin B in DSS-induced acute colitis (see physical interactions, pathways and co-localisation in Figure 4A). Obviously, 60.38% showed similar co-expression characteristics, and 30.91% shared the same protein domain. Then, AMPK/Nrf2 signaling for target fishing and pathway was analyzed. Schisandrin B induced p-AMPK and Nrf2 protein expression in colon tissue and intestinal epithelial cells by LPS+ATP (Figure 4B–4E). As shown by these results, Schisandrin B suppressed NLRP3 inflammasome in intestinal epithelial cells by AMPK/Nrf2 signaling.
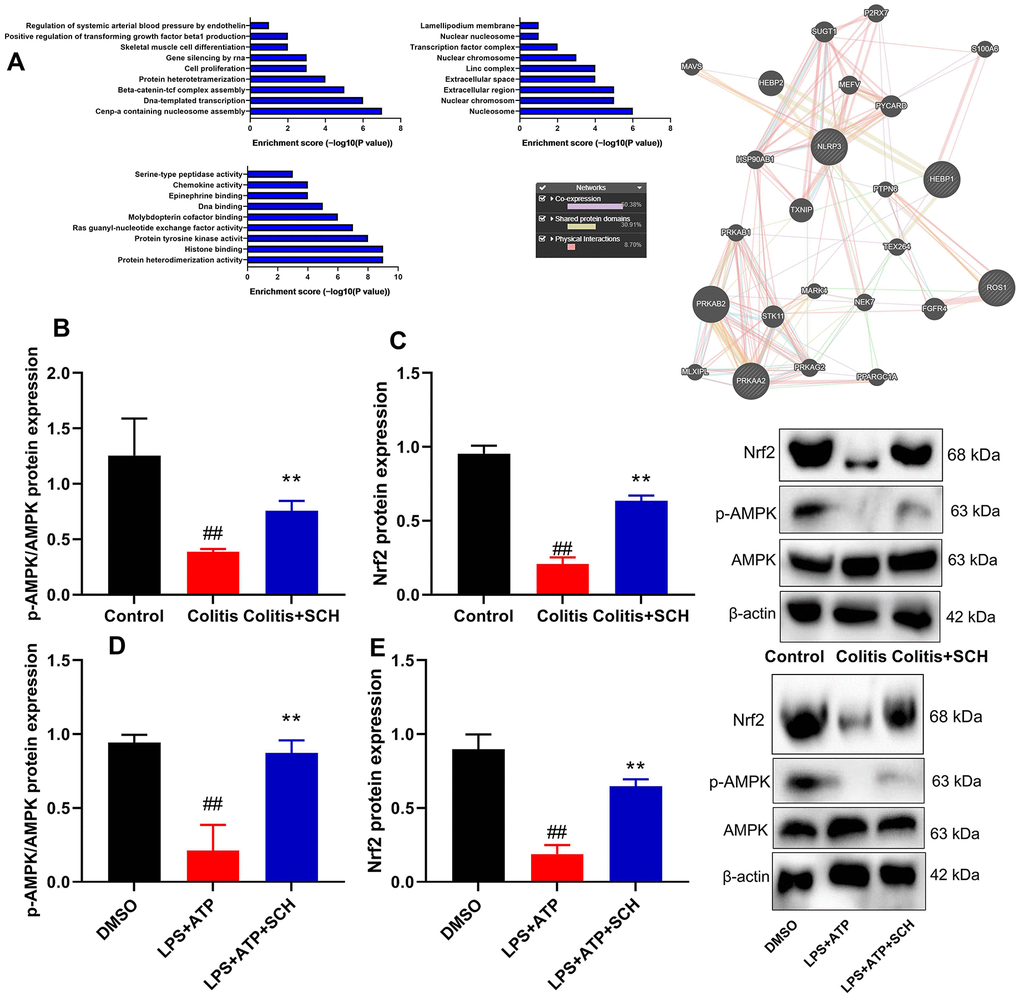
Figure 4. Systematic understanding of Schisandrin B induced AMPK/Nrf2 signaling pathway in vivo and vitro model of colitis. (A) Network of potential Schisandrin B targets; (B, C) p-AMPK and Nrf2 protein expression in mouse colon tissue; (D, E) p-AMPK and Nrf2 protein expression in intestinal epithelial cells induced by LPS + ATP. ##P<0.01 vs control group; **P<0.01 vs DSS- induced colitis group. Control: blank control group; Colitis: DSS- induced colitis group; Colitis+SCH: DSS- induced colitis mice with Schisandrin. ##P<0.01 vs MDSO group; **P<0.01 vs LPS+ATP induced intestinal epithelial cells group. MDSO: blank control group; LPS+ATP: intestinal epithelial cells with LPS+ATP group; LPS+ATP +SCH: intestinal epithelial cells induced by LPS+ATP with Schisandrin. SCH, Schisandrin B. Data were expressed as mean ± SEM.
Schisandrin B regulated AMPK protein at 316 sites
The study explored how Schisandrin B induced AMPK protein
According to the degree value and frequency, AMPK corresponding to only one active component in network screening target was selected for molecular docking. Indeed, the binding energy of receptor and ligand is less than - 6.3 kcal / mol, so that it can be considered to have a strong binding activity. Beyond that, the docking results were imported into Python PyMOL 2.5.0 Open Source Software for final visualization and local optimization. The results are displayed in Figure 5A. Furthermore, we conducted protein structure modification and reformed at 316-SER (Serine) of AMPK protein (Figure 5B, 5C). Compared with WT protein expression, p-AMPK protein expression was more significantly suppressed in 316-mutant protein (Figure 5D). Taken together, these results revealed that Schisandrin B combined AMPK protein at 316-SER.
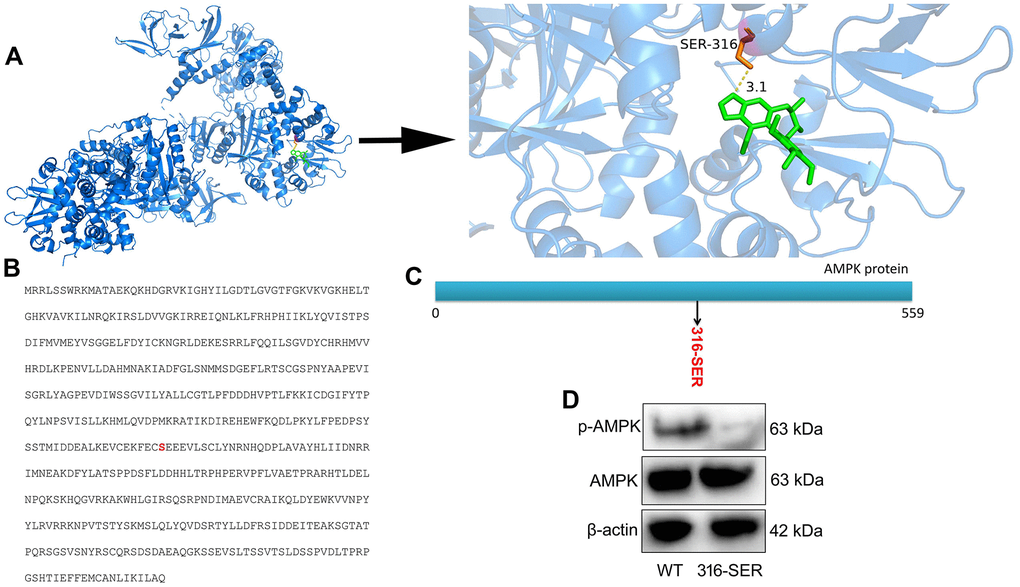
Figure 5. Schisandrin B regulated AMPK protein at 316 site. (A) protein spatial structure and drug structure; (B) amino acid sequence for AMPK protein; (C) Schematic diagram of protein mutation; (D) p- AMPK protein expression.
The inhibition of AMPK reduced the anti-inflammation effects of Schisandrin B on NLRP3 inflammasome
AMPK inhibitor (Dorsomorphin, 10 mg/kg) not only suppressed p-AMPK and Nrf2 protein expression levels, but also induced NLRP3 and Caspase-1 protein expression levels in DSS-induced colitis mice treated with Schisandrin B, in comparison to the group treated with Schisandrin B (Figure 6A–6D). Next, AMPK inhibitor (Dorsomorphin, 10 μM) suppressed p-AMPK and Nrf2 protein expression levels, and induced NLRP3 and Caspase-1 protein expression levels in intestinal epithelial cells treated with LPS+ATP and Schisandrin B, in comparison to the group treated with Schisandrin B (Figure 6E–6H). As revealed by these results, AMPK was the target spot of Schisandrin B on NLRP3 inflammasome in colitis model.
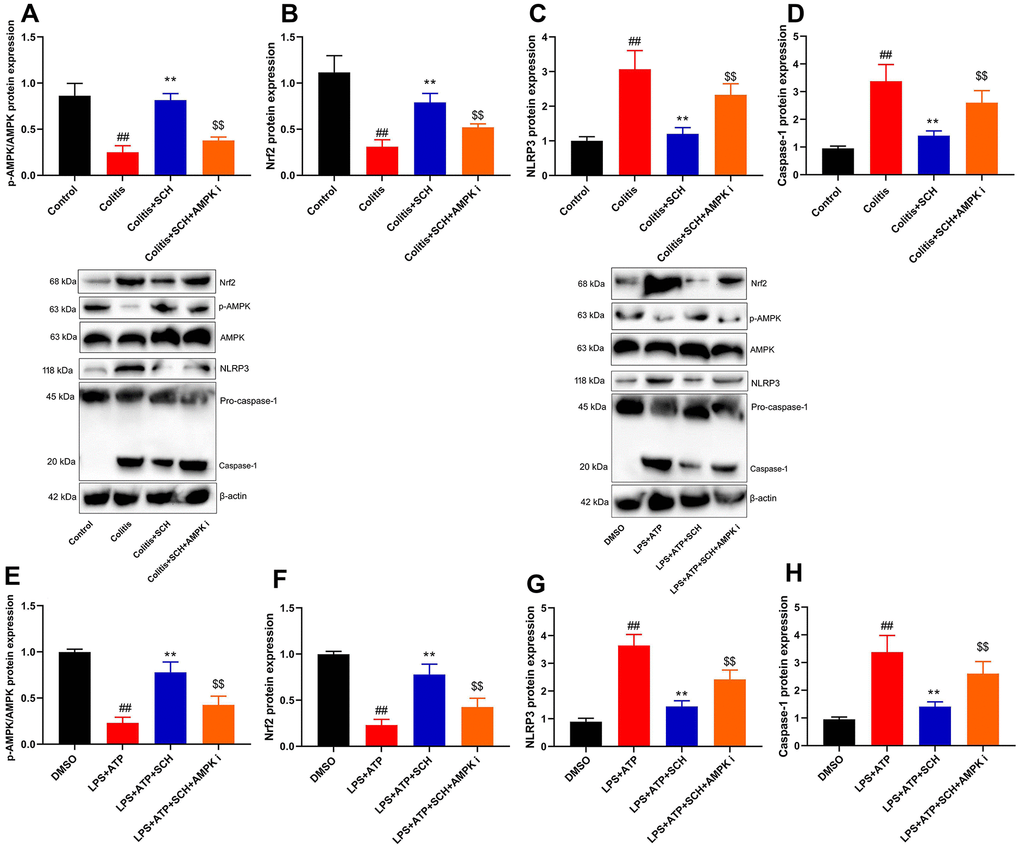
Figure 6. The inhibition of AMPK reduced the anti-inflammation effects of Schisandrin B on NLRP3 inflammasome. (A–D) p-AMPK, Nrf2, NLRP3 and Caspase-1 protein expressions in mouse colon tissue; (E–H) p-AMPK, Nrf2, NLRP3 and Caspase-1 protein expressions in intestinal epithelial cells induced by LPS + ATP. ##P<0.01 vs control group; **P<0.01 vs DSS- induced colitis group; $$P<0.01 vs Colitis+SCH group. Control: blank control group; Colitis: DSS- induced colitis group; Colitis+SCH: DSS- induced colitis mice with Schisandrin; Colitis+SCH+ AMPK i: DSS- induced colitis mice with Schisandrin and AMPK inhibitor. ##P<0.01 vs MDSO group; **P<0.01 vs LPS+ATP induced intestinal epithelial cells group; $$P<0.01 vs LPS+ATP+SCH group. MDSO: blank control group; LPS+ATP: intestinal epithelial cells with LPS+ATP group; LPS+ATP +SCH: intestinal epithelial cells induced by LPS+ATP with Schisandrin; LPS+ATP+SCH+AMPK i: intestinal epithelial cells induced by LPS+ATP, Schisandrin and AMPK inhibitor. SCH, Schisandrin B. Data were expressed as mean ± SEM.
Schisandrin B reduced ROS-induced mitochondrial damage
It is believed that ROS-induced mitochondrial damage is a common trigger of various NLRP3-activating agents. Thus, we analyzed whether Schisandrin B suppressed ROS-induced mitochondrial damage in intestinal epithelial cells by LPS+ATP. Obviously, LPS+ATP caused JC-1 aggregates and ROS production levels to increase in intestinal epithelial cells (Figure 7A, 7B). At the same time, Schisandrin B reduced JC-1 aggregates and ROS production levels in intestinal epithelial cells by LPS+ATP (Figure 8A, 8B). AMPK inhibitor (Dorsomorphin, 10 μM) increased JC-1 aggregates and ROS production levels in intestinal epithelial cells by LPS+ATP and Schisandrin B, in comparison to the group treated with Schisandrin B (Figure 7A, 7B). According to immunofluorescence, LPS+ATP enhanced ROS production level (green) and activated mitochondrial damage (red) in intestinal epithelial cells. ROS coincided with mitochondrial damage in intestinal epithelial cells (Figure 8C). In addition, Schisandrin B inhibited ROS production level (green) and reduced mitochondrial damage (red) in intestinal epithelial cells by LPS+ATP (Figure 8C). AMPK inhibitor (Dorsomorphin, 10 μM) promoted ROS production level (green) and mitochondrial damage (red) in intestinal epithelial cells by LPS+ATP and Schisandrin B, in comparison to the group treated with Schisandrin B (Figure 8C). More importantly, electron microscope indicated that LPS+ATP induced mitochondrial damage in intestinal epithelial cells (Figure 8D). Schisandrin B restored mitochondrial damage in intestinal epithelial cells by LPS+ATP (Figure 8D). AMPK inhibitor (Dorsomorphin, 10 μM) amplified mitochondrial damage in intestinal epithelial cells LPS+ATP and Schisandrin B, in comparison to the group treated with Schisandrin B (Figure 8D). Taken together, these results indicated that ROS-induced mitochondrial damage might be a dominant mechanism of underlying activation of the NLRP3 inflammasome in colitis by treatment with Schisandrin B through AMPK/Nrf2 signaling pathway.
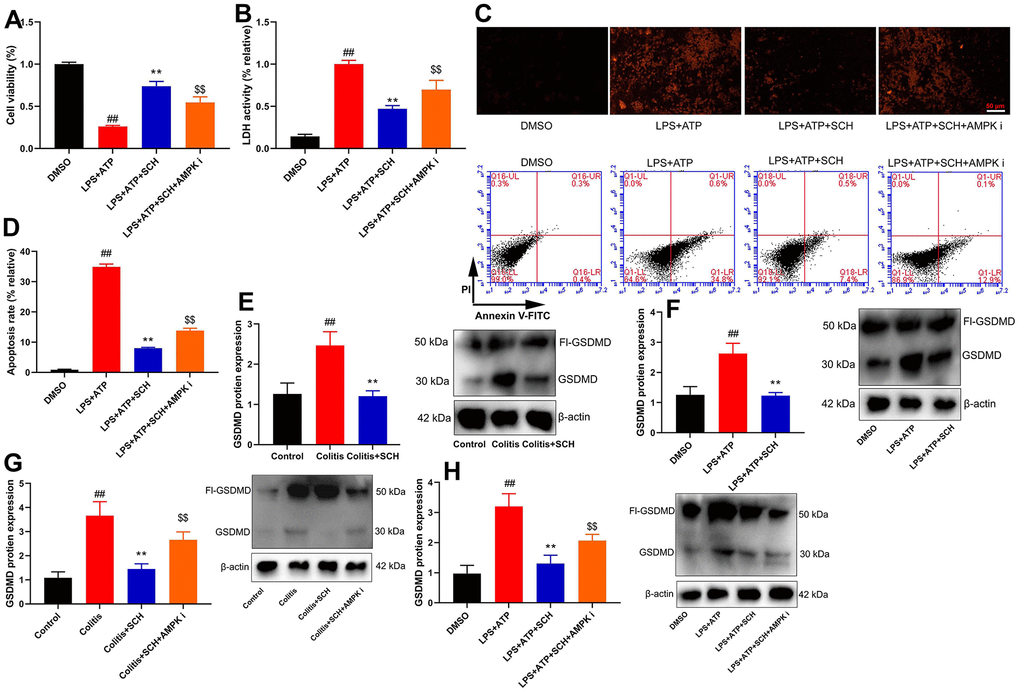
Figure 7. Schisandrin B reduced epithelial cells injury of colitis through regulating pyroptosis. (A) cell proliferation, (B) LDH activity, (C) PI staining, (D) cell apoptosis rate in epithelial cells induced by LPS + ATP; (E) GSDMD protein expression in mouse colon tissue; (F) GSDMD protein expression in epithelial cells induced by LPS + ATP; (G) GSDMD protein expression in mouse colon tissue by Schisandrin B and AMPK inhibitor; (H) GSDMD protein expression in epithelial cells induced by LPS + ATP, Schisandrin B and AMPK inhibitor. ##P<0.01 vs control group; **P<0.01 vs DSS- induced colitis group; $$P<0.01 vs Colitis+SCH group. Control: blank control group; Colitis: DSS- induced colitis group; Colitis+SCH: DSS- induced colitis mice with Schisandrin; Colitis+SCH+ AMPK i: DSS- induced colitis mice with Schisandrin and AMPK inhibitor. ##P<0.01 vs MDSO group; **P<0.01 vs LPS+ATP induced intestinal epithelial cells group; $$P<0.01 vs LPS+ATP+SCH group. MDSO: blank control group; LPS+ATP: intestinal epithelial cells with LPS+ATP group; LPS+ATP +SCH: intestinal epithelial cells induced by LPS+ATP with Schisandrin; LPS+ATP+SCH+AMPK i: intestinal epithelial cells induced by LPS+ATP, Schisandrin and AMPK inhibitor. SCH, Schisandrin B. Data were expressed as mean ± SEM.
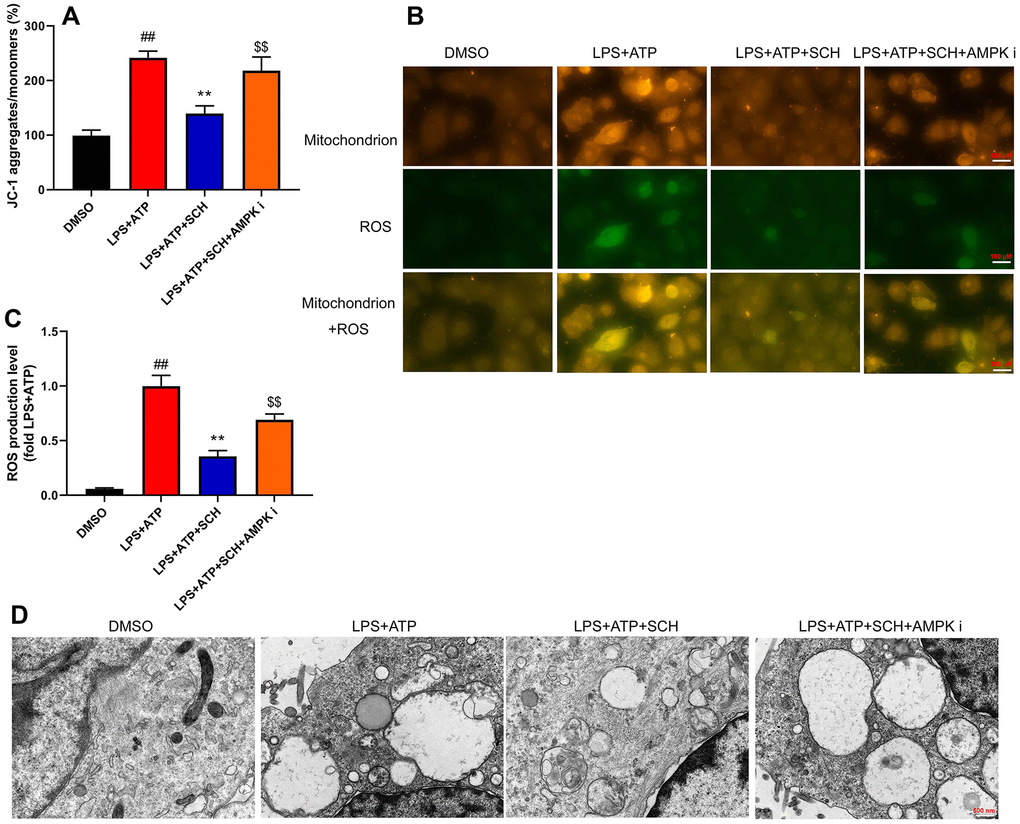
Figure 8. Schisandrin B reduced ROS-induced mitochondrial damage. (A) Methods: the total number of JC-1 in intestinal epithelial cells induced by LPS + ATP; (B) the level of ROS in intestinal epithelial cells induced by LPS + ATP; (C) mitochondria and ROS in intestinal epithelial cells by immunofluorescence; (D) mitochondria in intestinal epithelial cells by electron microscope. ##P<0.01 vs MDSO group; **P<0.01 vs LPS+ATP induced intestinal epithelial cells group; $$P<0.01 vs LPS+ATP+SCH group. MDSO: blank control group; LPS+ATP: intestinal epithelial cells with LPS+ATP group; LPS+ATP +SCH: intestinal epithelial cells induced by LPS+ATP with Schisandrin; LPS+ATP+SCH+AMPK i: intestinal epithelial cells induced by LPS+ATP, Schisandrin and AMPK inhibitor. SCH, Schisandrin B. Data were expressed as mean ± SEM.
Schisandrin B reduced epithelial cells damage of colitis through regulating pyroptosis
Pyroptosis is an important factor for intestinal cell damage or intestinal cell deaths to lead the occurrence and development of diseases of colitis. Cell viability was reduced, and LDH activity level and cell apoptosis were increased in intestinal epithelial cells by LPS+ATP (Figure 7A–7D). Schisandrin B not only increased cell viability but also reduced LDH activity level and cell apoptosis in intestinal epithelial cells by LPS+ATP (Figure 7A–7D). AMPK inhibitor (Dorsomorphin, 10 μM) decreased cell viability and increased LDH activity level and cell apoptosis in intestinal epithelial cells by LPS+ATP and Schisandrin B, in comparison to the group treated with Schisandrin B (Figure 7A–7D). Figure 7D showed that AMPK inhibitor (Dorsomorphin, 10 μM) induced early apoptosis of intestinal epithelial cells by LPS+ATP and Schisandrin B, in comparison to the group treated with Schisandrin B. GSDMD protein expression was induced in intestinal epithelial cells by LPS+ATP (Figure 7E), and Schisandrin B suppressed GSDMD protein expression in intestinal epithelial cells by LPS+ATP (Figure 7E). GSDMD protein expression in colon tissue of DSS induced colitis mice model is shown in Figure 7F. Administration of Schisandrin B suppressed GSDMD protein expression in colon tissue of DSS induced colitis mice model (Figure 7F). AMPK inhibitor (Dorsomorphin, 10 mg/kg) induced GSDMD protein expression in colon tissue of colitis mice treated with Schisandrin B more significantly in comparison to Schisandrin B group (Figure 7G). AMPK inhibitor (Dorsomorphin, 10 μM) induced GSDMD protein expression in intestinal epithelial cells treated by LPS+ATP and Schisandrin B, in comparison to Schisandrin B group (Figure 7H). Collectively, these data indicated that Schisandrin B reduced epithelial cells damage of colitis through regulating pyroptosis via AMPK/Nrf2 signaling pathway.
Discussion
UC is a chronic inflammatory disease with increasing incidence and prevalence in many countries [36]. It has been determined that the onset of UC is associated with multiple genetic and environmental factors [37]. Recent evidence indicates that intestinal permeability plays a vital role in host defense [38]. UC patients show increased intestinal mucosal permeability, which is associated with disease severity [39]. The disorder of the intestinal epithelial barrier plays an important role in the inflammatory process [40]. Therefore, it is necessary to clarify the potential mechanism of intestinal epithelial barrier damage induced by UC inflammation to identity novel therapeutic approaches [41]. In this study, we found that Schisandrin B presented colitis and reduced inflammation in vivo and vitro model of colitis. Li et al. revealed Schisandrin B prevents ulcerative colitis via gut microbiota in an in vivo and in vitro model [42]. Importantly, we identified that Schisandrin B may be a possible drug for the treatment of colitis.
UC is an autoimmune disease caused by excessive activation of the immune system. DSS-induced colitis is commonly used as the animal model for UC [43]. DSS can destroy the integrity of the intestinal epithelial barrier, increase epithelial permeability and cause the entrance of intestinal mucosal antigens and microorganisms into the mucosa, leading to the infiltration of immune cells (macrophages and neutrophils) in the lamina propria and submucosa [44, 45]. In addition, DSS can cause intense inflammation and the release of many pro-inflammatory cytokines [44]. We found that Schisandrin B suppressed NLRP3 inflammasone in vivo and vitro model of colitis. Chen et al. showed that Schisandrin B attenuates inflammation in asthma by inhibiting NLRP3 inflammasome activation [42]. Thus, our findings showed that NLRP3 inflammasone, a target spot of Schisandrin B on colitis to reduce inflammation factor of intestinal epithelial cells.
AMPK is mainly involved in regulating sugar, lipid and energy metabolism in the body [46]. Studies have demonstrated that AMPK activation can inhibit inflammation and oxidative stress [47, 48]. Nrf2 is a key molecule for the transcriptional regulation of antioxidant factors in the body [49]. The Nrf2 /HO-1 signaling pathway plays an important role in maintaining the antioxidant response of the body. Moreover, multiple studies have revealed that Nrf2 can be activated by AMPK [50]. The Nrf2 system can interact with NLRP3 inflammasome [51]. Moreover, the Nrf2 system can reduced the expression of downstream genes of the NLRP3 inflammasome, and the activation of NLRP3 inflammasome can also regulate the Nrf2 system [52]. The interaction between Nrf2 system and NLRP3 inflammasome is associated with acute and chronic inflammation, oxidative stress and autophagy [53]. In this study, we identified that Schisandrin B induced AMPK/Nrf2 signaling pathway and reduced ROS-induced mitochondrial damage in model of colitis. The inhibition of AMPK reduced the anti-inflammation effects of Schisandrin B on NLRP3 inflammasome. Zhao et al. suggested that Schisandrin B attenuates hypoxia/reoxygenation injury by activating the AMPK/Nrf2 signaling pathway [54]. Thus, Schisandrin B induced AMPK/Nrf2 signaling pathway to suppress NLRP3 inflammasome in model of colitis by the inhibition of ROS-induced mitochondrial damage.
NLRP3 is expressed by a variety of cells, mainly hematopoietic cells [17]. However, the expression of NLRP3 is also detected in glandular epithelial structures of the internal layer including the small intestine, stomach, and colon [17]. Pyroptosis is a novel type of pro-inflammatory programmed cell death, which is different from necrosis and apoptosis and is closely associated with the activation of NLRP3 inflammasome [11]. After stimulation, NLRP3 binds to ASC to recruit Caspase-1 to form an inflammasome complex [20]. Subsequently, it is cleaved to form active IL-1β. Caspase-1 also cleaves Gasdermin D to generate N-terminal fragments (GSDMD-N) and forms membrane pores to trigger the release of IL-1β [18]. Accumulative evidence suggests that the activation of NLRP3 inflammasomes and pyroptosis are associated with the inflammatory process of colitis [55]. Moreover, we also noticed that Schisandrin B reduced epithelial cells injury of colitis through regulating pyroptosis by AMPK/Nrf2/NLRP3 inflammasome. Guo et al. showed that Schisandrin B suppressed NLRP3 inflammasome activation-mediated IL-1β level and pyroptosis [30]. Thus, we conclude that Schisandrin B suppressed NLRP3 inflammasome activation-mediated IL-1β level and pyroptosis in intestinal epithelial cells of colitis model by the activation of AMPK/Nrf2 dependent signaling- ROS-induced mitochondrial damage.
In summary, our findings for first time indicated that Schisandrin B reduces the epithelial cells injury of colitis through regulating pyroptosis by AMPK/Nrf2/NLRP3 inflammasome (Figure 9). Our study also provides evidence that Schisandrin B may be of significant therapeutic approach in the treatment of acute colitis.
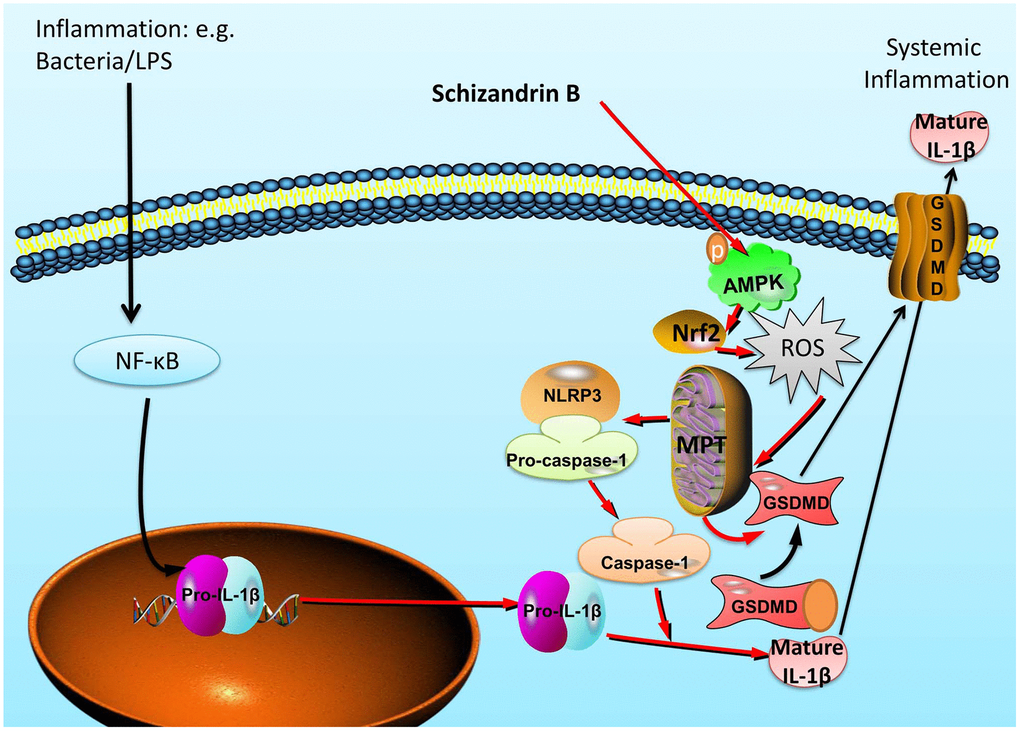
Figure 9. Schisandrin B reduces the epithelial cells injury of colitis through regulating pyroptosis by AMPK/Nrf2/NLRP3 inflammasome.
Author Contributions
Weiwei Zhang: Conceptualization, Methodology, Investigation, Formal analysis, Writing-original draft, Visualization, Project administration. Chaozhuang shen: Conceptualization, Writing - original draft, Visualization. Xiaohu Wang: Methodology, Investigation, Formal analysis. Wusan Wang: Methodology, Visualization. Qin Yin: Writing - review and editing, Supervision, Funding acquisition.
Weiwei Zhang, Wusan Wang, Chaozhuang shen and Xiaohu Wang conducted the experiments and data analysis, involved in preparation of the figures and manuscript. Zhichen Pu and Qin Yin designed the study and prepared the manuscript. All authors have read the manuscript and agreed to the conclusion.
Acknowledgments
The authors thank for central laboratory of Yijishan Hospital of Wannan Medical College.
Conflicts of Interest
The authors declared that there is no conflicts of interest.
Funding
This work was supported by National Natural Science Foundation of China (81173134); Nature Science Research Project of Anhui province (2108085QH384); Talent Introduction Program of Yijishan Hospital of Wannan Medical College (YR202005), Science and Technology Innovation Team of Yijishan Hospital of Wannan Medical College (YPF2019016) and Key project research fund of Wannan Medical College (WK2021F03, WK2021F37, WK2020ZF03 and WK2020F23). Thanks to the central laboratory for its support to this research.
Editorial Note
This corresponding author has a verified history of publications using a personal email address for correspondence
References
- 1. Delisle M, Wilkens R, Novak KL. Keeping It Simple: Using Intestinal Ultrasound to Assess Ulcerative Colitis with a Novel UC-Ultrasound Index. J Crohns Colitis. 2021; 15:1250–52. https://doi.org/10.1093/ecco-jcc/jjab066 [PubMed]
- 2. Kasuga K, Yamada M, Shida D, Tagawa T, Takamaru H, Sekiguchi M, Sakamoto T, Uraoka T, Sekine S, Kanemitsu Y, Saito Y. Treatment outcomes of endoscopic submucosal dissection and surgery for colorectal neoplasms in patients with ulcerative colitis. United European Gastroenterol J. 2021. [Epub ahead of print]. https://doi.org/10.1002/ueg2.12118 [PubMed]
- 3. Lechner K, Mott S, Al-Saifi R, Knipfer L, Wirtz S, Atreya R, Vieth M, Rath T, Fraass T, Winter Z, August A, Luban J, Zimmermann VS, et al. Targeting of the Tec Kinase ITK Drives Resolution of T Cell-Mediated Colitis and Emerges as Potential Therapeutic Option in Ulcerative Colitis. Gastroenterology. 2021; 161:1270–87.e19. https://doi.org/10.1053/j.gastro.2021.06.072 [PubMed]
- 4. Liu C, Hua H, Zhu H, Cheng Y, Guo Y, Yao W, Qian H. Aloe polysaccharides ameliorate acute colitis in mice via Nrf2/HO-1 signaling pathway and short-chain fatty acids metabolism. Int J Biol Macromol. 2021; 185:804–12. https://doi.org/10.1016/j.ijbiomac.2021.07.007 [PubMed]
- 5. da Silva BC, Lyra AC, Rocha R, Santana GO. Epidemiology, demographic characteristics and prognostic predictors of ulcerative colitis. World J Gastroenterol. 2014; 20:9458–67. https://doi.org/10.3748/wjg.v20.i28.9458 [PubMed]
- 6. Siow VS, Bhatt R, Mollen KP. Management of acute severe ulcerative colitis in children. Semin Pediatr Surg. 2017; 26:367–72. https://doi.org/10.1053/j.sempedsurg.2017.10.006 [PubMed]
- 7. Yamamoto-Furusho JK, Gutiérrez-Grobe Y, López-Gómez JG, Bosques-Padilla F, Rocha-Ramírez JL, and Grupo del Consenso Mexicano de Colitis Ulcerosa Crónica Idiopática. The Mexican consensus on the diagnosis and treatment of ulcerative colitis. Rev Gastroenterol Mex (Engl Ed). 2018; 83:144–67. https://doi.org/10.1016/j.rgmx.2017.08.006 [PubMed]
- 8. Bohl JL, Sobba K. Indications and Options for Surgery in Ulcerative Colitis. Surg Clin North Am. 2015; 95:1211–32, vi. https://doi.org/10.1016/j.suc.2015.07.003 [PubMed]
- 9. Gao Q, Bi P, Luo D, Guan Y, Zeng W, Xiang H, Mi Q, Yang G, Li X, Yang B. Nicotine-induced autophagy via AMPK/mTOR pathway exerts protective effect in colitis mouse model. Chem Biol Interact. 2020; 317:108943. https://doi.org/10.1016/j.cbi.2020.108943 [PubMed]
- 10. Gurung P, Dahal S, Chaudhary P, Guragain D, Karmacharya U, Kim JA, Jeong BS. Potent Inhibitory Effect of BJ-3105, a 6-Alkoxypyridin-3-ol Derivative, on Murine Colitis Is Mediated by Activating AMPK and Inhibiting NOX. Int J Mol Sci. 2020; 21:3145. https://doi.org/10.3390/ijms21093145 [PubMed]
- 11. Deng Z, Ni J, Wu X, Wei H, Peng J. GPA peptide inhibits NLRP3 inflammasome activation to ameliorate colitis through AMPK pathway. Aging (Albany NY). 2020; 12:18522–44. https://doi.org/10.18632/aging.103825 [PubMed]
- 12. Deng Z, Rong Y, Teng Y, Mu J, Zhuang X, Tseng M, Samykutty A, Zhang L, Yan J, Miller D, Suttles J, Zhang HG. Broccoli-Derived Nanoparticle Inhibits Mouse Colitis by Activating Dendritic Cell AMP-Activated Protein Kinase. Mol Ther. 2017; 25:1641–54. https://doi.org/10.1016/j.ymthe.2017.01.025 [PubMed]
- 13. Pu Z, Liu Y, Li C, Xu M, Xie H, Zhao J. Using Network Pharmacology for Systematic Understanding of Geniposide in Ameliorating Inflammatory Responses in Colitis Through Suppression of NLRP3 Inflammasome in Macrophage by AMPK/Sirt1 Dependent Signaling. Am J Chin Med. 2020; 48:1693–713. https://doi.org/10.1142/S0192415X20500846 [PubMed]
- 14. Youm YH, Nguyen KY, Grant RW, Goldberg EL, Bodogai M, Kim D, D’Agostino D, Planavsky N, Lupfer C, Kanneganti TD, Kang S, Horvath TL, Fahmy TM, et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat Med. 2015; 21:263–69. https://doi.org/10.1038/nm.3804 [PubMed]
- 15. Yang F, Qin Y, Wang Y, Meng S, Xian H, Che H, Lv J, Li Y, Yu Y, Bai Y, Wang L. Metformin Inhibits the NLRP3 Inflammasome via AMPK/mTOR-dependent Effects in Diabetic Cardiomyopathy. Int J Biol Sci. 2019; 15:1010–19. https://doi.org/10.7150/ijbs.29680 [PubMed]
- 16. Qing L, Fu J, Wu P, Zhou Z, Yu F, Tang J. Metformin induces the M2 macrophage polarization to accelerate the wound healing via regulating AMPK/mTOR/NLRP3 inflammasome singling pathway. Am J Transl Res. 2019; 11:655–68. [PubMed]
- 17. Chao L, Li Z, Zhou J, Chen W, Li Y, Lv W, Guo A, Qu Q, Guo S. Shen-Ling-Bai-Zhu-San Improves Dextran Sodium Sulfate-Induced Colitis by Inhibiting Caspase-1/Caspase-11-Mediated Pyroptosis. Front Pharmacol. 2020; 11:814. https://doi.org/10.3389/fphar.2020.00814 [PubMed]
- 18. Wu X, Pan S, Luo W, Shen Z, Meng X, Xiao M, Tan B, Nie K, Tong T, Wang X. Roseburia intestinalis-derived flagellin ameliorates colitis by targeting miR-223-3p-mediated activation of NLRP3 inflammasome and pyroptosis. Mol Med Rep. 2020; 22:2695–704. https://doi.org/10.3892/mmr.2020.11351 [PubMed]
- 19. Zhang X, Chen Y, Yu S, Jin B, Liu W. Inhibition of C3a/C3aR Axis in Diverse Stages of Ulcerative Colitis Affected the Prognosis of UC by Modulating the Pyroptosis and Expression of Caspase-11. Inflammation. 2020; 43:2128–36. https://doi.org/10.1007/s10753-020-01280-3 [PubMed]
- 20. Jie F, Xiao S, Qiao Y, You Y, Feng Y, Long Y, Li S, Wu Y, Li Y, Du Q. Kuijieling decoction suppresses NLRP3-Mediated pyroptosis to alleviate inflammation and experimental colitis in vivo and in vitro. J Ethnopharmacol. 2021; 264:113243. https://doi.org/10.1016/j.jep.2020.113243 [PubMed]
- 21. Zhang JM, Yu RQ, Wu FZ, Qiao L, Wu XR, Fu YJ, Liang YF, Pang Y, Xie CY. BMP-2 alleviates heart failure with type 2 diabetes mellitus and doxorubicin-induced AC16 cell injury by inhibiting NLRP3 inflammasome-mediated pyroptosis. Exp Ther Med. 2021; 22:897. https://doi.org/10.3892/etm.2021.10329 [PubMed]
- 22. Zhang X, Zhao D, Feng J, Yang X, Lan Z, Yang T, Kong X, Qu H, Zhou H. LuQi Formula Regulates NLRP3 Inflammasome to Relieve Myocardial-Infarction-Induced Cardiac Remodeling in Mice. Evid Based Complement Alternat Med. 2021; 2021:5518083. https://doi.org/10.1155/2021/5518083 [PubMed]
- 23. Pu Z, Han C, Zhang W, Xu M, Wu Z, Liu Y, Wu M, Sun H, Xie H. Systematic understanding of the mechanism and effects of Arctigenin attenuates inflammation in dextran sulfate sodium-induced acute colitis through suppression of NLRP3 inflammasome by SIRT1. Am J Transl Res. 2019; 11:3992–4009. [PubMed]
- 24. Lowes DJ, Miao J, Al-Waqfi RA, Avad KA, Hevener KE, Peters BM. Identification of Dual-Target Compounds with Antifungal and Anti-NLRP3 Inflammasome Activity. ACS Infect Dis. 2021; 7:2522–35. https://doi.org/10.1021/acsinfecdis.1c00270 [PubMed]
- 25. Cai NN, Wang ZZ, Zhu XC, Jiang Y, Zhu WQ, Yang R, Zhang XM. Schisandrin A and B enhance the dentate gyrus neurogenesis in mouse hippocampus. J Chem Neuroanat. 2020; 105:101751. https://doi.org/10.1016/j.jchemneu.2020.101751 [PubMed]
- 26. Ji ZR, Xue WL, Zhang L. Schisandrin B Attenuates Inflammation in LPS-Induced Sepsis Through miR-17-5p Downregulating TLR4. Inflammation. 2019; 42:731–39. https://doi.org/10.1007/s10753-018-0931-3 [PubMed]
- 27. Lam HY, Liang TR, Jiang SJ, Peng SY. Albendazole-Schisandrin B Co-Therapy on Angiostrongylus cantonensis-Induced Meningoencephalitis in Mice. Biomolecules. 2020; 10:1001. https://doi.org/10.3390/biom10071001 [PubMed]
- 28. Leong PK, Ko KM. Schisandrin B: A Double-Edged Sword in Nonalcoholic Fatty Liver Disease. Oxid Med Cell Longev. 2016; 2016:6171658. https://doi.org/10.1155/2016/6171658 [PubMed]
- 29. Wang J, Fang Z, Song C, Kang H, Guo Q, Dong Y, Zhang Y, Peng R, Guan H, Li F. Schisandrin B Inhibits Osteoclastogenesis and Protects Against Ovariectomy-Induced Bone Loss. Front Pharmacol. 2020; 11:1175. https://doi.org/10.3389/fphar.2020.01175 [PubMed]
- 30. Guo M, An F, Yu H, Wei X, Hong M, Lu Y. Comparative effects of schisandrin A, B, and C on Propionibacterium acnes-induced, NLRP3 inflammasome activation-mediated IL-1β secretion and pyroptosis. Biomed Pharmacother. 2017; 96:129–36. https://doi.org/10.1016/j.biopha.2017.09.097 [PubMed]
- 31. Li YJ, Liu HT, Xue CJ, Xing XQ, Dong ST, Wang LS, Ding CY, Meng L, Dong ZJ. The synergistic anti-tumor effect of schisandrin B and apatinib. J Asian Nat Prod Res. 2020; 22:839–49. https://doi.org/10.1080/10286020.2019.1645131 [PubMed]
- 32. Guo BJ, Liu Z, Ding MY, Li F, Jing M, Xu LP, Wang YQ, Zhang ZJ, Wang Y, Wang D, Zhou GC, Wang Y. Andrographolide derivative ameliorates dextran sulfate sodium-induced experimental colitis in mice. Biochem Pharmacol. 2019; 163:416–24. https://doi.org/10.1016/j.bcp.2019.03.019 [PubMed]
- 33. Choi JS, Kim JH, Ali MY, Jung HJ, Min BS, Choi RJ, Kim GD, Jung HA. Anti-adipogenic effect of epiberberine is mediated by regulation of the Raf/MEK1/2/ERK1/2 and AMPKα/Akt pathways. Arch Pharm Res. 2015; 38:2153–62. https://doi.org/10.1007/s12272-015-0626-3 [PubMed]
- 34. Hao H, Cao L, Jiang C, Che Y, Zhang S, Takahashi S, Wang G, Gonzalez FJ. Farnesoid X Receptor Regulation of the NLRP3 Inflammasome Underlies Cholestasis-Associated Sepsis. Cell Metab. 2017; 25:856–67.e5. https://doi.org/10.1016/j.cmet.2017.03.007 [PubMed]
- 35. Xu W, Che Y, Zhang Q, Huang H, Ding C, Wang Y, Wang G, Cao L, Hao H. Apaf-1 Pyroptosome Senses Mitochondrial Permeability Transition. Cell Metab. 2021; 33:424–36.e10. https://doi.org/10.1016/j.cmet.2020.11.018 [PubMed]
- 36. Liu S, Li Q, Liu F, Cao H, Liu J, Shan J, Dan W, Yuan J, Lin J. Uncovering the Mechanism of Curcuma in the Treatment of Ulcerative Colitis Based on Network Pharmacology, Molecular Docking Technology, and Experiment Verification. Evid Based Complement Alternat Med. 2021; 2021:6629761. https://doi.org/10.1155/2021/6629761 [PubMed]
- 37. Narula N, Wong EC, Colombel JF, Riddell R, Marshall JK, Reinisch W, Dulai PS. Early Change in Epithelial Neutrophilic Infiltrate Predicts Long-Term Response to Biologics in Ulcerative Colitis. Clin Gastroenterol Hepatol. 2021; S1542-3565:00716–13. https://doi.org/10.1016/j.cgh.2021.07.005 [PubMed]
- 38. DE Angelis PM, Dorg L, Pham S, Andersen SN. DNA Repair Protein Expression and Oxidative/Nitrosative Stress in Ulcerative Colitis and Sporadic Colorectal Cancer. Anticancer Res. 2021; 41:3261–70. https://doi.org/10.21873/anticanres.15112 [PubMed]
- 39. Rotondo-Trivette S, Wang B, Gayer C, Parsana R, Luan Y, Sun F, Michail S. Decreased secondary faecal bile acids in children with ulcerative colitis and Clostridioides difficile infection. Aliment Pharmacol Ther. 2021; 54:792–804. https://doi.org/10.1111/apt.16496 [PubMed]
- 40. Roy BC, Ahmed I, Stubbs J, Zhang J, Attard T, Septer S, Welch D, Anant S, Sampath V, Umar S. DCLK1 isoforms and aberrant Notch signaling in the regulation of human and murine colitis. Cell Death Discov. 2021; 7:169. https://doi.org/10.1038/s41420-021-00526-9 [PubMed]
- 41. Singh S, Heien HC, Sangaralingham L, Shah ND, Sandborn WJ. Obesity Is Not Associated With an Increased Risk of Serious Infections in Biologic-Treated Patients With Inflammatory Bowel Diseases. Clin Transl Gastroenterol. 2021; 12:e00380. https://doi.org/10.14309/ctg.0000000000000380 [PubMed]
- 42. Li J, Lu Y, Wang D, Quan F, Chen X, Sun R, Zhao S, Yang Z, Tao W, Ding D, Gao X, Cao Q, Zhao D, et al. Schisandrin B prevents ulcerative colitis and colitis-associated-cancer by activating focal adhesion kinase and influence on gut microbiota in an in vivo and in vitro model. Eur J Pharmacol. 2019; 854:9–21. https://doi.org/10.1016/j.ejphar.2019.03.059 [PubMed]
- 43. Panpetch W, Hiengrach P, Nilgate S, Tumwasorn S, Somboonna N, Wilantho A, Chatthanathon P, Prueksapanich P, Leelahavanichkul A. Additional Candida albicans administration enhances the severity of dextran sulfate solution induced colitis mouse model through leaky gut-enhanced systemic inflammation and gut-dysbiosis but attenuated by Lactobacillus rhamnosus L34. Gut Microbes. 2020; 11:465–80. https://doi.org/10.1080/19490976.2019.1662712 [PubMed]
- 44. Macias-Ceja DC, Cosín-Roger J, Ortiz-Masiá D, Salvador P, Hernández C, Esplugues JV, Calatayud S, Barrachina MD. Stimulation of autophagy prevents intestinal mucosal inflammation and ameliorates murine colitis. Br J Pharmacol. 2017; 174:2501–11. https://doi.org/10.1111/bph.13860 [PubMed]
- 45. Yang H, Wang W, Romano KA, Gu M, Sanidad KZ, Kim D, Yang J, Schmidt B, Panigrahy D, Pei R, Martin DA, Ozay EI, Wang Y, et al. A common antimicrobial additive increases colonic inflammation and colitis-associated colon tumorigenesis in mice. Sci Transl Med. 2018; 10:eaan4116. https://doi.org/10.1126/scitranslmed.aan4116 [PubMed]
- 46. Xu X, Wang Y, Wei Z, Wei W, Zhao P, Tong B, Xia Y, Dai Y. Madecassic acid, the contributor to the anti-colitis effect of madecassoside, enhances the shift of Th17 toward Treg cells via the PPARγ/AMPK/ACC1 pathway. Cell Death Dis. 2017; 8:e2723. https://doi.org/10.1038/cddis.2017.150 [PubMed]
- 47. Zhou K, Cheng R, Liu B, Wang L, Xie H, Zhang C. Eupatilin ameliorates dextran sulphate sodium-induced colitis in mice partly through promoting AMPK activation. Phytomedicine. 2018; 46:46–56. https://doi.org/10.1016/j.phymed.2018.04.033 [PubMed]
- 48. Takahara M, Takaki A, Hiraoka S, Adachi T, Shimomura Y, Matsushita H, Nguyen TT, Koike K, Ikeda A, Takashima S, Yamasaki Y, Inokuchi T, Kinugasa H, et al. Berberine improved experimental chronic colitis by regulating interferon-γ- and IL-17A-producing lamina propria CD4+ T cells through AMPK activation. Sci Rep. 2019; 9:11934. https://doi.org/10.1038/s41598-019-48331-w [PubMed]
- 49. Wang R, Luo Y, Lu Y, Wang D, Wang T, Pu W, Wang Y. Maggot Extracts Alleviate Inflammation and Oxidative Stress in Acute Experimental Colitis via the Activation of Nrf2. Oxid Med Cell Longev. 2019; 2019:4703253. https://doi.org/10.1155/2019/4703253 [PubMed]
- 50. Qiu S, Li P, Zhao H, Li X. Maresin 1 alleviates dextran sulfate sodium-induced ulcerative colitis by regulating NRF2 and TLR4/NF-kB signaling pathway. Int Immunopharmacol. 2020; 78:106018. https://doi.org/10.1016/j.intimp.2019.106018 [PubMed]
- 51. Bai X, Gou X, Cai P, Xu C, Cao L, Zhao Z, Huang M, Jin J. Sesamin Enhances Nrf2-Mediated Protective Defense against Oxidative Stress and Inflammation in Colitis via AKT and ERK Activation. Oxid Med Cell Longev. 2019; 2019:2432416. https://doi.org/10.1155/2019/2432416 [PubMed]
- 52. Fernández-Ortiz M, Sayed RK, Fernández-Martínez J, Cionfrini A, Aranda-Martínez P, Escames G, de Haro T, Acuña-Castroviejo D. Melatonin/Nrf2/NLRP3 Connection in Mouse Heart Mitochondria during Aging. Antioxidants (Basel). 2020; 9:1187. https://doi.org/10.3390/antiox9121187 [PubMed]
- 53. Hou Y, Wang Y, He Q, Li L, Xie H, Zhao Y, Zhao J. Nrf2 inhibits NLRP3 inflammasome activation through regulating Trx1/TXNIP complex in cerebral ischemia reperfusion injury. Behav Brain Res. 2018; 336:32–39. https://doi.org/10.1016/j.bbr.2017.06.027 [PubMed]
- 54. Zhao B, Li GP, Peng JJ, Ren LH, Lei LC, Ye HM, Wang ZY, Zhao S. Schisandrin B attenuates hypoxia/reoxygenation injury in H9c2 cells by activating the AMPK/Nrf2 signaling pathway. Exp Ther Med. 2021; 21:220. https://doi.org/10.3892/etm.2021.9651 [PubMed]
- 55. Saber S, Abd El-Kader EM, Sharaf H, El-Shamy R, El-Saeed B, Mostafa A, Ezzat D, Shata A. Celastrol augments sensitivity of NLRP3 to CP-456773 by modulating HSP-90 and inducing autophagy in dextran sodium sulphate-induced colitis in rats. Toxicol Appl Pharmacol. 2020; 400:115075. https://doi.org/10.1016/j.taap.2020.115075 [PubMed]