



Introduction
Hepatocellular carcinoma (HCC) is one of the leading causes of cancer death worldwide, especially in less-developed countries [1]. It is well known that the main environmental risk factor of HCC is hepatitis virus infection. Approximately 80% cases of HCC are attributed to chronic infection by hepatitis B (HBV) or C virus (HCV). However, only a small number of infected people develop HCC in their lifetime, indicating the pivotal role of genetic factors in tumorigenesis [2,3].
MicroRNAs (miRNA) are a class of small non-coding RNAs containing approximately 22 nucleotides. These endogenous molecules regulate gene expression via post-transcriptional repression. This regulation modulates diverse biological processes including proliferation, differentiation, and apoptosis, which are key steps in tumorigenesis [4,5]. MiRNAs have been implicated in the pathogenesis of various forms of liver disease such as viral hepatitis, liver fibrosis, and cancer [6]. They can also serve as repressors of viral infection pathways, thus regulating the host-virus interaction [7]. Moreover, studies have revealed that miRNAs can act as tumor suppressors or oncogenes and participate in the HBV-related HCC development [8,9]. Of all the miRNAs, miR-146a and miR-196a-2 are the most commonly studied in HCC. The miR-146a gene resides on chromosome 5q33.3 and the miR-196a-2 gene is located in a region between homeobox (HOX) clusters HOXC10 and HOXC9 on chromosome 12. The expression of mature miR-146a is decreased while miR-196a-2 are over-expressed in hepatocellular carcinoma tissues, suggesting both of them play roles in the development of HCC [10,11]. Single nucleotide polymorphism (SNP) is the most common form of mutation in the genome. Any minute alteration in miRNAs can be amplified by the numerous target genes, thus considerably increasing the possibility of carcinogenesis. Hence, genetic variation of miRNAs may alter the maturation, expression or lead to dysfunction of these molecules and potentially contribute to hepatitis virus-related HCC risk [4,12,13]. A number of studies have shown that several SNPs in miRNAs including miR-146a C>G (rs2910164) and miR-196a-2 C>T (rs2910164) are linked to the predisposition for HCC [10,11,14-29].However, the results of these studies are inconsistent. In this study, we systematically analyzed all relevant studies to assess the association of two common polymorphisms (rs2910164 and rs11614913) with hepatitis virus-related HCC risk.
Results
Characteristics of included studies
A total of 392 potentially relevant studies were identified via a database search. Finally, 14 studies comprising 3852 hepatitis virus-related HCC patients and 5275 cancer-free controls were included. And Table 1 lists the characteristics of these studies. Of the 14 studies, 8 studies including 1717 cases and 2589 controls evaluated the relationship between rs2910164 and hepatitis virus-related HCC risk [10,15,16,19,22-25], while 3 included both HBV-related and HCV-related HCC. Nine studies including 3217 cases and 4163 controls evaluated the association of rs11614913 with hepatitis virus-related HCC risk [10,11,14,17-21,24], and 5 included both HBV and HCV-related HCC. All studies were conducted in China except 2 Turkish studies and 1 Korean study. Additionally, 12 studies were based on hospital data and 3 included the general population. Distributions of genotype in controls conformed to HWE in all the studies except for 2 [10,20]. The detailed data of the allele and genotype distributions as well as HWE from each study are shown in Table S1.
Table 1. Characteristics of the studies included in the meta-analysis.
| First author | Year | Country | Ethnicity | Genotyping method | Source of control | Number of case/control | NOS score | SNP | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Yan | 2015 | China | Asian | PCR-RFLP | HB | 227/287 | 6 | 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Zhou | 2014 | China | Asian | PCR-RFLP | HB | 184/281 | 7 | 1,2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Kou | 2014 | China | Asian | PCR-RFLP | HB | 208/532 | 6 | 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cong | 2014 | China | Asian | PCR-RFLP | HB | 104/218 | 6 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hao | 2013 | China | Asian | PCR-RFLP | HB | 169/282 | 7 | 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Zhang | 2013 | China | Asian | MassARRAY | PB | 771/998 | 7 | 1,2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Shan | 2013 | China | Asian | PCR-RFLP | HB | 71/185 | 8 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Han | 2013 | China | Asian | FQ-PCR | HB | 1017/1009 | 7 | 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Xiang | 2012 | China | Asian | PCR-RFLP | HB | 73/100 | 6 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Kim | 2012 | Korea | Asian | PCR-RFLP | PB | 127/201 | 6 | 1,2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Wang | 2011 | China | Asian | MassARRAY | HB | 199/384 | 8 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Akkız | 2011 | Turkey | Caucasian | PCR-RFLP | HB | 188/222 | 9 | 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Akkız | 2011 | Turkey | Caucasian | PCR-RFLP | HB | 153/185 | 9 | 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Qi | 2010 | China | Asian | PCR-LDR | HB | 361/391 | 8 | 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PCR: polymerase chain reaction; RFLP: restriction fragment length polymorphism; FQ: fluorescent-probe real-time quantitative; LDR: ligation detection reaction; PB: population based; HB: hospital based; NOS: Newcastle-Ottawa Quality Assessment Scale; SNP: single-nucleotide polymorphism; SNP No.1: rs2910164, No 2: rs1161491. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Meta-analysis results
As presented in Table 2, miR-146a rs291016 was found to increase hepatitis virus-related HCC risk in overall analysis under G versus (vs.) C (OR=1.13, 95% CI =1.04-1.24, P=0.006,), GG vs. CC (OR=1.28, 95% CI=1.06-1.55, P=0.01,Fig.1), CG vs. GG (OR=1.20, 95% CI=1.04-1.38, P=0.01) and CG+GG vs. CC (OR=1.22, 95% CI=1.06-1.39, P=0.004). Additionally, this risk is more significant in the Chinese population. After omitting the study which did not conform to HWE, the results were found to be consistent with the overall analysis. Additionally, subgroup analyses showed a higher HBV-related HCC risk in the same 4 genetic models, whereas no significance was found in the HCV-related HCC subgroup. When stratifying data by source of control, rs2910164 was also observed to significantly increase hepatitis virus-related HCC risk based on hospital data, but there was no statistical significance based on population data.
Table 2. Meta-analysis results of miR-146a and miR-196a-2 polymorphisms with hepatitis virus-related HCC risk.
| SNP | B vs. A | BB vs. AA | AB vs. AA | BB vs. AA+AB | AB+BB vs. AA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OR (95%CI) | P | I2 | Ph | OR (95%CI) | P | I2 | Ph | OR (95%CI) | P | I2 | Ph | OR (95%CI) | P | I2 | Ph | OR (95%CI) | P | I2 | Ph | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-146a C>G (rs2910164) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Overall | 1.13 (1.04-1.24) | 0.006 | 36% | 0.14 | 1.28 (1.06-1.55) | 0.01 | 20% | 0.27 | 1.20 (1.04-1.38) | 0.01 | 0% | 0.95 | 1.10 (0.93-1.29) | 0.26 | 35% | 0.15 | 1.22 (1.06-1.39) | 0.004 | 0% | 0.81 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HWE | 1.12 (1.02-1.24) | 0.02 | 44% | 0.10 | 1.23(1.01-1.51) | 0.04 | 18% | 0.29 | 1.18 (1.01-1.37) | 0.04 | 0% | 0.95 | 1.07(0.90-1.26) | 0.47 | 36% | 0.15 | 1.19 (1.03-1.37) | 0.02 | 0% | 0.82 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Chinese | 1.17 (1.06-1.29) | 0.002 | 39% | 0.15 | 1.32 (1.08-1.62) | 0.007 | 36% | 0.16 | 1.19 (1.03-1.39) | 0.02 | 0% | 0.84 | 1.19 (0.99-1.43) | 0.06 | 32% | 0.20 | 1.23 (1.06-1.41) | 0.005 | 0% | 0.61 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HB | 1.20 (1.05-1.36) | 0.006 | 47% | 0.09 | 1.51 (1.14-1.99) | 0.004 | 17% | 0.31 | 1.23 (0.99-1.52) | 0.06 | 0% | 0.84 | 1.18 (0.94-1.46) | 0.15 | 48% | 0.09 | 1.29 (1.06-1.58) | 0.01 | 0% | 0.69 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PB | 1.08 (0.95-1.22) | 0.25 | 0% | 0.76 | 1.11 (0.85-1.45) | 0.45 | 0% | 0.64 | 1.17 (0.97-1.42) | 0.10 | 0% | 0.97 | 1.01 (0.79-1.29) | 0.94 | 0% | 0.59 | 1.16 (0.97-1.39) | 0.11 | 0% | 0.93 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HBV- | 1.13 (1.03-1.23) | 0.01 | 39% | 0.12 | 1.26 (1.04-1.54) | 0.02 | 10% | 0.36 | 1.20 (1.04-1.39) | 0.01 | 0% | 0.95 | 1.07 (0.91-1.27) | 0.41 | 33% | 0.16 | 1.21 (1.06-1.39) | 0.005 | 0% | 0.81 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HCV- | 1.25 (0.89-1.76) | 0.20 | 0% | 0.42 | 1.98 (0.89-4.45) | 0.10 | 0% | 0.39 | 1.18 (0.62-2.23) | 0.62 | 0% | 0.92 | 1.67 (0.75-3.71) | 0.21 | 51% | 0.13 | 1.38 (0.76-2.52) | 0.30 | 0% | 0.97 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| miR-196a2 C>T (rs11614913) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Overall | 0.85 (0.74-0.98) | 0.02 | 73% | 0.00 | 0.71 (0.52-0.95) | 0.02 | 75% | 0.00 | 0.84 (0.70-1.01) | 0.07 | 57% | 0.02 | 0.82 (0.68-0.98) | 0.03 | 54% | 0.03 | 0.80 (0.65-0.99) | 0.04 | 71% | 0.00 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HWE | 0.88 (0.76-1.01) | 0.07 | 72% | 0.00 | 0.75 (0.55-1.02) | 0.07 | 74% | 0.00 | 0.87 (0.72-1.06) | 0.16 | 55% | 0.03 | 0.85 (0.71-1.02) | 0.08 | 50% | 0.05 | 0.83 (0.67-1.04) | 0.11 | 69% | 0.00 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Chinese | 0.85 (0.73-0.99) | 0.04 | 76% | 0.00 | 0.70 (0.50-0.97) | 0.03 | 78% | 0.00 | 0.82(0.67-1.02) | 0.07 | 64% | 0.01 | 0.83 (0.68-1.00) | 0.05 | 56% | 0.04 | 0.79 (0.62-1.00) | 0.05 | 75% | 0.00 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HB | 0.83 (0.69-0.99) | 0.04 | 78% | 0.00 | 0.66 (0.45-0.97) | 0.03 | 79% | 0.00 | 0.83 (0.67-1.04) | 0.11 | 60% | 0.02 | 0.77 (0.60-0.99) | 0.04 | 63% | 0.01 | 0.78 (0.60-1.01) | 0.06 | 74% | 0.00 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PB | 0.92 (0.70-1.22) | 0.57 | 65% | 0.09 | 0.85 (0.46-1.58) | 0.60 | 68% | 0.08 | 0.88 (0.53-1.45) | 0.62 | 63% | 0.10 | 0.86 (0.71-1.04) | 0.13 | 0% | 0.34 | 0.88 (0.51-1.52) | 0.64 | 71% | 0.06 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HBV- | 0.85 (0.73-0.98) | 0.03 | 75% | 0.00 | 0.70 (0.50-0.96) | 0.03 | 77% | 0.00 | 0.83 (0.69-1.01) | 0.06 | 56% | 0.02 | 0.82 (0.67-1.00) | 0.05 | 58% | 0.01 | 0.79 (0.63--0.99) | 0.04 | 71% | 0.00 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HCV- | 0.80 (0.65-0.98) | 0.03 | 0% | 0.92 | 0.61 (0.39-0.94) | 0.02 | 0% | 0.93 | 0.82 (0.59-1.12) | 0.21 | 0% | 0.69 | 0.69 (0.47-1.02) | 0.06 | 0% | 1.00 | 0.75 (0.55-1.02) | 0.07 | 0% | 0.72 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PB: population based; HB: hospital based; HWE: subgroup excluding the studies departing from HWE; HBV-: hepatitis B virus-related HCC; HCV-: hepatitis C virus-related HCC; N: number of samples; A: the major allele; B: the minor allele; OR: odds ratio; CI: confidence interval; Ph: P-value of Q-test for heterogeneity. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
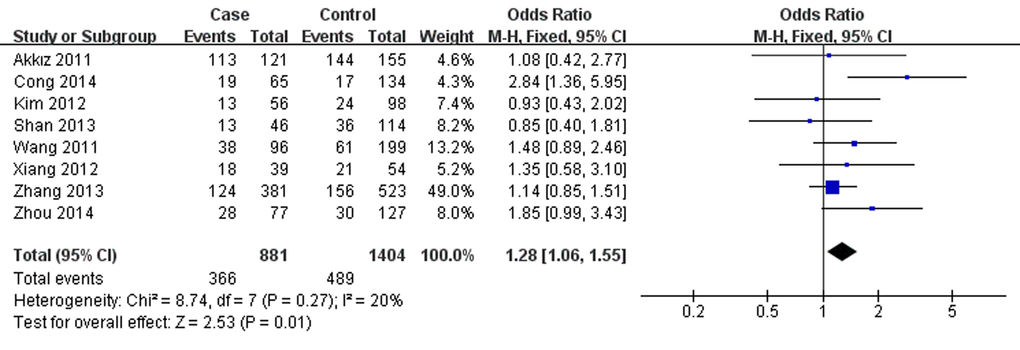
Figure 1. Forest plot describing the association between miR-146a rs2910164 and hepatitis virus-related HCC risk under homozygous model (GG vs. CC). The squares and horizontal lines correspond to the study-specific OR and 95% CI. The area of the squares reflects the weight (inverse of the variance). The diamond represents the summary OR and 95% CI. M–H: Mantel–Haenszel; df: degrees of freedom; Events: the number of GG genotypes.
Conversely, miR-196a2 rs11614913 was associated with lower hepatitis virus-related HCC risk in overall analysis under T vs. C (OR=0.85, 95% CI=0.74- 0.98, P=0.02), TT vs. CC (OR=0.71, 95% CI=0.52-0.95, P=0.02), TT vs. CT+CC (OR=0.82, 95% CI=0.68-0.98, P=0.03, Fig.2) and CT+TT vs. CC (OR=0.80, 95% CI =0.65-0.99, P=0.04). The results were similar in the overall population as well as particularly among the Chinese. However, further analysis of the studies which were in agreement with HWE showed no associations between rs11614913 and hepatitis virus-related HCC. Additionally, in subgroup analysis by source of control, decreased hepatitis virus-related HCC risk was observed based on hospital data, however, no significance was shown based on population data. Furthermore, in the stratified analysis according to etiology, rs11614913 was found to decrease both HBV- and HCV-related HCC risk in different genetic models.
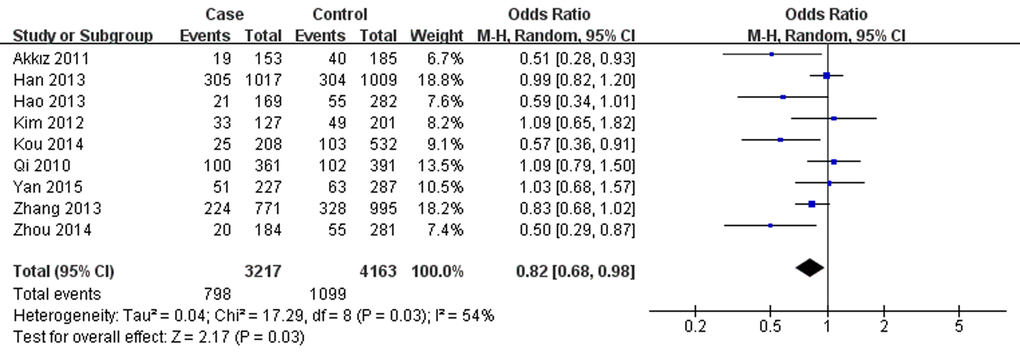
Figure 2. Forest plot describing the association between miR-196a-2 rs11614913 and hepatitis virus-related HCC risk under recessive model (TT vs. CC+CT). The squares and horizontal lines correspond to the study-specific OR and 95% CI. The area of the squares reflects the weight (inverse of the variance). The diamond represents the summary OR and 95% CI. M–H: Mantel–Haenszel; df: degrees of freedom; Events: the number of TT genotypes.
Heterogeneity analysis
No significant heterogeneity was observed for rs2910164 in all the genetic models analyzed. However, significant heterogeneity was detected for rs11614913 in each genetic model (Table 2). To explore the source of heterogeneity, we carried out subgroup analysis. After stratifying by etiology, the value of I2 in HCV-related HCC subgroup was reduced to zero. However, heterogeneity was still significant in other subgroups. We then used Galbraith plots to further investigate the heterogeneity and identified 3 three studies [10,17,20] as the outliers (Fig. S1). When these 3 studies were excluded, I2 values were <50% in heterozygous and recessive models in the overall populations and some subgroups (data not shown).
Sensitivity analysis and publication bias
The result of sensitivity analyses shows that the omission of any individual study did not substantially alter the pooled ORs, suggesting that the meta-analysis results are robust and statistically reliable (Fig. 3).
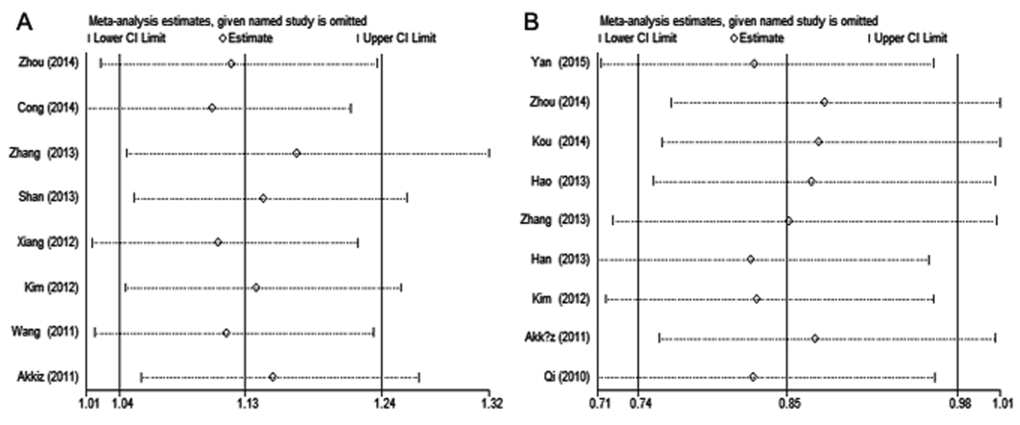
Figure 3. Sensitivity analysis of hepatitis virus-related HCC risk associated with (A) miR-146a rs2910164 and (B) miR-196a-2 rs11614913 under the allelic model. Pooled ORs were computed by omitting each study (left column) in turn. The two ends of the dotted lines represent the 95% CI.
Begg’s funnel plots did not show any obvious asymmetry in the overall analysis (Fig. 4). The Egger’s test results were not significant either (Table S1). Thus, no publication bias was revealed for the 2 polymorphisms and hepatitis virus-related HCC risk.
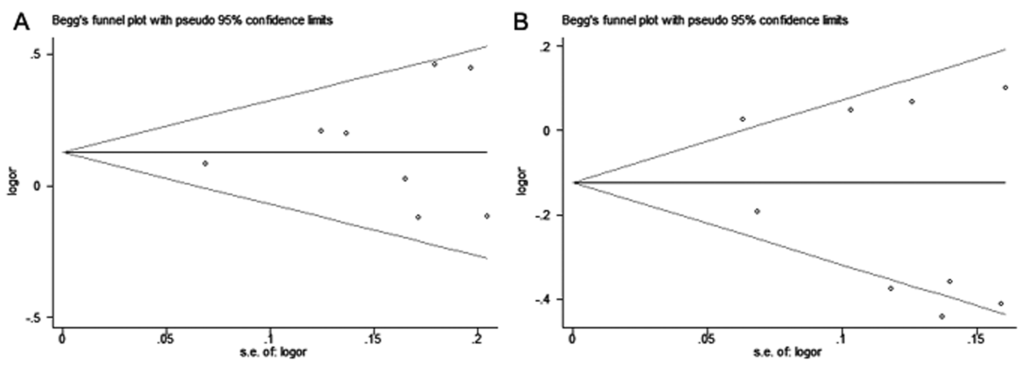
Figure 4. Begg’s funnel plots for publication bias of (A) miR-146a rs2910164 and (B) miR-196a-2 rs11614913 polymorphisms with hepatitis virus-related HCC risk under allelic model. Each point represents a single study for the indicated association.
Discussion
In recent years, several studies have focused on the SNPs in miRNAs and have investigated their potential role in HCC susceptibility [10,11,14-28]. A deeper understanding of the association between these polymorphisms and HCC would help early detection and prevention of this deadly disease in high-risk populations such as the Chinese. Amongst these SNPs, miR-146a C>G and miR-196a-2 C>T are the most commonly studied. Some case-control studies suggested these two SNPs contribute to HCC development [10,11,14,16,18,20,21,24,25], whereas other investigations failed to find any relationship between them [15,17,19,22,23]. The inconsistent results may originate from several factors including sample size, source of controls and genotype methods. Several meta-analyses have also explored the association between these 2 SNPs and HCC risk, but the results are inconsistent, while only few have focused specifically on hepatitis virus-related HCC [29-36]. Hence, we performed this updated meta-analysis to improve our understanding of the relationship between miR-146a C>G/miR-196a-2 C>T and hepatitis virus-related HCC risk, hoping to provide guidance for future studies.
Our results suggest that the G allele of rs2910164 increases the risk of hepatitis virus-related HCC, specifically for HBV-related HCC. This means that individuals carrying the G allele may be more susceptible to HBV-related HCC as compared to those carrying the C allele. Recent evidences indicated that miR-146a can promote apoptosis by inhibiting the NF-κB pathway and blocking its impact on cell proliferation, angiogenesis, metastasis and cancer cell survival. Loss of function of miR-146a could promote cancer cell migration and invasion [21]. And it has been reported that the rs2910164 polymorphism may reduce the production of mature miR-146a and lead to decreased suppression of its target genes including vascular endothelial growth factor, nuclear factor-κB, p65, and HAb18G, which are probably involved in hepatocarcinogenesis. Additionally, miR-146a expression in hepatoma cells and tissues is much lower than that in normal hepatic cells and tissues [37,38]. The results from the present meta-analysis confirmed the role of miR-146a rs2910164 in HCC development. Our results are consistent with that from previous meta-analyses [32-35], However, few differences were noted in our results compared with previous studies, since we specifically focused on hepatitis virus-related HCC. Although the P-value of HWE in the controls was <0.05 in one study [10], we included this study in the analysis since excluding it did not significantly influence the pooled OR. Additionally, neither significant heterogeneity nor publication bias was detected for this SNP, suggesting that our result regarding miR-146a C>G was statistically reliable.
Previous studies have revealed that miR-196a-2 deregulates numerous target genes such as HOX and ANXA1, which are considered to play crucial roles in the progression of HCC. Therefore, variations in miR-196a-2 may relate to HCC susceptibility [30,39]. Our data support this role of miR-196a-2 in HCC development. However, our results should be interpreted cautiously, due to obvious heterogeneity existed. Subgroup analysis suggested that the major source of heterogeneity may be the type of hepatitis virus, while Galbraith plots identified the studies conducted by Kou, Zhou and Han as the outliers. For this SNP, one study deviated from HWE [20]. After excluding this study, no significant association was identified in any model. This indicates that this study did not investigate a broad representative population, which has greatly impacted the pooled ORs. Therefore, epidemiological studies with broader population are needed to confirm our findings. The conclusions of previous meta-analyses for this SNP and HCC susceptibility were contradictory. Some studies reported no association between rs11614913 and HCC risk [29,30,33], whereas 2 recent meta-analyses published by Chen and Zhu showed that the C allele of rs11614913 may contribute to HCC susceptibility [35,36]. Our results are consistent with those reported by Chen and Zhu. Nevertheless, our meta-analysis differs from these studies. Chen et al. assessed HCC risk without stratifying by etiology while we focused on hepatitis virus-related HCC. Though Zhu et al. specifically analyzed HBV-related HCC, there were only 7 studies in their meta-analysis. On the other hand, our analysis included 2 additional studies and we estimated both HBV- and HCV-related HCC risk.
We attempted a comprehensive evaluation of the relationship between miR-146a/miR-196a-2 polymorphisms and hepatitis virus-related HCC risk. However, few limitations remain. First, the total sample size in this study was relatively small, especially for the HCV-related HCC subgroup. Additionally, we only examined the Chinese subpopulation as only one Korean and one Turkish study were included for both SNPs. Therefore, large-scale studies including multiple ethnicities are needed to provide sufficient statistical power. Secondly, heterogeneity still existed for rs11614913 in spite of our effort. And there may be a bias considering the representativeness of the samples. Therefore, it needs to be acknowledged that the result of this SNP, which may be affected, should be interpreted with caution. Thirdly, language bias may exist since only 2 languages were employed in the literature review. Lastly, lack of available original data prevented adjustment for other covariates such as age, gender, and lifestyle. As gene-environment interactions also affect cancer risk, these factors may potentially influence the results.
In summary, the results of our meta-analysis suggest that both miR-146a C>G and miR-196a-2 C>T polymorphisms associate with hepatitis virus-related HCC risk, especially in the Chinese population. MiR-146a C>G can increase HBV-related HCC risk while miR-196a-2 C>T may decrease the risk of both HBV-related and HCV-related HCC. In addition, large-scale studies with multi-ethnic groups are needed to confirm the findings. Detailed information about the effects of gene-environment interaction on HCC development is also required to further clarify these associations.
Materials and Methods
Search strategy
All relevant literature, in English or Chinese, was identified from databases including EMBASE, PubMed, Web of Science, Wanfang and Chinese National Knowledge Infrastructure (CNKI) as of 25th November, 2016. The search was performed using the following terms: “miR-146a OR microRNA-146a OR miR-196a-2 OR microRNA-196a-2” AND “polymorphisms OR variant OR SNP OR mutation” AND “hepatocellular carcinoma OR liver cell carcinoma OR liver cancer”. Additionally, references in retrieved articles were searched manually.
Criteria for selection
The studies that complied with the following criteria were included: 1) original studies investigating the association of rs2910164 or rs11614913 and hepatitis virus-related HCC risk; 2) case-control design studies conducted in humans; 3) patients were pathologically diagnosed, with cancer-free individuals used as controls; 4) availability of full text, with detailed data of the allele and genotype distributions. Studies without a control group, repeated publications, reviews, and conference abstracts were excluded. If two or more studies showed an overlapping study population, only the most comprehensive study was included. The details of included and excluded articles were shown in Fig.5.
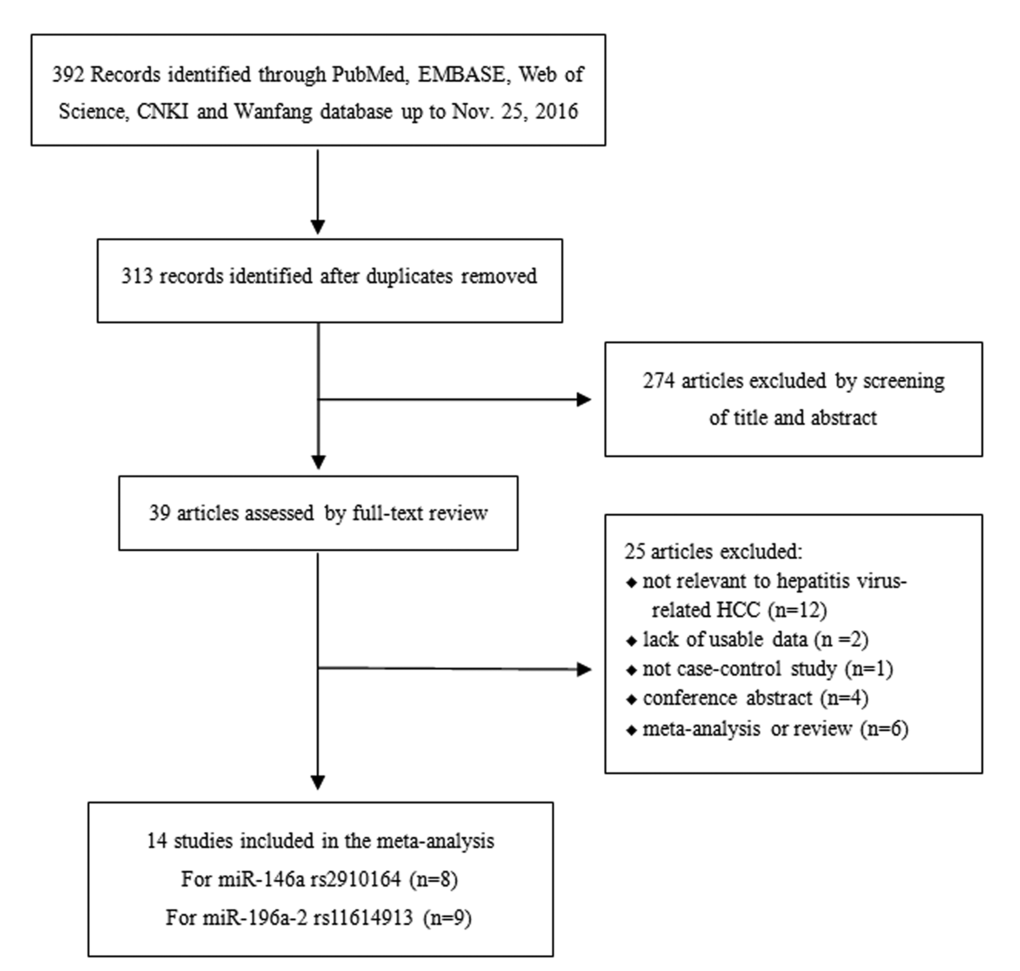
Figure 5. The flow chart illustrating the selection process of included studies.
Data extraction
Based on the criteria listed above, 2 authors independently reviewed the literature. The quality of each included article were assessed by Newcastle-Ottawa Quality Assessment Scale for case-control studies. For each eligible study, the raw data and information including: first author, publication year, country of origin, ethnicity, genotype methods, source of control, numbers of cases and controls, frequencies of allele and genotypes, and P value of Hardy-Weinberg equilibriums (HWE) in controls were collected. Any discrepancy was discussed between authors to reach a consensus.
Statistical analysis
Odds ratios (OR) with corresponding 95% confidence intervals (CI) for miR-146a and miR-196a-2 polymorphisms were calculated in each study under 5 different genetic models: allele comparison (B vs. A), homozygote (BB vs. AA), heterozygote (AB vs. AA), dominant model (BB+AB vs. AA), and recessive model (BB vs. AA+AB). The combined ORs were determined by Z test and P<0.05 was judged as statistically significant. Heterogeneity between studies was detected using I2 test and Q statistic, while significance was considered at I2>50%. A random-effects model was applied to analyze the pooled ORs if I2≥50%. Otherwise, a fixed-effects model was implemented. We carried out subgroup analyses to investigate the specific effects of ethnicity, source of control and etiology. Additionally, we assessed publication bias using Egger’s test and Begg’s funnel plot (significant bias was considered if P<0.05). We also performed a sensitivity analysis to assess the consistency and stability of our meta-analysis by removing each study in turn. All statistical analyses were accomplished with the software Review Manager 5.3 (Cochrane Collaboration, London, UK).and STATA (Version 12.0; Stata Corp, College Station, TX).
Supplementary Materials
Author Contributions
Zhi-Jun Dai and Tian Tian conceived and designed the study. Tian Tian, Meng Wang searched and selected literature, Tian Tian, Meng Wang, Zhi-Ming Dai, Shuai Lin, Peng-Tao Yang collected and assembled the data. Zhi-Jun Dai, Wenge Zhu, Tian Tian, Meng Wang analyzed and interpreted the data. Tian Tian wrote the manuscript. Xing-Han Liu, Kang Liu, Yu-Yao Zhu, Yi Zheng and Meng Liu contributed materials/analysis tools. Zhi-Jun Dai and Wenge Zhu reviewed drafts of the paper. Zhi-Jun Dai supervised the entire study.
Conflicts of Interest
The authors confirm no conflicts of interest.
Funding
This study was supported by National Natural Science Foundation, China (No. 81471670); China Postdoctoral Science Foundation (No. 2014M560791; 2015T81037); the Fundamental Research Funds for the Central Universities, China (No. 2014qngz-04) and Science and Technology Plan of Innovation Project, Shaanxi province, China (No. 2015KTCL03-06).
References
- 1. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65:87–108. https://doi.org/10.3322/caac.21262 [PubMed]
- 2. Yang JD, Roberts LR. Hepatocellular carcinoma: A global view. Nat Rev Gastroenterol Hepatol. 2010; 7:448–58. https://doi.org/10.1038/nrgastro.2010.100 [PubMed]
- 3. Bouchard MJ, Navas-Martin S. Hepatitis B and C virus hepatocarcinogenesis: lessons learned and future challenges. Cancer Lett. 2011; 305:123–43. https://doi.org/10.1016/j.canlet.2010.11.014 [PubMed]
- 4. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004; 116:281–97. https://doi.org/10.1016/S0092-8674(04)00045-5 [PubMed]
- 5. Fabian MR, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annu Rev Biochem. 2010; 79:351–79. https://doi.org/10.1146/annurev-biochem-060308-103103 [PubMed]
- 6. Wang XW, Heegaard NH, Orum H. MicroRNAs in liver disease. Gastroenterology. 2012; 142:1431–43. https://doi.org/10.1053/j.gastro.2012.04.007 [PubMed]
- 7. Chun HW, Hong R. Significance of the hedgehog pathway-associated proteins Gli-1 and Gli-2 and the epithelial-mesenchymal transition-associated proteins Twist and E-cadherin in hepatocellular carcinoma. Oncol Lett. 2016; 12:1753–62. [PubMed]
- 8. Esquela-Kerscher A, Slack FJ. Oncomirs - microRNAs with a role in cancer. Nat Rev Cancer. 2006; 6:259–69. https://doi.org/10.1038/nrc1840 [PubMed]
- 9. Yu K, Shi G, Li N. The function of MicroRNA in hepatitis B virus-related liver diseases: from Dim to Bright. Ann Hepatol. 2015; 14:450–56. [PubMed]
- 10. Zhou B, Dong LP, Jing XY, Li JS, Yang SJ, Wang JP, Zhao LF. Association between miR-146aG>C and miR-196a2C>T polymorphisms and the risk of hepatocellular carcinoma in a Chinese population. Tumour Biol. 2014; 35:7775–80. https://doi.org/10.1007/s13277-014-2020-z [PubMed]
- 11. Yan P, Xia M, Gao F, Tang G, Zeng H, Yang S, Zhou H, Ding D, Gong L. Predictive role of miR-146a rs2910164 (C>G), miR-149 rs2292832 (T>C), miR-196a2 rs11614913 (T>C) and miR-499 rs3746444 (T>C) in the development of hepatocellular carcinoma. Int J Clin Exp Pathol. 2015; 8:15177–83. [PubMed]
- 12. Negrini M, Nicoloso MS, Calin GA. MicroRNAs and cancer--new paradigms in molecular oncology. Curr Opin Cell Biol. 2009; 21:470–79. https://doi.org/10.1016/j.ceb.2009.03.002 [PubMed]
- 13. Wu M, Jolicoeur N, Li Z, Zhang L, Fortin Y, L’Abbe D, Yu Z, Shen SH. Genetic variations of microRNAs in human cancer and their effects on the expression of miRNAs. Carcinogenesis. 2008; 29:1710–16. https://doi.org/10.1093/carcin/bgn073 [PubMed]
- 14. Akkız H, Bayram S, Bekar A, Akgöllü E, Ulger Y. A functional polymorphism in pre-microRNA-196a-2 contributes to the susceptibility of hepatocellular carcinoma in a Turkish population: a case-control study. J Viral Hepat. 2011; 18:e399–407. https://doi.org/10.1111/j.1365-2893.2010.01414.x [PubMed]
- 15. Akkız H, Bayram S, Bekar A, Akgöllü E, Usküdar O, Sandıkçı M. No association of pre-microRNA-146a rs2910164 polymorphism and risk of hepatocellular carcinoma development in Turkish population: a case-control study. Gene. 2011; 486:104–09. https://doi.org/10.1016/j.gene.2011.07.006 [PubMed]
- 16. Cong N, Chen H, Bu WZ, Li JP, Liu N, Song JL. miR-146a G>C polymorphisms and risk of hepatocellular carcinoma in a Chinese population. Tumour Biol. 2014; 35:5669–73. https://doi.org/10.1007/s13277-014-1750-2 [PubMed]
- 17. Han Y, Pu R, Han X, Zhao J, Zhang Y, Zhang Q, Yin J, Xie J, Shen Q, Deng Y, Ding Y, Li W, Li J, et al. Associations of pri-miR-34b/c and pre-miR-196a2 polymorphisms and their multiplicative interactions with hepatitis B virus mutations with hepatocellular carcinoma risk. PLoS One. 2013; 8:e58564. https://doi.org/10.1371/journal.pone.0058564 [PubMed]
- 18. Hao Y-X, Wang J-P, Zhao L-F. Associations between three common MicroRNA polymorphisms and hepatocellular carcinoma risk in Chinese. Asian Pac J Cancer Prev. 2013; 14:6601–04. https://doi.org/10.7314/APJCP.2013.14.11.6601 [PubMed]
- 19. Kim WH, Min KT, Jeon YJ, Kwon CI, Ko KH, Park PW, Hong SP, Rim KS, Kwon SW, Hwang SG, Kim NK. Association study of microRNA polymorphisms with hepatocellular carcinoma in Korean population. Gene. 2012; 504:92–97. https://doi.org/10.1016/j.gene.2012.05.014 [PubMed]
- 20. Kou JT, Fan H, Han D, Li L, Li P, Zhu J, Ma J, Zhang ZH, He Q. Association between four common microRNA polymorphisms and the risk of hepatocellular carcinoma and HBV infection. Oncol Lett. 2014; 8:1255–60. [PubMed]
- 21. Qi P, Dou TH, Geng L, Zhou FG, Gu X, Wang H, Gao CF. Association of a variant in MIR 196A2 with susceptibility to hepatocellular carcinoma in male Chinese patients with chronic hepatitis B virus infection. Hum Immunol. 2010; 71:621–26. https://doi.org/10.1016/j.humimm.2010.02.017 [PubMed]
- 22. Shan YF, Huang YH, Chen ZK, Huang KT, Zhou MT, Shi HQ, Song QT, Yu ZP, Deng AM, Zhang QY. miR-499A>G rs3746444 and miR-146aG>C expression and hepatocellular carcinoma risk in the Chinese population. Genet Mol Res. 2013; 12:5365–71. https://doi.org/10.4238/2013.November.7.11 [PubMed]
- 23. Xiang Y, Fan S, Cao J, Huang S, Zhang LP. Association of the microRNA-499 variants with susceptibility to hepatocellular carcinoma in a Chinese population. Mol Biol Rep. 2012; 39:7019–23. https://doi.org/10.1007/s11033-012-1532-0 [PubMed]
- 24. Zhang J, Wang R, Ma Y-Y, Chen L-Q, Jin B-H, Yu H, Wang J-C, Gao C-F, Liu J. Association between single nucleotide polymorphisms in miRNA196a-2 and miRNA146a and susceptibility to hepatocellular carcinoma in a Chinese population. Asian Pac J Cancer Prev. 2013; 14:6427–31. https://doi.org/10.7314/APJCP.2013.14.11.6427 [PubMed]
- 25. Wang W. Association study of miR-146a single nucleotide polymorphisms with susceptibility to hepatocellular carcinoma and microarray analysis of tumor associated microRNAs [D]. 2011.
- 26. Chu YH, Hsieh MJ, Chiou HL, Liou YS, Yang CC, Yang SF, Kuo WH. MicroRNA gene polymorphisms and environmental factors increase patient susceptibility to hepatocellular carcinoma. PLoS One. 2014; 9:e89930. https://doi.org/10.1371/journal.pone.0089930 [PubMed]
- 27. Ma Y, Wang R, Zhang J, Li W, Gao C, Liu J, Wang J. Identification of miR-423 and miR-499 polymorphisms on affecting the risk of hepatocellular carcinoma in a large-scale population. Genet Test Mol Biomarkers. 2014; 18:516–24. https://doi.org/10.1089/gtmb.2013.0510 [PubMed]
- 28. Wang XH, Wang FR, Tang YF, Zou HZ, Zhao YQ. Association of miR-149C>T and miR-499A>G polymorphisms with the risk of hepatocellular carcinoma in the Chinese population. Genet Mol Res. 2014; 13:5048–54. https://doi.org/10.4238/2014.July.4.20 [PubMed]
- 29. Wang BS, Liu Z, Xu WX, Sun SL. Functional polymorphisms in microRNAs and susceptibility to liver cancer: a meta-analysis and meta-regression. Genet Mol Res. 2014; 13:5426–40. https://doi.org/10.4238/2014.July.24.22 [PubMed]
- 30. Wang Z, Cao Y, Jiang C, Yang G, Wu J, Ding Y. Lack of association of two common polymorphisms rs2910164 and rs11614913 with susceptibility to hepatocellular carcinoma: a meta-analysis. PLoS One. 2012; 7:e40039. https://doi.org/10.1371/journal.pone.0040039 [PubMed]
- 31. Hu M, Zhao L, Hu S, Yang J. The association between two common polymorphisms in MicroRNAs and hepatocellular carcinoma risk in Asian population. PLoS One. 2013; 8:e57012. https://doi.org/10.1371/journal.pone.0057012 [PubMed]
- 32. Liu J, Xie B, Chen S, Jiang F, Meng W. Association study of two inflammation-related polymorphisms with susceptibility to hepatocellular carcinoma: a meta-analysis. BMC Med Genet. 2014; 15:92. https://doi.org/10.1186/s12881-014-0092-7 [PubMed]
- 33. Peng Q, Li S, Lao X, Chen Z, Li R, Deng Y, Qin X. The association of common functional polymorphisms in mir-146a and mir-196a2 and hepatocellular carcinoma risk: evidence from a meta-analysis. Medicine (Baltimore). 2014; 93:e252. https://doi.org/10.1097/MD.0000000000000252 [PubMed]
- 34. Chen J, Cao X, Zhang H. MiR-146a rs2910164 polymorphism is associated with hepatocellular carcinoma: a meta-analysis. Int J Clin Exp Med. 2015; 8:15852–56. [PubMed]
- 35. Chen M, Luo F, Yu J, Xiang G, Jiang D, Pu X. Common functional polymorphism within miR-146a and miR-196a-2 as susceptibility loci for hepatocellular carcinoma: an updated meta-analysis. Meta Gene. 2015; 7:40–47. https://doi.org/10.1016/j.mgene.2015.11.002 [PubMed]
- 36. Zhu SL, Zhong JH, Gong WF, Li H, Li LQ. Association of the miR-196a2 C>T and miR-499 A>G polymorphisms with hepatitis B virus-related hepatocellular carcinoma risk: an updated meta-analysis. Onco Targets Ther. 2016; 9:2111–19. https://doi.org/10.2147/OTT.S96738 [PubMed]
- 37. Jazdzewski K, Murray EL, Franssila K, Jarzab B, Schoenberg DR, de la Chapelle A. Common SNP in pre-miR-146a decreases mature miR expression and predisposes to papillary thyroid carcinoma. Proc Natl Acad Sci USA. 2008; 105:7269–74. https://doi.org/10.1073/pnas.0802682105 [PubMed]
- 38. Zhang Z, Zhang Y, Sun XX, Ma X, Chen ZN. microRNA-146a inhibits cancer metastasis by downregulating VEGF through dual pathways in hepatocellular carcinoma. Mol Cancer. 2015; 14:5. https://doi.org/10.1186/1476-4598-14-5 [PubMed]
- 39. Kanai M, Hamada J, Takada M, Asano T, Murakawa K, Takahashi Y, Murai T, Tada M, Miyamoto M, Kondo S, Moriuchi T. Aberrant expressions of HOX genes in colorectal and hepatocellular carcinomas. Oncol Rep. 2010; 23:843–51. [PubMed]