



Introduction
Inflammatory demyelinating diseases (IDD) include a large group of nervous system disorders, such as multiple sclerosis (MS), neuromyelitis optica spectrum disorders (NMOSD) and acute disseminated encephalomyelitis (ADEM) [1], which have different clinical characteristics in terms of onset age, MRI and cerebrospinal fluid (CSF) features, clinical course and morbidity [1]. MS is considered the prototype [2] of IDD and affects over 2 million people worldwide [3]. The main manifestations of MS are transient and recurrent bouts of handicap [2], affecting mostly young adults with a strong sex bias of female to male about 4:1 [4], but the molecular mechanisms of IDD are essentially unclear.
Complex interactions between genetic and environmental factors play key roles in the inflammatory process of the diseases [2] or influence the susceptibility to these diseases [5]. Many lines of clinical and experimental evidence have shown the involvement of the immune system in IDD, especially MS [6]. So far, genetic linkage analyses and genome-wide association studies of MS have revealed that a large number of genes related to immune functions are associated with MS or other IDD [7]. Among the well investigated factors, interleukin-23A (IL-23A) is known to coordinate the activity of the immune cells and play a key role in the pathogenesis of immune inflammation diseases [8]. By supporting the Th17 cells, IL-23A is involved in chronic or autoimmune inflammations [9]. Additionally, IL-23A also has an important role in mediating some other autoimmune inflammation diseases, such as those in the brain and colon [10,11]. When combined with the heterodimeric receptor, which consists of interleukin-23A receptor (IL-23R) and interleukin 12 receptor beta 1 (IL-12RB1), IL-23A performs its functions [12] and cells responding to the IL-23A signal are mainly determined by their expression of IL-23R [12]. Several variants in the IL-23R gene have been reported for their associations with human autoimmune disorders, such as psoriasis and inflammatory bowel disease [13,14]. IDD, deemed a kind of autoimmune diseases, may involve IL23A in the pathogenesis.
In the present study, we analyzed the transcribed regions and splicing sites of the genes coding for IL-23A and its receptors IL-23R and IL-12RB1 and made comparisons between 206 Chinese Han IDD diseases (including 84 MS) and 300 controls. We also compared the serum levels of IL-23A in different genotype groups of the patients. Our results indicated that three variants rs2066808, rs2371494 and rs11575248 in the IL-23A gene and one variant rs1884444 in the IL-23R gene were associated with the risk of MS or other IDD diseases. Of considerable significance, the serum level of IL-23A in the MS patients was altered by the variants, strongly supporting the involvement of IL-23A in these diseases suggested by genomic analyses.
Results
Clinical data
Two specialists in neurology at the Second Affiliated Hospital of Harbin Medical University, Harbin, China, conducted the diagnosis. There were no other systemic abnormalities in these IDD patients and they had no previous familial history of these diseases. The IDD patients (n = 206, male 60, female 146, the min and max ages were 15 and 73 respectively, and the average age was 41.45 years) included 84 MS, 14 myelitis, 8 NMO, 4 optic neyritis and 96 radiologically isolated syndrome (RIS) diseases that were not classified as MS, myelitis, NMO or optic neyritis. The recruited normal controls (n = 300, male 87, female 213, the min and max ages were 23 and 50 respectively, and the average age was 41.70 years) had no statistical differences in the gender composition or age with the IDD patients (Table 1).
Table 1. Clinical characteristics of study population.
| Parameter | IDD | Control | F | t | P | 95%CI | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Up | Low | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Sample (n) | 206 | 300 | - | - | - | - | - | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Male/Female (n) | 60/146 | 87/213 | - | - | 0.975 | - | - | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Age (years) | 41.45±13.30 | 41.70±7.06 | 115.938 | -0.278 | 0.781 | -2.04458 | 1.53778 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Data are shown as mean±SD; between the two groups, there were no statistical differences of the age and gender composition. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
SNP analyses
We sequenced the transcribed regions and splicing sites of the IL-23A, IL-23R and IL-12RB1 genes to test the hypothesis that common genetic variants in those genes may confer the susceptibility to IDD diseases. We found variants within or near genes of IL-23A (rs2066808, rs2371494, rs11575248 and rs11171806), IL-23R (rs1884444, rs7530511, rs10889677 and rs76418789), and IL-12RB1 (rs11575934 and rs401502). We compared genetic heterozygosity of the SNPs and found that the genetic heterozygosity of rs2066808, rs2371494, rs11575248, rs1884444, rs7530511, rs10889677, rs401502 and rs11575934 was very high (Fig. S1), whereas that of rs11171806 and rs76418789 was very low.
Polymorphism-disease association analyses
In order to test any possible associations between IL-23A, IL-23R or IL-12RB1 and IDD diseases, we conducted analyses on the SNPs and found that the variants rs2066808, rs2371494, rs11575248 in the IL-23A gene and rs1884444 in the IL-23R gene were associated with the risk of IDD diseases in the Chinese Han population (Tables 2, 3); however, the variants rs7530511, rs10889677 in the IL-23R gene and rs401502, rs11575934 in the IL-12RB1 gene were not. Of special significance, the variants rs2066808, rs2371494, rs11575248 and rs1884444 were closely associated with the risk of MS but not RIS (Tables 2, 3).
Table 2. The genotype and allele frequency of rs2066808, rs2371494, rs11575248 and rs1884444 variants in 206 Chinese Han Neuroinflammatory demyelinating diseases of the central nervous system patients and 300 non- IDD controls.
| Genes | Variants | Group | Genotype frequency (%) | Allele frequency (%) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IL23A | Rs2066808 | Genotype | T/T | T/C | C/C | T | C | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IDD | 206 | 183(88.8) | 23(11.2) | 0(0.0) | 389(94.4) | 23(5.6) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MS | 84 | 72(85.7) | 12(14.3) | 0(0.0) | 156(92.9) | 12(7.1) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| RIS | 96 | 87(90.6) | 9(9.4) | 0(0.0) | 183(95.3) | 9(4.7) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Controls | 300 | 287(95.7) | 11(3.7) | 2(0.7) | 585(97.5) | 15(2.5) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs2371494 | Genotype | C/C | C/A | A/A | C | A | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IDD | 206 | 180(87.4) | 26(12.6) | 0(0.0) | 386(93.7) | 26(6.3) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MS | 84 | 72(85.7) | 12(14.3) | 0(0.0) | 156(92.9) | 12(7.1) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| RIS | 96 | 85(88.5) | 11(11.5) | 0(0.0) | 181(94.3) | 11(5.7) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Controls | 300 | 283(94.3) | 16(5.3) | 1(0.3) | 582(97.0) | 18(3.0) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs11575248 | Genotype | C/C | C/A | A/A | C | A | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IDD | 206 | 182(88.3) | 24(11.7) | 0(0.0) | 388(94.2) | 24(5.8) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MS | 84 | 71(84.5) | 13(15.5) | 0(0.0) | 155(92.3) | 13(7.7) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| RIS | 96 | 86(89.6) | 10(10.4) | 0(0.0) | 182(94.8) | 10(5.2) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Controls | 300 | 283(94.3) | 17(5.7) | 0(0.0) | 583(97.2) | 17(2.8) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IL23R | Rs1884444 | Genotype | T/T | T/G | G/G | T | G | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IDD | 206 | 76(11.0) | 108(74.0) | 22(15.1) | 260(63.1) | 152(36.9) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MS | 84 | 35(41.7) | 42(50.0) | 7(8.3) | 112(66.7) | 56(33.3) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| RIS | 96 | 33(34.4) | 52(54.2) | 11(11.5) | 118(61.5) | 74(38.5) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Controls | 300 | 101(33.7) | 133(44.3) | 66(22.0) | 335(55.8) | 265(44.2) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IDD: inflammatory demyelinating diseases; MS: multiple sclerosis; RIS: radiologically isolated syndrome. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Table 3. Associations of rs2066808, rs2371494, rs11575248 and rs1884444 variants within IL23A or IL23R with risk of IDD in Chinese populations.
| Titles | Pearson Chi-square | Pearson’s R | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Genes | Genotyped SNP | Diseases | Statistical Types | Value | Min counta | df | Asymp. Sig. (2-sided) | Value | Asymp. Std. errorb | Approx. Tc | Approx. Sig | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IL23A | Rs2066808 | IDD | Genotype | 12.207 | 0.81 | 2 | 0.002 | -0.109 | 0.047 | -2.459 | 0.014d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 6.422 | 15.47 | 1 | 0.011 | -0.080 | 0.032 | -2.540 | 0.011d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MS | Genotype | 13.610 | 0.44 | 2 | 0.001 | -0.139 | 0.062 | -2.752 | 0.006d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 8.341 | 5.91 | 1 | 0.004 | -0.104 | 0.044 | -2.900 | 0.004d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| RIS | Genotype | 5.528 | 0.48 | 2 | 0.063 | -0.072 | 0.056 | -1.441 | 0.150d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 2.369 | 5.82 | 1 | 0.124 | -0.055 | 0.040 | -1.539 | 0.124d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs2371494 | IDD | Genotype | 9.148 | 0.41 | 2 | 0.010 | -0.113 | 0.046 | -2.546 | 0.011d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 6.438 | 17.91 | 1 | 0.011 | -0.080 | 0.032 | -2.543 | 0.011d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MS | Genotype | 8.020 | 0.22 | 2 | 0.018 | -0.123 | 0.060 | -2.428 | 0.016d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 6.001 | 6.56 | 1 | 0.014 | -0.088 | 0.043 | -2.456 | 0.014d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| RIS | Genotype | 4.584 | 0.24 | 2 | 0.101 | -0.087 | 0.056 | -1.726 | 0.085d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 3.071 | 7.03 | 1 | 0.080 | -0.062 | 0.040 | -1.754 | 0.080d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs11575248 | IDD | Genotype | 5.873 | 16.69 | 1 | 0.015 | -0.108 | 0.045 | -2.433 | 0.015d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 5.625 | 16.69 | 1 | 0.018 | -0.075 | 0.032 | -2.376 | 0.018d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MS | Genotype | 8.768 | 6.56 | 1 | 0.003 | -0.151 | 0.061 | -2.988 | 0.003d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 8.412 | 6.56 | 1 | 0.004 | -0.105 | 0.044 | -2.912 | 0.004d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| RIS | Genotype | 2.583 | 6.55 | 1 | 0.108 | -0.081 | 0.056 | -1.608 | 0.109d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 2.492 | 6.55 | 1 | 0.114 | -0.056 | 0.040 | -1.579 | 0.115d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IL23R | Rs1884444 | IDD | Genotype | 11.043 | 35.83 | 2 | 0.004 | -0.102 | 0.043 | -2.298 | 0.022d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 5.334 | 169.77 | 1 | 0.021 | 0.073 | 0.031 | 2.313 | 0.021d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MS | Genotype | 8.096 | 15.97 | 2 | 0.017 | 0.125 | 0.046 | 2.453 | 0.015d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 6.332 | 70.22 | 1 | 0.012 | 0.091 | 0.035 | 2.523 | 0.012d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| RIS | Genotype | 5.672 | 18.67 | 2 | 0.059 | 0.067 | 0.046 | 1.340 | 0.181d | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Allele | 1.880 | 82.18 | 1 | 0.170 | 0.049 | 0.035 | 1.371 | 0.171d | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| a the minimum expected count; b not assuming the null hypothesis; c using the asymptotic standard error assuming the null hypothesis; d based on normal approximation; IDD: inflammatory demyelinating diseases; MS: multiple sclerosis; RIS: radiologically isolated syndrome. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The genotype frequencies in the IDD diseases and control groups were further analyzed by three genetic models: additive, dominant, and recessive. The variants rs2066808, rs2371494, rs11575248 and rs1884444 were associated with the risk of IDD diseases, however rs2066808 and rs2371494 were not in the recessive model and rs1884444 was not in the dominant model (Table 4). The experiment-wide significance threshold of the variants rs2066808, rs2371494, rs11575248 is 0.019. The Haploview software was used to conduct LD analysis in the IL23A variants, and the results from the LD analysis of the variants (rs2066808, rs2371494 and rs11575248) in the present study and the data from the HapMap CHB population were shown in Fig. 1. The data from the HapMap CHB and this work were consistent. We conducted the Hardy-Weinberg equilibrium test for the IDD diseases and controls and it was in line with equilibrium.
Table 4. Analysis of rs2066808, rs2371494, rs11575248 and rs1884444 variants in the IDD and control groups based on three genetic models.
| Genes | Variants | Additive model (P value) | Dominant model (P value) | Recessive model (P value) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IL23A | Rs2066808 | 0.01731 | 0.004455 | 0.5163 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs2371494 | 0.01761 | 0.008755 | 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs11575248 | 0.01537 | 0.02242 | 0.0197 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IL23R | Rs1884444 | 0.02291 | 0.5067 | 0.001176 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IDD: inflammatory demyelinating diseases | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
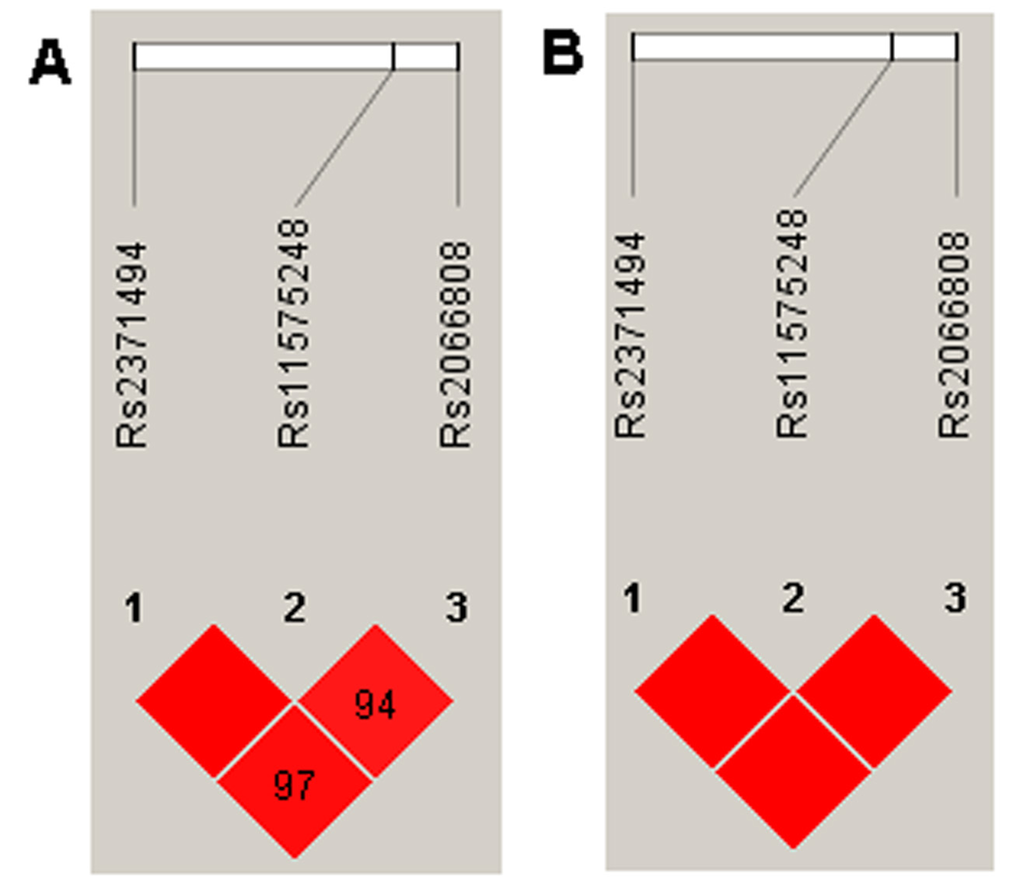
Figure 1. LD analysis of the variants in the IL23A gene region, and the LD plots were generated using the Haploview software v4.2. (A) Data analysis between IDD patients and controls from the present study. (B) Data from the HapMap CHB. The data from the HapMap CHB and this work were very similar.
Gene expression analysis
We conducted the ELISA analysis to measure the serum level of IL23A in the IDD participants and controls between the wild type, heterozygous mutation and homozygous mutation type groups. Firstly, we drew a standard curve and calculated the regression equation (Y = 0.0385X + 0.0393, R2=0.9969; Fig. S2). We found that the serum levels of IL23A in the MS patients were higher than those in the normal group (statistically significant, two asterisks). Notably, when both the IL-23A and IL-23R genes were altered, the serum levels of IL23A were significantly higher than those in the groups in which the IL-23A and IL-23R genes were not altered or only one of them was altered (Fig.2). Remarkably, although the serum levels of IL23A in the RIS patients were higher than those in the normal group (statistically significant), there was no statistically significant difference among the three groups that had the IL-23A and IL-23R genes both altered, only one altered or neither altered (Fig. 2).
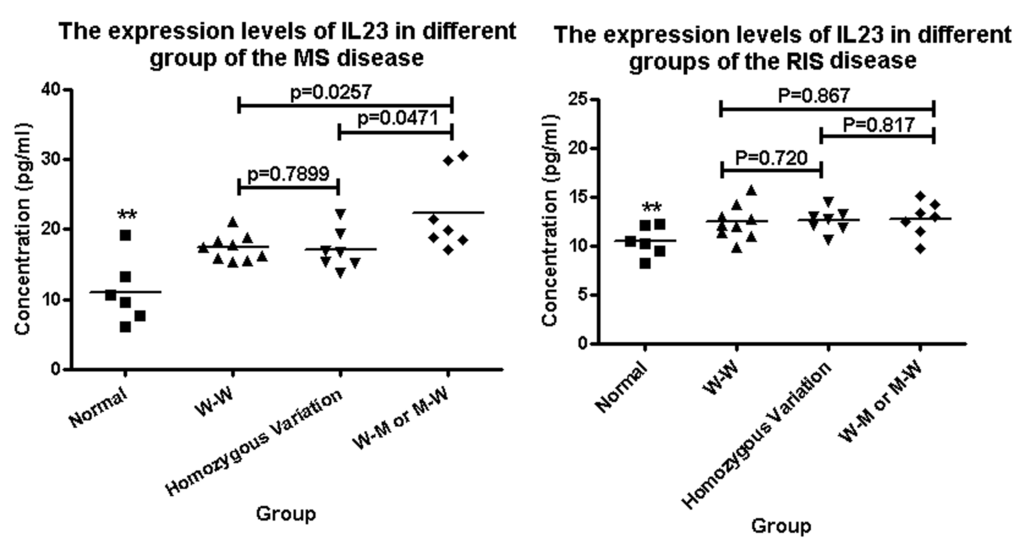
Figure 2. The serum levels of IL23A in IDD and normal groups were detected by ELISA. (A) The expression levels of IL23A in the MS patients were higher than that in the control group (two asterisk); and when both the IL-23A and IL-23R genes were altered, the serum levels of IL23A were higher than those in the groups in which neither the IL-23A nor the IL-23R gene was altered or only one of them was altered. (B) The expression levels of IL23A in the RIS patients were higher than those in the control group (two asterisks); however there was no difference between the three groups that had the IL-23A and IL-23R genes both altered, only one altered or neither altered. “WW” means neither the IL-23A nor the IL-23R gene was altered; “MM” means the IL-23A and IL-23R genes were both altered; “Homozygous variant” means the homozygous variant G/G of the rs1884444 variant in the IL-23R gene.
Discussion
In this study, we investigated possible associations of variants in the IL-23A and IL-23R genes with the risk of inflammatory demyelinating diseases (IDD). We found that three variants (rs2066808, rs2371494, rs11575248) in the IL-23A gene and one variant (rs1884444) in the IL-23R gene were associated with the risk of IDD, especially MS; the serum levels of IL23A in the MS patients were altered by the co-effects of the variants.
IDDs, especially MS, are deemed to be a kind of autoimmune diseases that develop in individuals with genetic predisposition [15]. The cytokine IL23A has important effects in the differentiation of pro- inflammatory cells, and these cells have been considered to be the main factors in the MS pathogenesis [16]. It has also been reported that more than 90% of MS patients have humoral responses [17], and the serum level of IL-23A may reflect autoimmune disease progression and types [18]. Additionally, oligoclonal IgG bands in the cerebrospinal fluid of MS patients are strong predictors for disease course progression [19], and oligoclonal IgM bands in the cerebrospinal fluid of MS patients also are correlated with worse prognosis of the disease [20]. The increased humoral responses also are correlated with MS progression [21], and the subunits of neuro-filament protein may be potential serum and cerebrospinal fluid biomarkers for MS disease progression [22]. In the present work, we found that the serum levels of IL-23A in the MS patients were higher than those in the normal groups. More interestingly, in the group in which both the IL-23A and IL-23R genes were altered, the serum levels of IL23A were higher than those in the groups in which neither the IL-23A nor IL-23R genes was altered or only one of them was altered.
Although we found that the serum levels of IL-23A in the MS patients were affected by the variants in IL-23A and IL-23R, these changes were not found in the RIS patients, strongly suggesting differences between MS and RIS in pathogenesis in addition to their differences in clinical characteristics such as onset age, MRI and cerebrospinal fluid (CSF) features, clinical course and morbidity [1] [23] [24]. The reasons underlying those differences between MS and other demyelinating syndromes may be in the balance status of effectors and regulatory immune cells, immune activation, and age-related immune cell access into the central nervous system [25].
As functional genetic variants within the IL-23A gene have significant impacts on the host immune response, they could be excellent candidate targets for genetic association studies [26]. The variant rs11171806 located within exon3 of the IL-23A gene is significantly associated with the susceptibility to the autoimmune disorder Graves’ disease [18]. Although the genetic heterozygosity of the rs11171806 characterized in this study was too low to be useful for susceptibility analysis to MS or other IDD disease, three variants (rs2066808, rs2371494, rs11575248) within the 5-‘UTR or near the 3-‘UTR of the IL-23A gene were significantly associated with the risk of MS or other IDD, providing opportunities for etiological and pathogenesis studies.
In addition to the IL-23A gene, some genes related to the IL-23A functions also have been investigated for their possible associations with the risk of autoimmune diseases [27,28], such as the variant rs10889677 in the IL-23R gene associated with MS [29] (consensus not reached yet [30]). In this work, we did not find statistical significance between rs10889677 and MS or other IDD, but another variant rs1884444 within exon1 of the IL-23R gene was associated with the risk of MS or other IDD. Of great importance, the variant rs1884444 was associated with changes of the serum levels of IL-23A in the MS patients.
In conclusion, we found that three variants (rs2066808, rs2371494, rs11575248) in the IL-23A gene and one variant (rs1884444) in the IL-23R gene were associated with the risk of MS and other IDD, and the serum levels of IL-23A in the MS patients were also altered by the variants, providing new evidence for the importance of IL-23A and IL-23R in the pathogenesis of the MS disease and demonstrating the differences between MS and other demyelinating syndromes in pathogenesis.
Materials and Methods
Study population
We included 206 IDD patients and 300 normal controls (Table 1), which were assembled at the Department of Neurology and Medical Examination Center of the Second Affiliated Hospital of Harbin Medical University, Harbin, China. In order to make the present work consistent with the 1975 Declaration of Helsinki, we obtained a written informed consent from each participant or their guardian, and this work also was approved by the Ethics Committee of Harbin Medical University. We also confirm that all experiments were performed in accordance with relevant guidelines and regulations [31]. Medical histories of the enrolled participants were recorded in detail, and the participants also received physical and neurological system examination. When necessary, some patients also received imageological examination or cerebrospinal fluid inspection. All the patients were diagnosed according to the McDonald's (2010) diagnostic criteria [32].
DNA analysis
Genomic DNA in peripheral blood leukocytes of each participant was extracted using standard protocols [33]. The transcribed regions and splicing sites of the IL-23A, IL-23R and IL-12RB1 genes were amplified by PCR with the primers shown in Table S1. The PCR products were sequenced for mutational analysis using standard protocols [34].
SNP genotyping analysis and statistical analysis
The variants within or near the genes of IL-23A (rs1884444, rs7530511, rs10889677 and rs76418789), IL-23R (rs2066808, rs2371494, rs11575248, rs11171806 and rs401502), and IL-12RB1 (rs11575934) were determined (Fig. 3) on 206 patients with IDD diseases and 300 normal controls. The DNA regions were amplified and the PCR products sequenced to determine the genotypes. And then overall IDD analysis was conducted according to the types of IDD and sample sizes.
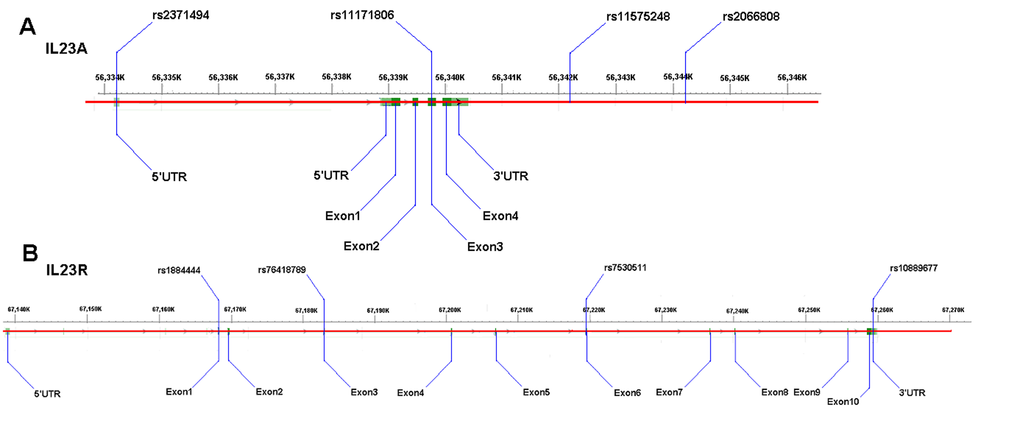
Figure 3. Schematic diagrams of the variants. (A) the IL-23A gene variants rs2066808, rs2371494 and rs11575248; (B): the IL-23R gene variant rs1884444.
Using the SPSS software (version 19.0), we conducted the statistical analyses [35]. The continuous variables (measurement data, such as age) statistical analyses were conducted using independent-samples T test and the discrete variables (enumeration data, such as gender composition and genotype frequency) statistical analyses were conducted using Chi-Square Tests to calculate P value. P values smaller than 0.05 were considered statistically significant [36]. The Hardy-Weinberg equilibrium tests of the IDD and control populations were conducted with the online software OEGE [37]. The Fisher test was used to compare the genotype frequencies between the IDD and control groups based on three genetic models using PLINK v1.07 software (http://pngu.mgh.harvard.edu/Bpurcell/plink/). We also determined experiment-wide significance threshold, matrix of mpirwise linkage-disequilibrium (LD) correlation for the markers and haplotype diagram of LD structure using SNPSpD software(http://neurogenetics.qimrberghofer.edu.au/SNPSpD/) [38] and Haploview software(http://www.broadinstitute.org/scientific-community/science/programs/medical-and-population-genetics/haploview/haploview) [39].
ELISA analysis and data calculation
We separated the serum and blood cells and stored the samples at -80°C prior to use. For ELISA assays, we used the Interleukins 23A (Human) ELISA testing kit (96T), No.: CK - E10077H, from Shanghai Yuanye Bio-Technology Co., Ltd. The IL23A levels were measured by ELISA using a microplate reader (BioTeK Epoch, USA).
To make a standard curve for measuring the levels of IL–23A, we put the OD values of six different concentrations of standard substance minus the OD value of blank hole on ordinate Y and the concentrations of the standard substance on the abscissa X, and then calculated the regression equation (Y = aX + b, R2 value). The concentrations of IL–23A in the samples were then determined according to their OD values. Finally, using GraphPad Prism, we conducted the T test.
Supplementary Materials
Acknowledgements
The authors thank the patients and the family members for their cooperation and participation in this study.
Funding
This work was supported by grants of National Natural Science Foundation of China to SLL (NSFC81271786, 81030029, 81671980) and JF (NSFC 81070964, 81171120), and a grant from Health and Family Planning Commission of Heilongjiang province Foundation (2016-189) to FFL. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflicts of Interest
All the authors have declared that no competing financial interests exist.
References
- 1. Berger T, Reindl M. Antibody biomarkers in CNS demyelinating diseases - a long and winding road. Eur J Neurol. 2015; 22:1162–68. https://doi.org/10.1111/ene.12759 [PubMed]
- 2. Legroux L, Arbour N. Multiple Sclerosis and T Lymphocytes: An Entangled Story. J Neuroimmune Pharmacol. 2015; 10:528–46. https://doi.org/10.1007/s11481-015-9614-0 [PubMed]
- 3. Browne P, Chandraratna D, Angood C, Tremlett H, Baker C, Taylor BV, Thompson AJ. Atlas of Multiple Sclerosis 2013: A growing global problem with widespread inequity. Neurology. 2014; 83:1022–24. https://doi.org/10.1212/WNL.0000000000000768 [PubMed]
- 4. Harbo HF, Gold R, Tintoré M. Sex and gender issues in multiple sclerosis. Ther Adv Neurol Disorder. 2013; 6:237–48. https://doi.org/10.1177/1756285613488434 [PubMed]
- 5. Koch MW, Metz LM, Agrawal SM, Yong VW. Environmental factors and their regulation of immunity in multiple sclerosis. J Neurol Sci. 2013; 324:10–16. https://doi.org/10.1016/j.jns.2012.10.021 [PubMed]
- 6. Cross AH, Naismith RT. Established and novel disease-modifying treatments in multiple sclerosis. J Intern Med. 2014; 275:350–63. https://doi.org/10.1111/joim.12203 [PubMed]
- 7. Andreassen OA, Harbo HF, Wang Y, Thompson WK, Schork AJ, Mattingsdal M, Zuber V, Bettella F, Ripke S, Kelsoe JR, Kendler KS, O’Donovan MC, Sklar P, et al, and Psychiatric Genomics Consortium (PGC) Bipolar Disorder and Schizophrenia Work Groups, and International Multiple Sclerosis Genetics Consortium (IMSGC). Genetic pleiotropy between multiple sclerosis and schizophrenia but not bipolar disorder: differential involvement of immune-related gene loci. Mol Psychiatry. 2015; 20:207–14. https://doi.org/10.1038/mp.2013.195 [PubMed]
- 8. Zhao J, Liu YH, Reichert P, Pflanz S, Pramanik B. Glycosylation analysis of interleukin-23 receptor: elucidation of glycosylation sites and characterization of attached glycan structures. J Mass Spectrom. 2010; 45:1416–25. https://doi.org/10.1002/jms.1858 [PubMed]
- 9. McGeachy MJ, Cua DJ. The link between IL-23 and Th17 cell-mediated immune pathologies. Semin Immunol. 2007; 19:372–76. https://doi.org/10.1016/j.smim.2007.10.012 [PubMed]
- 10. Chen Y, Langrish CL, McKenzie B, Joyce-Shaikh B, Stumhofer JS, McClanahan T, Blumenschein W, Churakovsa T, Low J, Presta L, Hunter CA, Kastelein RA, Cua DJ. Anti-IL-23 therapy inhibits multiple inflammatory pathways and ameliorates autoimmune encephalomyelitis. J Clin Invest. 2006; 116:1317–26. https://doi.org/10.1172/JCI25308 [PubMed]
- 11. Elson CO, Cong Y, Weaver CT, Schoeb TR, McClanahan TK, Fick RB, Kastelein RA. Monoclonal anti-interleukin 23 reverses active colitis in a T cell-mediated model in mice. Gastroenterology. 2007; 132:2359–70. https://doi.org/10.1053/j.gastro.2007.03.104 [PubMed]
- 12. Parham C, Chirica M, Timans J, Vaisberg E, Travis M, Cheung J, Pflanz S, Zhang R, Singh KP, Vega F, To W, Wagner J, O’Farrell AM, et al. A receptor for the heterodimeric cytokine IL-23 is composed of IL-12Rbeta1 and a novel cytokine receptor subunit, IL-23R. J Immunol. 2002; 168:5699–708. https://doi.org/10.4049/jimmunol.168.11.5699 [PubMed]
- 13. Capon F, Di Meglio P, Szaub J, Prescott NJ, Dunster C, Baumber L, Timms K, Gutin A, Abkevic V, Burden AD, Lanchbury J, Barker JN, Trembath RC, Nestle FO. Sequence variants in the genes for the interleukin-23 receptor (IL23R) and its ligand (IL12B) confer protection against psoriasis. Hum Genet. 2007; 122:201–06. https://doi.org/10.1007/s00439-007-0397-0 [PubMed]
- 14. Duerr RH, Taylor KD, Brant SR, Rioux JD, Silverberg MS, Daly MJ, Steinhart AH, Abraham C, Regueiro M, Griffiths A, Dassopoulos T, Bitton A, Yang H, et al. A genome-wide association study identifies IL23R as an inflammatory bowel disease gene. Science. 2006; 314:1461–63. https://doi.org/10.1126/science.1135245 [PubMed]
- 15. Garg N, Reddel SW, Miller DH, Chataway J, Riminton DS, Barnett Y, Masters L, Barnett MH, Hardy TA. The corpus callosum in the diagnosis of multiple sclerosis and other CNS demyelinating and inflammatory diseases. J Neurol Neurosurg Psychiatry. 2015; 86:1374–82. [PubMed]
- 16. McFarland HF, Martin R. Multiple sclerosis: a complicated picture of autoimmunity. Nat Immunol. 2007; 8:913–19. https://doi.org/10.1038/ni1507 [PubMed]
- 17. Kabat EA, Glusman M, Knaub V. Quantitative estimation of the albumin and gamma globulin in normal and pathologic cerebrospinal fluid by immunochemical methods. Am J Med. 1948; 4:653–62. https://doi.org/10.1016/S0002-9343(48)90389-1 [PubMed]
- 18. Jia H, Tao F, Liu C, Guo T, Zhu W, Wang S, Cui B, Ning G. Both interleukin-23A polymorphism and serum interlukin-23 expression are associated with Graves’ disease risk. Cell Immunol. 2015; 294:39–43. https://doi.org/10.1016/j.cellimm.2015.01.015 [PubMed]
- 19. Kuhle J, Disanto G, Dobson R, Adiutori R, Bianchi L, Topping J, Bestwick JP, Meier UC, Marta M, Dalla Costa G, Runia T, Evdoshenko E, Lazareva N, et al. Conversion from clinically isolated syndrome to multiple sclerosis: A large multicentre study. Mult Scler. 2015; 21:1013–24. https://doi.org/10.1177/1352458514568827 [PubMed]
- 20. Villar LM, Masjuan J, González-Porqué P, Plaza J, Sádaba MC, Roldán E, Bootello A, Alvarez-Cermeño JC. Intrathecal IgM synthesis predicts the onset of new relapses and a worse disease course in MS. Neurology. 2002; 59:555–59. https://doi.org/10.1212/WNL.59.4.555 [PubMed]
- 21. Olsson JE, Link H, Müller R. Immunoglobulin abnormalities in multiple sclerosis. Relation to clinical parameters: disability, duration and age of onset. J Neurol Sci. 1976; 27:233–45. https://doi.org/10.1016/0022-510X(76)90064-2 [PubMed]
- 22. Teunissen CE, Malekzadeh A, Leurs C, Bridel C, Killestein J. Body fluid biomarkers for multiple sclerosis--the long road to clinical application. Nat Rev Neurol. 2015; 11:585–96. https://doi.org/10.1038/nrneurol.2015.173 [PubMed]
- 23. Lebrun C. The radiologically isolated syndrome. Rev Neurol (Paris). 2015; 171:698–706. https://doi.org/10.1016/j.neurol.2015.05.001 [PubMed]
- 24. Azevedo CJ, Overton E, Khadka S, Buckley J, Liu S, Sampat M, Kantarci O, Lebrun Frenay C, Siva A, Okuda DT, Pelletier D. Early CNS neurodegeneration in radiologically isolated syndrome. Neurol Neuroimmunol Neuroinflamm. 2015; 2:e102. https://doi.org/10.1212/NXI.0000000000000102 [PubMed]
- 25. Brenton JN, Banwell BL. Therapeutic Approach to the Management of Pediatric Demyelinating Disease: Multiple Sclerosis and Acute Disseminated Encephalomyelitis. Neurotherapeutics. 2016; 13:84–95. https://doi.org/10.1007/s13311-015-0396-0 [PubMed]
- 26. Tindall EA, Hayes VM. Comprehensive sequence analysis of the human IL23A gene defines new variation content and high rate of evolutionary conservation. DNA Res. 2010; 17:117–22. https://doi.org/10.1093/dnares/dsq003 [PubMed]
- 27. Huber AK, Jacobson EM, Jazdzewski K, Concepcion ES, Tomer Y. Interleukin (IL)-23 receptor is a major susceptibility gene for Graves’ ophthalmopathy: the IL-23/T-helper 17 axis extends to thyroid autoimmunity. J Clin Endocrinol Metab. 2008; 93:1077–81. https://doi.org/10.1210/jc.2007-2190 [PubMed]
- 28. Yan N, Yu YL, Yang J, Qin Q, Zhu YF, Wang X, Song RH, Zhang JA. Association of interleukin-17A and -17F gene single-nucleotide polymorphisms with autoimmune thyroid diseases. Autoimmunity. 2012; 45:533–39. https://doi.org/10.3109/08916934.2012.702814 [PubMed]
- 29. Illes Z, Safrany E, Peterfalvi A, Magyari L, Farago B, Pozsonyi E, Rozsa C, Komoly S, Melegh B. 3'UTR C2370A allele of the IL-23 receptor gene is associated with relapsing-remitting multiple sclerosis. Neurosci Lett. 2008; 431:36–38. https://doi.org/10.1016/j.neulet.2007.11.015 [PubMed]
- 30. Huang J, Yang Y, Zhou F, Liang Z, Kang M, Kuang Y, Li F. Meta-analysis of the IL23R and IL12B polymorphisms in multiple sclerosis. Int J Neurosci. 2016; 126:205–12. https://doi.org/10.3109/00207454.2015.1007508 [PubMed]
- 31. Tan ZX, Li FF, Qu YY, Liu J, Liu GR, Zhou J, Zhu YL, Liu SL. Identification of a known mutation in Notch 3 in familiar CADASIL in China. PLoS One. 2012; 7:e36590. https://doi.org/10.1371/journal.pone.0036590 [PubMed]
- 32. Polman CH, Reingold SC, Banwell B, Clanet M, Cohen JA, Filippi M, Fujihara K, Havrdova E, Hutchinson M, Kappos L, Lublin FD, Montalban X, O’Connor P, et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Ann Neurol. 2011; 69:292–302. https://doi.org/10.1002/ana.22366 [PubMed]
- 33. Li FF, Wang XD, Zhu MW, Lou ZH, Zhang Q, Zhu CY, Feng HL, Lin ZG, Liu SL. Identification of two novel critical mutations in PCNT gene resulting in microcephalic osteodysplastic primordial dwarfism type II associated with multiple intracranial aneurysms. Metab Brain Dis. 2015; 30:1387–94. https://doi.org/10.1007/s11011-015-9712-y [PubMed]
- 34. Li FF, Liu Z, Yan P, Shao X, Deng X, Sam C, Chen YG, Xu YP, Wang XS, Wang GY, Liu SL. Identification of a novel mutation associated with familial adenomatous polyposis and colorectal cancer. Int J Mol Med. 2015; 36:1049–56. [PubMed]
- 35. Deng X, Zhou J, Li FF, Yan P, Zhao EY, Hao L, Yu KJ, Liu SL. Characterization of nodal/TGF-lefty signaling pathway gene variants for possible roles in congenital heart diseases. PLoS One. 2014; 9:e104535. https://doi.org/10.1371/journal.pone.0104535 [PubMed]
- 36. Li FF, Zhou J, Zhao DD, Yan P, Li X, Han Y, Li XS, Wang GY, Yu KJ, Liu SL. Characterization of SMAD3 Gene Variants for Possible Roles in Ventricular Septal Defects and Other Congenital Heart Diseases. PLoS One. 2015; 10:e0131542. https://doi.org/10.1371/journal.pone.0131542 [PubMed]
- 37. Rodriguez S, Gaunt TR, Day IN. Hardy-Weinberg equilibrium testing of biological ascertainment for Mendelian randomization studies. Am J Epidemiol. 2009; 169:505–14. https://doi.org/10.1093/aje/kwn359 [PubMed]
- 38. Nyholt DR. A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. Am J Hum Genet. 2004; 74:765–69. https://doi.org/10.1086/383251 [PubMed]
- 39. Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005; 21:263–65. https://doi.org/10.1093/bioinformatics/bth457 [PubMed]