



Introduction
Histone deacetylase (HDAC) proteins are highly conserved enzymes whose major role is regulation of chromatin structure. Histone deacetylation increases chromatin compaction with subsequent inhibition of gene transcription. Class I HDACs are zinc-dependent histone deacetylases and include HDACs 1, 2, 3, and 8. HDAC1 affects a variety of processes such as proliferation, differentiation, and development [1,2]. Disruption of HDAC1 function has been associated with many disorders including cancer and neurodegenerative diseases including Alzheimer’s disease [3,4]. HDAC inhibitors are now used clinically for the treatment of several disorders including malignancies [5]. Rpd3, an orthologue of mammalian HDAC1, can also deacetylate lysine residues of various other proteins, such as P53, or bind to promoter region of other genes (p21 and p57) and affect their transcription [6]. Reduced expression of Rpd3 extends lifespan in yeast, worms and fruit flies [7–12]. Similar effects on Drosophila lifespan were achieved by feeding fruit flies 4-phenylbutyrate (PBA), a histone deacetylase inhibitor [3]. Despite all of these studies, the mechanism of the beneficial effects of decreased rpd3 on longevity remains poorly understood.
The Insulin/insulin-like growth factor signaling (IIS) pathway is a nutrient-sensing pathway that regulates growth and development, energy homeostasis, stress response, and reproduction. Notably, mutations that reduce IIS activity are associated with longer lifespan in yeast, worms, flies, and mice [14–17]. Drosophila has eight insulin-like peptides (Dilps) that activate downstream events by binding to the insulin receptor [18]. dFOXO is the downstream target of IIS in flies [17]. When IIS is active, dFOXO is phosphorylated by dAkt, which leads to its binding to 14-3-3 proteins and its degradation. Reduced IIS results in decreased phosphorylation of dFOXO that promotes dFOXO nuclear translocation. dFOXO is a transcription factor and its nuclear localization is key to its influences on growth, stress resistance, and metabolism [19]. The direct targets of dFOXO are conserved across several different mammalian tissues and species. Overexpression of nuclear localized dFOXO in fat body/gut extends longevity in flies and worms [20,21]. In addition, overexpressing dFOXO in fly muscle extends lifespan [22].
Here, we investigate the effects and the mechanism of rpd3 reduction on fly metabolism, stress resitance, and longevity. We found that flies with reduced rpd3 levels have increased energy storage illustrated by increased levels of glucose, glycogen, trehalose, and triglycerides, which is consistent with their increased resistance to starvation. rpd3 mutant flies have reduced IIS supported by decreased levels of InR, chico, and increased levels of dfoxo mRNA compared to controls. Genetic studies show an overlap between rpd3 and IIS longevity pathways supported by a shorter life and reduced stress resistance of male flies with mutations in both rpd3 and dfoxo compared to rpd3 single mutant flies. Our data confer a novel link between rpd3 and IIS and suggest IIS as a potential downstream mediator of the effects of rpd3 mutation on fly health and metabolism.
Results
rpd3 reduction affects energy storage in flies
To examine the mechanism of the longevity extension observed in rpd3 mutant flies we examined how Rpd3 reduction affects fly physiology by examining resistance to starvation, oxidative stress, and fly metabolism. We used two different heterozygous rpd3 mutant alleles and their genetic controls, since homozygous rpd3 mutation is embryonic lethal [23]. We used rpd3 deficient (rpd3def/+) and their genetic controls, F1 progeny of rpd3def/+ littermates. We also used rpd3P-UTR/+ flies, an rpd3 hypomorph, and rpd3P-1.8/+, which are genetic controls for rpd3P-UTR/+ flies and have rpd3 reduction only in the eyes [24]. Here we show that rpd3def/+ mutant flies have higher starvation resistance at 10 and 40 days of age compared to control flies (Fig. 1 A,B; Supplemental Table 1A). Male rpd3def/+ flies are 38% and 44% more resistant to starvation at ages 10 and 40 days, respectively. Female rpd3def/+ flies are 28% and 108% more resistant at 10 and 40 days, respectively. To examine the potential mechanism of increased starvation resistance in rpd3 alleles we examined the effects of rpd3 reduction on fly metabolism. We quantified various forms of energy storage for the two rpd3 mutant alleles. At 10 days of age rpd3def/+ females have increased triglyceride levels, but reduced glucose and glycogen levels (Fig. 1C-G). At 40 days, rpd3def/+ female flies have increased levels of glucose, glycogen, trehalose, and triglycerides. rpd3def/+ males have increased levels of glucose and trehalose at 40 days, while no changes were observed at 10 days (Fig. 1C-F). Consistent with increased energy storage rpd3def/+ flies weighed more than control flies (Fig. 1H).
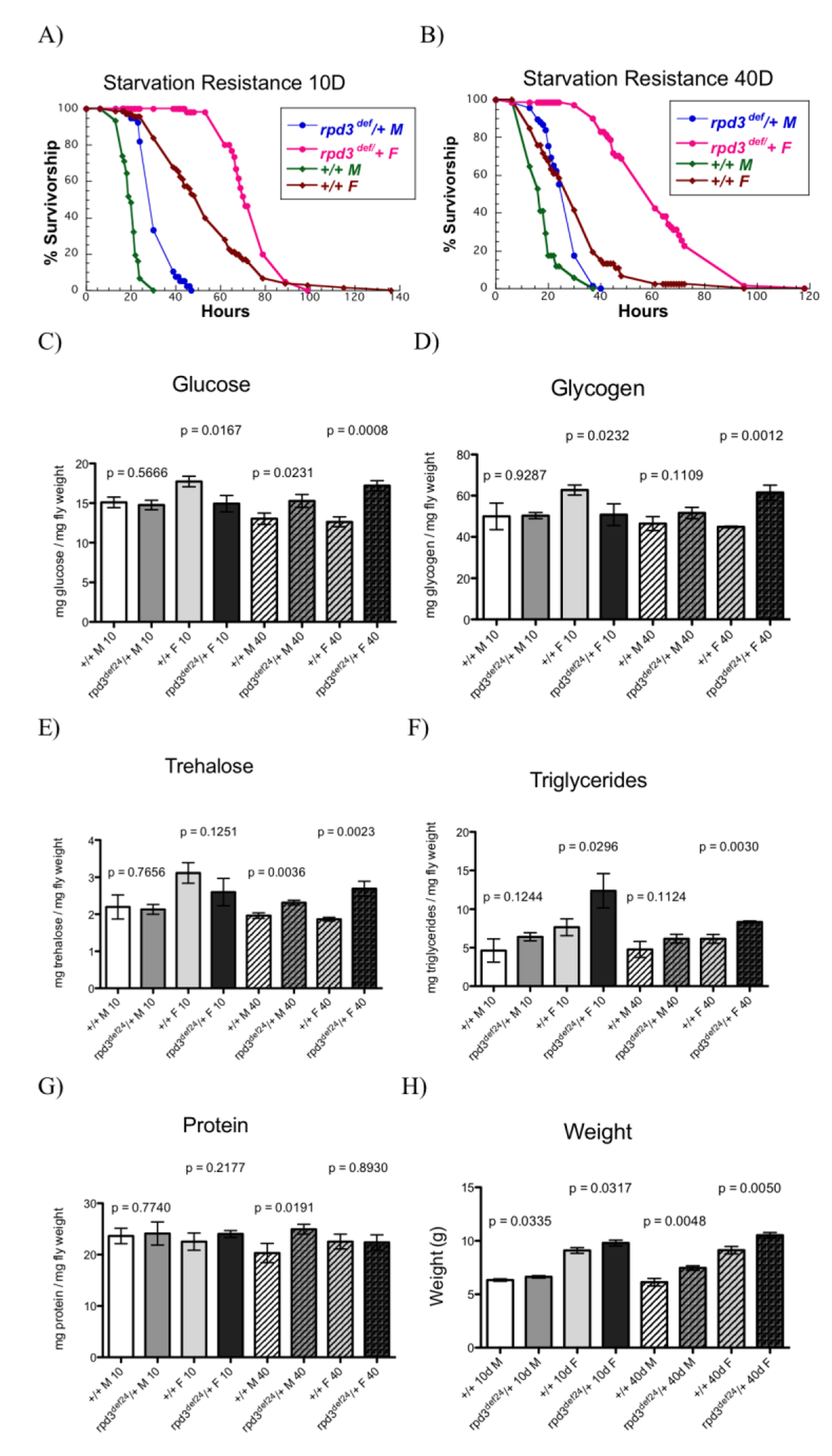
Figure 1. rpd3 reduction affects stress resistance and metabolism. (A,B) Reduced rpd3 levels increase stress resistance. Survival curves for male and female rpd3def/+ and control flies during starvation at age 10 (A) and 40 (B). (C-G) rpd3 reduction affects intermediary metabolism: Total levels of glucose (C), glycogen (D), trehalose (E), triglyceride (F) and protein (G) in rpd3def/+ and control male and female flies at age 10 and 40 days. (H) Weight of rpd3def and control flies used in C-G. Data are presented as means + SD (n=3, 30 flies per replicate. t test).
rpd3P-UTR /+ females live longer during starvation due to metabolic adaptation
At age 10, male and female rpd3P-UTR/+ flies have the same levels of energy stores as controls (Fig. 2A-F). At age 40, male rpd3P-UTR/+ flies have increased levels of triglycerides but reduced trehalose, glucose, and glycogen. When kept on standard lab food, female rpd3P-UTR/+ have the same triglyceride levels as controls at 40 days. To examine how rpd3 reduction affects fly adaptation to starvation we kept flies at high calorie food (1.5N) for 10 or 40 days and then starved them for 24 hours. Triglyceride levels were much higher in rpd3P-UTR/+ female, compared to levels found in rpd3P-1.8/+ control female flies (Fig. 2G). Consistently, an increase in starvation resistance was observed in rpd3P-UTR/+ flies when compared to rpd3P-1.8/+ flies (Fig. 2H, Supplemental Table 1B). These data suggest that reduction in rpd3 levels facilitates female adaptation to starvation, most likely by increasing efficiency in using metabolic reserves [25]. This is supported by our findings that rpd3P-UTR/+ flies have increased starvation resistance and consistent with our recent report that rpd3P-UTR/+ flies live longer compared to controls in conditions similar to starvation [11].
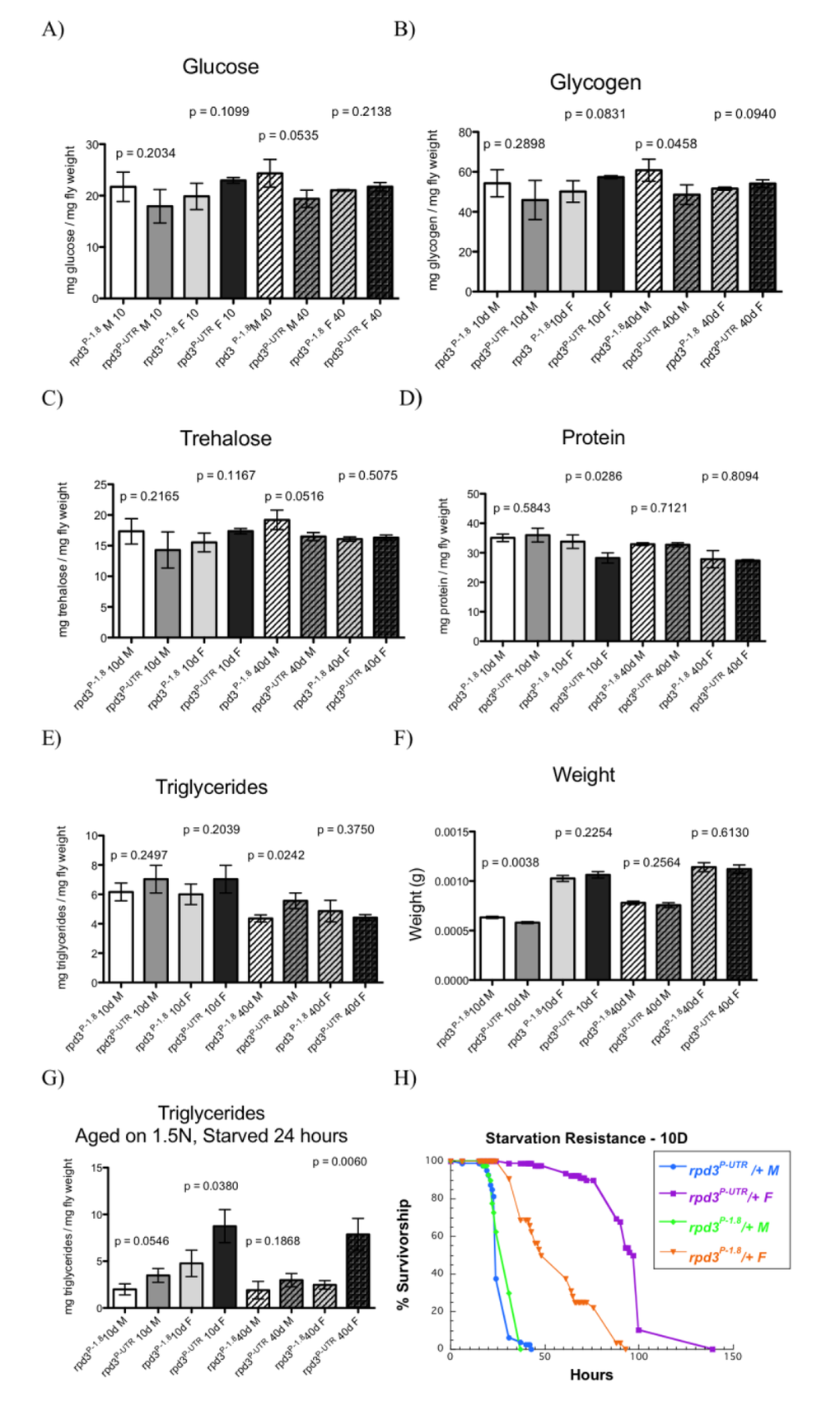
Figure 2. rpd3 reduction affects fly intermediary metabolism. (A-E) Total glucose (A), glycogen (B), trehalose (C), protein (D) and triglyceride (E) levels in rpd3P-UTR/+ (experimental) and rpd3P-1.8/+ (control) male and female flies at age 10 and 40 days. F) Weight of male and female flies used in (C-E). (G). Total triglyceride levels in rpd3P-UTR/+ and rpd3P-1.8 /+ flies aged on 1.5N for 10 or 40 days and then starved for 24 hours. Data are presented as means + SD (n=3, 30 flies per replicate. t test) (H) rpd3 reduction increases stress resistance in flies. Survival curves of rpd3P-UTR/+ and rpd3P-1.8/+ male and female flies during starvation at 10 of age.
rpd3-mutant flies have altered gene expression of components of the IIS pathway
The IIS pathway plays an important role in metabolism, stress resistance, and aging. Decreasing IIS by inhibiting InR, chico, or ablating the IPCs of the brain has been shown to extend the lifespan of fruit flies [26]. Overexpressing dFOXO, a downstream target of the IIS pathway, increases fly stress resistance. Since both reduction of rpd3 and of different members of IIS pathway extend fly longevity and increase stress resistance, we examined if these two longevity pathways overlap. We first examined how aging affects transcriptional levels of rpd3 and different members of IIS in a control strain of Drosophila, Canton-S (CS) at age 10, 20, 40, and 60 days. We found age-related increase in the levels of rpd3 and dInR mRNA in the heads and thoraces of CS flies and no changes in mRNA levels of dfoxo (Fig. 3A-C). Next we examined the status of IIS in rpd3 mutant flies and determined the levels of genes of members of the IIS pathway in the heads and thoraces (H + T) of rpd3def/+ flies at 20 and 40 days of age. The mRNA levels of InR and chico were significantly decreased in heads and thoraces of rpd3def mutants compared to their genetic controls at age 40 (Fig. 3D-I). The mRNA levels of PI3K, another member of IIS pathway, were the same as in controls (Fig. 3G). dFOXO is a downstream effector of the IIS pathway, whose activity is inversely related to IIS. Consistently with reduced IIS in rpd3 mutant flies, we found increased dfoxo mRNA expression in both rpd3def/+ flies and rpd3P-UTR/+ flies at 40 of age (Fig. 3H,I). We used only thoraces of rpd3P-UTR because their genetic controls rpd3P-1.8 flies have reduced levels of rpd3 mRNA in the eyes. Similarly, increased expression of dfoxo mRNA was found in whole body RNA isolated from heart-specific rpd3 downregulation [12].
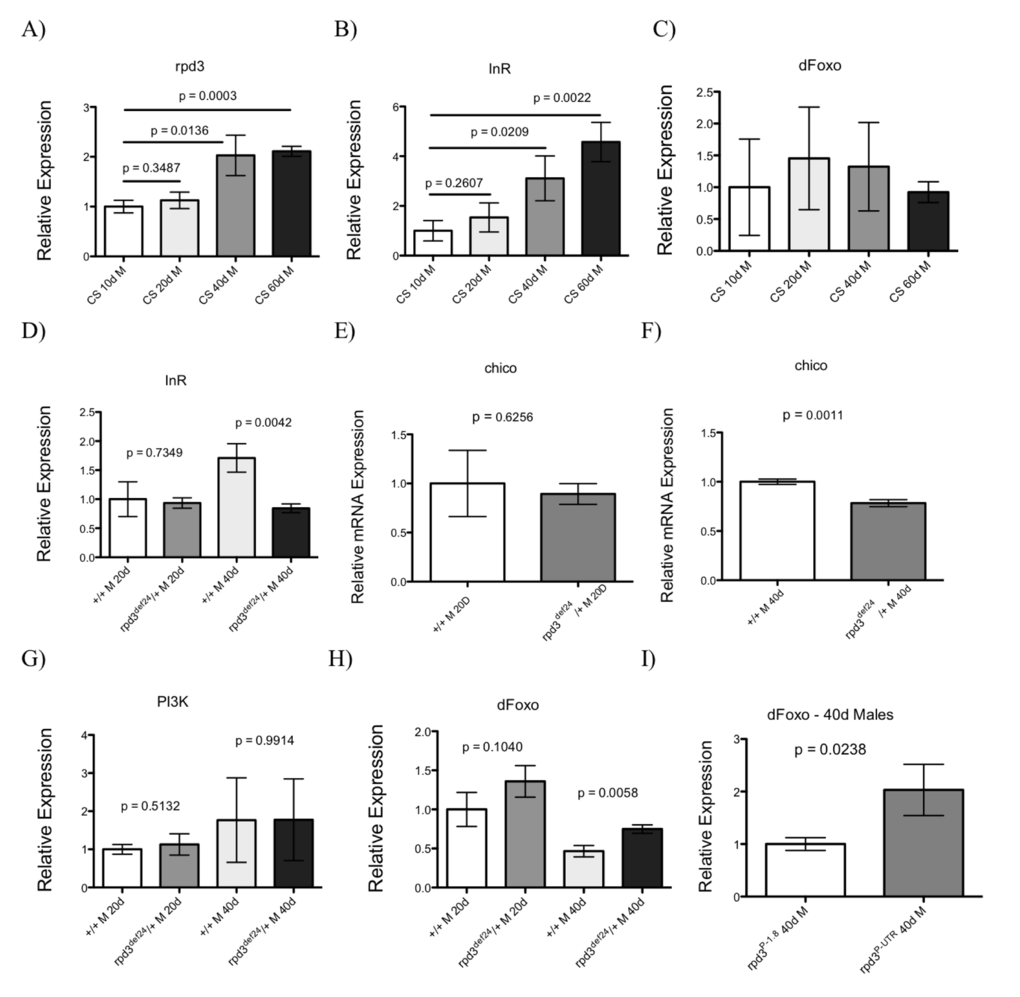
Figure 3. rpd3 reduction decreases IIS. (A-C) Aging affects rpd3 and InR mRNA levels. The levels of rpd3 (A), InR (B) or dfoxo (C) mRNA in the heads and thoraces of Canton S (CS) wild type male flies at 10, 20, 40 or 60 days determined by qPCR. Controls show an age-related increase in rpd3 and InR mRNA levels. (D-I) The levels of InR (D), chico (E,F), PI3K (G), and dfoxo (H) mRNA in heads and thoraces of rpd3def/yw male flies at ages 20 and 40 and their genetic controls determined by qPCR. (I) The levels of dfoxo mRNA are increased in thoraces of rpd3P-UTR/CS male flies compared to rpd3P-1.8/CS controls at 40 days (n=3, A-H: 30 heads and thoraces per replicate. I: 30 thoraces per replicate. p as noted, t test).
When the IIS is active, dFOXO is phosphorylated by Akt and localized in cytoplasm. Reduction in IIS decreases dFOXO phosphorylation, which allows nuclear dFOXO localization and activation of transcription of many genes known to mediate beneficial effects of reduced IIS. Our Western blots did not reveal any difference in the levels of phosphorylated dFOXO or in the ratio of nuclear and cytoplasmic fraction (Supplemental Figure 1A-C). Nevertheless, this result could be due to low stability of the FOXO protein or possibly due to the quality of our anti-dFOXO antibody. Our data show that rpd3 reduction prevents age-related increase in InR and reduction in dfoxo mRNA and suggest that decreased IIS could mediate the effects of rpd3 mutation on Drosophila longevity.
dfoxo partially mediates lifespan extension and increased stress resistance in rpd3-mutant flies
To strenghten the link between rpd3 mutation and reduced IIS, we generated flies that were double mutants for both rpd3 and dfoxo and examined their longevity and stress resistance. dfoxo is the downstream IIS target that mediates beneficial effects of reduced IIS signaling. We confirmed that the levels of rpd3 and dfoxo expression were reduced based on the presence of the mutations in male flies (Fig. 4A,B). rpd3def/yw mutants and rpd3def/dfoxoc01841 double mutants had lower levels of rpd3 mRNA expression compared to dfoxoc01841/yw mutant flies (Fig. 4A). dfoxoc01841/yw and rpd3def/dfoxoc01841 males have decreased levels of dfoxo mRNA expression compared to rpd3def/yw flies (Fig. 4B). Notably, male rpd3def/dfoxoc01841 double mutant flies have higher levels of dfoxo mRNA compared to single dfoxoc01841/yw flies, consistent with our data that rpd3 mutation increases dfoxo mRNA levels (Fig. 4B). Lifespan experiments were performed on these flies. Heterozygous rpd3def/+ males lived the longest (Fig. 4C, Supplemental Table 2) dfoxoc01841/yw males had the shortest lifespan, whereas rpd3def/dfoxoc01841 double mutants had a longevity in the middle of these two. These data suggest that the full beneficial effects of rpd3 mutation on fly longevity require both copies of dfoxo, and that small increases in dfoxo levels in rpd3def/dfoxoc01841may increase longevity of double mutant flies compared to dfoxoc01841/yw males. In contrast, female rpd3def/dfoxoc01841 flies live longer compared to single mutant flies, which have similar lifespans. The differences between male and female in longevity effects of double mutations may be due to sexual dimorphism previously described to be associated with IIS, FOXO, and p53 [27]. Similar differences were observed in stress response, which is shown in next paragraph and discussed later.
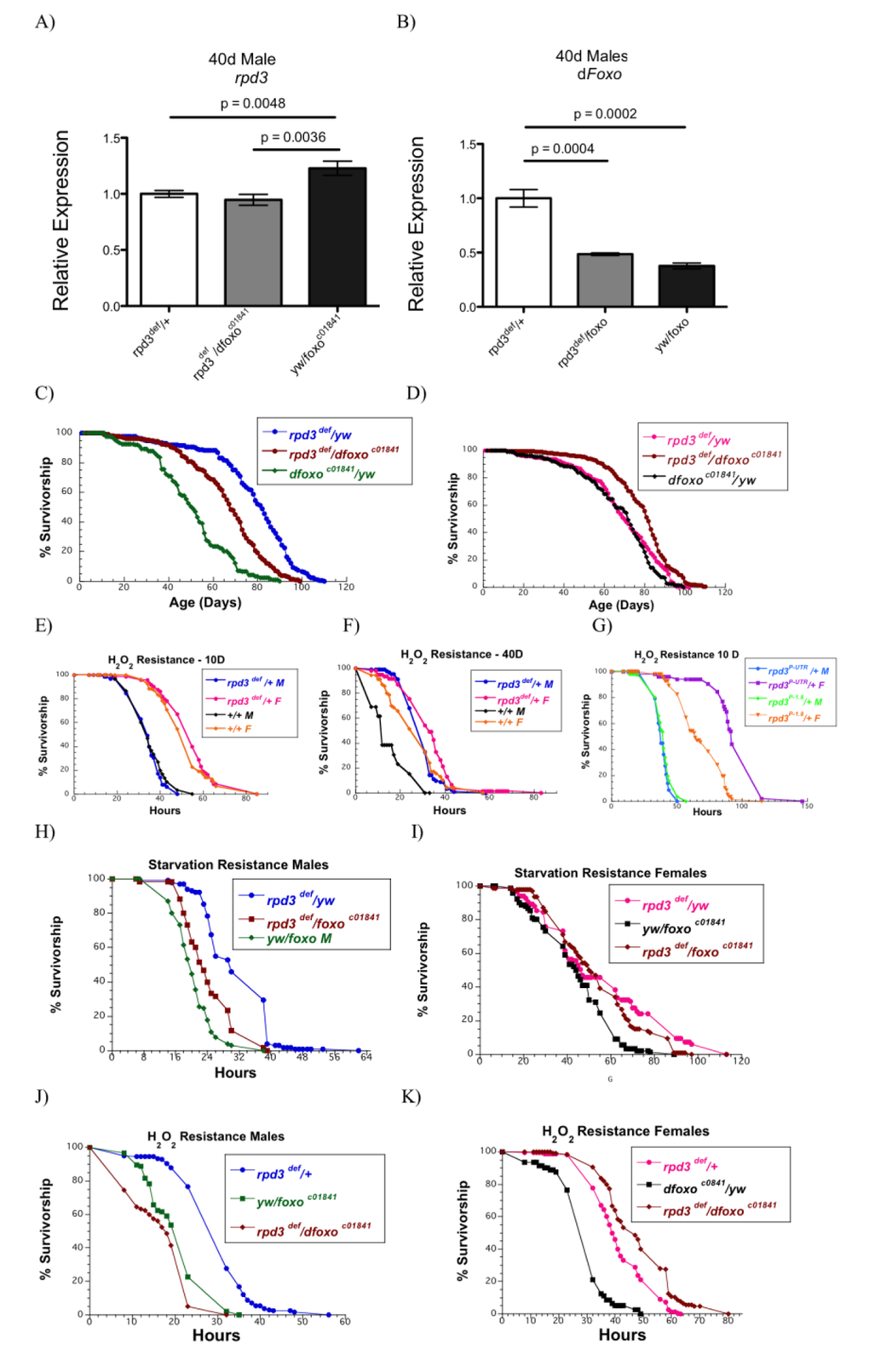
Figure 4. dfoxo partially mediates effects on longevity and stress resistance observed in male rpd3 mutant flies. (A, B) rpd3 (A) and dfoxo (B) mRNA levels in the heads and thoraces of rpd3def/yw, rpd3def/dfoxoc01841and dfoxoc01841/yw male flies at 40 days determined by qPCR (n=3, 30 Heads and thoraces per replicate. p as noted, t test, error bars represent SEM). (C,D) Survival curves of male (C) and female (D) rpd3def/yw, rpd3def/dfoxoc01841 and dfoxoc01841/yw flies. (E-F) Survival curves of rpd3def/+ and control flies male and female flies on 5% H2O2 at age 10 (E) and 40 (F) days. (G) Survival curves of rpd3P-UTR/+ and rpd3P-1.8/+ male and female flies exposed to 5% H2O2 at age 10 days of age. (H, K) Survival curves for male (H, J) and female (I, K) rpd3def/yw, rpd3def/dfoxoc01841 and dfoxoc01841/yw flies during starvation (H, I) or on 5% H2O2 (J, K) at age 40.
Increased stress resistance in flies with reduced rpd3 and IIS
Reduced IIS is associated with increased resistance to oxidative stress. Since rpd3 reduction is associated with IIS reduction we examined if flies with reduced rpd3 levels are more resistant to hydrogen peroxide (H2O2). Male and female rpd3def/+ are more resistant to H2O2 compared to control flies at 40 days of age, while no difference was observed at age 10 days (Fig. 4E,F; Supplemental Table 3). Female rpd3P-UTR/+ flies are more resistant to H2O2 at both ages but no difference was observed in male rpd3P-UTR/+ flies (Fig 4G, Supplemental Table 3 and data not shown). To examine the role of dfoxo in increased stress resistance of rpd3 mutant flies, we determined starvation and H2O2 resistance of rpd3 mutant flies with or without dfoxo mutation. Similarly to longevity studies, rpd3def/+ male flies had the highest mean survival rate when exposed to starvation at 40d of age (Fig. 4H,I; SSupplemental Table 4). rpd3def/dfoxoc01841 double mutants had a lower mean survival, but they were more resistant to starvation than dfoxoc01841/yw flies (Fig. 4H,J). Similarly, male rpd3def/+ mutants are more resistant to H2O2 compared to rpd3def/dfoxoc01841 or dfoxoc01841/yw flies. These results suggest that dfoxo mediates some of the effects of rpd3 mutation on lifespan in Drosophila males and is required for full beneficial effects of rpd3 mutation on longevity and stress resistance. Female flies double mutant for rpd3def and dfoxoc01841 live longer and have similar starvation and H2O2 resistance compared to single rpd3def/+, but are more resistance compared to dfoxoc01841/yw mutant flies (Fig. 4D,I,K; Supplemental Table 5).
Discussion
rpd3 reduction affects fly metabolism and IIS
Members of the class I HDAC family are vital regulators of chromatin structure and gene expression. They have multiple functions including a role in development, metabolism, and aging [1,28]. Deletion of rpd3 in yeast extends their replicative lifespan [7]. Flies heterozygous for a null or a hypomorphic rpd3 mutant alleles had an extended lifespan compared to genetic controls [9,11]. Moreover, heart-specific rpd3 downregulation in flies increases heart function, stress resistance, and extends longevity [12]. In addition, HDAC inhibitors trichostatin A and butyrate also extend fly lifespan [13,29]. Longevity extension of rpd3 mutant flies was not further extended by dietary restriction (DR) and was absent in flies double mutant for dSir2 and rpd3 mutations [30]. Both DR flies and flies with reduced rpd3 have increased dSir2 levels [12,30]. These findings suggested that the mechanism of longevity extension in rpd3 mutant and DR flies overlap. However, the full understanding of the mechanism of longevity extension associated with rpd3 reduction is missing. Therefore we examined intermediary metabolism and starvation resistance of two rpd3 heterozygous mutant alleles. Increased starvation resistance in female rpd3def/+ flies is the result of increased energy storage in forms of triglyceride at young and old age, as well as increased carbohydrate levels at age 40. Metabolic adaptation to fasting is key to preserving homeostasis of an organism. This adaptation includes mobilization of lipids followed by their oxidation into ketone bodies, which are used as a source of energy in other tissues. Metabolic adaptation to starvation in rpd3 mutant flies is illustrated by the findings that rpd3 reduction prevents a fasting-induced decrease in triglycerides in female rpd3P-UTR/+ flies. This is consistent with our recent report that rpd3P-UTR/+ flies live longer compared to controls in conditions similar to starvation and findings that heart-specific Rpd3 downregulation increases fly resistance to starvation at age 2 days [11,12]. To get insights into the mechanism associated with rpd3 reduction we examined if these changes are mediated by IIS. The IIS pathway is a nutrient sensing pathway, which also affects the activity of metabolic enzymes. When nutrients are abundant IIS is active, dFOXO is phosphorylated, and it is localized in the cytoplasm. Reduced IIS decreases phosphorylation of dFOXO, which promotes dFOXO nuclear translocation. In the nucleus, dFOXO regulates glucose, glycogen, and lipid metabolism by activating transcription of key enzymes involved in these metabolic pathways. For instance, dFOXO activates glycogenolysis and gluconeogenesis by activating transcription of glucose-6-phosphatase (G6P) and phosphoenolpyruvate carboxykinase (PEPCK) mRNA, respectively [31]. dFOXO regulates autophagy in response to starvation, which promotes recycling of the cellular components [32]. Intriguingly, adult dFOXO null mutant flies have no difference in starvation resistance or energy storage [33]. Under conditions when nutrients are limited, activated dFOXO upregulates InR transcription. This allows the cells to accumulate InR mRNA, and prime them to respond quickly when nutrients become available [34]. Once IIS is activated, it will upregulate growth and inhibit dFOXO activity via its phosphorylation. Here we show that in a control strain of Drosophila, CS, the levels of InR mRNA expression gradually increase throughout the lifespan. We found that rpd3 reduction affects the IIS pathway and prevents age-associated changes in the transcript levels of IIS genes. rpd3def flies have reduced levels of InR and chico and increased dfoxo mRNA at 40 days. These changes in IIS are consistent with metabolic changes found in 40 day old rpd3 mutant flies.
dFOXO mediates some longevity effects observed in rpd3 mutant flies
Our genetic studies suggest that dFOXO mediates some longevity effects of rpd3def mutant flies. This is illustrated by shorter lifespan of male flies double mutant for rpd3 and dfoxo compared to rpd3def/+ flies. rpd3def/dfoxoc0184 flies live longer compared to dfoxo single mutant flies, which is most likely mediated by higher dfoxo mRNA expression in double mutants compared to dfoxo single mutants. Similarly, rpd3def/dfoxoc0184 flies have reduced resistance to H2O2 compared rpd3def/+ indicating that increased dFOXO mediates resistance to stress in rpd3 mutant male flies. This is consistent with findings that overexpression of nuclear localized dFOXO mediates increased resistance to oxidative stress in flies and mammalian cells [33]. Moreover, treatment with PBA, a HDAC1 inhibitor, increases expression of genes that have been implicated in response to oxidative stress, such as SOD, gluthathione S-transferase, and heat-shock protein. However, female rpd3def/dfoxoc01841 live longer and are similarly more resistant to H2O2 compared to both single rpd3def/+ and dfoxoc01841/+ mutant flies. It is possible that reduced rpd3 levels in rpd3def/+ female flies increase dfoxo levels of the remaining wild type copy of the gene, which contribute to longer lifespan. The differences may be also due to sexual dimorphism previously described to be associated with IIS, FOXO, and p53. It was found that nervous-system specific overexpression of p53 increases female lifespan in a foxo null background. In contrast, in males foxo null mutation caused the tissue-specific effects of p53 [27]. Several studies have examined the relationship between IIS and HDAC inhibition. β−hydroxybutyrate (βOHB) is an endogeneous inhibitor of HDACs 1, 3, and 4 (Class I and Class II). βOHB is one of the ketone bodies released during fasting and exercise. βOHB inhibition of HDAC1 and HDAC2 activity results in increased histone acetylation and gene expression. Particularly important is induction of Foxo3, the mammalian orthologue of the dFOXO, expression by removing HDAC-mediated Foxo3 repression via hypoacetylation of its promoter [35]. Ye reviewed the effects of an HDAC1 inhibitor on energy metabolism and insulin sensitivity [36]. It was also reported that use of butyrate, a class I and II HDAC inhibitor, improves glucose metabolism and prevents age-related atrophy [37]. However, less is known about the specific role of rpd3 reduction on metabolism in flies. The data presented here provide new information about the effects of rpd3 reduction on fly metabolism and link the changes in metabolism to a reduction in IIS. Taken together, our genetic data strenghten the link between rpd3 reduction and reduced IIS.
Longevity extension in rpd3 mutant flies has been linked to dSir2, 4E-BP, dFOXO, and CR longevity pathways [9,11,12,30]. The data presented here add to our understanding of the mechanism of rpd3’s effects on longevity by identifying a novel genetic link between rpd3 reduction and IIS (Fig. 5). However, considering that rpd3 has multiple targets, the reduction of rpd3 does not completely reproduce phenotypes of the flies with reduced IIS. In Drosophila, reduction in IIS results in female sterility and longevity extension independent of fertility [15,16]. Chico mutants are smaller and sterile, but they contain twice as much lipid content per weight as do genetic controls [38]. Overexpression of dfoxo in adult fat body and the gut has no effect on fecundity, total triglyceride, total trehalose, or total glycogen content, but it reduced fly weight and total protein content [39]. rpd3 mutant flies have reduced levels of InR and chico, increased levels of dfoxo mRNA, and increased resistance to starvation and oxidative stress. However, rpd3 flies are as fertile as controls on a standard diet, and their weight is higher or the same compared to controls. These differences highlight the complex mechanism of the beneficial effects of rpd3 reduction on fly health and longevity.
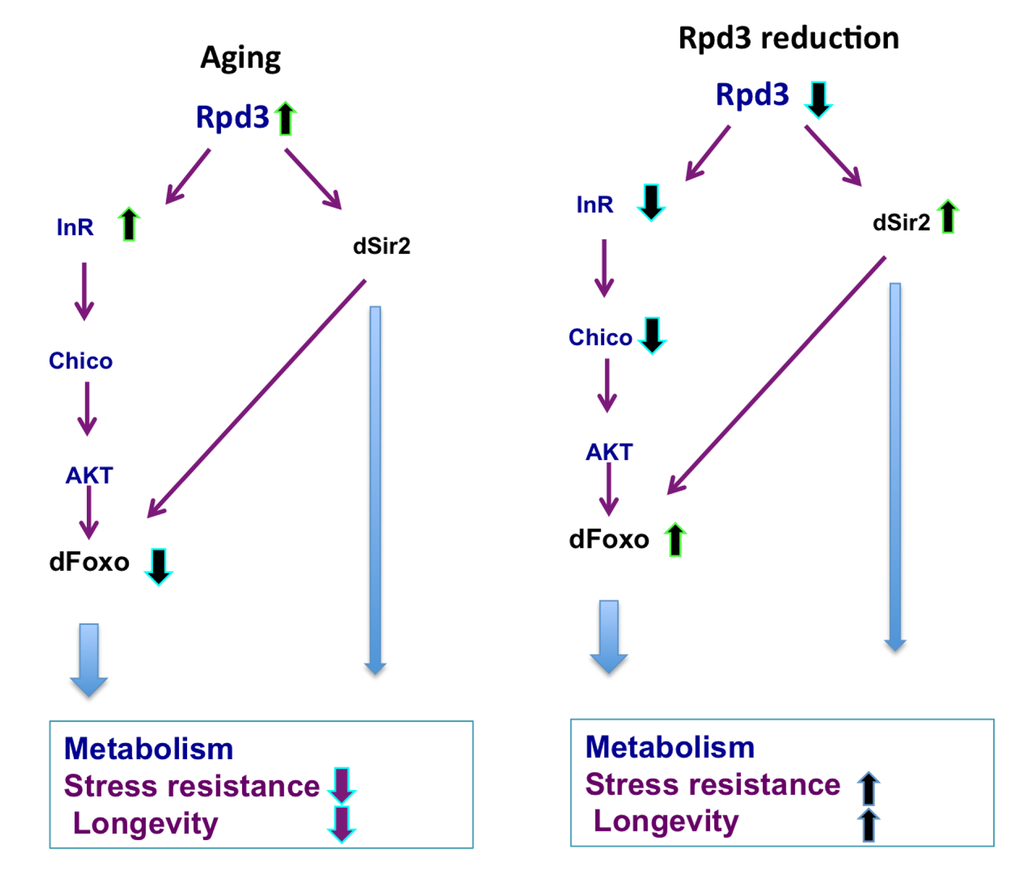
Figure 5. rpd3 reduction prevents age-associated changes in IIS. Age-associated increases in the rpd3, InR, and a decrease in dfoxo mRNA are observed in wild type flies, Reduced rpd3 activity decreases InR and chico mRNA, while increases dfoxo and dSir2 mRNA levels. Reduced rpd3 affects metabolism, increases stress resistance and longevity by reducing IIS and increasing dSir2 levels. Purple and light blue arrows indicate downstream effects, green arrows represent increase and blue reduction in mRNA levels.
Here we show that rpd3 reduction affects fly metabolism characterized by increased energy storages, higher resistance to starvation and oxidative stress, and increased longevity. These effects are mediated at least partially by reduced IIS, confirmed by genetic studies showing that longevity extension requires increased dFOXO (Fig. 5). Previous studies highlight the role of dSir2 in rpd3 effects on lifespan [12,30]. Histone modifying enzymes link changes in nutrient availability to changes in intermediary metabolism by affecting the activity and stability of the enzymes involved in glycogenesis, glycolysis, gluconeogenesis, and β-oxidation through acetylation [40–42]. The acetylation pattern differs in tissues and cell types, suggesting a complex, highly orchestrated regulation of acetylation levels at the organismal level. Future studies on the acetylation status of enzymes involved in intermediary metabolism in different tissues of rpd3 mutant flies would further expand our knowledge of the effects of rpd3 reduction on metabolism, health, and longevity. Our data illustrates how these complex interactions result in phenotypic changes at the organismal level. Further studies are necessary to examine how tissue-specific alterations in rpd3 levels orchestrate these changes.
Materials and Methods
Fly strains and maintenance
rpd3-deficient (rpd3def24) and rpd3–hypomorphic (rpd3P-UTR) flies and their genetic controls were used in the experiments. Genetic controls for rpd3def24 were progeny generated by crossing F1 rpd3def24/+ littermates. The hypomorphic rpd3P-UTR allele has a P-element inserted in the 5’UTR region of the rpd3 genes, which affects expression throughout the fly’s bod. The control rpd3P-1.8 allele has a P-element inserted 1.8 Kb upstream from the transcriptional start site, which only decreases expression in the eye [24]. Canton S, yw, and dfoxoc01841 were kindly provided by the Bloomington Stock center. dfoxoc01841 flies were backcrossed to yw strain for 10 generations to eliminate difference in genetic background. Flies were collected within 24 hours of eclosion and maintained using standard culture media in plastic vials. They were kept at 25oC in a humidified incubator. About 25 males and 25 females were kept together in each vial, and they were passed to a fresh vial every Monday, Wednesday, and Friday. Standard corn and 1.5N (High calorie diet) diet was prepared as previously reported [43].
Lifespan studies
Flies were collected within 24 hours of eclosion and maintained as described above. 25 males or 25 females were placed in each vial. They were passed every 2 days up to age 30 days and every day after that and the number of dead flies were counted. The number of flies in each survivorship study is listed in Tables 1-7.
Starvation and oxidative stress resistance
Flies were collected as described above and aged until 10 or 40 days of age. They were separated into vials of 20 males or 20 females and transferred into new vials containing 2 filter papers with 300 μl of DI H2O for starvation studies. 300 μl of 5% H2O2 with 5% sucrose was used instead of water for oxidative stress studies following initial 6 hours of starvation. The number of dead flies was counted at regular intervals, and the vials were changed ever 24 hours.
Resistance to paraquat: flies were collected as described above. 20 flies were transferred to a vial containing filter paper soaked with 300 μL of 20 mM paraquat following initial starvation for 6 hours. The number of dead flies was counted hourly during the day and twice overnight until no flies remained alive. Stress resistance data were analyzed by log-rank tests using the JMP 10 program. Total number of flies per experiment is listed in Supplemental Tables 1, 2 and 3.
Biochemistry
Flies were collected and aged as described above. At ages 10 and 40 days, flies were weighed, homogenized, and 25 μl of homogenate was aliquoted into 96-well plates. For glucose, PGO enzyme plus color reagent was added to each well, the plate was incubated at 37oC, and the optic density was read at 450 nm. For glycogen, the procedure was the same except amyloglucosidase was added to each well in addition to the other enzyme. For trehalose, the procedure was the same as glucose except the samples were incubated with trehalase before addition of PGO. For protein, BioRad protein solution was diluted 1:5, added to each well, and then the plate was read at 595 nm. For triglycerides, free glycerol and triglyceride reagents were added to each well, the plate was incubated at 37oC, and the optic density was read at 540 nm [45,46].
Quantitative PCR (qPCR)
Flies were frozen at the appropriate age, and RNA was isolated from the heads and thoraces, thoraces, or heads. cDNA was synthesized from RNA. Using TaqMan primers and the Applied Biosystems Thermal Cycler, levels of gene expression were determined. The data were normalized to the results of the control flies using ankryn as standard.
Western blots
40 day old flies were dissected on CO2 and placed in tubes containing Kinase Lysis Buffer. The tissue was homogenized using Teflon pestles. Protein concentrations were quantified using BioRad Protein determination Kit. Lysates were cleared by centrifugation, run on a Lithium Dodecyl Sulfate (LDS) PAGE using NuPAGE NOVEX 4-12% gradient gels (Life Technologies), and transferred overnight onto nitrocellulose membranes in transfer buffer (20mM CAPS pH 11, 20% MeOH). Western blotting was performed using standard procedures with washes done in TBST. Membranes were blocked in 5% milk for an hour followed by overnight, 1:500 primary antibody incubation at 4oC. Secondary antibody incubation was done for an hour at room temperature at a concentration of 1:5,000. Secondary antibodies were labeled with Horseradish Peroxidase. Blots were imaged using the Kodak Image Station 4000 MM following application of Enhanced Chemiluminensence reagent (Perkin Elmer). The dFOXO antibody was a gift from Mark Tatar, and the two anti-phospho-Foxo3A antibodies (Cat #9466) were gifts from Cell Signaling Technology to test if they would work in Drosophila tissue. Nuclear and cytoplasmic fractions were isolated from 40-day old flies following instruction for The Active Motif Nuclear Extract Kit (Carlsbad, CA. USA, Cat #40010).
Statistical analysis
Significance was determined using a two-tailed, unpaired t-test from at least three independent experiments and expressed as P values. P < 0.05 is considered to be significant. P values are specifically indicated in each figure. Error bars represent standard deviation (SD). Longevity data were censored for early mortality (1-9 Days) and analyzed by log-rank tests using the JMP 12 program.
Supplementary Materials
Acknowledgements
We would like to thank Suzanne Kowalski, Alexander Pokorski, Bhavin Gupta and Dr. Ryan Rogers for excellent technical assistance. We thank Dr. Stewart Frankel for critical reading of the manuscript.
Conflicts of Interest
The authors have no conflict of interests to declare.
Funding
This work was supported by grant from the National Institute on Health RO1AG 023088 to B. Rogina. B. Rogina is a recipient of a Glenn Award for Research in Biological Mechanisms of Aging.
References
- 1. Seto E, Yoshida M. Erasers of histone acetylation: the histone deacetylase enzymes. Cold Spring Harb Perspect Biol. 2014; 6:a018713. https://doi.org/10.1101/cshperspect.a018713 [PubMed]
- 2. Moser MA, Hagelkruys A, Seiser C. Transcription and beyond: the role of mammalian class I lysine deacetylases. Chromosoma. 2014; 123:67–78. https://doi.org/10.1007/s00412-013-0441-x [PubMed]
- 3. Willis-Martinez D, Richards HW, Timchenko NA, Medrano EE. Role of HDAC1 in senescence, aging, and cancer. Exp Gerontol. 2010; 45:279–85. https://doi.org/10.1016/j.exger.2009.10.001 [PubMed]
- 4. Woods JK, Rogina B. The effects of Rpd3 on fly metabolism, health, and longevity. Exp Gerontol. 2016; :S0531-5565(16)30056-0. https://doi.org/10.1016/j.exger.2016.02.015 [PubMed]
- 5. Drummond DC, Noble CO, Kirpotin DB, Guo Z, Scott GK, Benz CC. Clinical development of histone deacetylase inhibitors as anticancer agents. Annu Rev Pharmacol Toxicol. 2005; 45:495–528. https://doi.org/10.1146/annurev.pharmtox.45.120403.095825 [PubMed]
- 6. Juan LJ, Shia WJ, Chen MH, Yang WM, Seto E, Lin YS, Wu CW. Histone deacetylases specifically down-regulate p53-dependent gene activation. J Biol Chem. 2000; 275:20436–43. https://doi.org/10.1074/jbc.M000202200 [PubMed]
- 7. Kim S, Benguria A, Lai C-Y, Jazwinski SM. Modulation of life-span by histone deacetylase genes in Saccharomyces cerevisiae.. Mol Biol Cell. 1999; 10:3125–36. https://doi.org/10.1091/mbc.10.10.3125 [PubMed]
- 8. Edwards C, Canfield J, Copes N, Rehan M, Lipps D, Bradshaw PC. D-beta-hydroxybutyrate extends lifespan in C. elegans.. Aging (Albany NY). 2014; 6:621–44. https://doi.org/10.18632/aging.100683 [PubMed]
- 9. Rogina B, Helfand SL, Frankel S. Longevity regulation by Drosophila Rpd3 deacetylase and caloric restriction. Science. 2002; 298:1745. https://doi.org/10.1126/science.1078986 [PubMed]
- 10. Pallos J, Bodai L, Lukacsovich T, Purcell JM, Steffan JS, Thompson LM, Marsh JL. Inhibition of specific HDACs and sirtuins suppresses pathogenesis in a Drosophila model of Huntington’s disease. Hum Mol Genet. 2008; 17:3767–75. https://doi.org/10.1093/hmg/ddn273 [PubMed]
- 11. Frankel S, Woods J, Ziafazeli T, Rogina B. RPD3 histone deacetylase and nutrition have distinct but interacting effects on Drosophila longevity. Aging (Albany NY). 2015; 7:1112–29. https://doi.org/10.18632/aging.100856 [PubMed]
- 12. Kopp ZA, Hsieh JL, Li A, Wang W, Bhatt DT, Lee A, Kim SY, Fan D, Shah V, Siddiqui E, Ragam R, Park K, Ardeshna D, et al. Heart-specific Rpd3 downregulation enhances cardiac function and longevity. Aging (Albany NY). 2015; 7:648–63. https://doi.org/10.18632/aging.100806 [PubMed]
- 13. Kang H-L, Benzer S, Min K-T. Life extension in Drosophila by feeding a drug. Proc Natl Acad Sci USA. 2002; 99:838–43. https://doi.org/10.1073/pnas.022631999 [PubMed]
- 14. Kenyon C, Chang J, Gensch E, Rudner A, Tabtiang R. A C. elegans mutant that lives twice as long as wild type. Nature. 1993; 366:461–64. https://doi.org/10.1038/366461a0 [PubMed]
- 15. Clancy DJ, Gems D, Harshman LG, Oldham S, Stocker H, Hafen E, Leevers SJ, Partridge L. Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein. Science. 2001; 292:104–06. https://doi.org/10.1126/science.1057991 [PubMed]
- 16. Tatar M, Kopelman A, Epstein D, Tu M-P, Yin C-M, Garofalo RS. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science. 2001; 292:107–10. https://doi.org/10.1126/science.1057987 [PubMed]
- 17. Yamamoto R, Tatar M. Insulin receptor substrate chico acts with the transcription factor FOXO to extend Drosophila lifespan. Aging Cell. 2011; 10:729–32. https://doi.org/10.1111/j.1474-9726.2011.00716.x [PubMed]
- 18. Kannan K, Fridell YW. Functional implications of Drosophila insulin-like peptides in metabolism, aging, and dietary restriction. Front Physiol. 2013; 4:288. https://doi.org/10.3389/fphys.2013.00288 [PubMed]
- 19. Webb AE, Kundaje A, Brunet A. Characterization of the direct targets of FOXO transcription factors throughout evolution. Aging Cell. 2016; 15:673–85. https://doi.org/10.1111/acel.12479 [PubMed]
- 20. Giannakou ME, Goss M, Jünger MA, Hafen E, Leevers SJ, Partridge L. Long-lived Drosophila with overexpressed dFOXO in adult fat body. Science. 2004; 305:361. https://doi.org/10.1126/science.1098219 [PubMed]
- 21. Hwangbo DS, Gershman B, Tu MP, Palmer M, Tatar M. Drosophila dFOXO controls lifespan and regulates insulin signalling in brain and fat body. Nature. 2004; 429:562–66. https://doi.org/10.1038/nature02549 [PubMed]
- 22. Demontis F, Perrimon N. FOXO/4E-BP signaling in Drosophila muscles regulates organism-wide proteostasis during aging. Cell. 2010; 143:813–25. https://doi.org/10.1016/j.cell.2010.10.007 [PubMed]
- 23. Mannervik M, Levine M. The Rpd3 histone deacetylase is required for segmentation of the Drosophila embryo. Proc Natl Acad Sci USA. 1999; 96:6797–801. https://doi.org/10.1073/pnas.96.12.6797 [PubMed]
- 24. Mottus R, Sobel RE, Grigliatti TA. Mutational analysis of a histone deacetylase in Drosophila melanogaster: missense mutations suppress gene silencing associated with position effect variegation. Genetics. 2000; 154:657–68. [PubMed]
- 25. Rion S, Kawecki TJ. Evolutionary biology of starvation resistance: what we have learned from Drosophila.. J Evol Biol. 2007; 20:1655–64. https://doi.org/10.1111/j.1420-9101.2007.01405.x [PubMed]
- 26. Kenyon CJ. The genetics of ageing. Nature. 2010; 464:504–12. https://doi.org/10.1038/nature08980 [PubMed]
- 27. Shen J, Tower J. Drosophila foxo acts in males to cause sexual-dimorphism in tissue-specific p53 life span effects. Exp Gerontol. 2010; 45:97–105. https://doi.org/10.1016/j.exger.2009.10.009 [PubMed]
- 28. Mihaylova MM, Shaw RJ. Metabolic reprogramming by class I and II histone deacetylases. Trends Endocrinol Metab. 2013; 24:48–57. https://doi.org/10.1016/j.tem.2012.09.003 [PubMed]
- 29. Zhao Y, Sun H, Lu J, Li X, Chen X, Tao D, Huang W, Huang B. Lifespan extension and elevated hsp gene expression in Drosophila caused by histone deacetylase inhibitors. J Exp Biol. 2005; 208:697–705. https://doi.org/10.1242/jeb.01439 [PubMed]
- 30. Rogina B, Helfand SL. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc Natl Acad Sci USA. 2004; 101:15998–6003. https://doi.org/10.1073/pnas.0404184101 [PubMed]
- 31. Barthel A, Schmoll D, Unterman TG. FoxO proteins in insulin action and metabolism. Trends Endocrinol Metab. 2005; 16:183–89. https://doi.org/10.1016/j.tem.2005.03.010 [PubMed]
- 32. Martins R, Lithgow GJ, Link W. Long live FOXO: unraveling the role of FOXO proteins in aging and longevity. Aging Cell. 2016; 15:196–207. https://doi.org/10.1111/acel.12427 [PubMed]
- 33. Jünger MA, Rintelen F, Stocker H, Wasserman JD, Végh M, Radimerski T, Greenberg ME, Hafen E. The Drosophila forkhead transcription factor FOXO mediates the reduction in cell number associated with reduced insulin signaling. J Biol. 2003; 2:20. https://doi.org/10.1186/1475-4924-2-20 [PubMed]
- 34. Puig O, Tjian R. Transcriptional feedback control of insulin receptor by dFOXO/FOXO1. Genes Dev. 2005; 19:2435–46. https://doi.org/10.1101/gad.1340505 [PubMed]
- 35. Shimazu T, Hirschey MD, Newman J, He W, Shirakawa K, Le Moan N, Grueter CA, Lim H, Saunders LR, Stevens RD, Newgard CB, Farese RV
Jr , de Cabo R, et al. Suppression of oxidative stress by β-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science. 2013; 339:211–14. https://doi.org/10.1126/science.1227166 [PubMed] - 36. Ye J. Improving insulin sensitivity with HDAC inhibitor. Diabetes. 2013; 62:685–87. https://doi.org/10.2337/db12-1354 [PubMed]
- 37. Walsh ME, Bhattacharya A, Sataranatarajan K, Qaisar R, Sloane L, Rahman MM, Kinter M, Van Remmen H. The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell. 2015; 14:957–70. https://doi.org/10.1111/acel.12387 [PubMed]
- 38. Böhni R, Riesgo-Escovar J, Oldham S, Brogiolo W, Stocker H, Andruss BF, Beckingham K, Hafen E. Autonomous control of cell and organ size by CHICO, a Drosophila homolog of vertebrate IRS1-4. Cell. 1999; 97:865–75. https://doi.org/10.1016/S0092-8674(00)80799-0 [PubMed]
- 39. Alic N, Tullet JM, Niccoli T, Broughton S, Hoddinott MP, Slack C, Gems D, Partridge L. Cell-nonautonomous effects of dFOXO/DAF-16 in aging. Cell Reports. 2014; 6:608–16. https://doi.org/10.1016/j.celrep.2014.01.015 [PubMed]
- 40. Zhao S, Xu W, Jiang W, Yu W, Lin Y, Zhang T, Yao J, Zhou L, Zeng Y, Li H, Li Y, Shi J, An W, et al. Regulation of cellular metabolism by protein lysine acetylation. Science. 2010; 327:1000–04. https://doi.org/10.1126/science.1179689 [PubMed]
- 41. Peleg S, Feller C, Forne I, Schiller E, Sévin DC, Schauer T, Regnard C, Straub T, Prestel M, Klima C, Schmitt Nogueira M, Becker L, Klopstock T, et al. Life span extension by targeting a link between metabolism and histone acetylation in Drosophila.. EMBO Rep. 2016; 17:455–69. https://doi.org/10.15252/embr.201541132 [PubMed]
- 42. Gut P, Verdin E. The nexus of chromatin regulation and intermediary metabolism. Nature. 2013; 502:489–98. https://doi.org/10.1038/nature12752 [PubMed]
- 43. Woods JK, Kowalski S, Rogina B. Determination of the spontaneous locomotor activity in Drosophila melanogaster.. J Vis Exp. 2014; 86. https://doi.org/10.3791/51449 [PubMed]
- 44. Bross TG, Rogina B, Helfand SL. Behavioral, physical, and demographic changes in Drosophila populations through dietary restriction. Aging Cell. 2005; 4:309–17. https://doi.org/10.1111/j.1474-9726.2005.00181.x [PubMed]
- 45. Wang PY, Neretti N, Whitaker R, Hosier S, Chang C, Lu D, Rogina B, Helfand SL. Long-lived Indy and calorie restriction interact to extend life span. Proc Natl Acad Sci USA. 2009; 106:9262–67. https://doi.org/10.1073/pnas.0904115106 [PubMed]
- 46. Rogers RP, Rogina B. Increased mitochondrial biogenesis preserves intestinal stem cell homeostasis and contributes to longevity in Indy mutant flies. Aging (Albany NY). 2014; 6:335–50. https://doi.org/10.18632/aging.100658 [PubMed]