



Introduction
Acute ischemic stroke (AIS) is a major cerebrovascular disease ascribing to the sudden reduction of cerebral blood flow, characterized by a series of cellular and molecular disturbances. With a mortality rate of 10.3% and a morbidity rate of 19.7% in China [1], ischemic stroke is the leading cause of death and disability worldwide [2]. Several common risk factors, including hypertension, diabetes, dyslipidemia, alcohol, smoking, and inflammation [3–5], have been identified to be related to the pathogenesis of AIS. It is generally accepted that impaired neurovascular repair, especially angiogenesis, plays crucial roles in the development of ischemic cerebral injury [6].
Angiogenesis is the physiological process through which new blood vessels form by the extension or elaboration of existing vessels [7]. This process depending on endothelial cells is under the control of an extensive variety of angiogenic stimulators and inhibitors [8]. A growing number of studies have shown that microRNAs (miRNAs) are involved in the regulation of angiogenesis in ischemic diseases, including AIS [9–11].
MiRNAs are ~22nt RNAs that mediate posttranscriptional regulation of mRNA targets by binding to their 3′ un-translated region (UTRs) in diverse eukaryotic lineages [12]. Changes of MiRNA expression profile have been detected in AIS patients [13], middle cerebral artery occlusion (MCAO) rats [14] and oxygen-glucose deprivation/reoxygenation (OGD/R) cells [15–17]. Particularly, several angiogenesis-associated miRNAs, such as miR-210 [18, 19], miR-21 [20, 21], miR-126 [22, 23], miR-155 [24, 25], miR-31 [26], miR-223-3p [27], miR-191 [28] and miR-361 [29], are altered in patients with AIS and injured endothelial cells. However, the mechanism underlying the role of angiogenesis-associated miRNAs in AIS remains to be further explored.
Thus, the aim of the present study was to investigate the effects of angiogenesis-associated miRNAs in the angiogenesis after AIS both in vitro and in vivo.
Results
Screening of miRNAs
To exclude tumor-related angiogenesis, we retrieved all published miRNAs related to the angiogenesis of endothelial cell in Pubmed (32 miRNAs, Table 1). After excluded 24 miRNAs that have been shown to play definite roles in acute stroke, we selected 8 miRNAs for further study (Table 2).
Table 1. Endothelial angiogenesis associated miRNAs.
| miRNA names | Cell types | miRNA target(s) | Impact on angiogenesis | References(PMID) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-210 | HUVEC | EFNA3 | promote | 18417479 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-424 | HUVEC;BOEC;MVEC | CUL2 | promote | 20972335 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-200b | HMEC | ETS1 | inhibit | 21081489 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-24 | HUVEC | GATA2;PAK4 | inhibit | 21788589 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-21 | HPAEC | RhoB; Rho-kinase | promote | 22371328 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-125b | HUVEC | VE-cadherin | inhibit | 22391569 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-361 | HUVEC | VEGF | inhibit | 23128854 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Let-7;miR-103 | HUVEC | AGO1 | promote | 23426184 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-29a | HUVEC | HBP1 | promote | 23541945 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-93 | HUVEC | P21; E2F-1; P53 | promote | 23559675 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-320 | HUVEC | NRP1 | inhibit | 24114198 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-31 | EPC | FAT4;TBXA2R | promote | 24558106 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-101 | HUVEC | Cul3 | promote | 24844779 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-221 | EPC | PIK3R1 | inhibit | 25236949 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-222 | EPC | ETS1 | inhibit | 25236949 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-223-3p | CMEC | RPS6KB1;HIF-1α | inhibit | 25313822 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-429 | HUVEC | HIF-1α | inhibit | 25550463 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-107 | RBMECs; HUVECs | Dicer-1 | promote | 26294080 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-185 | HMEC-1 | STIM1 | inhibit | 26694763 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-640 | HUVEC | HIF-1α | inhibit | 26879375 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-140-5p | HUVEC | VEGF | inhibit | 27035554 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-126 | EPC;ESC | SPRED1; PIK3R2/p85β | promote | 27180261 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-195 | hEPC | GABARAPL1 | inhibit | 27623937 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-155 | HUVEC | E2F2 | promote | 27731397 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-193a-3p | ECFC | HMGB1 | inhibit | 28276476 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-186 | HUVEC | HIF-1α | inhibit | 28571741 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-153-3p | HUVEC | HIF-1α | inhibit | 28985553 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-191 | HUVEC | HIF-2α | inhibit | 30090327 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-503 | EPC | Apelin | promote | 29800588 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-19b | HUVEC | TGFβ2 | inhibit | 30425199 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-19a | HUVEC | EDNRB | inhibit | 30550764 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: HUVEC: human umbilical vein endothelial cells; BOEC: blood outgrowth endothelial cells; MVEC: microvasculature endothelial cells; HMEC: human microvascular endothelial cells; HPAECs: human pulmonary arterial endothelial cells; EPC: endothelial progenitor cells; CMEC: cardiac microvascular endothelial cells; RBMEC: rat brain microvascular endothelial cells; ESC: embryonic stem cell; ECFC: circulating endothelial colony forming cells; EFNA3: Ephrin-A3; CUL2/3: cullin 2/3; Ets-1: v-ets erythroblastosis virus E26 oncogene homolog 1; GATA2: Endothelial transcription factor GATA2; PAK4: Serine/threonine-protein kinase PAK4; VE- cadherin: vascular endothelial (VE)-cadherin; VEGF: vascular endothelial growth factor; AGO1: argonaute 1; HBP1:HMG-box transcription factor 1; P21: cyclin dependent kinase inhibitor 1A; E2F-1/2: E2F transcription factor 1/2; P53: tumor protein p53; NRP1: Neuropilin 1; FAT4: FAT atypical cadherin 4; TBXA2R: thromboxane A2 receptor; PIK3R1: phosphoinositide-3-kinase regulatory subunit 1; RPS6KB1: ribosomal protein S6 kinase B1; HIF-1/2α: hypoxia inducible factor 1/2 subunit alpha; STIM1: stromal interaction molecule 1; SPRED1: Sprouty-related protein; PIK3R2/p85β: phosphoinositol-3 kinase regulatory subunit 2; GABARAPL1: GABA type A receptor associated protein like 1; HMGB1: high mobility group box 1; TGFβ2: transforming growth factor beta 2; EDNRB: endothelin receptor type B. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 2. Endothelial angiogenesis associated miRNAs without AIS study.
| miRNA names | Cell types | miRNA target(s) | Impact on angiogenesis | References | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-361 | HUVEC | VEGF | inhibit | [25] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-29a | HUVEC | HBP1 | promote | [26] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-31 | EPC | FAT4;TBXA2R | promote | [22] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| mir-223-3p | CMEC | RPS6KB1;HIF-1α | inhibit | [23] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-640 | HUVEC | HIF-1α | inhibit | [27] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-193a-3p | ECFC | HMGB1 | inhibit | [28] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-191 | HUVEC | HIF-2α | inhibit | [24] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-503 | EPC | Apelin | promote | [29] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: AIS: acute ischemic stroke; HUVEC: human umbilical vein endothelial cells; EPC: endothelial progenitor cells; CMEC: cardiac microvascular endothelial cells; ECFC: circulating endothelial colony forming cells; VEGF: vascular endothelial growth factor; HBP1:HMG-box transcription factor 1; FAT4: FAT atypical cadherin 4; TBXA2R: thromboxane A2 receptor; RPS6KB1: ribosomal protein S6 kinase B1; HIF-1/2α: hypoxia inducible factor 1/2 subunit alpha; HMGB1: high mobility group box 1. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Characteristics of enrollment patients
We enrolled 6 AIS patients and 6 control subjects as Cohort A, and another 12 AIS patients and 12 control subjects as Cohort B. Characteristics of Cohort A and B were shown in Table 3 and Table 4, respectively. There was no statistically difference of demographic or vascular risk factors between AIS patients and controls.
Table 3. Characteristics of acute ischemic stroke and control subjects in Cohort A.
| Characteristic | Healthy controls (M±SEM) | Stroke patients (M±SEM) | P value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (years) | 74.5±3.9 | 71.3±4.2 | 0.592 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gender (%) | 50F/50M | 50F/50M | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CAD history, n(%) | 2(33.3) | 1(16.7) | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HT history, n(%) | 5(83.3) | 3(50) | 0.545 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DM history, n(%) | 2(33.3) | 2(33.3) | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Smoking history, n(%) | 1(16.7) | 1(16.7) | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BMI(kg/m2) | 26.8±1.3 | 25.2±0.6 | 0.297 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SBP(mmHg) | 145.7±8.6 | 148.3±7.5 | 0.820 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DBP(mmHg) | 73.5±5.8 | 74.2±3.6 | 0.204 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| mRS score | 2.67±0.8 | - | - | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HBA1c(%) | 6.5±0.5 | 6.8±0.5 | 0.694 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FBG(mmol/L) | 5.7±0.5 | 6.6±0.6 | 0.307 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TG(mmol/L) | 1.3±0.3 | 1.6±0.6 | 0.719 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TC(mmol/L) | 4.4±0.7 | 4.7±0.5 | 0.789 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LDL-C(mmol/L) | 2.5±0.5 | 2.8±0.4 | 0.617 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HDL-C(mmol/L) | 1.6±0.3 | 1.4±0.2 | 0.732 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Creatinine(μmol/L) | 80.3±9.0 | 73.8±7.1 | 0.584 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BUN(mmol/L) | 5.6±0.4 | 6.2±0.8 | 0.526 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LP(α)(mg/L) | 249.3±120.1 | 288.3±139.0 | 0.836 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hcy(μmol/L) | 15.8±2.1 | 13.8±0.8 | 0.341 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AST(U/L) | 18.4±2.3 | 17.9±1.5 | 0.861 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ALT(U/L) | 21.4±3.0 | 16.6±2.6 | 0.256 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| γGT(U/L) | 41.8±14.5 | 16.8±1.7 | 0.118 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CRP(mg/L) | 2.4±1.1 | 4.7±3.0 | 0.481 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| WBC count(×109/L) | 5.7±0.7 | 6.2±0.6 | 0.605 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hemoglobin(g/L) | 133.5±6.3 | 139.3±5.7 | 0.507 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Platelets(×109/L) | 169.7±13.3 | 177.8±32.5 | 0.821 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PT(s) | 11.5±0.6 | 10.5±0.5 | 0.230 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| APTT(s) | 33.1±2.0 | 30.1±2.0 | 0.311 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FIB(g/L) | 2.9±0.2 | 3.0±0.1 | 0.609 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: CAD: coronary artery disease; HT: hypertension; DM: diabetes mellitus; BMI: body mass index; SBP: systolic blood pressure; DBP: diastolic blood pressure; HBA1c: glycolated hemoglobin A1c; FBG: fasting blood glucose; TG: total triglyceride; TC: total cholesterol; LDL-C: low density lipoprotein cholesterol; HDL-C: high density lipoprotein cholesterol; BUN: blood urea nitrogen; LP(α): lipoprotein; Hcy: homocysteine; AST: aspartate aminotransferase; ALT: alanine aminotransferase; γGT: γ-aminobutyric acid; CRP: C-reactive protein; WBC: white blood cell; PT: prothrombin time; APTT: activated partial thromboplastin time; FIB: fibrinogen. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 4. Characteristics of acute ischemic stroke and control subjects in Cohort B.
| Characteristic | Healthy controls (M±SEM) | Stroke patients (M±SEM) | P value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (years) | 75.4±2.8 | 75.3±3.0 | 0.968 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gender (%) | 50F/50M | 48.7F/51.3M | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CAD history, n(%) | 2(16.7) | 2(16.7) | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HT history, n(%) | 8(66.7) | 9(75) | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DM history, n(%) | 2(16.7) | 3(25) | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Smoking history, n(%) | 3(25) | 4(33.3) | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BMI(kg/m2) | 24.7±0.7 | 23.2±1.2 | 0.273 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SBP(mmHg) | 131.8±5.0 | 135.1±4.2 | 0.614 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DBP(mmHg) | 72.5±3.4 | 76.7±2.9 | 0.357 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| mRS | 2.58±0.8 | - | - | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HBA1c(%) | 5.8±0.2 | 5.9±0.8 | 0.838 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FBG(mmol/L) | 6.0±0.6 | 7.5±1.1 | 0.246 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TG(mmol/L) | 1.4±0.1 | 1.7±0.3 | 0.205 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TC(mmol/L) | 4.2±0.2 | 4.6±0.2 | 0.212 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LDL-C(mmol/L) | 2.7±0.2 | 2.9±0.2 | 0.411 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HDL-C(mmol/L) | 1.2±0.1 | 1.3±0.1 | 0.578 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Creatinine(μmol/L) | 82.8±8.8 | 88.8±8.6 | 0.631 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BUN(mmol/L) | 6.7±1.3 | 6.8±1.0 | 0.926 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LP(α)(mg/L) | 181.4±54.8 | 273.6±137.3 | 0.545 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hcy(μmol/L) | 18.9±3.2 | 17.2±2.6 | 0.741 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AST(U/L) | 23.8±2.7 | 24.5±2.8 | 0.860 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ALT(U/L) | 20.3±2.6 | 20.0±2.8 | 0.925 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| γGT(U/L) | 53.1±23.7 | 43.4±10.3 | 0.713 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CRP(mg/L) | 15.0±9.2 | 19.8±12.3 | 0.759 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| WBC count(×109/L) | 7.0±0.9 | 6.9±1.1 | 0.890 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hemoglobin(g/L) | 125.2±5.7 | 116.6±4.3 | 0.242 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Platelets(×109/L) | 177.3±17.2 | 262.9±59.0 | 0.177 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PT(s) | 12.5±1.0 | 11.4±0.2 | 0.317 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| APTT(s) | 33.7±1.5 | 31.9±1.1 | 0.355 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FIB(g/L) | 3.2±0.2 | 4.0±0.6 | 0.256 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: CAD: coronary artery disease; HT: hypertension; DM: diabetes mellitus; BMI: body mass index; SBP: systolic blood pressure; DBP: diastolic blood pressure; HBA1c: glycolated hemoglobin A1c; FBG: fasting blood glucose; TG: total triglyceride; TC: total cholesterol; LDL-C: low density lipoprotein cholesterol; HDL-C: high density lipoprotein cholesterol; BUN: blood urea nitrogen; LP(α): lipoprotein; Hcy: homocysteine; AST: aspartate aminotransferase; ALT: alanine aminotransferase; γGT: γ-aminobutyric acid; CRP: C-reactive protein; WBC: white blood cell; PT: prothrombin time; APTT: activated partial thromboplastin time; FIB: fibrinogen. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Expressions of miRNAs in AIS patients
Eight angiogenesis-associated miRNAs (miR-361 [29], miR-29a [30], miR-31 [26], miR-223-3p [27], miR-640 [31], miR-193a-3p [32], miR-191 [28], miR-503 [33]) levels were first measured in Cohort A. The expression of miR-361, miR-31, miR-223-3p, and miR-191 were changed in AIS patients when compared to the controls (Supplementary Figure 1B, 1C, 1E, 1F). These four miRNAs, including miR-31 (Figure 1A), miR-191 (Figure 1B), miR-223-3p (Figure 1C), and miR-361 (Figure 1D) were further detected in Cohort B. . However, only miR-191 was higher in AIS patients than those controls (Figure 1B). The Chart 1 showed the screening process.
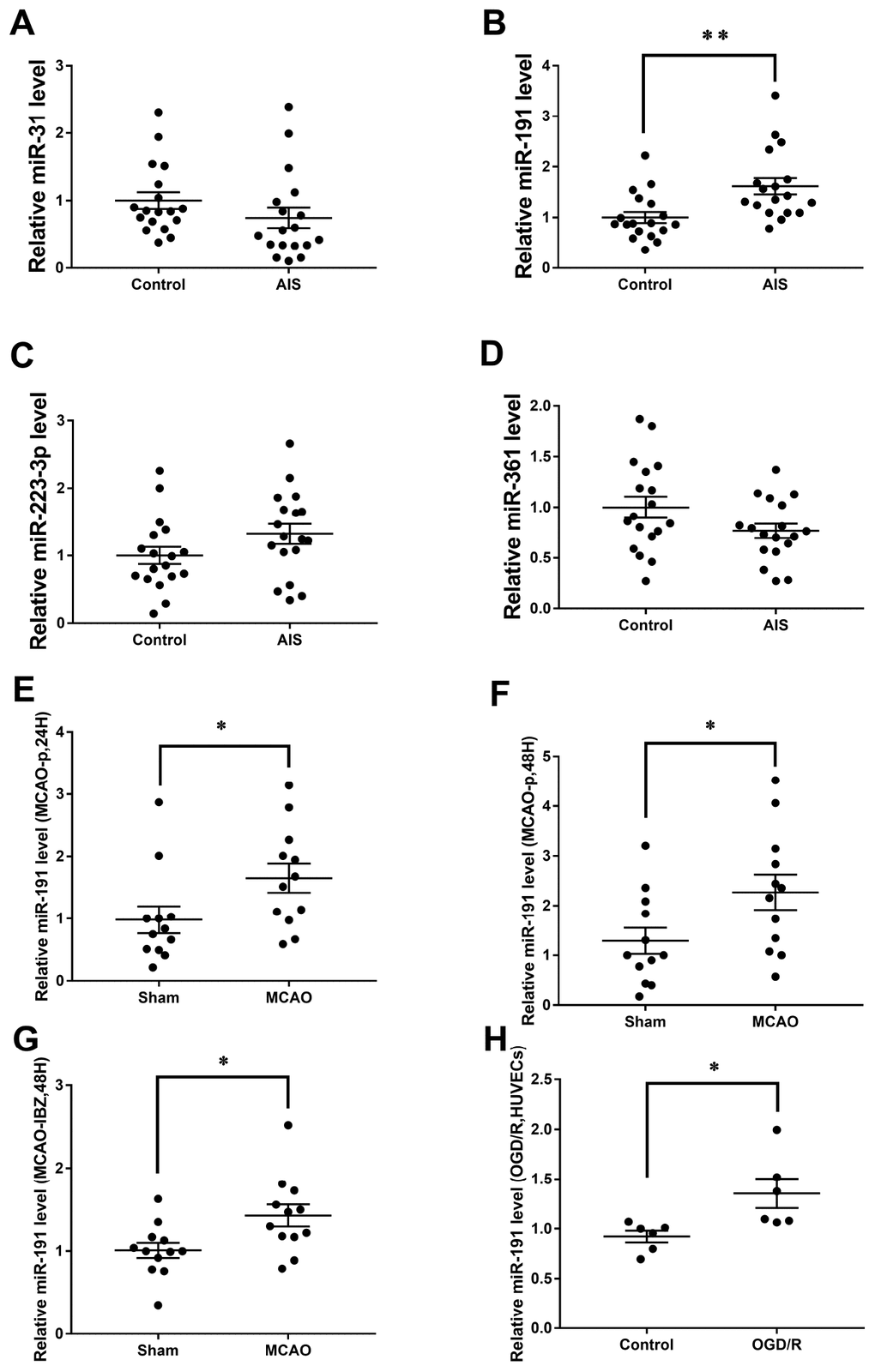
Figure 1. Relative miRNAs levels. Expression levels of miRNAs in Cohort A+B (n=18) (A) miR-31, (B) miR-191, (C) miR-223-3p, (D) miR-361; (E) Expression level of miR-191 in rat MCAO plasma after 24h reperfusion (n=12); (F) Expression level of miR-191 in rat MCAO plasma after 48h reperfusion (n=12); (G) Expression level of miR-191 in rat MCAO brains (n=12); (H) Expression level of miR-191 in OGD HUVECs (n=6). Means ± SEM. * P< 0.05,** P< 0.01 vs. NCm or NCi.
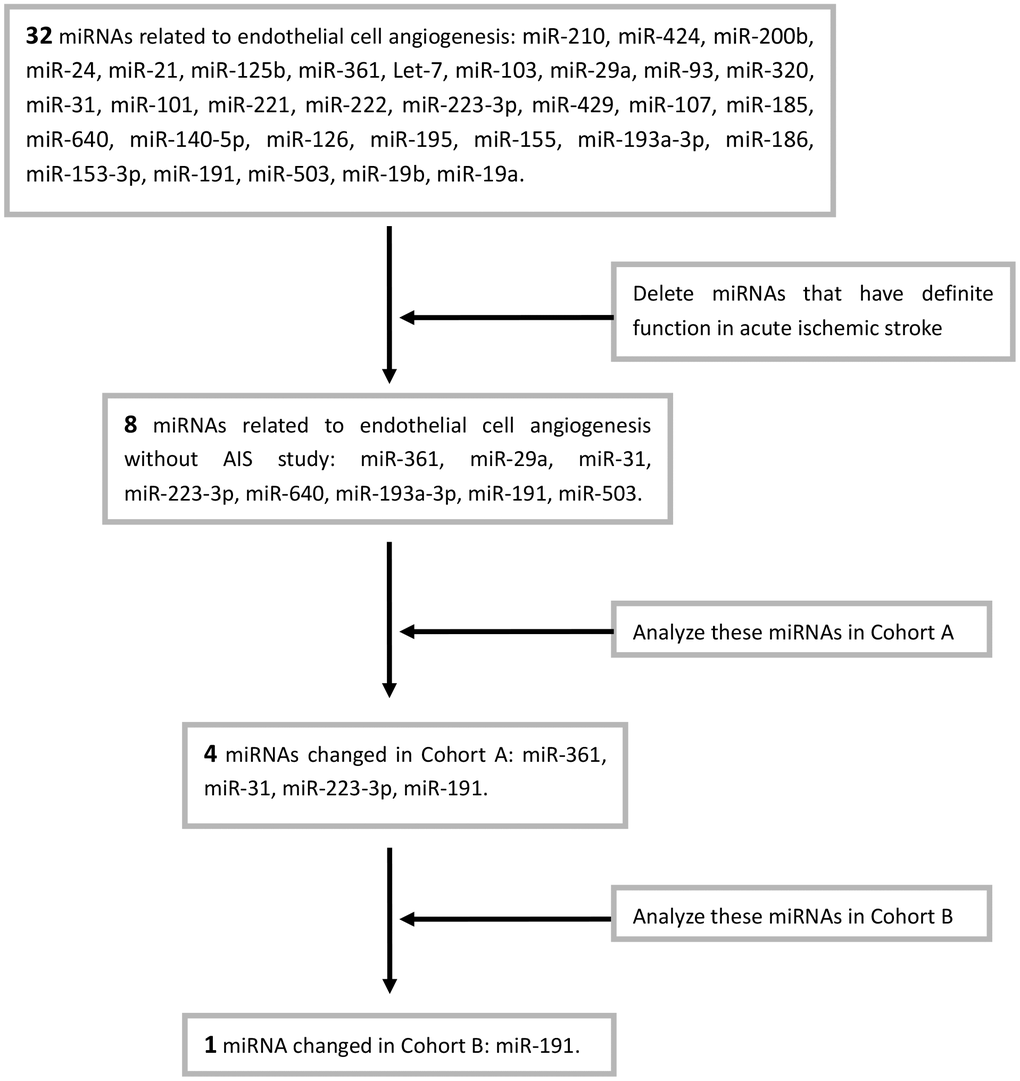
Chart 1. Screening process of miRNAs.
Expression of miR-191 in rat MCAO model and OGD/R HUVECs
Consistently, miR-191 levels were increased in the plasma of rat MCAO model both at 24h and 48h after reperfusion [14, 34] (Figure 1E, 1F). However, no significant difference of miR-191 levels was observed between the two time points (data not shown). The expression of miR-191 was also increased in the ischemic boundary zone (IBZ) (Figure 1G). We further detected miR-191 in HUVECs and found that miR-191 expression was elevated in OGD/R group (Figure 1H).
Function of miR-191 in HUVECs proliferation
We transfected HUVECs with 50nM miR-191 mimic and 100nM miR-191 inhibitor to up and down-regulate the expression of miR-191, respectively (Figure 2A, 2B). HUVECs were then subjected to reoxygenation for 18h after 2h of OGD. We found that up-regulation of miR-191 significantly reduced HUVEC proliferation (Figure 2C), while down-regulation of miR-191 promoted the proliferation (Figure 2D).
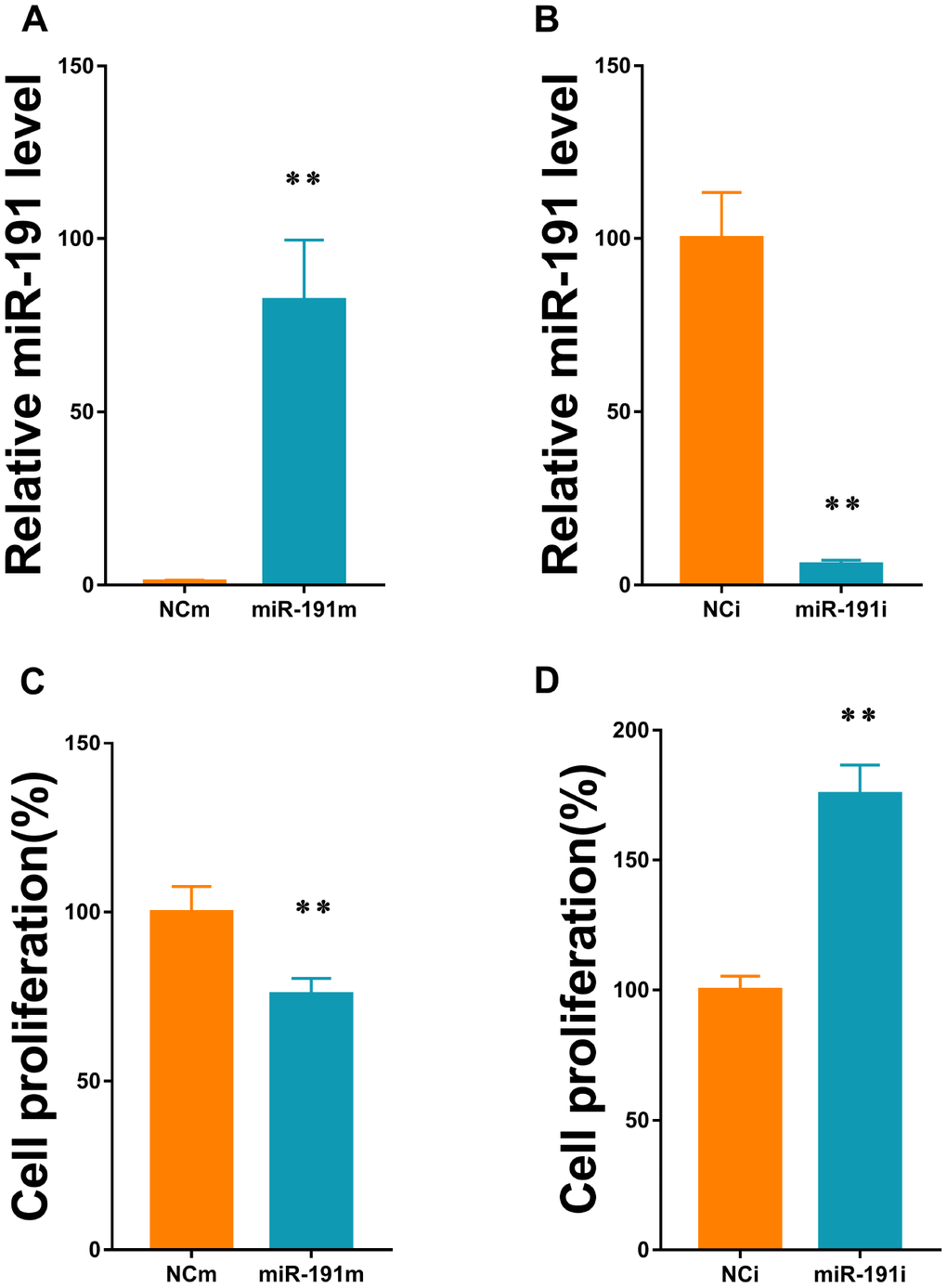
Figure 2. MiR-191 transfection efficiency and cell proliferation. (A, B) Expression level of miR-191 (in fold of NCm or percentage of NCi) in HUVECs that were transfected with miR-191 mimic (A) or miR-191 inhibitor (B) as assessed by real-time PCR (n=6 per group); (C, D) Proliferation (percentage of NCm or NCi) of HUVECs transfected with miR-191 mimic (C) or miR-191 inhibitor (D) as assessed by CCK-8 assay (n =6 per group). After transfection, the cells were reseeded into 96-well plates and incubated for another 48 h. Means ± SEM. ** P< 0.01 vs. NCm or NCi.
Function of miR-191 in HUVECs apoptosis and cell cycle
By using flow cytometry, we showed that up-regulation of miR-191 increased the apoptosis rate of HUVECs (Figure 3A, 3C), while down-regulation of miR-191 ameliorated the apoptosis induced by OGD/R (Figure 3B, 3D). We also found that over-expression of miR-191 blocked the cell cycle in the S phase (Figure 3E, 3G), which is consistent with the study of Gu, Y., et al. [35]. However, silence of miR-191 only slightly increased the number of G2 cells (Figure 3F, 3H).
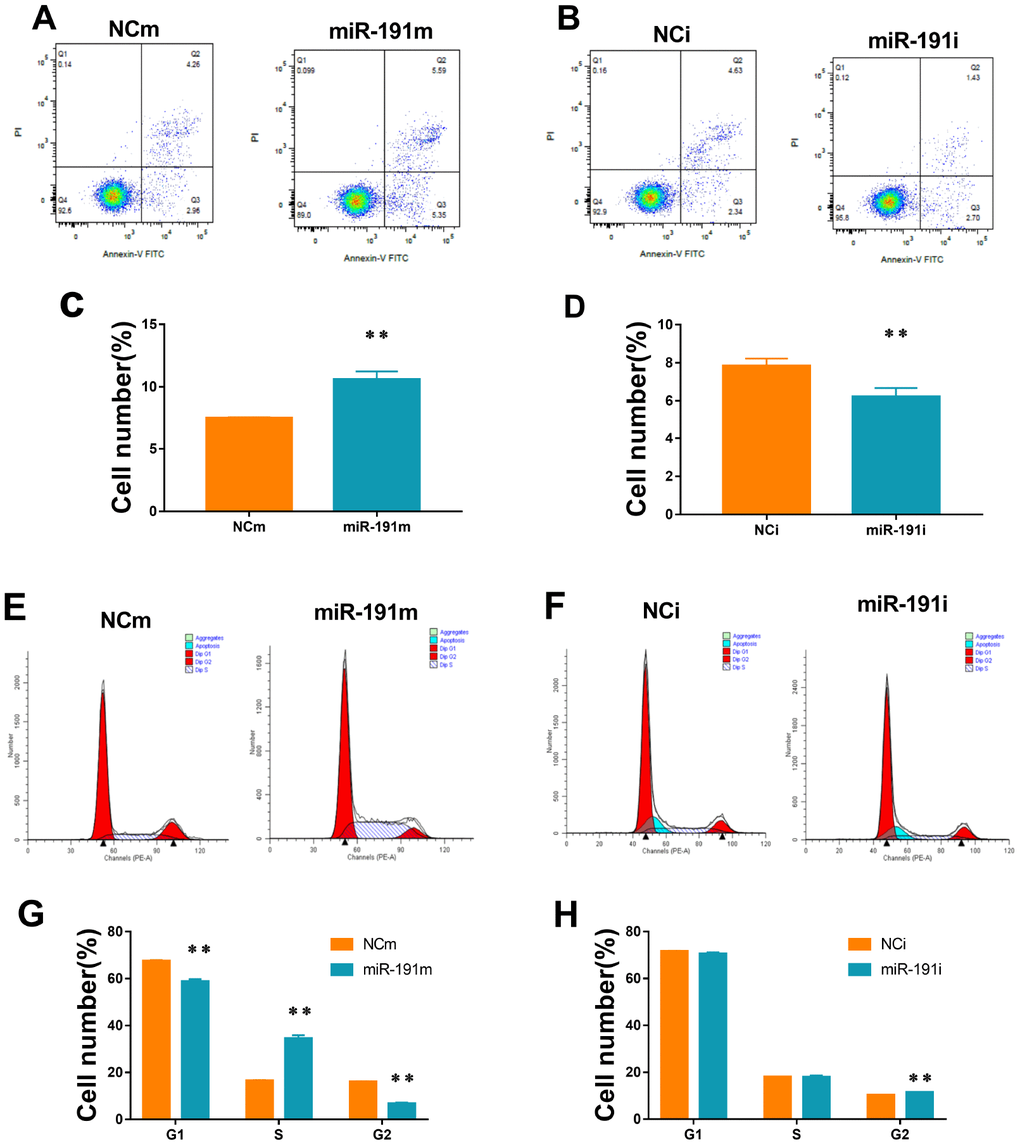
Figure 3. Cell apoptosis and cell cycle. (A, B) Cell apoptosis analysis of HUVECs transfected with miR-191 mimic (A) or miR-191 inhibitor (B)as assessed by flow cytometry; (C, D) Percentage of apoptosis HUVECs transfected with miR-191 mimic (C) or miR-191 inhibitor (D) (n =6 per group); (E, F) Cell cycle analysis of HUVECs transfected with miR-191 mimic (E) or miR-191 inhibitor (F) as assessed by flow cytometry; (G, H) Percentage of HUVECs transfected with miR-191 mimic (G) or miR-191 inhibitor (H) in different stages (n=6 per group). Means ± SEM.,** P< 0.01 vs. NCm or NCi.
Function of miR-191 in HUVECs migration
To investigate the function of miR-191 in HUVECs migration in OGD/R, scratch wound healing assay and transwell migration assay were performed. Over-expression of miR-191 significantly delayed the closure of scratch wounds (Figure 4A, 4C) and markedly reduced the number of migrated cells (Figure 4E, 4G). In contrast, inhibition of miR-191 promoted the healing of scratch wounds (Figure 4B, 4D) and enhanced cell migration (Figure 4F, 4H).
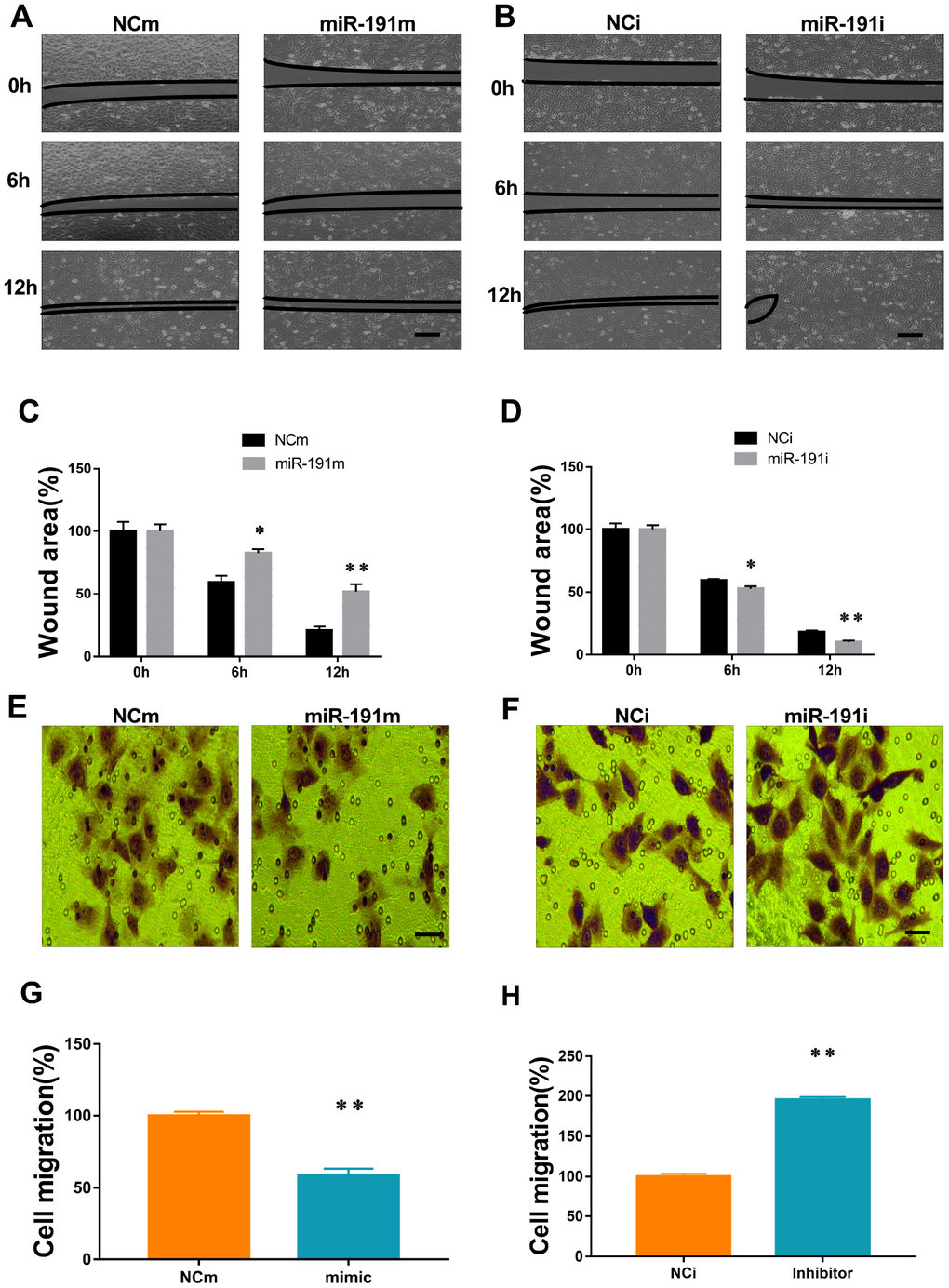
Figure 4. MiR-191 inhibited cell migration. (A, B) Phase contrast microscopic images of HUVECs at 0, 6, and 12 h after scratching. The cells were transfected with miR-191 mimic (A), miR-191 inhibitor (B), or the corresponding scrambled NCm (A) and NCi (B). Black lines indicate the wound area. Scale bars, 100 μm. (C, D) Size of wound area (percentage of 0 h) created by scratching HUVECs transfected with miR-191 mimic (C) or miR-191 inhibitor (D) (n =6 per group). (E, F) Phase contrast microscopic images of HUVECs migrated and attached to the bottom membrane of a transwell. The cells were transfected with miR-191 mimic (E), miR-191 inhibitor (F), or the corresponding scrambled NCm (E) and NCi (F). Scale bars, 20μm. (G, H) Number of migrated HUVECs (percentage of NCm or NCi) transfected with miR-191 mimic (G) or miR-191 inhibitor (H) (n =6 per group). Means ± SEM. * P< 0.05,** P< 0.01 vs. NCm or NCi.
Function of miR-191 in HUVECs tube-forming activity
We found that transfection of HUVECs with miR-191 mimic reduced the number of newly developed tube meshes when compared with controls (Figure 5A, 5C). In contrast, transfection of cells with miR-191 inhibitor promoted tube formation (Figure 5B, 5D)
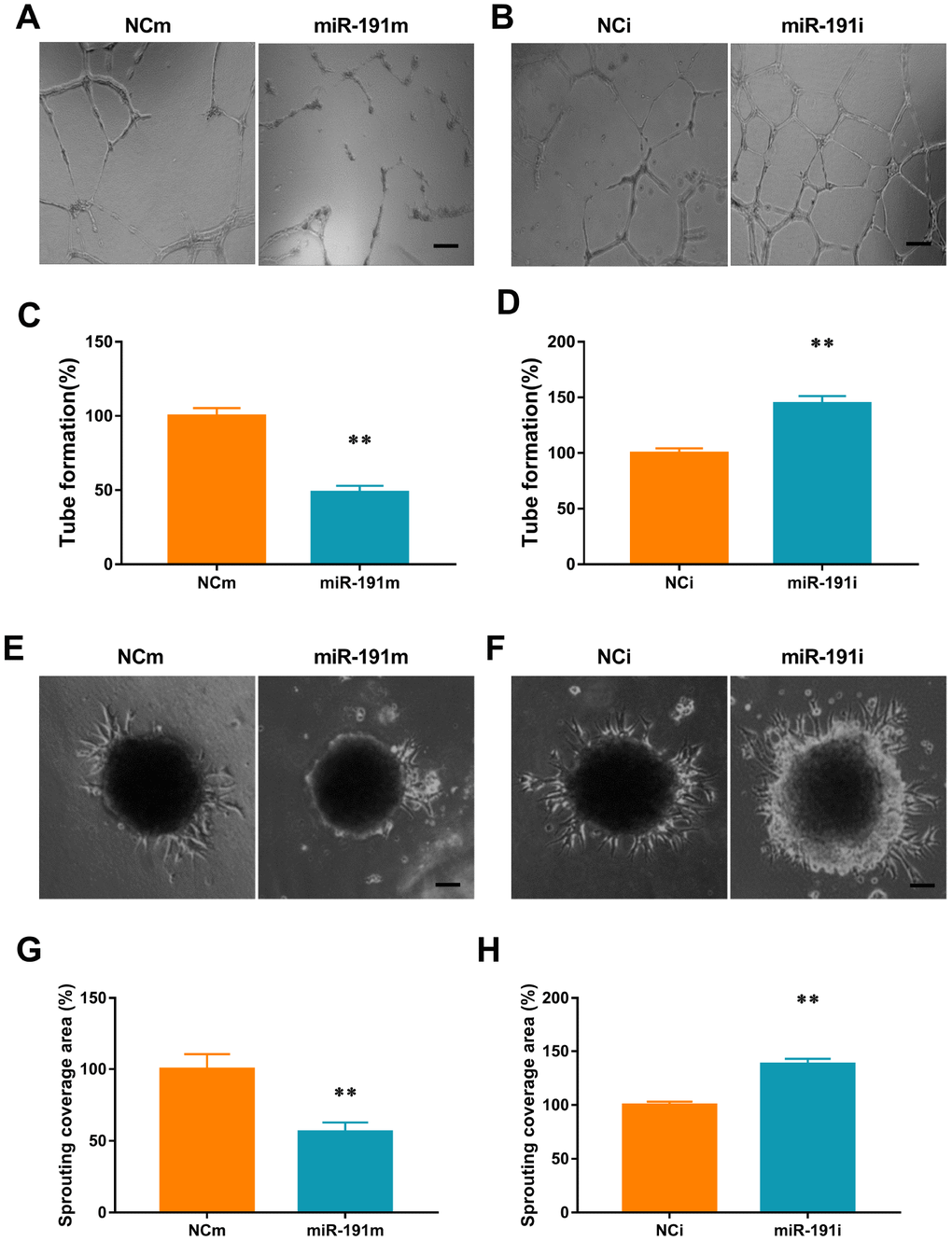
Figure 5. miR-191 inhibited tube formation and spheroid sprouting. (A, B) Phase-contrast microscopic images of tubeforming HUVECs that were transfected with miR-191 mimic (A), miR-191 inhibitor (B), or the corresponding scrambled NCm (A) and NCi (B). Scale bars, 20μm. (C, D) Tube formation of HUVECs (percentage of NCm or NCi) transfected with miR-191 mimic (C) or miR-191 inhibitor (D) as assessed by tube formation assay (n =6 per group). (E, F) Phase-contrast microscopic images of sprouting HUVECs spheroids. HUVECs were transfected with miR-191 mimic (E), miR-191 inhibitor (F), or the corresponding scrambled NCm (E) and NCi (F). Scale bars, 20μm. (G, H) Sprouting coverage area of HUVECs (percentage of NCm or NCi) transfected with miR-191 mimic (G) or miR-191 inhibitor (H) as assessed by spheroid sprouting assay (n = 6 per group). Means ± SEM. ** P< 0.01 vs. NCm or NCi.
Function of miR-191 in HUVECs spheroid sprouting activity
We performed a 3-dimensional spheroid sprouting assay and demonstrated that over-expression of miR-191 significantly decreased the sprouting coverage area of HUVECs spheroids when compared with controls (Figure 5E, 5G). In contrast, transfection of cells with miR191 inhibitor markedly enhanced the sprouting coverage area (Figure 5F, 5H).
Validation of predictive target gene of miR-191
Vascular endothelial zinc finger 1 (VEZF1) is one of the target genes of miR-191 predicted by TargetScan 7.2 and miRDB (Figure 6A). We found that VEZF1 mRNA levels were not influenced by miR-191 mimic or miR-191 interference (Figure 6B). Since VEZF1 is a nucleus transcription factor, we then extracted nucleoprotein and found that over-expression of miR-191 decreased VEZF1 protein levels in nucleus, while miR-191 inhibitor increased the protein levels of VEZF1 (Figure 6C, 6D). Further luciferase assay showed a significant decrease in the luciferase activity of wild-type VEZF1 3’ UTR (Figure 6E, 6F), indicating that VEZF1 is the target of miR-191.
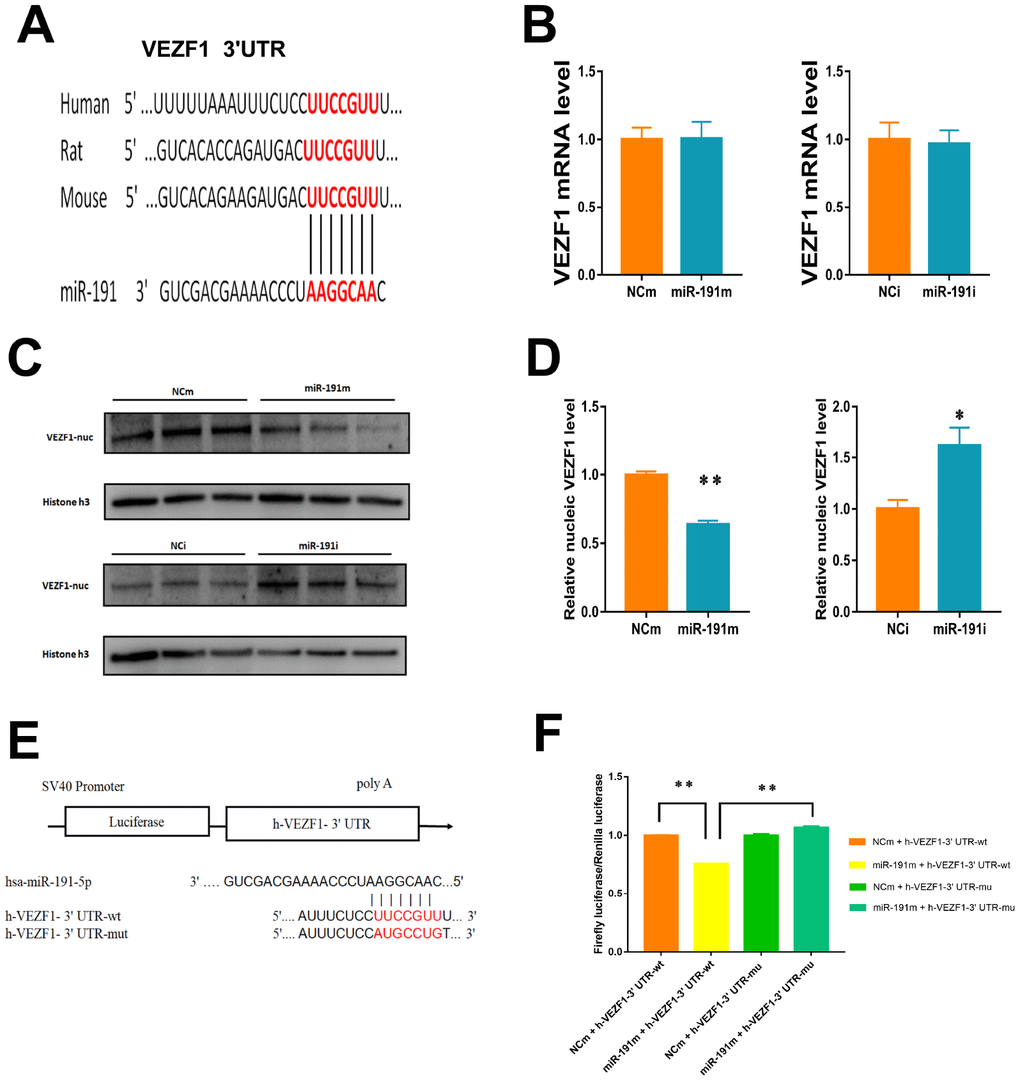
Figure 6. Regulation of VEZF1 by miR-191. (A) MiR-191 and its putative binding sequence in the 3'-UTR of VEZF1. (B) Real-time PCR analysis of VEZF1 expression in HUVECs by miR-191 interference. (n =6 per group); (C, D) Western blot analysis of VEZF1 expression in HUVECs transfected by miR-191. (E) Construction of Plasmid; (F) Luciferase activities were measured to evaluate the binding of miR-191 to the candidate binding sequence of VEZF1 after transfection of miR-191 mimic or NCm. Means ± SEM. * P< 0.05,** P< 0.01 vs. NCm or NCi.
Action of miR-191 on VEZF1 signaling
VEZF1 has been reported to regulate several genes that are involved in angiogenesis, such as endothelin 1 (EDN1) [36], matrix metalloproteinase 2 (MMP2) [36], stathmin 1 (STMN1) [37], matrix metalloproteinase (MMPs) [38], Cbp/p300 interacting transactivator with Glu/Asp rich carboxy-terminal domain 2 (CITED2) [39]. In the present study, we found that miR-191 over-expression significantly suppressed the mRNA levels of EDN1, MMP1, and STMN1 and but increased the mRNA level of CITED2 (Figure 7A, 7C, 7E, 7G). Inhibition of miR-191 showed an opposite trend of the changes of these genes (Figure 7B, 7D, 7F, 7H). We did not find any effects of miR-191 on the mRNA levels of MMP2 and MMP9 (Figure 7I–7L).
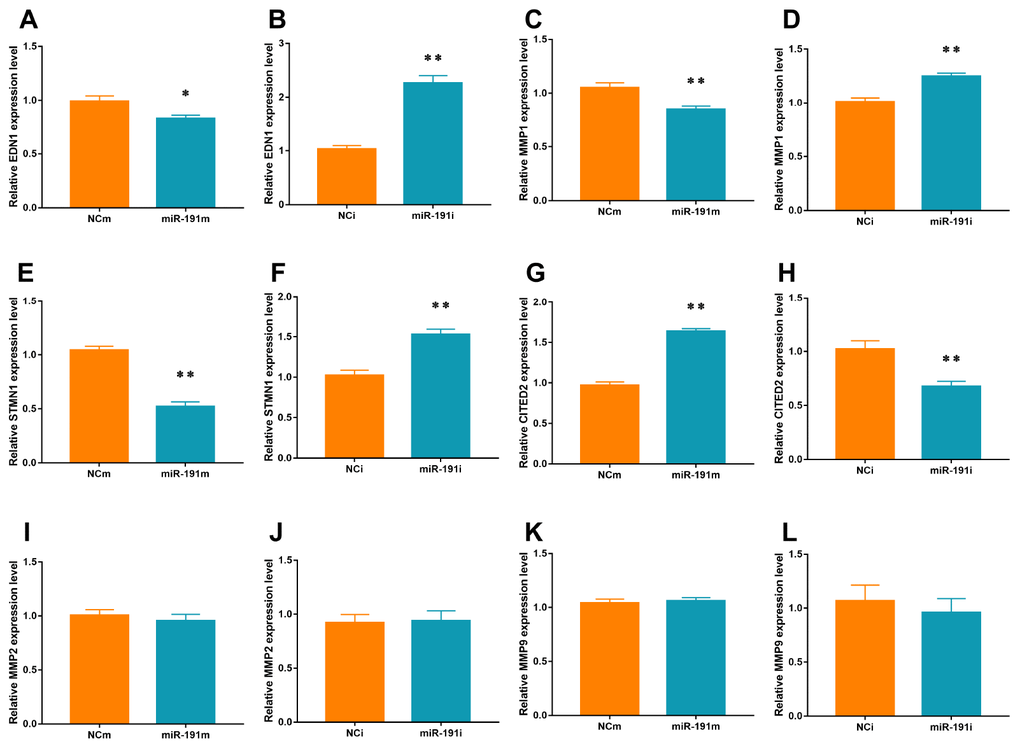
Figure 7. mRNA levels of Targets of VEZF1. Real-time PCR for EDN1 (A, B), MMP1 (C, D), STMN1 (E, F), CITED2 (G, H), MMP2 (I, J), and MMP9 (K, L) mRNA of HUVECs transfected with miR-191 mimic or miR-191 inhibitor (n = 6 per group). Means ± SEM. * P< 0.05,** P< 0.01 vs. NCm or NCi.
Inhibition of miR-191 reduced infarction volume of MCAO rats
The rats randomly received an intracerebroventricular infusion of miR-191/NC antagomir or blank control 3 days prior to MCAO. Compared to the NC antagomir, miR-191 antagomir significantly reduced the miR-191 levels both in plasma and IBZ at 48 h after reperfusion in MCAO rats (Figure 8A, 8B). We found that VEZF1 mRNA levels of IBZ were not influenced by miR-191 antagomir (Figure 8C) which was consistent with the results of cell experiments. However, the protein levels of VEZF1 were increased significantly (Figure 8D, 8E). We also found that rats receiving miR-191 antagomir had smaller brain infarct volumes than those with NC antagomir (Figure 8F, 8G).
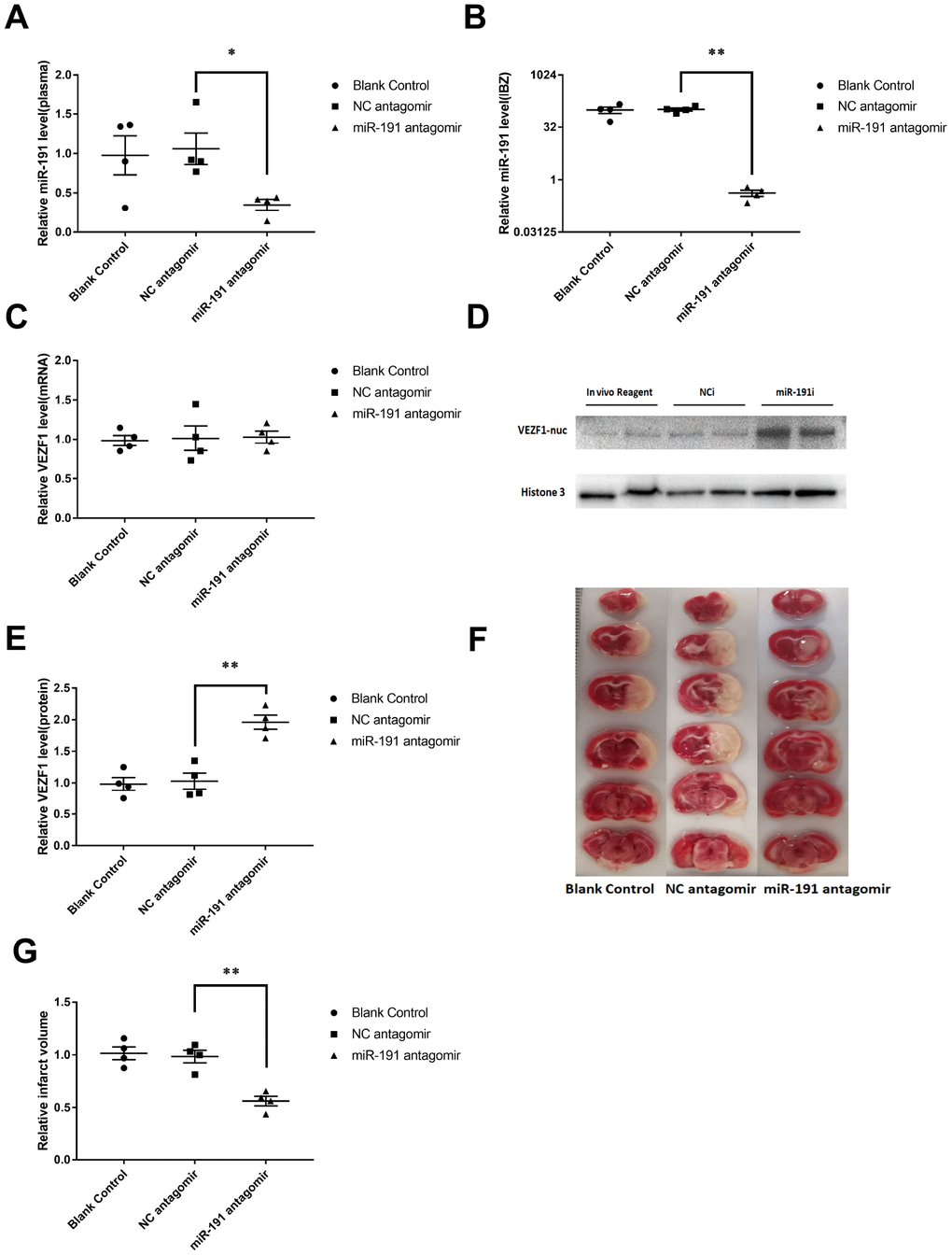
Figure 8. Inhibition of miR-191 reduced infarction volume of MCAO rats after the injection of miR-191 antagomir compared to NC antagomir and blank control. Relative miR-191 levels in (A) plasma and (B) IBZ (n =4 per group); (C) Real-time PCR analysis of VEZF1 expression in IBZ (n =4 per group); (D, E) Western blot analysis of VEZF1 expression in IBZ (n =4 per group); (F, G) Relative infarct volume of MCAO rats (n =4 per group). Means ± SEM. * P< 0.05,** P< 0.01 vs. NC antagomir.
Discussion
Impaired angiogenesis plays a crucial role in cerebral injury after acute ischemic attack [6]. MiRNAs have been shown to be important regulators involved in the process of angiogenesis [10, 40]. Here we showed for the first time that miR-191 was elevated in the plasma of AIS patients as well as in MCAO rat model and OGD/R HUVEC model. Over-expression of miR-191 promoted apoptosis but inhibited proliferation, migration, tube-forming and spheroid sprouting activity in HUVECs, while silence of miR-191 displayed opposite results. Mechanistically, we found that miR-191 directly regulated VEZF1, leading to the changes of a variety of angiogenesis-associated genes targeted by VEZF1. In vivo studies demonstrated that Inhibition of miR-191 could reduce the infarction volume induced by MCAO in rats. Therefore, miR-191 might be a novel therapeutic target for the treatment of AIS.
Angiogenesis is one of the key repair mechanisms for the ischemic injury induced by acute stroke [7]. Gu et al. [35] demonstrated that miR-191 was preferentially expressed in endothelial cells compared to other types of human cells and displayed antiangiogenic effect. In the present study, we performed a series of well-established angiogenesis assays and demonstrated that miR-191 is an inhibitor of angiogenesis with effects of suppressing proliferation, migration, tube formation and spheroid sprouting in HUVECs. Knockdown of miR-191 could reduce the infarction area induced by MCAO in rats and promote proliferation, migration, tube formation and spheroid sprouting in HUVEC. Our results complement nicely with a previous report showing that knockout of miR-191 reduced hepatic ischemia-reperfusion injury through inhibiting inflammatory responses and cell death [41]. Another study also showed that up-regulated miR-191 participated in renal ischemia-reperfusion injury via inducing apoptosis of renal tubular epithelial cells [42]. These results indicate that lowing miR-191 might be a potential therapy for ischemia-reperfusion injury. However, although we found that plasma levels of miR-191 were increased in patients with AIS, there might be false positive and negative results in the process of miRNA screening because of the relative small sample size. Future studies with larger sample size will be needed to verify the exact roles of miR-191 in the diagnosis and prediction of AIS.
VEZF1 encodes a zinc finger transcription factor which is essential for developmental angiogenesis and lymphangiogenesis. Mammalian VEZF1 is expressed in the anterior-most mesoderm at E7.5 during development and is later restricted in the vascular endothelium [43]. VEZF1 knockout mice showed embryonic lethality caused by vascular remodeling defects and loss of vascular integrity, indicating that VEZF1 is a critical regulator of vascular development [44]. VEZF1 is thus proposed to act as a transcriptional activator of pro-angiogeneic genes including EDN1 [36], MMP2 [36], STMN1 [37], MMPs [38], CITED2 [39]. VEZF1 can be epigenetically regulated by histone acetylation and deacetylation [43]. VEZF1 is specifically expressed in endothelial cells and correlated with the differentiation and proliferation of endothelial cells in the embryonic vascular system [45]. A series of studies showed that VEZF1 could activate angiogenesis by promoting endothelial cells proliferation, migration and vessel network formation [37, 39, 46, 47]. Our data demonstrated that VEZF1 is regulated by miR-191 at post-translational level. Luciferase reporter assay validated that VEZF1 is a direct target of miR-191. To further prove miR-191 inhibit HUVECs angiogenesis by targeting VEZF1, we measured the expression levels of the VEZF1 targets [36–39] after miR-191 interference. Our data shown that over-expression of miR-191 inhibited the expression of angiogenesis-related genes including EDN1, MMP1, and STMN1. However, further intervention studies are needed to elucidate whether miR-191 inhibited angiogenesis via targeting VEZF1.
In conclusion, our data reveal a novel role of miR-191 in promoting ischemic brain injury through inhibiting angiogenesis via targeting VEZF1, which in turn resulted in up-regulation of CITED2 and down-regulation of MMP-1, STMN1.(Figure 9). MiR-191 might be served as a promising effective biomarker and therapeutic target for AIS.
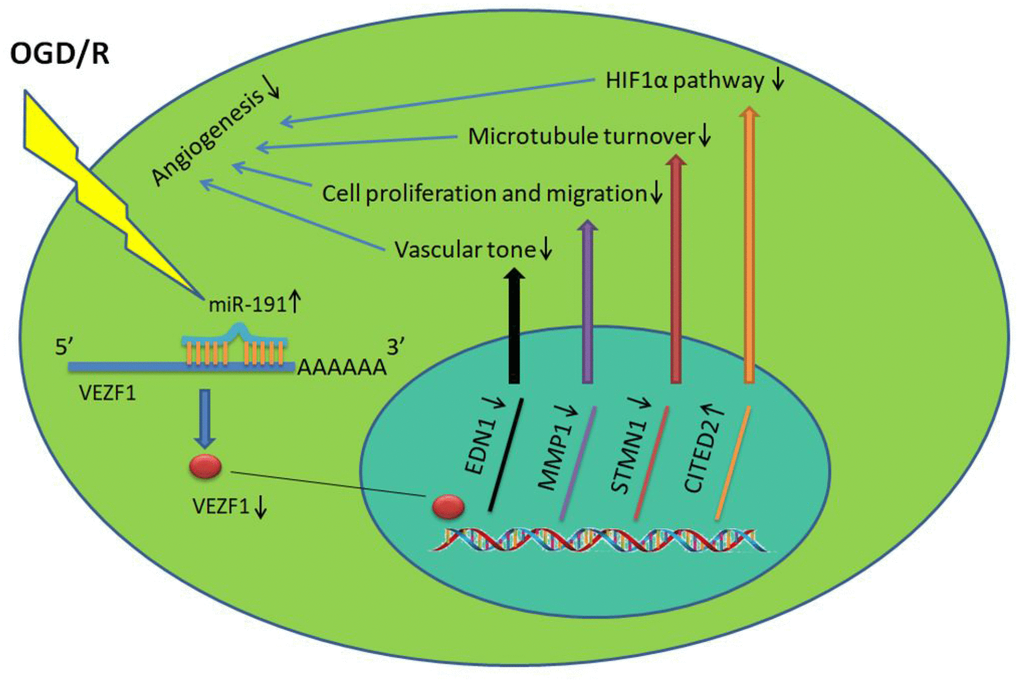
Figure 9. miR-191 reduces the nucleoprotein VEZF1, resulting in up-regulation of CITED2 and down-regulation of MMP-1, STMN1 mRNA. This, in turn, suppresses HUVEC proliferation and migration and thus prevents endothelial tube formation and spheroid sprouting. Moreover, miR-191 suppresses the expression of EDN1 which functions as maintaining vascular tension.
Methods
Ethics statement
Investigation was conducted in accordance with the ethical standards and according to the Declaration of Helsinki and was approved by the Ethics Committee of Sir Run Run Hospital, Nanjing Medical University (Protocol Numbers: 2018-SR-25, Supplementary Figure 2) and the Animal Ethical and Welfare Committee of Nanjing Medical University (Protocol Numbers: IACUC-1806010, Supplementary Figure 3).
Patient enrollment
Patients were recruited consecutively from the department of geriatrics of Sir Run Run Hospital, Nanjing Medical University from January to June 2018. There were total 18 patients with AIS and 18 controls. AIS patients were recruited from stroke center within six hours from the onset of the symptoms before thrombolytic therapy. AIS diagnosis was confirmed using clinical features and brain MRI by two investigators in a double-blinded manner according to the 2018 Guidelines for the Early Management of Patients With Acute Ischemic Stroke [2]. The control subjects were selected during the same period in the same hospital from the health examination center. Exclusion criteria including: under 18 or over 90 years old; history of intracranial hemorrhage; craniocerebral trauma; giant intracranial aneurysms; recent (within 3 months) history of intracranial surgery; malignant tumors. Informed consents were obtained from all subjects.
MCAO
Male Sprague Dawley rats (7-8w, 230 g–280 g) randomly divided into two groups (12 in each group) were obtained from Shanghai Sippr-BK laboratory animal Co. Ltd. Right MCAO was induced using an intraluminal filament as Longa, E.Z. et al [48] described. The body temperature was maintained at 37 °C with a homothermal blanket and physiological parameters were monitored during the surgical procedure. After anesthetized with 1 % pentobarbital sodium (0.5ml/100g), the right common carotid artery (CCA), external carotid artery (ECA) and internal carotid artery (ICA) were sequentially isolated. An incision was made in the distal region of the CCA, then a 40mm long (diameter: 0.26-0.28mm) poly-L-lysine coated nylon monofilament (Beijing Shadong Biotechnology Co., Ltd.) was inserted into the ICA, and the monofilament was advanced approximately 18-20mm beyond the carotid bifurcation until mild resistance was encountered. The occlusion was sustained for 2 h. The sham group underwent similar procedures, but the monofilament was not advanced into the CCA.
Neurological evaluations were performed according to an established graded scoring system at 24 h after reperfusion to verify the modeling success in function [49]. Briefly, neurological deficits were scored as follows: 0, no deficit; 1, failure to extend left forepaw upon lifting the whole animal by tail; 2, grip strength weakening of the left forepaw; 3, circling to the left when held by the tail; and 4, spontaneous circling.
Peripheral venous blood was collected at 24 and 48 hours after reperfusion and the brains were removed and frozen at −20 °C for 15 min at 48 hours after reperfusion when sacrificed (Supplementary Figure 3). To histologically verify the success of the model, coronal sections were cut into 2 mm thick slices, stained with 1% 2,3,5-triphenyltetrazolium chloride (TTC) at 37 °C in the dark for 20 min, and photographed (Supplementary Figure 4).
Plasma and brain fragments collection and storage
Peripheral blood samples were collected in tubes anticoagulated with ethylenediamine tetraacetic acid (EDTA) dipotassium salt after a definite diagnosis before any medication is administered. The samples were centrifuged (1000×g, 5minutes, 4 °C, Beckman Coulter) to remove blood cells and debris, and then transferred to 1.5ml microtubes (Axygen, MCT-150-C) for storage at –80°C until further processing. Brain fragments were collected and stored in liquid nitrogen.
Primary HUVECs isolation and culture
Fresh umbilical cords were obtained following delivery of healthy babies to healthy mothers. HUVECs were isolated from umbilical cords according to a previously described method [50]. Briefly, the umbilical vein was inserted an intravenous needle for single use with tube and flushed with 30ml 37 °C phosphate buffer saline (PBS), following which the cord was clamped at the distal end and the vessel filled with collagenase (Type Ia, 1 mg/mL, Sigma, C9891), until mildly distended. Following incubation at 37°C for 10-15 min, the cord was unclamped and the digest drained. The vessel was gently massaged and flushed through with 20ml endothelial cell medium (ECM, Science, 1001), and the digests were pooled. The endothelial cell suspension was centrifuged (500×g, 5 min), and the cell pellet was resuspended in ECM. This suspension was seeded into a 25ml flask (Corning, 430639).
The cells were incubated at 37°C, 5% CO2 until they are 85% - 90% confluent. Then, cells were detached from the substratum by exposure to 0.25% trypsin-EDTA (Gibco, 25200072, US) for 1 min at 37°C, pelleted (500×g, 5 min) and passaged at a ratio of 1:3-5. Confluent cells at passage 3-6 were used for all experiments.
OGD/R
The OGD/R protocol was performed to mimic ischemia in vitro. Briefly, culture medium was removed and rinsed with PBS for three times. HUVECs were placed into a tri-gas incubator (memmert, Eastern Friesland, Germany) containing 1% O2, 5% CO2, 94% N2 at 37 °C with glucose-free Dulbecco's Modified Eagle Medium (DMEM, Gibco, 11966025, US). After two hours challenge, DMEM was replaced with ECM. The cells were maintained for further 24 h at 37 °C in a humidified 5% CO2 incubator to generate reperfusion.
Total RNA isolation
Total RNA was isolated from plasma, brain and cell samples using TRIzon (CWBio, CW0580) reagent following the manufacturer’s instructions. RNA concentration and purity were determined with one drop spectrophotometer (OD-1000+, Nanjing wuyi Science and Technology Co., Ltd., China).
Quantitative real-time polymerase chain reaction (qRT-PCR)
CDNA was generated from 1 μg RNA using miRNA 1st Strand cDNA Synthesis Kit (by stem-loop) (vazyme, MR101-02, Nanjing, China) for miRNA or PrimeScript™ RT reagent Kit (Perfect Real Time) (Takara, RR047A, Japan) for mRNA. Real-time PCR was performed using QuantStudio 5 (Applied Biosystems, US) with miRNA Universal SYBR qPCR Master Mix (vazyme, MQ101-02) for miRNA or Maxima SYBR Green/ROX qPCR Master Mix (2X) (Thermo Scientific™, K0221, US) for mRNA according to the manufacturer’s protocol. All reactions were run in triplicate and relative gene expression was calculated using the comparative threshold cycle (Ct) method (relative gene expression = 2− (ΔCtsample-ΔCtcontrol)). The U6 snRNA was used as an internal control for miRNA while TATA-binding protein (TBP) used for mRNA. The gene-specific primers sequences are listed in Table 5.
Table 5. Primer sets for real-time PCR analyses.
| Gene | Forward primer (5′ to 3′) | Reverse Primer (5′ to 3′) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| U6 (MIM:180692) | CTCGCTTCGGCAGCACA | AACGCTTCACGAATTTGCGT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TBP (NM_003194) | CCACTCACAGACTCTCACAAC | CTGCGGTACAATCCCAGAACT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| VEZF1 (NM_007146) | GGACAGCTATCACCTGAGGC | GCGATGGTAGAGATAAGGGGAA | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MMP1 (NM_002421) | AAAATTACACGCCAGATTTGCC | GGTGTGACATTACTCCAGAGTTG | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MMP2 (NM_004530) | TACAGGATCATTGGCTACACACC | GGTCACATCGCTCCAGACT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MMP9 (NM_004994) | TGTACCGCTATGGTTACACTCG | GGCAGGGACAGTTGCTTCT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EDN1 (NM_001168319) | AGAGTGTGTCTACTTCTGCCA | CTTCCAAGTCCATACGGAACAA | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CITED2 (NM_006079) | CCTAATGGGCGAGCACATACA | GGGGTAGGGGTGATGGTTGA | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| STMN1 (NM_203401) | TCAGCCCTCGGTCAAAAGAAT | TTCTCGTGCTCTCGTTTCTCA | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miRNA | Accession | Reverse transcription primers (5′ to 3′) | Forward primers (5′ to 3′) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-29a | MIMAT0004503 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTGAAC | GCGCGACTGATTTCTTTTGGT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-31 | MIMAT0000089 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGCTAT | GCGAGGCAAGATGCTGGC | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-138 | MIMAT0000430 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCGGCCT | GCGAGCTGGTGTTGTGAATC | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-191 | MIMAT0000440 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCAGCTG | CGCAACGGAATCCCAAAAG | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-193a-3p | MIMAT0000459 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACACTGGG | CGCGAACTGGCCTACAAAGT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| mir-223-3p | MIMAT0000280 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTGGGGT | GCGCGTGTCAGTTTGTCAAAT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-361 | MIMAT0000703 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACGTACCC | GCGCGTTATCAGAATCTCCAG | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-503 | MIMAT0002874 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTGCAG | CGTAGCAGCGGGAACAGTT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-640 | MIMAT0003310 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGAGGC | CGCGATGATCCAGGAACCT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviation: TBP: TATA-box binding protein; MMP1/2/9: matrix metallopeptidase 1/2/9; EDN1: endothelin 1; CITED2: Cbp/p300 interacting transactivator with Glu/Asp rich carboxy-terminal domain 2; STMN1: stathmin 1. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Transfection of miR-191 mimic or inhibitor into HUVECs
The cells were transfected for 24 h with 50 nM miR-191 mimic (Ribobio, Guangzhou, China) or 100 nM miR-191 inhibitor (Ribobio) using Lipofectamine 3000(Invitrogen, US) according to the manufacturer’s protocol. Cells transfected with negative control of mimic (NCm) (Ribobio) or negative control of inhibitor (NCi) (Ribobio) served as controls.
Cell proliferation assay
To assess the proliferation rate of HUVECs, Cell Counting Kit-8(CCK-8) assays (Dojindo, Japan) were performed according to the manufacturer’s instructions. Briefly, 3000 HUVECs in 100 μl of cell suspension were seeded in 96-well flat-bottomed plates. After 8 hours incubation, CCK-8 reagent was added to each well, and the absorbance of each well was measured at 450 nm after 3 hours incubation by a microplate reader (Synergy H1, BioTek, US). A value of 100% was assigned to the respective control group.
Flow cytometric analysis of apoptosis
HUVECs were treated with miR-191mimic/ihibitor for 24 h, and then subjected to OGD/R for 2 h/18 h. The apoptotic cell death rate was examined with Annexin V-FITC and PI double staining using the Annexin V-FITC apoptosis detection kit (Beyotime Biotechnology, C1063, Shanghai, China) according to the manufacturer’s instructions. Briefly, 5×104 cells were collected with 0.25% EDTA free trypsin (Gibco, 15050065, US), washed and resuspended in PBS. After staining with Annexin V-FITC/PI, flow cytometric analysis was performed and data were analyzed using FlowJo software.
Flow cytometric analysis of cell cycle
HUVECs were treated with miR-191 mimic/inhibitor for 24 h, and then subjected to OGD/R for 2 h/18 h. HUVECs were collected and fixed in 70% ethanol overnight at 4 °C by using the cell cycle detection kit (KeyGen BioTech, KGA512, Jiangsu, China). Single-cell suspensions were labeled with PI for 30 min at 4 °C and analyzed by flow cytometry (BD, New York, US). The data were analyzed with FlowJo software.
Cell migration assay
Scratch wound healing assay and transwell migration assay were performed to evaluate the motility of HUVECs. For the scratch wound healing assay, 5×105 of HUVECs were seeded in a 6-well plate. After reaching confluence, the cell monolayer was scratched with a pipette tip (10 μl) to generate 4 scratch wounds and then rinsed twice with PBS to remove nonadherent cells. Phase-contrast light micrographs were taken immediately after scratching (0 h) as well as after 6h and 12 h with ×200 magnification using a CKX41 microscope (Olympus, Japan).
For the transwell migration assay, 2×105 of HUVECs in 500μl 1% FBS ECM were seeded into a 24-well insert (costar, 3422), and 750μl of 15% FBS ECM was added to the lowerwell. After 24h of incubation, nonmigrated cells were removed with cotton swabs, and migrated cells were stained with 0.1% crystal violet (Solarbio, C8470, Beijing, China). The number of migrated cells was determined in 3 microscopic regions of interest at ×200 magnification using a CKX41 microscope.
Tube formation assay
To analyze the function of miR-191 in the tube-forming activity of HUVECs, 50 μl of Matrigel (8-12mg/ml; Corning) was plated in each well of a 96-well plate. After Matrigel polymerization, 3×104 of HUVECs in 100μl ECM were added into each well. After 12 h, vessel-like network structures were examined under CKX41 microscope. Tube formation was quantified by measuring the number of meshes using ImageJ software [National Institutes of Health (NIH), Bethesda, MD, USA].
Spheroid sprouting assay
HUVECs were suspended in ECM containing 0.25% (w/v) methylcellulose (Sigma-Aldrich) and seeded (1000 cells/100μl) in low attachment, round-bottom, 96-well spheroid microplates (Corning, 4520). After incubation for 24 h, spheroids were harvested and resuspended in 20μl Matrigel. The spheroid-containing Matrigel was rapidly transferred to 24-well plates and allowed to polymerize for 30 min, after which 500μl ECM was added to each well. After 24h of incubation, the spheroid-sprouting capacity was quantified by measuring the sprouting coverage area of the sprouts using imageJ software.
Luciferase reporter assays
293T cells (3 × 105 cells per well) were plated onto 24-well plates. pSV40-VEZF1-wt or pSV40-VEZF1-mut was cotransfected with miRNA-191 mimic or NCm into 293T cells by Lipofectamine 3000. The relative luciferase activity was normalized to Renilla luciferase activity 48h after transfection.
Protein extraction and Western blots
Nuclear extracts were prepared with NE-PER™ Nuclear and Cytoplasmic Extraction Reagents according to the manufacturer's protocol (Thermo Scientific, 78833, US). Protein concentrations were measured by BCA protein assay (Beyotime Biotechnology, Shanghai, China). Equal amounts (20 μg) of protein were separated by 4-20% GenScript SurePAGE, Bis-Tris, precast polyacrylamide gels (GenScript Biotechnology, Nanjing, China) electrophoresis and transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). Membranes were then incubated overnight at 4 °C with a 1:1000 dilution of anti- Histone H3 (Cell Signaling Technology, 4499s, US) and anti-VEZF1 (Proteintech, 19003-1-AP, US). After additional incubation with a 1:5000 dilution of anti-rabbit IgG (heavy and light chain) antibody (CST, 7074S) for 2 h, the immune complexes were detected by Immobilon Western HRP Substrate Peroxide Solution (Millipore Corporation, Billerica, MA 01821, USA). And images were acquired using ChemiDocTM XRS+ Imaging System (Bio-rad, US). The intensity of immunoreactivity was assessed using Image Lab 6.0 software.
Intracerebroventricular injection of the miR-191 antagomir
The miR-191 antagomir and NC antagomir were purchased from RiboBio(Guangzhou, China). The NC and miR-191 antagomir (2.5 μg/2.5 μl) were diluted with 1.25 μl of EntransterTM in vivo transfection reagent (Engreen,18668-11-1, Beijing, China). The solution was mixed with 1.25μl PBS gently, kept at room temperature for 5 min and then injected intracerebroventricularly (i.c.v.) using a microsyringe (KD Scientific Inc., USA) under the guidance of a stereotaxic instrument (RWD Life Science). A solution of 3.75μl PBS added with 1.25 μl of EntransterTM in vivo transfection reagent was acted as blank control. Intracerebroventricular injection was performed according to a previously described method [51]. The stereotaxic coordinates the right lateral ventricle: ML: -1.40mm, AP: -0.36mm, DV: -3.90mm.
Statistics
Differences between the two groups were analyzed by the unpaired Student’s t test or Mann-Whitney test after testing the distribution of the data. Differences between multiple groups were analyzed by one-ANOVA followed by the Student-Newman-Keuls post hoc test (Graphpad Prism 7.0, USA) after testing the data for equal variance. All values were expressed as mean ± SEM. Statistical significance was accepted at P<0.05.
Supplementary Materials
Author Contributions
Wei Gao and Xiang Lu designed the research, interpreted the data, and contributed to revising the manuscript. Kang Du and Can Zhao performed the research, analyzed the data, and wrote the manuscript. Li Wang and Yue Wang contributed to data collection and performance of rats MCAO model. Kangzhen Zhang, Xiyu Shen and Huixian Sun contributed to recruitment of patients and clinical diagnosis of disease.
Acknowledgments
The authors thank Ms. Weiwei Wu for her great advice and Ms. Ping Zhou for her excellent technical assistance.
Conflicts of Interest
The authors have announced no conflicts of interest.
Funding
This work was supported by the Natural Science Foundation of China (81770440, 81700331); Postgraduate Research & Practice Innovation Program of Jiangsu Province (SJCX18_0426); Natural Science Foundation of the Higher Education Institutions of Jiangsu Province (17KJB320003); Jiangsu Province Health Development Project with Science and Education (QNRC2016857); Natural Science Foundation of Jiangsu Province (BK20171051);The Six One Project of Jiangsu Province (LGY2018100).
References
- 1. Wei JW, Heeley EL, Wang JG, Huang Y, Wong LK, Li Z, Heritier S, Arima H, Anderson CS, and ChinaQUEST Investigators. Comparison of recovery patterns and prognostic indicators for ischemic and hemorrhagic stroke in China: the ChinaQUEST (QUality Evaluation of Stroke Care and Treatment) Registry study. Stroke. 2010; 41:1877–83. https://doi.org/10.1161/STROKEAHA.110.586909 [PubMed]
- 2. Powers WJ, Rabinstein AA, Ackerson T, Adeoye OM, Bambakidis NC, Becker K, Biller J, Brown M, Demaerschalk BM, Hoh B, Jauch EC, Kidwell CS, Leslie-Mazwi TM, et al, and American Heart Association Stroke Council. 2018 Guidelines for the Early Management of Patients With Acute Ischemic Stroke: A Guideline for Healthcare Professionals From the American Heart Association/American Stroke Association. Stroke. 2018; 49:e46–110. https://doi.org/10.1161/STR.0000000000000158 [PubMed]
- 3. Licata G, Tuttolomondo A, Corrao S, Di Raimondo D, Fernandez P, Caruso C, Avellone G, Pinto A. Immunoinflammatory activation during the acute phase of lacunar and non-lacunar ischemic stroke: association with time of onset and diabetic state. Int J Immunopathol Pharmacol. 2006; 19:639–46. https://doi.org/10.1177/039463200601900320 [PubMed]
- 4. Tuttolomondo A, Di Sciacca R, Di Raimondo D, Pedone C, La Placa S, Pinto A, Licata G. Effects of clinical and laboratory variables and of pretreatment with cardiovascular drugs in acute ischaemic stroke: a retrospective chart review from the GIFA study. Int J Cardiol. 2011; 151:318–22. https://doi.org/10.1016/j.ijcard.2010.06.005 [PubMed]
- 5. Di Raimondo D, Tuttolomondo A, Buttà C, Miceli S, Licata G, Pinto A. Effects of ACE-inhibitors and angiotensin receptor blockers on inflammation. Curr Pharm Des. 2012; 18:4385–413. https://doi.org/10.2174/138161212802481282 [PubMed]
- 6. Ruan L, Wang B, ZhuGe Q, Jin K. Coupling of neurogenesis and angiogenesis after ischemic stroke. Brain Res. 2015; 1623:166–73. https://doi.org/10.1016/j.brainres.2015.02.042 [PubMed]
- 7. Fraisl P, Mazzone M, Schmidt T, Carmeliet P. Regulation of angiogenesis by oxygen and metabolism. Dev Cell. 2009; 16:167–79. https://doi.org/10.1016/j.devcel.2009.01.003 [PubMed]
- 8. Phng LK, Gerhardt H. Angiogenesis: a team effort coordinated by notch. Dev Cell. 2009; 16:196–208. https://doi.org/10.1016/j.devcel.2009.01.015 [PubMed]
- 9. Suárez Y, Fernández-Hernando C, Yu J, Gerber SA, Harrison KD, Pober JS, Iruela-Arispe ML, Merkenschlager M, Sessa WC. Dicer-dependent endothelial microRNAs are necessary for postnatal angiogenesis. Proc Natl Acad Sci USA. 2008; 105:14082–87. https://doi.org/10.1073/pnas.0804597105 [PubMed]
- 10. Poliseno L, Tuccoli A, Mariani L, Evangelista M, Citti L, Woods K, Mercatanti A, Hammond S, Rainaldi G. MicroRNAs modulate the angiogenic properties of HUVECs. Blood. 2006; 108:3068–71. https://doi.org/10.1182/blood-2006-01-012369 [PubMed]
- 11. Suárez Y, Fernández-Hernando C, Pober JS, Sessa WC. Dicer dependent microRNAs regulate gene expression and functions in human endothelial cells. Circ Res. 2007; 100:1164–73. https://doi.org/10.1161/01.RES.0000265065.26744.17 [PubMed]
- 12. Bartel DP. Metazoan MicroRNAs. Cell. 2018; 173:20–51. https://doi.org/10.1016/j.cell.2018.03.006 [PubMed]
- 13. Tiedt S, Prestel M, Malik R, Schieferdecker N, Duering M, Kautzky V, Stoycheva I, Böck J, Northoff BH, Klein M, Dorn F, Krohn K, Teupser D, et al. RNA-Seq Identifies Circulating miR-125a-5p, miR-125b-5p, and miR-143-3p as Potential Biomarkers for Acute Ischemic Stroke. Circ Res. 2017; 121:970–80. https://doi.org/10.1161/CIRCRESAHA.117.311572 [PubMed]
- 14. Jeyaseelan K, Lim KY, Armugam A. MicroRNA expression in the blood and brain of rats subjected to transient focal ischemia by middle cerebral artery occlusion. Stroke. 2008; 39:959–66. https://doi.org/10.1161/STROKEAHA.107.500736 [PubMed]
- 15. Kulshreshtha R, Ferracin M, Wojcik SE, Garzon R, Alder H, Agosto-Perez FJ, Davuluri R, Liu CG, Croce CM, Negrini M, Calin GA, Ivan M. A microRNA signature of hypoxia. Mol Cell Biol. 2007; 27:1859–67. https://doi.org/10.1128/MCB.01395-06 [PubMed]
- 16. Lee HT, Chang YC, Tu YF, Huang CC. VEGF-A/VEGFR-2 signaling leading to cAMP response element-binding protein phosphorylation is a shared pathway underlying the protective effect of preconditioning on neurons and endothelial cells. J Neurosci. 2009; 29:4356–68. https://doi.org/10.1523/JNEUROSCI.5497-08.2009 [PubMed]
- 17. Wu CC, Chen YC, Chang YC, Wang LW, Lin YC, Chiang YL, Ho CJ, Huang CC. Human umbilical vein endothelial cells protect against hypoxic-ischemic damage in neonatal brain via stromal cell-derived factor 1/C-X-C chemokine receptor type 4. Stroke. 2013; 44:1402–09. https://doi.org/10.1161/STROKEAHA.111.000719 [PubMed]
- 18. Fasanaro P, D’Alessandra Y, Di Stefano V, Melchionna R, Romani S, Pompilio G, Capogrossi MC, Martelli F. MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3. J Biol Chem. 2008; 283:15878–83. https://doi.org/10.1074/jbc.M800731200 [PubMed]
- 19. Lou YL, Guo F, Liu F, Gao FL, Zhang PQ, Niu X, Guo SC, Yin JH, Wang Y, Deng ZF. miR-210 activates notch signaling pathway in angiogenesis induced by cerebral ischemia. Mol Cell Biochem. 2012; 370:45–51. https://doi.org/10.1007/s11010-012-1396-6 [PubMed]
- 20. Parikh VN, Jin RC, Rabello S, Gulbahce N, White K, Hale A, Cottrill KA, Shaik RS, Waxman AB, Zhang YY, Maron BA, Hartner JC, Fujiwara Y, et al. MicroRNA-21 integrates pathogenic signaling to control pulmonary hypertension: results of a network bioinformatics approach. Circulation. 2012; 125:1520–32. https://doi.org/10.1161/CIRCULATIONAHA.111.060269 [PubMed]
- 21. Buller B, Liu X, Wang X, Zhang RL, Zhang L, Hozeska-Solgot A, Chopp M, Zhang ZG. MicroRNA-21 protects neurons from ischemic death. FEBS J. 2010; 277:4299–307. https://doi.org/10.1111/j.1742-4658.2010.07818.x [PubMed]
- 22. Chistiakov DA, Orekhov AN, Bobryshev YV. The role of miR-126 in embryonic angiogenesis, adult vascular homeostasis, and vascular repair and its alterations in atherosclerotic disease. J Mol Cell Cardiol. 2016; 97:47–55. https://doi.org/10.1016/j.yjmcc.2016.05.007 [PubMed]
- 23. Long G, Wang F, Li H, Yin Z, Sandip C, Lou Y, Wang Y, Chen C, Wang DW. Circulating miR-30a, miR-126 and let-7b as biomarker for ischemic stroke in humans. BMC Neurol. 2013; 13:178. https://doi.org/10.1186/1471-2377-13-178 [PubMed]
- 24. Yang D, Wang J, Xiao M, Zhou T, Shi X. Role of Mir-155 in Controlling HIF-1α Level and Promoting Endothelial Cell Maturation. Sci Rep. 2016; 6:35316. https://doi.org/10.1038/srep35316 [PubMed]
- 25. Liu DZ, Tian Y, Ander BP, Xu H, Stamova BS, Zhan X, Turner RJ, Jickling G, Sharp FR. Brain and blood microRNA expression profiling of ischemic stroke, intracerebral hemorrhage, and kainate seizures. J Cereb Blood Flow Metab. 2010; 30:92–101. https://doi.org/10.1038/jcbfm.2009.186 [PubMed]
- 26. Wang HW, Huang TS, Lo HH, Huang PH, Lin CC, Chang SJ, Liao KH, Tsai CH, Chan CH, Tsai CF, Cheng YC, Chiu YL, Tsai TN, et al. Deficiency of the microRNA-31-microRNA-720 pathway in the plasma and endothelial progenitor cells from patients with coronary artery disease. Arterioscler Thromb Vasc Biol. 2014; 34:857–69. https://doi.org/10.1161/ATVBAHA.113.303001 [PubMed]
- 27. Dai GH, Ma PZ, Song XB, Liu N, Zhang T, Wu B. MicroRNA-223-3p inhibits the angiogenesis of ischemic cardiac microvascular endothelial cells via affecting RPS6KB1/hif-1a signal pathway. PLoS One. 2014; 9:e108468. https://doi.org/10.1371/journal.pone.0108468 [PubMed]
- 28. Xu W, Luo F, Sun B, Ye H, Li J, Shi L, Liu Y, Lu X, Wang B, Wang Q, Liu Q, Zhang A. HIF-2α, acting via miR-191, is involved in angiogenesis and metastasis of arsenite-transformed HBE cells. Toxicol Res (Camb). 2015; 5:66–78. https://doi.org/10.1039/C5TX00225G [PubMed]
- 29. Dal Monte M, Landi D, Martini D, Bagnoli P. Antiangiogenic role of miR-361 in human umbilical vein endothelial cells: functional interaction with the peptide somatostatin. Naunyn Schmiedebergs Arch Pharmacol. 2013; 386:15–27. https://doi.org/10.1007/s00210-012-0808-1 [PubMed]
- 30. Yang Z, Wu L, Zhu X, Xu J, Jin R, Li G, Wu F. MiR-29a modulates the angiogenic properties of human endothelial cells. Biochem Biophys Res Commun. 2013; 434:143–49. https://doi.org/10.1016/j.bbrc.2013.03.054 [PubMed]
- 31. Zhou Y, Li XH, Zhang CC, Wang MJ, Xue WL, Wu DD, Ma FF, Li WW, Tao BB, Zhu YC. Hydrogen sulfide promotes angiogenesis by downregulating miR-640 via the VEGFR2/mTOR pathway. Am J Physiol Cell Physiol. 2016; 310:C305–17. https://doi.org/10.1152/ajpcell.00230.2015 [PubMed]
- 32. Khoo CP, Roubelakis MG, Schrader JB, Tsaknakis G, Konietzny R, Kessler B, Harris AL, Watt SM. miR-193a-3p interaction with HMGB1 downregulates human endothelial cell proliferation and migration. Sci Rep. 2017; 7:44137. https://doi.org/10.1038/srep44137 [PubMed]
- 33. Wen Y, Chen R, Zhu C, Qiao H, Liu Y, Ji H, Miao J, Chen L, Liu X, Yang Y. MiR-503 suppresses hypoxia-induced proliferation, migration and angiogenesis of endothelial progenitor cells by targeting Apelin. Peptides. 2018; 105:58–65. https://doi.org/10.1016/j.peptides.2018.05.008 [PubMed]
- 34. Zhang ZG, Zhang L, Tsang W, Soltanian-Zadeh H, Morris D, Zhang R, Goussev A, Powers C, Yeich T, Chopp M. Correlation of VEGF and angiopoietin expression with disruption of blood-brain barrier and angiogenesis after focal cerebral ischemia. J Cereb Blood Flow Metab. 2002; 22:379–92. https://doi.org/10.1097/00004647-200204000-00002 [PubMed]
- 35. Gu Y, Ampofo E, Menger MD, Laschke MW. miR-191 suppresses angiogenesis by activation of NF-κB signaling. FASEB J. 2017; 31:3321–33. https://doi.org/10.1096/fj.201601263R [PubMed]
- 36. Aitsebaomo J, Kingsley-Kallesen ML, Wu Y, Quertermous T, Patterson C. Vezf1/DB1 is an endothelial cell-specific transcription factor that regulates expression of the endothelin-1 promoter. J Biol Chem. 2001; 276:39197–205. https://doi.org/10.1074/jbc.M105166200 [PubMed]
- 37. Miyashita H, Kanemura M, Yamazaki T, Abe M, Sato Y. Vascular endothelial zinc finger 1 is involved in the regulation of angiogenesis: possible contribution of stathmin/OP18 as a downstream target gene. Arterioscler Thromb Vasc Biol. 2004; 24:878–84. https://doi.org/10.1161/01.ATV.0000126373.52450.32 [PubMed]
- 38. Miyashita H, Sato Y. Metallothionein 1 is a downstream target of vascular endothelial zinc finger 1 (VEZF1) in endothelial cells and participates in the regulation of angiogenesis. Endothelium. 2005; 12:163–70. https://doi.org/10.1080/10623320500227101 [PubMed]
- 39. AlAbdi L, He M, Yang Q, Norvil AB, Gowher H. The transcription factor Vezf1 represses the expression of the antiangiogenic factor Cited2 in endothelial cells. J Biol Chem. 2018; 293:11109–18. https://doi.org/10.1074/jbc.RA118.002911 [PubMed]
- 40. Suárez Y, Sessa WC. MicroRNAs as novel regulators of angiogenesis. Circ Res. 2009; 104:442–54. https://doi.org/10.1161/CIRCRESAHA.108.191270 [PubMed]
- 41. Pan W, Wang L, Zhang XF, Zhang H, Zhang J, Wang G, Xu P, Zhang Y, Hu P, Zhang XD, Du RL, Wang H. Hypoxia-induced microRNA-191 contributes to hepatic ischemia/reperfusion injury through the ZONAB/Cyclin D1 axis. Cell Death Differ. 2019; 26:291–305 https://doi.org/10.1038/s41418-018-0120-9 [PubMed]
- 42. Wu XQ, Tian XY, Wang ZW, Wu X, Wang JP, Yan TZ. miR-191 secreted by platelet-derived microvesicles induced apoptosis of renal tubular epithelial cells and participated in renal ischemia-reperfusion injury via inhibiting CBS. Cell Cycle. 2019; 18:119–29. https://doi.org/10.1080/15384101.2018.1542900 [PubMed]
- 43. Bruderer M, Alini M, Stoddart MJ. Role of HOXA9 and VEZF1 in endothelial biology. J Vasc Res. 2013; 50:265–78. https://doi.org/10.1159/000353287 [PubMed]
- 44. Kuhnert F, Campagnolo L, Xiong JW, Lemons D, Fitch MJ, Zou Z, Kiosses WB, Gardner H, Stuhlmann H. Dosage-dependent requirement for mouse Vezf1 in vascular system development. Dev Biol. 2005; 283:140–56. https://doi.org/10.1016/j.ydbio.2005.04.003 [PubMed]
- 45. Xiong JW, Leahy A, Lee HH, Stuhlmann H. Vezf1: A Zn finger transcription factor restricted to endothelial cells and their precursors. Dev Biol. 1999; 206:123–41. https://doi.org/10.1006/dbio.1998.9144 [PubMed]
- 46. Zou Z, Ocaya PA, Sun H, Kuhnert F, Stuhlmann H. Targeted Vezf1-null mutation impairs vascular structure formation during embryonic stem cell differentiation. Arterioscler Thromb Vasc Biol. 2010; 30:1378–88. https://doi.org/10.1161/ATVBAHA.109.200428 [PubMed]
- 47. Gerald D, Adini I, Shechter S, Perruzzi C, Varnau J, Hopkins B, Kazerounian S, Kurschat P, Blachon S, Khedkar S, Bagchi M, Sherris D, Prendergast GC, et al. RhoB controls coordination of adult angiogenesis and lymphangiogenesis following injury by regulating VEZF1-mediated transcription. Nat Commun. 2013; 4:2824. https://doi.org/10.1038/ncomms3824 [PubMed]
- 48. Longa EZ, Weinstein PR, Carlson S, Cummins R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke. 1989; 20:84–91. https://doi.org/10.1161/01.STR.20.1.84 [PubMed]
- 49. Schäbitz WR, Weber J, Takano K, Sandage BW, Locke KW, Fisher M. The effects of prolonged treatment with citicoline in temporary experimental focal ischemia. J Neurol Sci. 1996; 138:21–25. https://doi.org/10.1016/0022-510X(95)00341-X [PubMed]
- 50. Ferro A, Queen LR, Priest RM, Xu B, Ritter JM, Poston L, Ward JP. Activation of nitric oxide synthase by beta 2-adrenoceptors in human umbilical vein endothelium in vitro. Br J Pharmacol. 1999; 126:1872–80. https://doi.org/10.1038/sj.bjp.0702512 [PubMed]
- 51. Zhao C, Li H, Zhao XJ, Liu ZX, Zhou P, Liu Y, Feng MJ. Heat shock protein 60 affects behavioral improvement in a rat model of Parkinson’s disease grafted with human umbilical cord mesenchymal stem cell-derived dopaminergic-like neurons. Neurochem Res. 2016; 41:1238–49. https://doi.org/10.1007/s11064-015-1816-6 [PubMed]