



Introduction
Rheumatoid arthritis (RA) is an autoimmune disease characterized by synovial inflammation and hyperplasia, pannus formation, and erosion of adjacent articular cartilage and bone leading to joint dysfunction [1, 2]. Although its pathogenesis remains unclear, an increasing number of studies indicate that fibroblast-like synoviocytes (FLS) are a key component of invasive pannus and contribute to the development of RA [3, 4]. In RA, activated FLS invade the extracellular matrix and secrete proinflammatory factors [5], matrix metalloproteinases (MMPs) [6, 7], chemokines [8], and cell adhesion molecules [9], which destroy articular cartilage and bone and exacerbate joint dysfunction. Therefore, inhibition of FLS migration and the inflammatory response could be a target in the treatment of RA.
FLS secrete several cytokines, including the proinflammatory factors TNF-α, IL-6, and IL-1β [10–12]; transforming growth factor-β (TGF-β) [13, 14]; chemokines [8]; and cell adhesion molecules [15]. TGF-β/Smad not only regulates several physiologic processes of bone development, but also is involved in inflammation and fibrosis of FLS in RA [14, 16]. Bone morphogenetic proteins (BMPs), as members of the TGF-β superfamily, are secreted signaling proteins [17]. They also contribute to the development of cartilage and bone [18]. Accumulating evidence indicates that BMPs participate in regulating the inflammatory response, migration, and invasion of FLS [14, 16, 19].
BMP3, which is an antagonist of osteogenic BMPs [20, 21], inhibits the proliferation of colorectal cancer cells [22]. Similarly, forced expression of BMP3 decreases cell proliferation and viability and increases apoptosis of biliary cancer cells [23]. More importantly, expression of BMP3 mRNA has been found to be decreased in arthritic synovium compared with nonarthritic synovium [24].
To further elucidate the relationship between BMP3 and proinflammatory cytokines and chemokines and migration of FLS in RA, we studied the effect of BMP3 expression on the formation of proinflammatory cytokines and chemokines and the migration of FLS.
Results
BMP3 expression is significantly downregulated in RA
To explore the role of BMP3 in RA, we collected synovial tissues from patients with osteoarthritis (OA) and RA. We then performed histopathologic analysis (Figure 1A), and the results indicated that inflammatory cell infiltration and synovial hyperplasia were significantly increased in RA synovial tissues. In addition, immunohistochemical analysis (Figure 1B) demonstrated that BMP3 expression was significantly downregulated in RA synovial tissues compared to expression in OA synovial tissues. Immunofluorescence staining also verified that BMP3 expression was notably decreased in RA synovial tissues (Figure 1C) and in RA FLS treated with TNF-α (Figure 1D). Western blot results also demonstrated that BMP3 expression was significantly downregulated in RA synovial tissues (Figure 1E). Conversely, expression of IL-6, IL-1β, IL-17A, and TNF-α was significantly increased in RA synovial tissues. Importantly, after RA FLS were treated with various inflammatory factors in vitro, such as IL-6, IL-1β, IL-17A, TNF-α, IFN-γ, and LPS, western blot results showed that BMP3 expression was decreased to different degrees (Figure 1F). Thus, these results indicate that BMP3 expression is significantly decreased in RA synovial tissues and FLS.
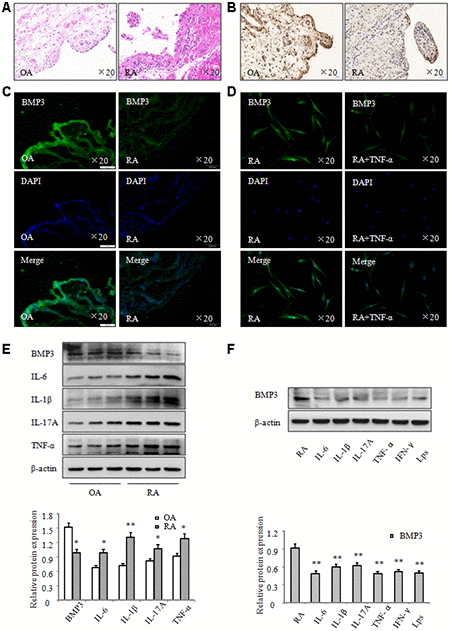
Figure 1. BMP3 expression was significantly downregulated in RA. (A) Representative H&E staining of OA and RA synovial tissues (original magnification, ×20). (B) BMP3 expression in OA and RA synovial tissues was analyzed using IHC staining (original magnification, ×20). (C) BMP3 expression in OA and RA synovial tissues was analyzed using immunofluorescence staining (original magnification, ×20). (D) BMP3 expression in RA FLS treated with TNF-α was analyzed using immunofluorescence staining (original magnification, ×10). (E) The protein levels of BMP3 in OA and RA synovial tissues were analyzed using western blot. (F) The protein levels of BMP3 in RA FLS treated with various inflammation factors were analyzed using western blot. All values are expressed as the mean ± SD. *P < 0.05, **P < 0.01 vs control group.
BMP3 expression is significantly downregulated in AIA
To further investigate the role of BMP3 in RA, we also created an AIA model using injection with the complete Freund’s adjuvant. Histopathologic analysis (Figure 2A) confirmed that the AIA model was successfully established. Our results showed that inflammatory cell infiltration and synovial hyperplasia were significantly increased in the synovial tissue of AIA rats. Furthermore, immunohistochemical analysis (Figure 2B) showed that BMP3 expression was significantly downregulated in AIA synovial tissue. We also measured BMP3 protein expression in synovial tissues and FLS using immunofluorescence staining (Figure 2C and 2D). Our results showed that BMP3 expression was decreased in the AIA synovial tissues compared with that in normal synovial tissues (Figure 2C). In addition, BMP3 expression was lower in AIA FLS treated with TNF-α (Figure 2D). Western blot analysis also demonstrated that BMP3 expression was significantly downregulated in AIA rat synovial tissues (Figure 2E). However, the expression of IL-6, IL-1β, IL-17A, and TNF-α was concomitantly increased in AIA synovial tissues. More significantly, after the AIA FLS were treated with various inflammation factors in vitro, such as IL-6, IL-1β, IL-17A, TNF-α, IFN-γ, and LPS, western blot results demonstrated that BMP3 expression was decreased to different degrees (Figure 2F). Thus, these results indicate that BMP3 expression is reduced in AIA synovial tissues and FLS.
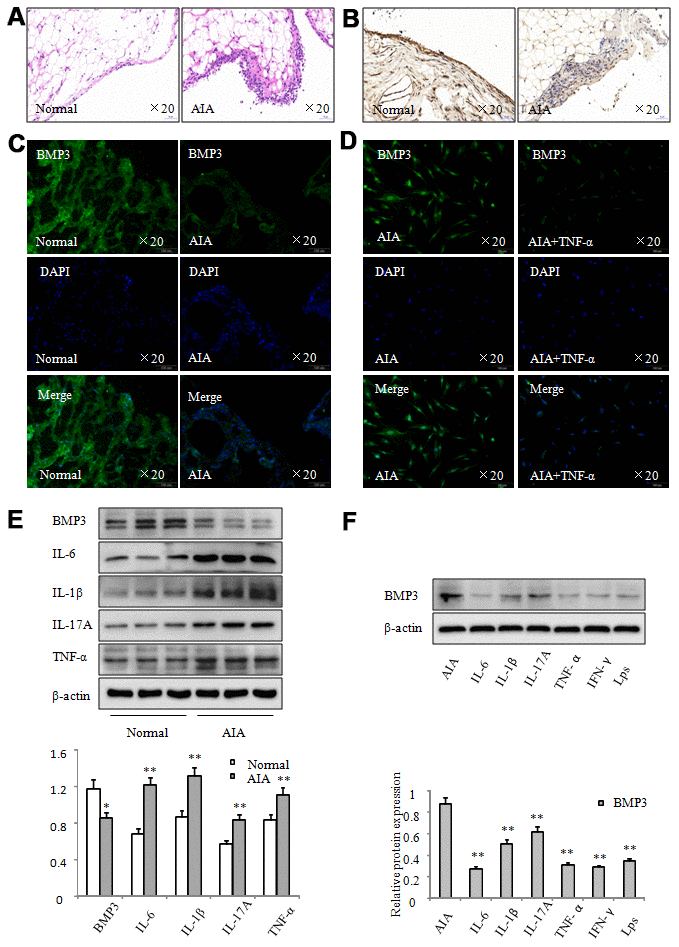
Figure 2. BMP3 expression was significantly downregulated in AIA. (A) Representative H&E staining of normal and AIA rat synovial tissues (original magnification, ×20). (B) BMP3 expression in normal and AIA rat synovial tissues was analyzed using IHC staining (original magnification, ×20). (C) BMP3 expression in normal and AIA rat synovial tissues was analyzed using immunofluorescence staining (original magnification, ×20). (D) BMP3 expression in FLS from AIA rats treated with TNF-α was analyzed using immunofluorescence staining (original magnification, ×10). (E) BMP3 protein levels in normal and AIA synovial tissues were analyzed using western blot. (F) BMP3 protein levels in AIA FLS treated with various inflammation factors were analyzed using western blot. All values are expressed as the mean ± SD. *P < 0.05, **P < 0.01 vs control group.
Inhibition of BMP3 expression increases proinflammatory cytokines and chemokines in RA and AIA FLS
To investigate the significance of BMP3 in RA FLS, specific siRNA for BMP3 was used to knock down gene expression in TNF-α–treated RA FLS. As shown in Figure 3A and 3C, western blot and quantitative polymerase chain reaction (qPCR) results showed that BMP3 expression was significantly reduced in TNF-α–treated RA FLS transfected with BMP3-RNAi compared to the cells transfected with NC-RNAi. In addition, the results of Western blot and qPCR also showed that BMP3 expression was notably reduced in TNF-α–treated AIA FLS transfected with BMP3-RNAi (Figure 3B and 3D). More significantly, after treatment with TNF-α, the results of western blot and qPCR indicated that expression of the proinflammatory cytokines IL-6, IL-1β, and IL-17A was markedly upregulated by BMP3-RNAi in RA FLS (Figure 3A and 3C). As expected, the results of western blot and qPCR indicated that the expression of the proinflammatory cytokines IL-6, IL-1β, and IL-17A was also markedly upregulated by BMP3-RNAi in TNF-α–treated AIA FLS (Figure 3B and 3D). Moreover, qPCR results showed that CCL-2, CCL-3, and VCAM-1 mRNA expression was upregulated in TNF-α–treated RA FLS after transfection with BMP3-RNAi (Figure 3E). mRNA expression of CCL-2, CCL-3, and VCAM-1 was also significantly upregulated in TNF-α–treated AIA FLS after transfection with BMP3-RNAi (Figure 3F). Thus, these results demonstrate that inhibition of BMP3 expression increases proinflammatory cytokines and chemokines in RA and AIA FLS.
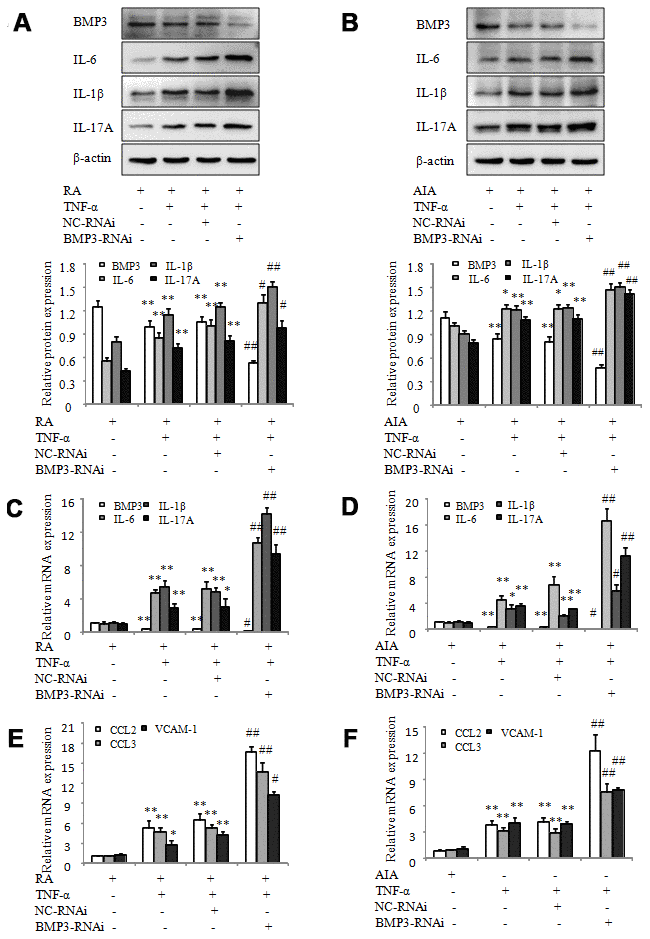
Figure 3. BMP3 siRNA silencing increases the proinflammatory cytokines and chemokines in RA and AIA FLS. (A) The protein levels of BMP3, IL-6, IL-1β, and IL-17A in TNF-α–treated RA FLS transfected with BMP3 siRNA were analyzed using western blot. (B) The protein levels of BMP3, IL-6, IL-1β, and IL-17A in TNF-α–treated AIA FLS transfected with BMP3 siRNA were analyzed using western blot. (C) The mRNA levels of BMP3, IL-6, IL-1β, and IL-17A in TNF-α–treated RA FLS transfected with BMP3 siRNA were analyzed using qPCR. (D) The mRNA levels of BMP3, IL-6, IL-1β, and IL-17A in TNF-α–treated AIA FLS transfected with BMP3 siRNA were analyzed using qPCR. (E) The mRNA levels of CCL2, CCL3, and VCAM-1 in TNF-α–treated RA FLS transfected with BMP3 siRNA were analyzed using qPCR. (F) The mRNA levels of CCL2, CCL3, and VCAM-1 in TNF-α–treated AIA FLS transfected with BMP3 siRNA were analyzed using qPCR. All values are expressed as the mean ± SD. *P < 0.05, **P < 0.01 vs AIA group. #P < 0.05, ##P < 0.01 vs NC-RNAi group.
Inhibition of BMP3 expression promotes the migration of RA and AIA FLS
Based on the previous results, we explored the effect of BMP3 on the migration of RA and AIA FLS after transfection with specific siRNA for BMP3. As shown in Figure 4A and 4C, the results of western blot and qPCR demonstrated that the protein and mRNA levels of MMP-3 and MMP-9 were higher in TNF-α–treated RA FLS transfected with BMP3-RNAi compared to the cells transfected with NC-RNAi. Conversely, the protein and mRNA levels of TIMP-1 were downregulated in RA FLS treated with TNF-α after transfection with BMP3-RNAi (Figure 4A and 4C). In addition, the results of Western blot and qPCR also showed that MMP-3 and MMP-9 expression was significantly upregulated in TNF-α–treated AIA FLS transfected with BMP3-RNAi (Figure 4B and 4D), whereas the protein and mRNA expression of TIMP-1 was downregulated in AIA FLS under the same conditions (Figure 4B and 4D). We also performed a wound-healing assay, and the results showed that the migration ability of RA FLS was significantly increased after transfection with BMP3-RNAi (Figure 4E). The migration of AIA FLS was also increased after BMP3-RNAi transfection (Figure 4F). Therefore, our findings suggest that inhibition of BMP3 expression promotes the migration of RA and AIA FLS and contributes to the progression of RA.
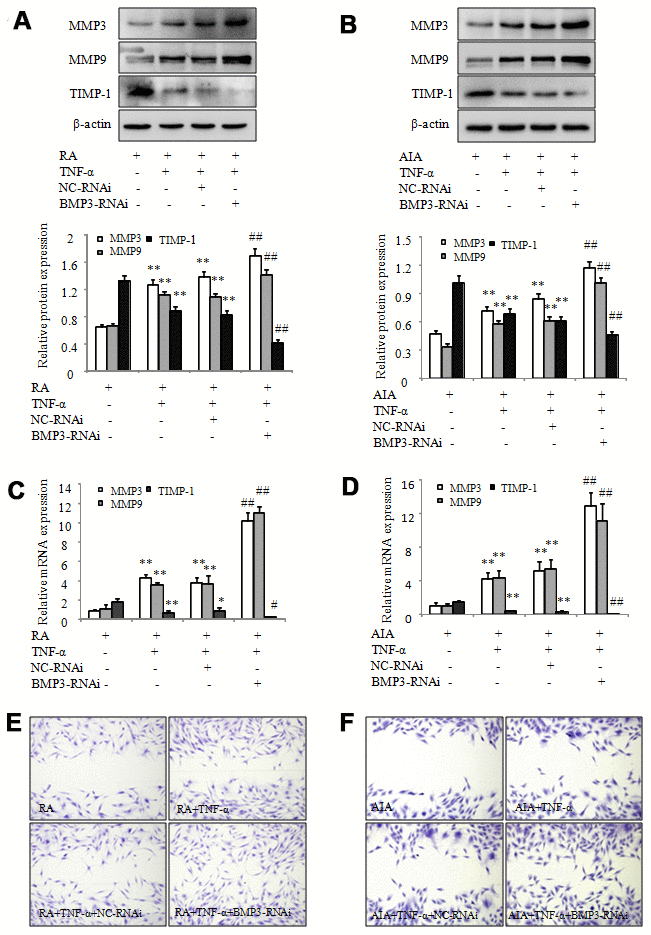
Figure 4. BMP3 siRNA silencing promotes the migration of RA and AIA FLS. (A) The protein levels of MMP-3, MMP-9, and TIMP-1 in TNF-α–treated RA FLS transfected with BMP3 siRNA were analyzed using western blot. (B) The protein levels of MMP-3, MMP-9, and TIMP-1 in TNF-α–treated AIA FLS transfected with BMP3 siRNA were analyzed using western blot. (C) The mRNA levels of MMP-3, MMP-9, and TIMP-1 in TNF-α–treated RA FLS transfected with BMP3 siRNA were analyzed using qPCR. (D) The mRNA levels of MMP-3, MMP-9, and TIMP-1 in TNF-α–treated AIA FLS transfected with BMP3 siRNA were analyzed using qPCR. (E) TNF-α–treated AIA FLS were transfected with BMP3 siRNA, and their migration into the wound-healing site after 24 hours was photographed (original magnification, ×10). (F) TNF-α–treated AIA FLS were transfected with BMP3 siRNA, and their migration into the wound-healing site after 24 hours was photographed (original magnification, ×10). All values are expressed as the mean ± SD. *P < 0.05, **P < 0.01 vs RA group. #P < 0.05, ##P < 0.01 vs NC-RNAi group.
Overexpression of BMP3 decreases proinflammatory cytokines and chemokines in RA and AIA FLS
To further study the effect of BMP3 on the proinflammatory cytokines and chemokines of RA FLS, we used the overexpression plasmids BMP3-pcDNA3.1 for humans and BMP3-PEX for rats to induce overexpression of BMP3 in TNF-α–treated RA and AIA FLS, respectively. As shown in Figure 5A and 5C, the western blot and qPCR results showed that BMP3 expression was higher in TNF-α–treated RA FLS transfected with BMP3-pcDNA3.1, compared to the cells transfected with NC-pcDNA3.1 (the negative control). Furthermore, the protein and mRNA levels of the proinflammatory cytokines IL-6, IL-1β, and IL-17A were significantly downregulated by BMP3-pcDNA3.1 in RA FLS treated with TNF-α (Figure 5A and 5C). In addition, the western blot and qPCR results showed that the expression of BMP3 was significantly higher in TNF-α–treated AIA FLS transfected with BMP3-PEX, compared to the cells transfected with empty vector (NC-PEX) (Figure 5B and 5D). The protein and mRNA levels of the proinflammatory cytokines IL-6, IL-1β, and IL-17A were significantly decreased by BMP3-PEX in AIA FLS treated with TNF-α (Figure 5B and 5D). More significantly, as shown in Figure 5E, after transfection with BMP3-pcDNA3.1, the qPCR results showed that the chemokines CCL-2, CCL-3, and VCAM-1 were notably reduced in TNF-α–treated RA FLS. Also, the mRNA expression of the chemokines CCL-2, CCL-3, and VCAM-1 was significantly reduced after transfection with BMP3-PEX in TNF-α–treated AIA FLS (Figure 5F). Thus, these results demonstrate that BMP3 overexpression decreases the levels of proinflammatory cytokines and chemokines in RA.
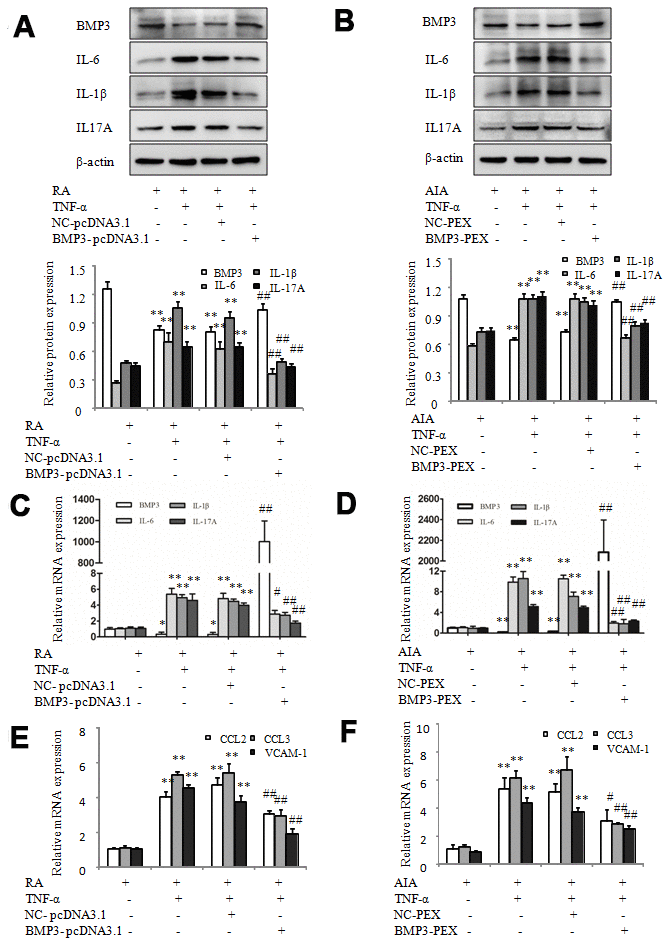
Figure 5. Overexpression of BMP3 decreases proinflammatory cytokines and chemokines in RA and AIA FLS. (A) The protein levels of BMP3, IL-6, IL-1β, and IL-17A in TNF-α–treated RA FLS transfected with BMP3-pcDNA3.1 were analyzed using western blot. (B) The protein levels of BMP3, IL-6, IL-1β, and IL-17A in TNF-α–treated AIA FLS transfected with BMP3-PEX were analyzed using western blot. (C) The mRNA levels of BMP3, IL-6, IL-1β, and IL-17A in TNF-α–treated RA FLS transfected with BMP3-pcDNA3.1 were analyzed using qPCR. (D) The mRNA levels of BMP3, IL-6, IL-1β, and IL-17A in TNF-α–treated AIA FLS transfected with BMP3-PEX were analyzed using qPCR. (E) The mRNA levels of CCL2, CCL3, and VCAM-1 in TNF-α–treated RA FLS transfected with BMP3-pcDNA3.1 were analyzed using qPCR. (F) The mRNA levels of CCL2, CCL3, and VCAM-1 in TNF-α–treated AIA FLS transfected with BMP3-PEX were analyzed using qPCR. All values are expressed as the mean ± SD. *P < 0.05, **P < 0.01 vs AIA group. #P < 0.05, ##P < 0.01 vs NC-PEX group.
Overexpression of BMP3 inhibits the migration of RA and AIA FLS
We measured the effect of BMP3 on the migration of RA and AIA FLS after transfection with BMP3 overexpression plasmids. As expected, the western blot and qPCR results showed that MMP-3 and MMP-9 expression was significantly upregulated in RA FLS stimulated by TNF-α. However, compared with the cells transfected with NC-pcDNA3.1, the increases in MMP-3 and MMP-9 were significantly inhibited by BMP3-pcDNA3.1 in TNF-α–treated RA FLS (Figure 6A and 6C). Conversely, the expression of TIMP-1 was significantly increased in RA FLS treated with TNF-α after transfection with BMP3-pcDNA3.1 (Figure 6A and 6C). In TNF-α–treated AIA FLS transfected with BMP3-PEX, the protein and mRNA levels of MMP-3 and MMP-9 were significantly downregulated (Figure 6B and 6D), whereas the expression of TIMP-1 was significantly upregulated (Figure 6B and 6D). In addition, the wound-healing assay indicated that the migration of RA FLS was suppressed after overexpression of BMP3 (Figure 6E). The migration of AIA FLS was also inhibited by BMP3-PEX transfection, compared to the negative control (Figure 6F). Taken together, these data demonstrate that BMP3 overexpression inhibits the migration of FLS in RA and AIA and that BMP3 may play a role in the pathogenesis of RA.
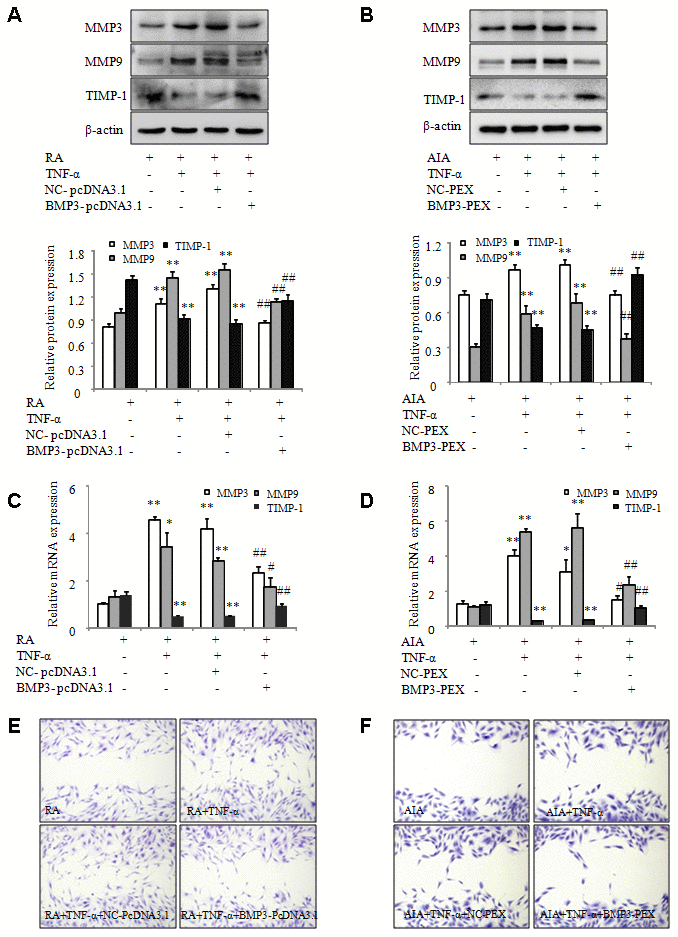
Figure 6. Overexpression of BMP3 inhibits the migration of RA and AIA FLS. (A) The protein levels of MMP-3, MMP-9, and TIMP-1 in TNF-α–treated RA FLS transfected with BMP3-pcDNA3.1 were analyzed using western blot. (B) The protein levels of MMP-3, MMP-9, and TIMP-1 in TNF-α–treated AIA FLS transfected with BMP3-PEX were analyzed using western blot. (C) The mRNA levels of MMP-3, MMP-9, and TIMP-1 in TNF-α–treated RA FLS transfected with BMP3-pcDNA3.1 were analyzed using qPCR. (D) The mRNA levels of MMP-3, MMP-9, and TIMP-1 in TNF-α–treated AIA FLS transfected with BMP3-PEX were analyzed using qPCR. (E) TNF-α–treated RA FLS were transfected with BMP3-pcDNA3.1, and their migration into the wound-healing site after 24 hours was photographed (original magnification, ×10). (F) TNF-α–treated AIA FLS were transfected with BMP3-PEX, and their migration into the wound-healing site after 24 hours was photographed (original magnification, ×10). All values are expressed as the mean ± SD. *P < 0.05, **P < 0.01 vs RA group. #P < 0.05, ##P < 0.01 vs NC-pcDNA3.1 group.
BMP3 regulates the migration and proinflammatory response of FLS and may be associated with the TGF-β1/Smad signaling pathway
BMP3 is associated with the TGF-β1/Smad signaling pathway in pathophysiologic mechanisms [25]. To investigate whether BMP3 regulates proinflammatory cytokines, chemokines, and migration through the Smad signaling pathway in FLS, we measured the protein levels of phosphorylated (p)-Smad2, a TGF-β1/Smad pathway activation marker. Western blot analysis demonstrated upregulation of p-Smad2 in RA and AIA when FLS were treated with TNF-α. In particular, expression of p-Smad2 was significantly increased after BMP3 silencing (Figure 7A and 7C) but reduced after BMP3 overexpression (Figure 7B and 7D). These results indicate that BMP3 may regulate FLS migration and secretion of proinflammatory cytokines through the Smad signaling pathway.
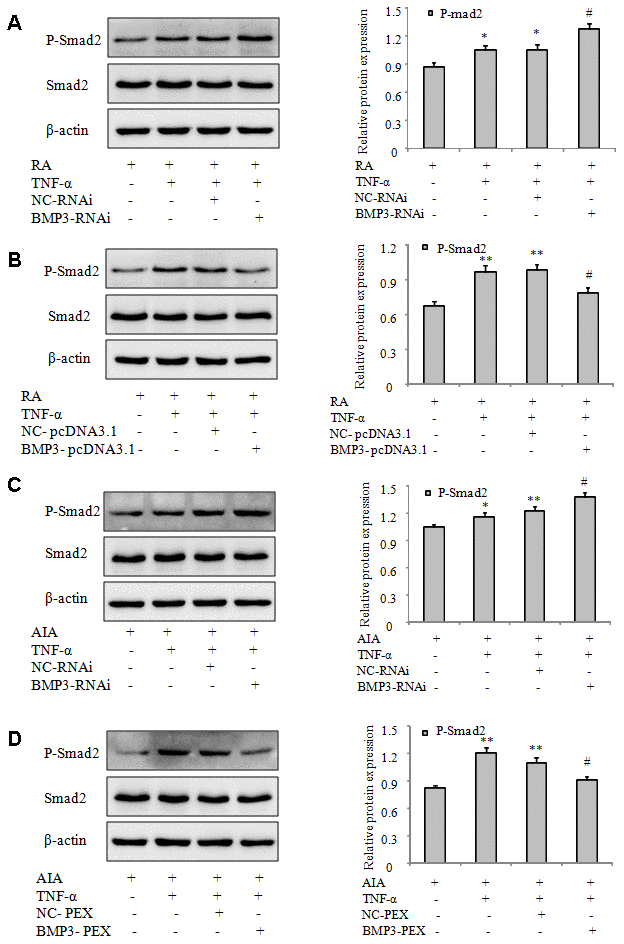
Figure 7. BMP3 regulates the proinflammatory response and migration of FLS and may be associated with the TGF-β1/Smad signaling pathway. (A) The protein levels of p-Smad2 in RA FLS transfected with BMP3 siRNA were analyzed using western blot. (B) The protein levels of p-Smad2 in RA FLS transfected with BMP3-PEX were analyzed using western blot. (C) The protein levels of p-Smad2 in AIA FLS transfected with BMP3-RNAi were analyzed using western blot. (D) The protein levels of p-Smad2 in AIA FLS transfected with BMP3-pcDNA3.1 were analyzed using western blot. All values are expressed as the mean ± SD. *P < 0.05, **P < 0.01 vs normal group. #P < 0.05, ##P < 0.01 vs model group.
Induction of BMP3 overexpression by adenovirus in vivo alleviated arthritis severity in AIA rats
To explore the effect of BMP3 in synovial tissues of AIA in vivo, adenovirus carrying Rattus BMP3 (ad-BMP3) was injected intra-articularly into the knees of AIA rats to induce overexpression of BMP3, and adenovirus carrying LUC (ad-LUC) was used as a control. First, in vivo imaging showed that adenovirus carrying BMP3 and LUC replicated successfully in rat joints (Figure 8A). Second, using immunofluorescence staining and immunohistochemical analysis to detect BMP3 expression in synovial tissues of AIA, we found that BMP3 expression was significantly higher in the synovial tissues of ad-BMP3–injected joints compared with ad-LUC–injected joints (Figure 8B and 8C). Overexpression of BMP3 also reduced paw swelling caused by AIA (Figure 9A) and the articular index (Figure 9B). More importantly, histopathologic analysis confirmed that intra-articular injection of ad-BMP3 significantly reduced inflammatory cell infiltration and synovial hyperplasia in the synovial tissue of AIA rats (Figure 9C). In addition, the results of western blot analysis showed that the proinflammatory cytokines IL-6, IL-1β, IL-17A, and TNF-α were all significantly downregulated by intra-articular injection of ad-BMP3 in AIA synovial tissues (Figure 9D). IL-6, IL-1β, and TNF-α expression was also significantly downregulated in the serum of AIA rats injected with ad-BMP3 (Figure 9E). Thus, our results confirm that overexpression of BMP3 induced by intra-articular injection of ad-BMP3 alleviates arthritis severity in AIA rats.
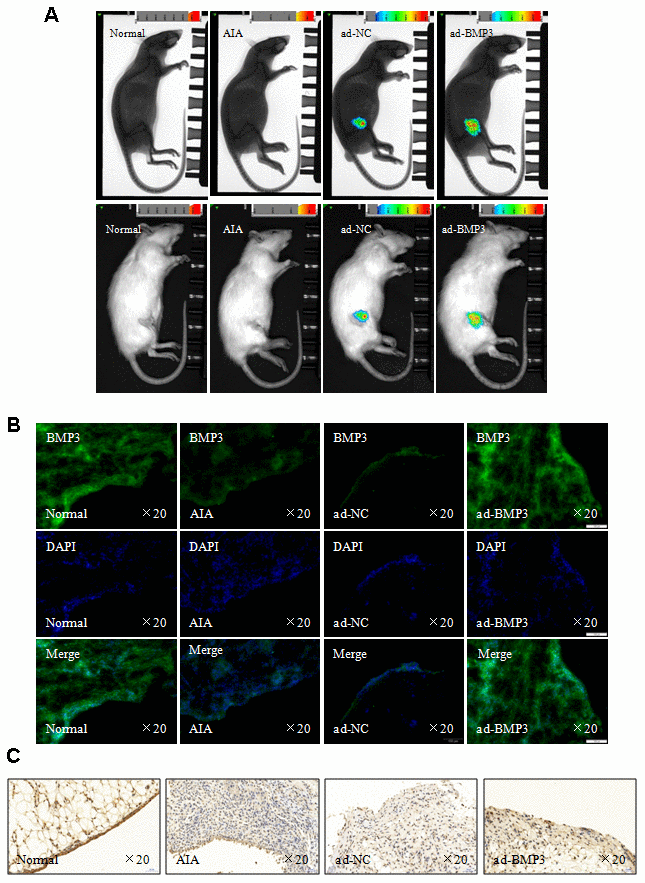
Figure 8. Overexpression of BMP3 by adenovirus in vivo alleviated arthritis severity in AIA rats. (A) In vivo imaging of normal and AIA rats. (B) Overexpression of BMP3 in AIA rat synovial tissues injected with ad-BMP3 was analyzed using immunofluorescence staining (original magnification, ×20). (C) BMP3 expression in AIA rat synovial tissues injected with ad-BMP3 was analyzed using IHC staining (original magnification, ×20).
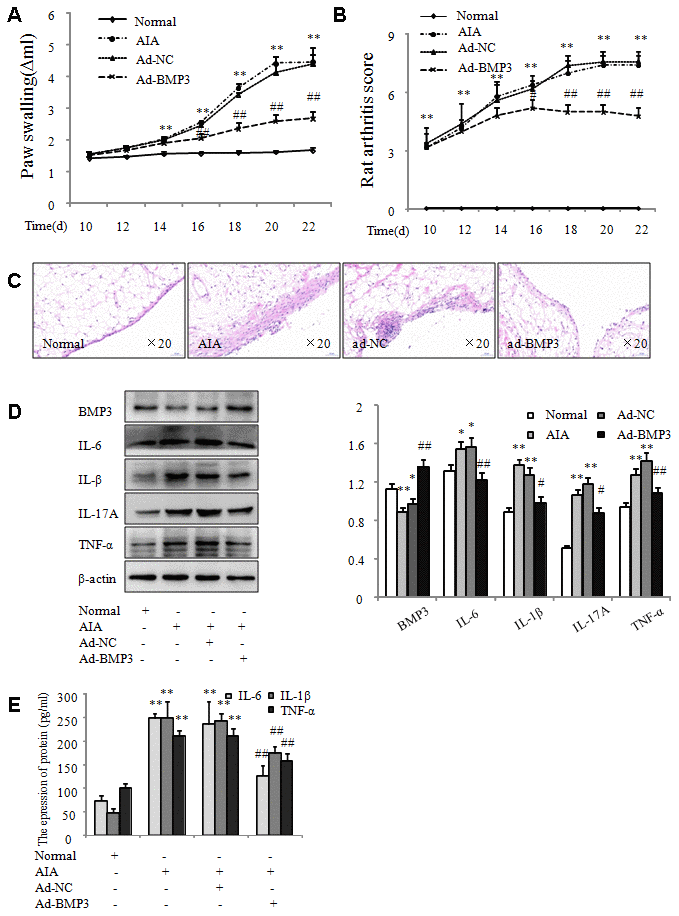
Figure 9. Overexpression of BMP3 by adenovirus in vivo alleviated arthritis severity in AIA rats. (A) The swelling of ankle joints was quantified using a plethysmometer. (B) Arthritis scores were reduced in AIA rats injected with ad-BMP3. (C) Representative H&E staining of normal and AIA rat synovial tissues (original magnification, ×20). (D) BMP3 protein levels in normal and AIA synovial tissues were analyzed using western blot. (E) Levels of IL-6, IL-1β, and TNF-α expression in the serum of AIA rats injected with ad-BMP3. All values are expressed as the mean ± SD. *P < 0.05, **P < 0.01 vs normal group. #P < 0.05, ##P < 0.01 vs ad-NC group.
Discussion
The results of this study demonstrated the following: (1) BMP3 expression is significantly downregulated in RA and AIA synovial tissues; (2) BMP3-RNAi increased the proinflammatory response and migration of RA and AIA FLS; (3) the overexpression vector of BMP3 suppressed the proinflammatory response and migration of RA and AIA FLS; (4) overexpression of BMP3 decreases the proinflammatory response and migration of FLS and BMP3 expression may be closely associated with the TGF-β1/Smad signaling pathway; and (5) induction of BMP3 overexpression by adenovirus in vivo alleviated arthritis severity in AIA rats.
The AIA rat model is widely used to study the pathogenesis of RA [26]. The AIA model has similar characteristics as RA with respect to histology and immunology and is a well-established model for evaluating treatment for RA [27]. Hence, we chose AIA to investigate the role of BMP3 in RA. RA is a systemic autoimmune disease characterized by synovial inflammation and hyperplasia and erosion of the synovial joints, leading to articular destruction and functional disability [28]. It is clear that various inflammatory cells are activated in RA, such as T cells, B cells, and FLS [29]. Accumulating evidence indicates that hyperplastic FLS potentially promote macrophage and lymphocyte infiltration, recruitment, and retention via the production of chemokines, cell adhesion molecules, proinflammatory cytokines, and extracellular matrix proteins [30–32]. Although FLS have been shown to play an important role in RA, the mechanisms underlying the activation of FLS remain unresolved.
BMP3, a member of the BMP family, has been shown to inhibit the proliferation of colorectal cancer cells [22] and biliary cancer cells [23]. BMP3 mRNA expression has also been shown to be lower in arthritic synovium than in the synovium of nonarthritic controls [24].
In this study, we explored whether BMP3 plays a role in regulating the proinflammatory response and migration of FLS. In this study, we show that BMP3 expression is significantly reduced in RA and AIA synovial tissues using western blot, qRT-PCR, immunohistochemical, and immunofluorescence analysis. Notably, after AIA and RA FLS were treated with various inflammation factors in vitro, the expression of BMP3 was decreased to different degrees. TNF-α is one of the major cytokines involved in RA. It has been shown that stimulation using 10 ng/mL of TNF-α results in the activation of RA FLS and increases production of inflammatory cytokines [10]. It has also been reported that TNF-α stimulation affects the expression levels of certain genes in RA FLS [33]. In the current study, we explored whether BMP3 affects the proinflammatory response and migration of RA and AIA FLS.
Western blot and qRT-PCR analyses revealed that inhibition of BMP3 expression using BMP3-RNAi resulted in the overexpression of the proinflammatory cytokines IL-6, IL-1β, and IL-17A in RA and AIA FLS treated with TNF-α. Studies indicate that the secretion of inflammatory factors leads to the activation and migration of RA FLS and exacerbates the inflammatory response [30, 34]. Furthermore, inhibition of BMP3 expression using BMP3-RNAi increases the expression of CCL-2, CCL-3, VCAM-1, MMP-3, and MMP-9 in RA and AIA FLS treated with TNF-α, whereas TIMP-1 expression is decreased. Macrophages and FLS in synovial tissues are a major source of chemokines [35]. Many chemokines produced by RA synovium may facilitate the recruitment and activation of FLS in synovial tissue and subsequently aggravate inflammation and bone destruction in the progression of RA [36, 37]. MMP-9 promotes the migration and invasion of FLS in collagen-induced arthritis in mice [38], whereas MMP-3 causes FLS to migrate to adjacent cartilage and induces articular cartilage degradation [39]. Significantly, our wound-healing assay results also suggest that inhibition of BMP3 expression increases the migration of RA and AIA FLS.
Our goal was to determine whether BMP3 overexpression reduces the proinflammatory response and migration of FLS. In this study, RA and AIA FLS treated with TNF-α were transfected with the overexpression vectors BMP3-PEX and BMP3-pcDNA3.1. Our results indicate that BMP3 overexpression downregulates the expression of proinflammatory cytokines, chemokines, MMP-3, and MMP-9, and upregulates TIMP-1 expression. Thus, BMP3 overexpression significantly reduces the proinflammatory response and migration of AIA FLS.
More importantly, inflammatory cells infiltration, arthritis scores, and paw swelling were significantly decreased by intra-articular injection of ad-BMP3 in AIA knees in vivo. The expression of IL-6, IL-1β, and TNF-α was also significantly downregulated in AIA serum. Thus, BMP3 overexpression may improve RA by suppressing the production of proinflammatory cytokines, chemokines, and MMPs of FLS.
BMP3 mediates cell activation by negatively regulating the TGF-β1/Smad signaling pathway [25]. The activation of p-Smad kinases plays a vital role in cell growth, proliferation, and invasion [14]. Our data also indicate that inhibition of BMP3 by BMP3-RNAi significantly increases p-Smad2 expression in AIA and RA FLS. In particular, overexpression of BMP3 suppressed Smad signaling, with a substantial decrease in p-Smad2 expression in AIA and RA FLS. Taken together, these data indicate that BMP3 mediates the proinflammatory cytokines and migration of FLS and might regulate the activation of the TGF-β1/Smad signaling pathway.
In conclusion, we found that BMP3 expression was significantly reduced in AIA and RA synovial tissues. BMP3 silencing or overexpression resulted in promotion or reduction of the proinflammatory response and migration of FLS, respectively. Our findings suggest that BMP3 plays a pivotal role in the proinflammatory response and migration of FLS through activation of the TGF-β1/Smad signaling pathway. Thus, we believe that BMP3 could be a new target for the treatment of RA.
Materials and Methods
Materials and reagents
Complete Freund’s adjuvant (CFA) was obtained from Chondrex, Inc. (USA). D-luciferin was purchased from Abcam (UK). Rabbit anti-BMP3, anti-TNF-a, and anti-TIMP metallopeptidase inhibitor 1 (TIMP-1) antibodies were obtained from Abcam (UK). Rabbit anti-MMP-3 and anti-MMP-9 antibodies were acquired from Merck Millipore (USA). Rabbit anti-IL-6, anti-IL-1β, and anti-IL-17A were purchased from Bioworld (USA). The rat anti-β-actin antibody was purchased from CST (USA). A horseradish peroxidase (HRP)-labeled goat anti-rabbit immunoglobulin (IgG) was procured from Zhongshan Biotechnology Corporation (China). BMP3, MMP-3, MMP-9, TIMP-1, TNF-a, IL-6, IL-1β, IL-17A, CCL-2, CCL-3, VCAM-1, and β-actin primers were synthesized by the Shanghai Sangon Biological and Technological Company (China).
Human synovial tissue
Human synovial tissue was obtained from patients with RA (n = 5) according to the RA criteria during joint synovectomies revised in 1987 by the American College of Rheumatology. Osteoarthritis (OA, n = 5) patients served as controls. All patients signed informed consent forms to participate in the study. The ethics board of Anhui Medical University approved the study protocol, and synovial tissue specimens were collected according to organizational guidelines. Written informed consent was obtained from all subjects.
Adjuvant-induced arthritis (AIA) model
All Sprague-Dawley (SD) rats (120–160 g, female) were purchased from the Laboratory Animal Center of Anhui Medical University. The animals’ experimental protocols were approved by the Animal Care and Use Committee of Anhui Medical University. Adult SD rats were intradermally injected with 0.1 mL of CFA per 100 g of body weight in the left paw to induce AIA. At the same time, control group rats were treated with normal saline on day 0. The right knees of AIA rats were intra-articularly injected with 0.1 mL of adenovirus carrying Rattus BMP3 (ad-BMP3) or ad-LUC on day 8. All rats were killed on day 23 after adjuvant injection.
Histopathology
Rat and human synovium specimens were fixed with 4% paraformaldehyde for 24 hours and then embedded in paraffin. Immunohistochemistry analysis and hematoxylin and eosin (H&E) staining were performed according to a standard procedure. The Olympus BX-51 microscope was used to photograph pathologic changes.
ELISA assay
Rats were killed at 23 days, and serum was collected through the abdominal aorta. The levels of IL-6, IL-1β, and TNF-α in serum or FLS supernatant were analyzed using ELISA Kit (Wuhan ColorfulGene Biological Technology Co., Ltd., China) according to the manufacturer’s procedure.
Cell culture and treatment
FLS were derived from the synovial tissues of the RA patients and AIA rats. The cells were cultured in cell culture flasks in high-glucose DMEM medium (Hyclone, USA) containing 20% (v/v) fetal bovine serum (FBS) (Gibco, USA), 100 mg/mL of streptomycin, and 100 U/mL of penicillin (both from Beyotime, China). All cells were cultured at 37°C at an atmosphere of 5% CO2. The FLS of 3-8 generation were used in the subsequent experiments.
We explored the effects of various inflammatory treatments on the expression of BMP3 in FLS. First, we seeded FLS into 6-well plate for 24 hours. Subsequently, the FLS were incubated with IL-6 (5 ng/mL), IL-1β (2 ng/mL), IL-17A (10 ng/mL), TNF-α (10 ng/mL), IFN-γ (10 ng/mL), and LPS (1 μg/mL) for 48 hours, respectively. Then the FLS were collected and analyzed using qPCR and Western blot.
Immunofluorescence staining
FLS were cultured in DMEM with 20% FBS at a density of 1–2 × 105 cells/mL. FLS were stimulated with TNF-α for 48 hours and then fixed with methanol. Immunofluorescence staining was performed with rabbit anti-BMP3 (Abcam, UK) and anti-Vimentin (Cell Signaling, USA). Alexa Fluor 488–conjugated goat anti-rabbit IgG (Zhongshan Biotechnology Corporation, China) was used as the secondary antibody. Counterstaining of nuclei was performed with 4′,6-diamidino-2-phenylindole (DAPI; Beyotime, China). Pathologic changes were observed and photographed using an Olympus BX-51 microscope.
Small interfering RNA transfection
Small interfering RNA targeting BMP3 (BMP3-RNAi) was purchased from GenePharma Corporation (China). The BMP3-RNAi (rat) sense strand is 5′-CCAAAGUCUUUGAAGCCAUTT-3′, and antisense strand is 5′-AUGGCUUCAAAGACUUUGGTT-3′. The BMP3-RNAi (human) sense strand is 5′-GCAAGACAAGGUCUCUGAAdTdT-3′, and antisense strand is 5′-UUCAGAGACCUUGUCUUGCdTdT-3′. The cultured FLS were transfected with 100 nM of small interfering RNA (RNAi) using Lipofectamine 2000 (Invitrogen, USA), according to the manufacturer’s instructions. Negative scrambled RNAi was used as control in the experiment. After transfection for 4–6 hours, we replaced the Opti-MEM with DMEM containing 20% FBS and co-cultured with TNF-α (10 ng/mL) for 48 hours, and then transfected FLS were collected for the subsequent experiments.
Plasmid construction and transfection
Overexpression plasmid for rat BMP3-PEX was purchased from GenePharma Corporation (Shanghai, China), and human BMP3-pcDNA3.1 was purchased from Hanbio (Shanghai, China). The FLS were transfected with BMP3-PEX and BMP3-pcDNA3.1 to induce the ectopic expression of BMP3 and with the empty PEX vector (PEX) or pcDNA3.1 as a control. The transfection experiments were performed using Lipofectamine 2000 (Invitrogen, USA) according to the manufacturer’s instructions. After incubation of transfected FLS with TNF-α (10 ng/mL) for 48 hours, cells were collected for subsequent tests, such as qRT-PCR and Western blot analyses.
Ad-BMP3 overexpression
BMP3 adenovirus (ad-BMP3) and the negative control adenovirus (ad-LUC) were purchased from Hanbio (Shanghai, China). The stock solutions of ad-BMP3 and ad-LUC contained 1 × 1010 plaque formation units (PFU)/mL, respectively.
In vivo imaging
D-luciferin was dissolved in DMSO and finally formulated into a 50 mmol/L solution. The rats were anesthetized with 2% isoflurane, and 50 μL of D-luciferin solution was injected intra-articularly into the right knee of each rat. After 5–10 minutes, the rats were placed right side up in the imaging chamber. The images were captured after a continuous exposure time of 15 seconds in brightfield or x-ray using SI Imaging Amix (SI Imaging Services Co., Ltd, USA).
Wound-healing assay
FLS were cultured in 6-well plates (1.0 × 106 cells/well) in DMEM containing 20% FBS for 24 h. Subsequently, cells were scratched with a pipette tip, serum deprived, and treated with BMP3-RNAi or BMP3-PEX/BMP3-pcDNA3.1 for 24 hours. Then, the cells were fixed with methanol, stained with crystal violet, and photographed using an Olympus BX-51 microscope.
Real-time quantitative PCR
Total RNA was extracted from FLS using TRIzol reagent (Invitrogen) according to the manufacturer’s manuals/protocol. Then, cDNA was reverse transcribed using a TAKARA kit system (Japan). The reaction was performed according to the manufacturer’s protocol using TB Green qPCR Master Mix (TAKARA, Japan). Furthermore, the mRNA expression of the genes was detected in a Pikoreal 96 real-time PCR system (Thermo Scientific, USA). All the primers we used are shown in Tables 1 and 2. The relative gene mRNA expression was analyzed using the 2−ΔΔCt method and normalized to β-actin mRNA expression.
Table 1. Primers sequences used for quantitative real-time PCR (Rat)
| Gene | Primer sequence |
| BMP3 | forward: 5′- CCTACCCTACCTAGCTACGTAT -3′ |
| reverse: 5′- GCTGGTGACATTGTTACTCATG -3′ | |
| IL-6 | forward: 5′- GAGCCCACCAGGAACGAAAGTC -3′ |
| reverse: 5′-TGTTGTGGGTGGTATCCTCTGTGAA-3′ | |
| IL-1β | forward: 5′- TGACCCATGTGAGCTGAAAG -3′ |
| reverse: 5′- AGGGATTTTGTCGTTGCTTG - 3′ | |
| IL-17A | forward: 5′-TGCCTGATGCTGTTGCTGCTAC-3′ |
| reverse: 5′-GGTGAAGTGGAACGGTTGAGGTAG-3′ | |
| TNF-α | forward: 5′-ACTCCCAGAAAAGCAAGCAA-3′ |
| reverse: 5′-CAGTTCCACATCTCGGATCA-3′ | |
| CCL-2 | forward: 5′-TAGCATCCACGTGCTGTCTC-3′ |
| reverse: 5′-TGCTGCTGGTGATTCTCTTG-3′ | |
| CCL-3 | forward: 5′-ACTGCCTGCTGCTTCTCCTA-3′ |
| reverse: 5′-CGGTTTCTCTTGGTCAGGAA-3′ | |
| VCAM-1 | forward: 5′-GTCAGCGAAGGAAACTGGAG-3′ |
| reverse: 5′-ACCGTGCAGTTGACAGTGAC-3′ | |
| MMP- 3 | forward: 5′- ATGATGAACGATGGACAGATGA - 3′ |
| reverse: 5′- CATTGGCTGAGTGAAAGAGACC -3′ | |
| MMP-9 | forward: 5′-CACTGTAACTGGGGGCAACT-3′ |
| reverse: 5′-CACTTCTTGTCAGCGTCGAA-3′ | |
| TIMP-1 | forward: 5′-CATCTCTGGCC TCTGGCATC-3′ |
| reverse: 5′-CATAACGCTGGTATAAGGTGGTCTC-3′ | |
| β-actin | forward: 5′-TTCGCCATGGATGACGATATC-3′ |
| reverse: 5′-TAGGAGTCCTTCTGACCCATAC-3′ |
Table 2. Primers sequences used for quantitative real-time PCR (Human).
| Gene | Primer sequence |
| BMP3 | forward: 5′-GCTCTACTGGGGTCTTGCTG -3′ |
| reverse: 5′-CTGAGCCTGAAGGGTCTTGT -3′ | |
| IL-6 | forward: 5′- CACACAGACAGCCACTCACC -3′ |
| reverse: 5′- AGTGCCTCTTTGCTGCTTTC -3′ | |
| IL-1β | forward: 5′- GGACAAGCTGAGGAAGATGC -3′ |
| reverse: 5′- TCGTTATCCCATGTGTCGAA - 3′ | |
| IL-17A | forward: 5′- CGGACTGTGATGGTCAACCTGAAC -3′ |
| reverse: 5′- GGTCCTCATTGCGGTGGAGATTC -3′ | |
| TNF-α | forward: 5′-AACCTCCTCTCTGCCATCAA -3′ |
| reverse: 5′-CTGAGTAGGTCACCCTTCTC -3′ | |
| CCL-2 | forward: 5′- CCTTCATTCCCCAAGGGCTC -3′ |
| reverse: 5′- CTTCTTTGGGACACTTGCTGC -3′ | |
| CCL-3 | forward: 5′- TGCAACCAGTTCTCTGCATC -3′ |
| reverse: 5′- TGGCTGCTCGTCTCAAAGTA -3′ | |
| VCAM-1 | forward: 5′-GTCAGCGAAGGAAACTGGAG -3′ |
| reverse: 5′-ACCGTGCAGTTGACAGTGAC -3′ | |
| MMP- 3 | forward: 5′- GGCCAGGGATTAATGGAGAT - 3′ |
| reverse: 5′- TGAAAGAGACCCAGGGAGTG -3′ | |
| MMP-9 | forward: 5′- GTACCACGGCCAACTACGAC -3′ |
| reverse: 5′- GCCTTGGAAGATGAATGGAA -3′ | |
| TIMP-1 | forward: 5′- TGACATCCGGTTCGTCTACA -3′ |
| reverse: 5′- TGATGTGCAAGAGTCCATCC -3′ | |
| β-actin | forward: 5′- GCCAACACAGTGCTGTCTGG -3′ |
| reverse: 5′- CTCAGGAGGAGCAATGATCTTG -3′ |
Western blotting
Cultured FLS in vitro and synovial tissues were lysed using the radioimmunoprecipitation assay (RIPA) reagent containing 1% phenylmethanesulfonyl fluoride (PMSF) (Beyotime, China). The whole-cell extracts (20 mg protein) were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and blotted onto PVDF membranes (Millipore, USA). After blocking with 5% milk for 2 hours, the membranes were incubated with the appropriate antibodies, which were diluted using an antibody dilution buffer (Beyotime, China). The primary antibodies against BMP3 (1:500), IL-6 (1:500), IL-1β (1:500), IL-17 (1:500), TNF-α (1:500), MMP-3 (1:500), MMP-9 (1:500), TIMP-1 (1:500), TGF-β1 (1:500), and β-actin (1:1000) were used. After incubation with the primary antibodies for 12 hours, the blot was washed three times using TBS/Tween-20 (TBST). The blot was then incubated with goat anti-mouse or rabbit HRP-conjugated antibodies at 1:5000 dilutions in TBST containing 5% skim milk for 2 hours. After washing three times with TBST, the protein bands in the western blot were analyzed using Immobilon Western Chemiluminescent HRP Substrate (Millipore).
Statistical analysis
All data are presented as the means ± standard deviations (SDs) and were analyzed by SPSS 16.0 software. The results were from at least three experiments. The Student-Newman-Keuls test or one-way analysis of variance (ANOVA) was used to compare the means of different values. P values < 0.05 were considered statistically significant.
Conflicts of Interest
The authors declare no conflicts of interest.
Funding
This project was supported by the National Natural Science Foundation of China (No. 81473268) and the University Synergy Innovation Program of Anhui Province (No. GXXT-2019-045).
References
- 1. McInnes IB, Schett G. The pathogenesis of rheumatoid arthritis. N Engl J Med. 2011; 365:2205–19. https://doi.org/10.1056/NEJMra1004965 [PubMed]
- 2. Scott DL, Wolfe F, Huizinga TW. Rheumatoid arthritis. Lancet. 2010; 376:1094–108. https://doi.org/10.1016/S0140-6736(10)60826-4 [PubMed]
- 3. Bartok B, Firestein GS. Fibroblast-like synoviocytes: key effector cells in rheumatoid arthritis. Immunol Rev. 2010; 233:233–55. https://doi.org/10.1111/j.0105-2896.2009.00859.x [PubMed]
- 4. Jia X, Wei F, Sun X, Chang Y, Xu S, Yang X, Wang C, Wei W. CP-25 attenuates the inflammatory response of fibroblast-like synoviocytes co-cultured with BAFF-activated CD4(+) T cells. J Ethnopharmacol. 2016; 189:194–201. https://doi.org/10.1016/j.jep.2016.05.034 [PubMed]
- 5. Kim EK, Kwon JE, Lee SY, Lee EJ, Kim DS, Moon SJ, Lee J, Kwok SK, Park SH, Cho ML. IL-17-mediated mitochondrial dysfunction impairs apoptosis in rheumatoid arthritis synovial fibroblasts through activation of autophagy. Cell Death Dis. 2017; 8:e2565. https://doi.org/10.1038/cddis.2016.490 [PubMed]
- 6. Lerner A, Neidhöfer S, Reuter S, Matthias T. MMP3 is a reliable marker for disease activity, radiological monitoring, disease outcome predictability, and therapeutic response in rheumatoid arthritis. Best Pract Res Clin Rheumatol. 2018; 32:550–62. https://doi.org/10.1016/j.berh.2019.01.006 [PubMed]
- 7. Miao CG, Yang YY, He X, Li XF, Huang C, Huang Y, Zhang L, Lv XW, Jin Y, Li J. Wnt signaling pathway in rheumatoid arthritis, with special emphasis on the different roles in synovial inflammation and bone remodeling. Cell Signal. 2013; 25:2069–78. https://doi.org/10.1016/j.cellsig.2013.04.002 [PubMed]
- 8. Sato H, Muraoka S, Kusunoki N, Masuoka S, Yamada S, Ogasawara H, Imai T, Akasaka Y, Tochigi N, Takahashi H, Tsuchiya K, Kawai S, Nanki T. Resistin upregulates chemokine production by fibroblast-like synoviocytes from patients with rheumatoid arthritis. Arthritis Res Ther. 2017; 19:263. https://doi.org/10.1186/s13075-017-1472-0 [PubMed]
- 9. Conde J, Scotece M, López V, Gómez R, Lago F, Pino J, Gómez-Reino JJ, Gualillo O. Adiponectin and leptin induce VCAM-1 expression in human and murine chondrocytes. PLoS One. 2012; 7:e52533. https://doi.org/10.1371/journal.pone.0052533 [PubMed]
- 10. Du H, Zhang X, Zeng Y, Huang X, Chen H, Wang S, Wu J, Li Q, Zhu W, Li H, Liu T, Yu Q, Wu Y, Jie L. A novel phytochemical, DIM, inhibits proliferation, migration, invasion and TNF-α induced inflammatory cytokine production of synovial fibroblasts from rheumatoid arthritis patients by targeting MAPK and AKT/mTOR signal pathway. Front Immunol. 2019; 10:1620. https://doi.org/10.3389/fimmu.2019.01620 [PubMed]
- 11. Zheng W, Pan H, Wei L, Gao F, Lin X. Dulaglutide mitigates inflammatory response in fibroblast-like synoviocytes. Int Immunopharmacol. 2019; 74:105649. https://doi.org/10.1016/j.intimp.2019.05.034 [PubMed]
- 12. Zhang Y, Dong J, He P, Li W, Zhang Q, Li N, Sun T. Genistein inhibit cytokines or growth factor-induced proliferation and transformation phenotype in fibroblast-like synoviocytes of rheumatoid arthritis. Inflammation. 2012; 35:377–87. https://doi.org/10.1007/s10753-011-9365-x [PubMed]
- 13. Song HY, Kim MY, Kim KH, Lee IH, Shin SH, Lee JS, Kim JH. Synovial fluid of patients with rheumatoid arthritis induces alpha-smooth muscle actin in human adipose tissue-derived mesenchymal stem cells through a TGF-beta1-dependent mechanism. Exp Mol Med. 2010; 42:565–73. https://doi.org/10.3858/emm.2010.42.8.057 [PubMed]
- 14. Zhu D, Zhao J, Lou A, Huang Q, OuYang Q, Zhu J, Fan M, He Y, Ren H, Yang M. Transforming growth factor β1 promotes fibroblast-like synoviocytes migration and invasion via TGF-β1/smad signaling in rheumatoid arthritis. Mol Cell Biochem. 2019; 459:141–50. https://doi.org/10.1007/s11010-019-03557-0 [PubMed]
- 15. Totoson P, Maguin-Gaté K, Nappey M, Prati C, Wendling D, Demougeot C. Microvascular abnormalities in adjuvant-induced arthritis: relationship to macrovascular endothelial function and markers of endothelial activation. Arthritis Rheumatol. 2015; 67:1203–13. https://doi.org/10.1002/art.39065 [PubMed]
- 16. Zhou G, Sun X, Qin Q, Lv J, Cai Y, Wang M, Mu R, Lan HY, Wang QW. Loss of Smad7 promotes inflammation in rheumatoid arthritis. Front Immunol. 2018; 9:2537. https://doi.org/10.3389/fimmu.2018.02537 [PubMed]
- 17. Katagiri T, Watabe T. Bone morphogenetic proteins. Cold Spring Harb Perspect Biol. 2016; 8:a021899. https://doi.org/10.1101/cshperspect.a021899 [PubMed]
- 18. Urist MR, Strates BS. Bone morphogenetic protein. J Dent Res. 1971; 50:1392–406. https://doi.org/10.1177/00220345710500060601 [PubMed]
- 19. Song B, Li XF, Yao Y, Xu QQ, Meng XM, Huang C, Li J. BMP9 inhibits the proliferation and migration of fibroblast-like synoviocytes in rheumatoid arthritis via the PI3K/AKT signaling pathway. Int Immunopharmacol. 2019; 74:105685. https://doi.org/10.1016/j.intimp.2019.105685 [PubMed]
- 20. Bahamonde ME, Lyons KM. BMP3: to be or not to be a BMP. J Bone Joint Surg Am. 2001; 83:S56–62. [PubMed]
- 21. Kokabu S, Gamer L, Cox K, Lowery J, Tsuji K, Raz R, Economides A, Katagiri T, Rosen V. BMP3 suppresses osteoblast differentiation of bone marrow stromal cells via interaction with Acvr2b. Mol Endocrinol. 2012; 26:87–94. https://doi.org/10.1210/me.2011-1168 [PubMed]
- 22. Loh K, Chia JA, Greco S, Cozzi SJ, Buttenshaw RL, Bond CE, Simms LA, Pike T, Young JP, Jass JR, Spring KJ, Leggett BA, Whitehall VL. Bone morphogenic protein 3 inactivation is an early and frequent event in colorectal cancer development. Genes Chromosomes Cancer. 2008; 47:449–60. https://doi.org/10.1002/gcc.20552 [PubMed]
- 23. Kisiel JB, Li J, Zou H, Oseini AM, Strauss BB, Gulaid KH, Moser CD, Aderca I, Ahlquist DA, Roberts LR, Shire AM. Methylated bone morphogenetic protein 3 (BMP3) gene: evaluation of tumor suppressor function and biomarker potential in biliary cancer. J Mol Biomark Diagn. 2013; 4:1000145. https://doi.org/10.4172/2155-9929.1000145 [PubMed]
- 24. Matzelle MM, Shaw AT, Baum R, Maeda Y, Li J, Karmakar S, Manning CA, Walsh NC, Rosen V, Gravallese EM. Inflammation in arthritis induces expression of BMP3, an inhibitor of bone formation. Scand J Rheumatol. 2016; 45:379–83. https://doi.org/10.3109/03009742.2015.1126347 [PubMed]
- 25. Yu X, Gu P, Huang Z, Fang X, Jiang Y, Luo Q, Li X, Zhu X, Zhan M, Wang J, Fan L, Chen R, Yu J, et al. Reduced expression of BMP3 contributes to the development of pulmonary fibrosis and predicts the unfavorable prognosis in IIP patients. Oncotarget. 2017; 8:80531–44. https://doi.org/10.18632/oncotarget.20083 [PubMed]
- 26. van den Berg WB. Lessons from animal models of arthritis over the past decade. Arthritis Res Ther. 2009; 11:250. https://doi.org/10.1186/ar2803 [PubMed]
- 27. Snekhalatha U, Anburajan M, Venkatraman B, Menaka M. Evaluation of complete freund’s adjuvant-induced arthritis in a wistar rat model. Comparison of thermography and histopathology. Z Rheumatol. 2013; 72:375–82. https://doi.org/10.1007/s00393-012-1083-8 [PubMed]
- 28. Bottini N, Firestein GS. Duality of fibroblast-like synoviocytes in RA: passive responders and imprinted aggressors. Nat Rev Rheumatol. 2013; 9:24–33. https://doi.org/10.1038/nrrheum.2012.190 [PubMed]
- 29. Lee DG, Woo JW, Kwok SK, Cho ML, Park SH. MRP8 promotes Th17 differentiation via upregulation of IL-6 production by fibroblast-like synoviocytes in rheumatoid arthritis. Exp Mol Med. 2013; 45:e20. https://doi.org/10.1038/emm.2013.39 [PubMed]
- 30. Sluzalska KD, Liebisch G, Lochnit G, Ishaque B, Hackstein H, Schmitz G, Rickert M, Steinmeyer J. Interleukin-1β affects the phospholipid biosynthesis of fibroblast-like synoviocytes from human osteoarthritic knee joints. Osteoarthritis Cartilage. 2017; 25:1890–99. https://doi.org/10.1016/j.joca.2017.07.011 [PubMed]
- 31. Smeets TJ, Barg EC, Kraan MC, Smith MD, Breedveld FC, Tak PP. Analysis of the cell infiltrate and expression of proinflammatory cytokines and matrix metalloproteinases in arthroscopic synovial biopsies: comparison with synovial samples from patients with end stage, destructive rheumatoid arthritis. Ann Rheum Dis. 2003; 62:635–38. https://doi.org/10.1136/ard.62.7.635 [PubMed]
- 32. Smeets TJ, Kraan MC, Galjaard S, Youssef PP, Smith MD, Tak PP. Analysis of the cell infiltrate and expression of matrix metalloproteinases and granzyme B in paired synovial biopsy specimens from the cartilage-pannus junction in patients with RA. Ann Rheum Dis. 2001; 60:561–65. https://doi.org/10.1136/ard.60.6.561 [PubMed]
- 33. Huang SH, Liu GW, Li JH, Xu JH, Xu DW, Zhang WQ, Huang JR. Expression of TREM-2 and its inhibitory effects on TNF-α induced inflammation in fibroblast-like synoviocytes via inhibiting p38 pathway activation. Clin Exp Rheumatol. 2018; 36:185–94. [PubMed]
- 34. Sun W, Qin R, Qin R, Wang R, Ding D, Yu Z, Liu Y, Hong R, Cheng Z, Wang Y. Sam68 promotes invasion, migration, and proliferation of fibroblast-like synoviocytes by enhancing the NF-κB/P65 pathway in rheumatoid arthritis. Inflammation. 2018; 41:1661–70. https://doi.org/10.1007/s10753-018-0809-4 [PubMed]
- 35. Tani K, Shimizu T, Motoki Y, Sone S. Chemokines in synovial inflammation in rheumatoid arthritis: basic and clinical aspects. Mod Rheumatol. 2002; 12:93–99. https://doi.org/10.3109/s101650200017 [PubMed]
- 36. Bustamante MF, Garcia-Carbonell R, Whisenant KD, Guma M. Fibroblast-like synoviocyte metabolism in the pathogenesis of rheumatoid arthritis. Arthritis Res Ther. 2017; 19:110. https://doi.org/10.1186/s13075-017-1303-3 [PubMed]
- 37. You S, Yoo SA, Choi S, Kim JY, Park SJ, Ji JD, Kim TH, Kim KJ, Cho CS, Hwang D, Kim WU. Identification of key regulators for the migration and invasion of rheumatoid synoviocytes through a systems approach. Proc Natl Acad Sci USA. 2014; 111:550–55. https://doi.org/10.1073/pnas.1311239111 [PubMed]
- 38. Shi Y, Wu Q, Xuan W, Feng X, Wang F, Tsao BP, Zhang M, Tan W. Transcription factor SOX5 promotes the migration and invasion of fibroblast-like synoviocytes in part by regulating MMP-9 expression in collagen-induced arthritis. Front Immunol. 2018; 9:749. https://doi.org/10.3389/fimmu.2018.00749 [PubMed]
- 39. Du F, Lü LJ, Teng JL, Shen N, Ye P, Bao CD. T-614 alters the production of matrix metalloproteinases (MMP-1 andMMP-3) and inhibits the migratory expansion of rheumatoid synovial fibroblasts, in vitro. Int Immunopharmacol. 2012; 13:54–60. https://doi.org/10.1016/j.intimp.2012.03.003 [PubMed]