



Introduction
Wt p53 can induce apoptosis, cell cycle arrest and senescence, which are sufficient to explain tumor suppression by p53 [1]. A recent paper in Cell described that these activities are dispensable for tumor suppression [2]. Mutant p53 (p533KR) that cannot cause arrest, senescence and apoptosis still suppressed tumors in mice [2, 3]. Why do then wt p53 induce apoptosis, cell cycle arrest and senescence? Before entertaining this intriguing question, I will focus on suppression of senescence (gerosuppression) by p53, overlapping with its anti-hypertrophic, anti-hypermetabolic, anti-inflammatory and anti-secretory effects.
P53 suppresses the conversion from arrest to senescence (geroconversion)
How can p53 suppress senescence, if it also can cause senescence? As recently suggested, induction of senescence is not an independent activity of p53 but a consequence of cell-cycle arrest [4-8]. This predicts that any mutant p53 that cannot cause arrest will not cause senescence too. In agreement, p533KR did not cause senescence [2]. This is not trivial. To create p533KR, wt p53 was altered to abolish apoptosis and cell-cycle arrest only [2]. Li et al did not modify p53 to abolish senescence as an independent activity. It was not needed, simply because p53 does not induce senescence as an independent effect. (Note: Seemingly in contrast, it was reported that mutant p53, which cannot induce arrest in response to DNA damage, can cause senescence [9]. Although this mutant p53 did not cause instant arrest, it still arrested proliferation later and then senescence developed [9]. So there is no exception). p53 cannot induce senescence without inducing arrest. But p53 can induce quiescence, a reversible condition characterized by low protein synthesis and metabolism (see detailed definitions in ref. [7, 8]). It was assumed that when p53 causes quiescence, it simply fails to induce senescence. But another possibility is that in such cases p53 suppresses the conversion from cell-cycle arrest to senescence (geroconversion). How can that be tested? In some cell lines, induction of ectopic p21 causes irreversible senescence, whereas induction of p53 causes quiescence [4]. Does p53 suppresses a senescent program? This question can be answered by simultaneously inducing both p53 and ectopic p21. When both p21 and p53 were induced, then cells become quiescent not senescent [4]. p53 was dominant, actively suppressing senescence caused by p21… or by something else? In fact, p21 merely causes cell cycle arrest and does not inhibit mitogen-activated, nutrient-sensing and growth-promoting pathways such as Target of Rapamycin (mTOR) [4]. During several days, these pathways (gerogenic pathways, for brevity) convert p21-induced arrest into senescence. Rapamycin can decelerate geroconversion [10-13]. Also, p53 can inhibit the mTOR pathway [4-6, 14-17]. In some conditions, p53 can suppress senescence during arrest [4-6]. Wt p53 induces arrest and then if it fails to suppress senescence, then senescence prevails. Rather than p53, gerogenic pathways drive senescence during cell-cycle arrest [18].
In summary, wt p53 seems to have three independent effects: apoptosis, cell-cycle arrest and gerosuppression. By inducing arrest, wt p53 primes cells for senescence, unless p53 is able or “willing” to suppress geroconversion. At high levels, gerosuppression by p53 is limited by apoptosis [6]. This predicts that p533KR would potently suppress senescence because gero-suppression by p533KR will not be limited by apoptosis.
Hyper-metabolic senescent phenotype
Senescent cells are hyper-functional: hypertrophic, hypermetabolic, hyper-secretory and hyper-inflammatory [8]. Also, senescent cells may accumulate lipids, becoming not only large but also “fat” (Figure1). Induction of p53 decreased both cellular hypertrophy and fat accumulation (Figure 1). This is in line with numerous metabolic effects of p53 including inhibition of glycolysis and stimulation of fatty acids oxidation [19-32]. Importantly, p533KR retained the ability to inhibit glycolysis and reactive oxygen species (ROS) [2]. (Noteworthy, ROS and mTOR co-activate each other [33] and N-Acetyl Cysteine (NAC), which decreases ROS, also inhibits mTOR [34]). Also, p53 decreases hyper-secretory phenotype also known as SASP [35] and suppresses a pro-inflammatory phenotype [36, 37]. How might gerosuppression contribute to tumor suppression? There are several overlapping explanations, from different points of view of the same process.
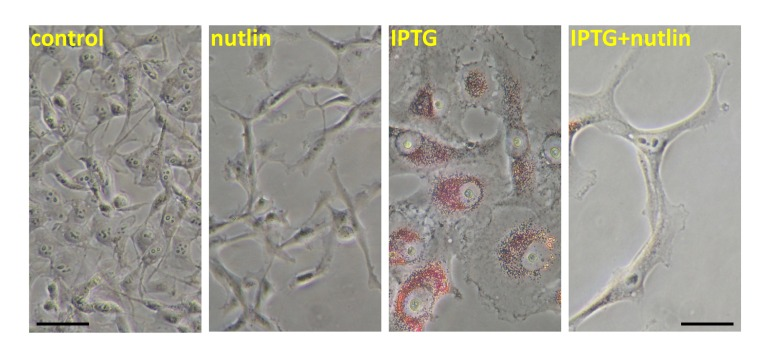
Figure 1. Nutlin-3a decreased lipid accumulation during IPTG-induced senescence HT-p21 cells were treated with IPTG, nutlin-3a and IPTG+nutlin-3a (as indicated) for 3 days as described previously [4-6] and cells were stained with “oil red O” for lipids. In HT-p21 cells, IPTG induces ectopic p21 and senescence. As described previously, nutlin-3a induces endogenous p53 and suppresses IPTG-induced senescence [4-6].
Gerogenic conversion and oncogenic transformation
In proliferating epithelial cells, pro-gerogenic conversion may contribute to carcinogenesis directly. The PI3K/mTOR pathway is universally activated in cancer [38-49]. p53 can inhibit the PI3K/mTOR pathway [4-6, 14-17, 50]. Like p53, many other tumor suppressors such as PTEN, AMPK, TSC2, LKB1, NF1 inhibit the PI3K/mTOR pathway [51].
Geroconversion of stromal cells creates carcinogenic microenvironment
First, senescence creates a selective disadvantage for normal cells, thus selecting for cancer [52-54]. Also, senescent stromal cells secrete factors that favors pre-cancer and cancer growth [37, 54-62]. Third, the senescent stroma is hyper-metabolic and thus promotes cancer by fueling cancer growth [59, 60, 63-71]. In a model of accelerated host aging, mTOR activity was increased in normal tissues [72]. This pro-senescent microenvironment accelerated growth of implanted tumors. The tumor-promoting effects of pro-senescent microenvironment were abrogated by rapamycin [72].
Is aging accelerated in p53-deficient mice?
Inactivation of tumor suppressors accelerates both aging and cancer [87]. It was thought that p53 is an exception. Yet, given that p53 can suppress geroconversion, it may not be the exception after all. A complex role of p53 in cellular senescence and organismal aging was discussed [88-91]. Mice with increased, but normally regulated, p53 lives longer [92]. p53 knockout mice have both accelerated carcinogenesis and decreased longevity [93-98]. p53−/− mice have a pro-inflammatory phenotype characteristic of accelerated aging [36, 37]. Also, atherosclerosis is accelerated in p53−/− animals [99-102]. While loss of p53 by itself makes cells prone to become tumorigenic, an increased rate of organismal aging in the absence of p53 may further accelerate carcinogenesis.
Rapalogs and p53
Rapamycin (sirolimus) and other rapalogs (everolimus and temsirolimus) are pharmacological tumor suppressors. Noteworthy, like p53, rapamycin decreases glycolysis [103] and lactate production [34] and stimulates oxidation of fatty acids [104, 105]. Furthermore, rapamycin slows cellular proliferation, and so, not surprisingly, p533KR inhibits clonogenicity too [2]. Yet, p53 affects metabolism and aging not only via mTOR but also via direct transactivation of metabolic enzymes, rendering it a more potent tumor suppressor.
Puzzles remain
Still, even if gerosuppression and anti-hypermetabolic effects can in part explain tumor suppression, puzzles remain. Why does wt p53 cause “unneeded” apoptosis and “instant” (p21-dependent) arrest? Why is p53 needed at all? In the wild, most mice die from external/accidental causes and only a few would live long enough to die from cancer, regardless of p53 status. In the wild, starvation (natural calorie restriction) would delay cancer further. Yet, p53 is also needed very early in life, or technically speaking, even before life has begun, because p53 plays role in fertility and reproduction [106-113]. And is tumor suppression a late life function?
Alternatively, tumor suppression is a primary function of p53. And each of the three activities (apoptosis, arrest, gerosuppression) is partially sufficient for cancer prevention. In their combination, these activities are the most effective tumor suppressor. And each activity may be partially dispensable in some mice strains and in some conditions. For example, the gerosuppressive activity of p53 may be preferentially important in peculiar strains of laboratory mice, or mice fed ad libitum, which constantly activates mTOR and accelerates aging. In fact, calorie restriction, which deactivates mTOR and decelerates aging, partially substitutes for the loss of p53 in mice.
Acknowledgments
I thank Wei Gu, Arnold Levine and Bert Vogelstein for critical reading of the manuscript and excellent suggestions.
Conflicts of Interest
The author of this manuscript has no conflict of interest to declare.
References
- 1. Vogelstein B, Lane DP, Levine AJ. Surfing the p53 network. Nature. 2000; 408: 307 -310. [PubMed] .
- 2. Li T, Kon N, Jiang L, Tan M, Ludwig T, Zhao Y, Baer R, Gu W. Tumor Suppression in the Absence of p53-Mediated Cell-Cycle Arrest, Apoptosis, and Senescence. Cell. 2012; 149: 1269 -1283. [PubMed] .
- 3. Hock AK and Vousden KH. Tumor Suppression by p53: Fall of the Triumvirate? Cell. 2012; 149: 1183 -1185. [PubMed] .
- 4. Demidenko ZN, Korotchkina LG, Gudkov AV, Blagosklonny MV. Paradoxical suppression of cellular senescence by p53. Proc Natl Acad Sci U S A. 2010; 107: 9660 -9664. [PubMed] .
- 5. Korotchkina LG, Leontieva OV, Bukreeva EI, Demidenko ZN, Gudkov AV, Blagosklonny MV. The choice between p53-induced senescence and quiescence is determined in part by the mTOR pathway. Aging. 2010; 2: 344 -352. [PubMed] .
- 6. Leontieva O, Gudkov A, Blagosklonny M. Weak p53 permits senescence during cell cycle arrest. Cell Cycle. 2010; 9: 4323 -4327. [PubMed] .
- 7. Blagosklonny MV. Cell cycle arrest is not senescence. Aging. 2011; 3: 94 -101. [PubMed] .
- 8. Blagosklonny MV. Cell cycle arrest is not yet senescence, which is not just cell cycle arrest: terminology for TOR-driven aging. Aging. 2012; 4: 159 -165. [PubMed] .
- 9. Brady CA, Jiang D, Mello SS, Johnson TM, Jarvis LA, Kozak MM, Kenzelmann Broz D, Basak S, Park EJ, McLaughlin ME, Karnezis AN, Attardi LD. Distinct p53 transcriptional programs dictate acute DNA-damage responses and tumor suppression. Cell. 2011; 145: 571 -583. [PubMed] .
- 10. Demidenko ZN and Blagosklonny MV. Growth stimulation leads to cellular senescence when the cell cycle is blocked. Cell Cycle. 2008; 7: 3355 -3361. [PubMed] .
- 11. Demidenko ZN, Zubova SG, Bukreeva EI, Pospelov VA, Pospelova TV, Blagosklonny MV. Rapamycin decelerates cellular senescence. Cell Cycle. 2009; 8: 1888 -1895. [PubMed] .
- 12. Demidenko ZN and Blagosklonny MV. Quantifying pharmacologic suppression of cellular senescence: prevention of cellular hypertrophy versus preservation of proliferative potential. Aging. 2009; 1: 1008 -1016. [PubMed] .
- 13. Pospelova TV, Demidenko ZN, Bukreeva EI, Pospelov VA, Gudkov AV, Blagosklonny MV. Pseudo-DNA damage response in senescent cells. Cell Cycle. 2009; 8: 4112 -4118. [PubMed] .
- 14. Feng Z and Levine AJ. The regulation of energy metabolism and the IGF-1/mTOR pathways by the p53 protein. Trends Cell Biol. 2010; 20: 427 -434. [PubMed] .
- 15. Feng Z, Hu W, de Stanchina E, Teresky AK, Jin S, Lowe S, Levine AJ. The regulation of AMPK beta1, TSC2, and PTEN expression by p53: stress, cell and tissue specificity, and the role of these gene products in modulating the IGF-1-AKT-mTOR pathways. Cancer Res. 2007; 67: 3043 -3053. [PubMed] .
- 16. Levine AJ, Feng Z, Mak TW, You H, Jin S. Coordination and communication between the p53 and IGF-1-AKT-TOR signal transduction pathways. Genes Dev. 2006; 20: 267 -275. [PubMed] .
- 17. Budanov AV and Karin M. p53 target genes sestrin1 and sestrin2 connect genotoxic stress and mTOR signaling. Cell. 2008; 134: 451 -460. [PubMed] .
- 18. Leontieva OV and Blagosklonny MV. DNA damaging agents and p53 do not cause senescence in quiescent cells, while consecutive re-activation of mTOR is associated with conversion to senescence. Aging. 2010; 2: 924 -935. [PubMed] .
- 19. Bensaad K, Tsuruta A, Selak MA, Vidal MN, Nakano K, Bartrons R, Gottlieb E, Vousden KH. TIGAR, a p53-inducible regulator of glycolysis and apoptosis. Cell. 2006; 126: 107 -120. [PubMed] .
- 20. Bensaad K and Vousden KH. p53: new roles in metabolism. Trends Cell Biol. 2007; 17: 286 -291. [PubMed] .
- 21. Kawauchi K, Araki K, Tobiume K, Tanaka N. p53 regulates glucose metabolism through an IKK-NF-kappaB pathway and inhibits cell transformation. Nat Cell Biol. 2008; 10: 611 -618. [PubMed] .
- 22. Vousden KH and Ryan KM. p53 and metabolism. Nat Rev Cancer. 2009; 9: 691 -700. [PubMed] .
- 23. Vigneron A and Vousden KH. p53, ROS and senescence in the control of aging. Aging. 2010; 2: 471 -474. [PubMed] .
- 24. Cheung EC and Vousden KH. The role of p53 in glucose metabolism. Curr Opin Cell Biol. 2010; 22: 186 -191. [PubMed] .
- 25. Suzuki S, Tanaka T, Poyurovsky MV, Nagano H, Mayama T, Ohkubo S, Lokshin M, Hosokawa H, Nakayama T, Suzuki Y, Sugano S, Sato E, Nagao T, Yokote K, Tatsuno I, Prives C. Phosphate-activated glutaminase (GLS2), a p53-inducible regulator of glutamine metabolism and reactive oxygen species. Proc Natl Acad Sci U S A. 2010; 107: 7461 -7466. [PubMed] .
- 26. Jiang P, Du W, Wang X, Mancuso A, Gao X, Wu M, Yang X. p53 regulates biosynthesis through direct inactivation of glucose-6-phosphate dehydrogenase. Nat Cell Biol. 2011; 13: 310 -316. [PubMed] .
- 27. Zhu Y and Prives C. p53 and Metabolism: The GAMT Connection. Mol Cell. 2009; 36: 351 -352. [PubMed] .
- 28. Bensaad K, Cheung EC, Vousden KH. Modulation of intracellular ROS levels by TIGAR controls autophagy. Embo J. 2009; 28: 3015 -3026. [PubMed] .
- 29. Hu W, Zhang C, Wu R, Sun Y, Levine A, Feng Z. Glutaminase 2, a novel p53 target gene regulating energy metabolism and antioxidant function. Proc Natl Acad Sci U S A. 2010; 107: 7455 -7460. [PubMed] .
- 30. Ide T, Brown-Endres L, Chu K, Ongusaha PP, Ohtsuka T, El-Deiry WS, Aaronson SA, Lee SW. GAMT, a p53-inducible modulator of apoptosis, is critical for the adaptive response to nutrient stress. Mol Cell. 2009; 36: 379 -392. [PubMed] .
- 31. Park JY, Wang PY, Matsumoto T, Sung HJ, Ma W, Choi JW, Anderson SA, Leary SC, Balaban RS, Kang JG, Hwang PM. p53 improves aerobic exercise capacity and augments skeletal muscle mitochondrial DNA content. Circ Res. 2009; 105: 705 -712. [PubMed] .
- 32. Madan E, Gogna R, Bhatt M, Pati U, Kuppusamy P, Mahdi AA. Regulation of glucose metabolism by p53: emerging new roles for the tumor suppressor. Oncotarget. 2011; 2: 948 -957. [PubMed] .
- 33. Blagosklonny MV. Aging: ROS or TOR. Cell Cycle. 2008; 7: 3344 -3354. [PubMed] .
- 34. Leontieva OV and Blagosklonny MV. Yeast-like chronological senescence in mammalian cells: phenomenon, mechanism and pharmacological suppression. Aging. 2011; 3: 1078 -1091. [PubMed] .
- 35. CoppŽ JP, Patil CK, Rodier F, Sun Y, Mu-oz DP, Goldstein J, Nelson PS, Desprez PY, Campisi J. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008; 6: 2853 -2868. [PubMed] .
- 36. Komarova EA, Krivokrysenko V, Wang K, Neznanov N, Chernov MV, Komarov PG, Brennan ML, Golovkina TV, Rokhlin OW, Kuprash DV, Nedospasov SA, Hazen SL, Feinstein E, Gudkov AV. p53 is a suppressor of inflammatory response in mice. Faseb J. 2005; 19: 1030 -1032. [PubMed] .
- 37. Gudkov AV, Gurova KV, Komarova EA. Inflammation and p53: A Tale of Two Stresses. Genes Cancer. 2011; 2: 503 -516. [PubMed] .
- 38. Vogelstein B and Kinzler KW. Cancer genes and the pathways they control. Nat Med. 2004; 10: 789 -799. [PubMed] .
- 39. Shaw RJ and Cantley LC. Ras, PI(3)K and mTOR signalling controls tumour cell growth. Nature. 2006; 441: 424 -430. [PubMed] .
- 40. Janes MR and Fruman DA. Targeting TOR dependence in cancer. Oncotarget. 2010; 1: 69 -76. [PubMed] .
- 41. Guertin DA and Sabatini DM. Defining the role of mTOR in cancer. Cancer Cell. 2007; 12: 9 -22. [PubMed] .
- 42. Schmidt-Kittler O, Zhu J, Yang J, Liu G, Hendricks W, Lengauer C, Gabelli SB, Kinzler KW, Vogelstein B, Huso DL, Zhou S. PI3Kalpha inhibitors that inhibit metastasis. Oncotarget. 2010; 1: 339 -348. [PubMed] .
- 43. Martelli AM, Evangelisti C, Chiarini F, McCubrey JA. The phosphatidylinositol 3-kinase/Akt/mTOR signaling network as a therapeutic target in acute myelogenous leukemia patients. Oncotarget. 2010; 1: 89 -103. [PubMed] .
- 44. Zawel L. P3Kalpha: a driver of tumor metastasis? Oncotarget. 2010; 1: 315 -316. [PubMed] .
- 45. Zhang Z, Stiegler AL, Boggon TJ, Kobayashi S, Halmos B. EGFR-mutated lung cancer: a paradigm of molecular oncology. Oncotarget. 2010; 1: 497 -514. [PubMed] .
- 46. Shahbazian D, Parsyan A, Petroulakis E, Hershey J, Sonenberg N. eIF4B controls survival and proliferation and is regulated by proto-oncogenic signaling pathways. Cell Cycle. 2010; 9: 4106 -4109. [PubMed] .
- 47. Zhao L and Vogt PK. Hot-spot mutations in p110alpha of phosphatidylinositol 3-kinase (pI3K): differential interactions with the regulatory subunit p85 and with RAS. Cell Cycle. 2010; 9: 596 -600. [PubMed] .
- 48. Bhatia B, Nahle Z, Kenney AM. Double trouble: when sonic hedgehog signaling meets TSC inactivation. Cell Cycle. 2010; 9: 456 -459. [PubMed] .
- 49. Fujishita T, Aoki M, Taketo MM. The role of mTORC1 pathway in intestinal tumorigenesis. Cell Cycle. 2009; 8: 3684 -3687. [PubMed] .
- 50. Galluzzi L, Kepp O, Kroemer G. TP53 and MTOR crosstalk to regulate cellular senescence. Aging. 2010; 2: 535 -537. [PubMed] .
- 51. Blagosklonny MV. Molecular damage in cancer: an argument for mTOR-driven aging. Aging. 2011; 3: 1130 -1141. [PubMed] .
- 52. Blagosklonny MV. NCI's provocative questions on cancer: some answers to ignite discussion. Oncotarget. 2011; 2: 1352 -1367. [PubMed] .
- 53. Henry CJ, Marusyk A, Zaberezhnyy V, Adane B, DeGregori J. Declining lymphoid progenitor fitness promotes aging-associated leukemogenesis. Proc Natl Acad Sci U S A. 2010; 107: 21713 -21718. [PubMed] .
- 54. Henry CJ, Marusyk A, DeGregori J. Aging-associated changes in hematopoiesis and leukemogenesis: what's the connection? Aging. 2011; 3: 643 -656. [PubMed] .
- 55. Parrinello S, Coppe JP, Krtolica A, Campisi J. Stromal-epithelial interactions in aging and cancer: senescent fibroblasts alter epithelial cell differentiation. J Cell Sci. 2005; 118: 485 -496. [PubMed] .
- 56. Coppe JP, Patil CK, Rodier F, Sun Y, Munoz DP, Goldstein J, Nelson PS, Desprez PY, Campisi J. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008; 6: 2853 -2868. [PubMed] .
- 57. Davalos AR, Coppe JP, Campisi J, Desprez PY. Senescent cells as a source of inflammatory factors for tumor progression. Cancer Metastasis Rev. 2011; 29: 273 -283. [PubMed] .
- 58. Coussens LM and Werb Z. Inflammation and cancer. Nature. 2002; 420: 860 -867. [PubMed] .
- 59. Lisanti MP, Martinez-Outschoorn UE, Pavlides S, Whitaker-Menezes D, Pestell RG, Howell A, Sotgia F. Accelerated aging in the tumor microenvironment: connecting aging, inflammation and cancer metabolism with personalized medicine. Cell Cycle. 2011; 10: 2059 -2063. [PubMed] .
- 60. Balliet RM, Capparelli C, Guido C, Pestell TG, Martinez-Outschoorn UE, Lin Z, Whitaker-Menezes D, Chiavarina B, Pestell RG, Howell A, Sotgia F, Lisanti MP. Mitochondrial oxidative stress in cancer-associated fibroblasts drives lactate production, promoting breast cancer tumor growth: understanding the aging and cancer connection. Cell Cycle. 2011; 10: 4065 -4073. [PubMed] .
- 61. Campisi J. Senescent cells, tumor suppression, and organismal aging: good citizens, bad neighbors. Cell. 2005; 120: 513 -522. [PubMed] .
- 62. Vicente-Duenas C, Abollo-Jimenez F, Ruiz-Roca L, Alonso-Escudero E, Jimenez R, Cenador MB, Criado FJ, Cobaleda C, Sanchez-Garcia I. The age of the target cell affects B-cell leukaemia malignancy. Aging. 2010; 2: 908 -913. [PubMed] .
- 63. Bonuccelli G, Whitaker-Menezes D, Castello-Cros R, Pavlides S, Pestell RG, Fatatis A, Witkiewicz AK, Vander Heiden MG, Migneco G, Chiavarina B, Frank PG, Capozza F, Flomenberg N, Martinez-Outschoorn UE, Sotgia F, Lisanti MP. The reverse Warburg effect: glycolysis inhibitors prevent the tumor promoting effects of caveolin-1 deficient cancer associated fibroblasts. Cell Cycle. 2010; 9: 1960 -1971. [PubMed] .
- 64. Castello-Cros R, Bonuccelli G, Molchansky A, Capozza F, Witkiewicz AK, Birbe RC, Howell A, Pestell RG, Whitaker-Menezes D, Sotgia F, Lisanti MP. Matrix remodeling stimulates stromal autophagy, “fueling” cancer cell mitochondrial metabolism and metastasis. Cell Cycle. 2011; 10: 2021 -2034. [PubMed] .
- 65. Chiavarina B, Whitaker-Menezes D, Martinez-Outschoorn UE, Witkiewicz AK, Birbe RC, Howell A, Pestell RG, Smith J, Daniel R, Sotgia F, Lisanti MP. Pyruvate kinase expression (PKM1 and PKM2) in cancer-associated fibroblasts drives stromal nutrient production and tumor growth. Cancer Biol Ther. 2011; 12 .
- 66. Bonuccelli G, Tsirigos A, Whitaker-Menezes D, Pavlides S, Pestell RG, Chiavarina B, Frank PG, Flomenberg N, Howell A, Martinez-Outschoorn UE, Sotgia F, Lisanti MP. Ketones and lactate “fuel” tumor growth and metastasis: Evidence that epithelial cancer cells use oxidative mitochondrial metabolism. Cell Cycle. 2010; 9: 3506 -3514. [PubMed] .
- 67. Migneco G, Whitaker-Menezes D, Chiavarina B, Castello-Cros R, Pavlides S, Pestell RG, Fatatis A, Flomenberg N, Tsirigos A, Howell A, Martinez-Outschoorn UE, Sotgia F, Lisanti MP. Glycolytic cancer associated fibroblasts promote breast cancer tumor growth, without a measurable increase in angiogenesis: evidence for stromal-epithelial metabolic coupling. Cell Cycle. 2010; 9: 2412 -2422. [PubMed] .
- 68. Ko YH, Lin Z, Flomenberg N, Pestell RG, Howell A, Sotgia F, Lisanti MP, Martinez-Outschoorn UE. Glutamine fuels a vicious cycle of autophagy in the tumor stroma and oxidative mitochondrial metabolism in epithelial cancer cells: Implications for preventing chemotherapy resistance. Cancer Biol Ther. 2011; 12 .
- 69. Martinez-Outschoorn UE, Pestell RG, Howell A, Tykocinski ML, Nagajyothi F, Machado FS, Tanowitz HB, Sotgia F, Lisanti MP. Energy transfer in “parasitic” cancer metabolism: mitochondria are the powerhouse and Achilles’ heel of tumor cells. Cell Cycle. 2011; 10: 4208 -4216. [PubMed] .
- 70. Martinez-Outschoorn UE, Whitaker-Menezes D, Lin Z, Flomenberg N, Howell A, Pestell RG, Lisanti MP, Sotgia F. Cytokine production and inflammation drive autophagy in the tumor microenvironment: role of stromal caveolin-1 as a key regulator. Cell Cycle. 2011; 10: 1784 -1793. [PubMed] .
- 71. Capparelli C, Guido C, Whitaker-Menezes D, Bonuccelli G, Balliet R, Pestell TG, Goldberg AF, Pestell RG, Howell A, Sneddon S, Birbe R, Tsirigos A, Martinez-Outschoorn U, Sotgia F, Lisanti MP. Autophagy and senescence in cancer-associated fibroblasts metabolically supports tumor growth and metastasis via glycolysis and ketone production. Cell Cycle. 2012; 11: 2285 -2302. [PubMed] .
- 72. Mercier I, Camacho J, Titchen K, Gonzales DM, Quann K, Bryant KG, Molchansky A, Milliman JN, Whitaker-Menezes D, Sotgia F, Jasmin JF, Schwarting R, Pestell RG, Blagosklonny MV, Lisanti MP. Caveolin-1 and Accelerated Host Aging in the Breast Tumor Microenvironment: Chemoprevention with Rapamycin, an mTOR Inhibitor and Anti-Aging Drug. Am J Pathol. 2012; 181: 278 -293. [PubMed] .
- 73. Blagosklonny MV. Prevention of cancer by inhibiting aging. Cancer Biol Ther. 2008; 7: 1520 -1524. [PubMed] .
- 74. Hursting SD, Lavigne JA, Berrigan D, Perkins SN, Barrett JC. Calorie restriction, aging, and cancer prevention: mechanisms of action and applicability to humans. Annu Rev Med. 2003; 54: 131 -152. [PubMed] .
- 75. Longo VD and Fontana L. Calorie restriction and cancer prevention: metabolic and molecular mechanisms. Trends Pharmacol Sci. 2010; 31: 89 -98. [PubMed] .
- 76. Blagosklonny MV. Calorie restriction: Decelerating mTOR-driven aging from cells to organisms (including humans). Cell Cycle. 2010; 9: 683 -688. [PubMed] .
- 77. Hursting SD, Perkins SN, Phang JM. Calorie restriction delays spontaneous tumorigenesis in p53-knockout transgenic mice. Proc Natl Acad Sci U S A. 1994; 91: 7036 -7040. [PubMed] .
- 78. Berrigan D, Perkins SN, Haines DC, Hursting SD. Adult-onset calorie restriction and fasting delay spontaneous tumorigenesis in p53-deficient mice. Carcinogenesis. 2002; 23: 817 -822. [PubMed] .
- 79. Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandezr E, Miller RA. Rapamycin fed late in life extends lifespan in genetically heterogenous mice. Nature. 2009; 460: 392 -396. [PubMed] .
- 80. Anisimov VN, Zabezhinski MA, Popovich IG, Piskunova TS, Semenchenko AV, Tyndyk ML, Yurova MN, Antoch MP, Blagosklonny MV. Rapamycin extends maximal lifespan in cancer-prone mice. Am J Pathol. 2010; 176: 2092 -2097. [PubMed] .
- 81. Anisimov VN, Zabezhinski MA, Popovich IG, Piskunova TS, Semenchenko AV, Tyndyk ML, Yurova MN, Rosenfeld SV, Blagosklonny MV. Rapamycin increases lifespan and inhibits spontaneous tumorigenesis in inbred female mice. Cell Cycle. 2011; 10: 4230 -4236. [PubMed] .
- 82. Mathew T, Kreis H, Friend P. Two-year incidence of malignancy in sirolimus-treated renal transplant recipients: results from five multicenter studies. Clin Transplant. 2004; 18: 446 -449. [PubMed] .
- 83. Kauffman HM, Cherikh WS, Cheng Y, Hanto DW, Kahan BD. Maintenance immunosuppression with target-of-rapamycin inhibitors is associated with a reduced incidence of de novo malignancies. Transplantation. 2005; 80: 883 -889. [PubMed] .
- 84. Yakupoglu YK, Buell JF, Woodle S, Kahan BD. Individualization of Immunosuppressive Therapy. III. Sirolimus Associated With a Reduced Incidence of Malignancy. Transplant Proc. 2006; 38: 358 -361. [PubMed] .
- 85. Campistol JM, Eris J, Oberbauer R, Friend P, Hutchison B, Morales JM, Claesson K, Stallone G, Russ G, Rostaing L, Kreis H, Burke JT, Brault Y, Scarola JA, Neylan JF. Sirolimus Therapy after Early Cyclosporine Withdrawal Reduces the Risk for Cancer in Adult Renal Transplantation. J Am Soc Nephrol. 2006; 17: 581 -589. [PubMed] .
- 86. Stallone G, Schena A, Infante B, Di Paolo S, Loverre A, Maggio G, Ranieri E, Gesualdo L, Schena FP, Grandaliano G. Sirolimus for Kaposi's sarcoma in renal-transplant recipients. N Engl J Med. 2005; 352: 1317 -1323. [PubMed] .
- 87. Pinkston JM, Garigan D, Hansen M, Kenyon C. Mutations that increase the life span of C. elegans inhibit tumor growth. Science. 2006; 313: 971 -975. [PubMed] .
- 88. Poyurovsky MV and Prives C. P53 and aging: A fresh look at an old paradigm. Aging. 2010; 2: 380 -382. [PubMed] .
- 89. Blagosklonny MV. Revisiting the antagonistic pleiotropy theory of aging: TOR-driven program and quasi-program. Cell Cycle. 2010; 9: 3151 -3156. [PubMed] .
- 90. de Keizer PL, Laberge RM, Campisi J. p53: Pro-aging or pro-longevity? Aging. 2010; 2: 377 -379. [PubMed] .
- 91. Chao SK, Horwitz SB, McDaid HM. Insights into 4E-BP1 and p53 mediated regulation of accelerated cell senescence. Oncotarget. 2011; 2: 89 -98. [PubMed] .
- 92. Matheu A, Maraver A, Klatt P, Flores I, Garcia-Cao I, Borras C, Flores JM, Vi-a J, Blasco MA, Serrano M. Delayed ageing through damage protection by the Arf/p53 pathway. Nature. 2007; 448: 375 -379. [PubMed] .
- 93. Donehower LA, Harvey M, Slagle BL, McArthur MJ, Montgomery CA Jr., Butel JS, Bradley A. Mice deficient for p53 are developmentally normal but susceptible to spontaneous tumours. Nature. 1992; 356: 215 -221. [PubMed] .
- 94. Harvey M, McArthur MJ, Montgomery CA Jr., Butel JS, Bradley A, Donehower LA. Spontaneous and carcinogen-induced tumorigenesis in p53-deficient mice. Nat Genet. 1993; 5: 225 -229. [PubMed] .
- 95. Jacks T, Remington L, Williams BO, Schmitt EM, Halachmi S, Bronson RT, Weinberg RA. Tumor spectrum analysis in p53-mutant mice. Curr Biol. 1994; 4: 1 -7. [PubMed] .
- 96. Donehower LA, Harvey M, Vogel H, McArthur MJ, Montgomery CA Jr., Park SH, Thompson T, Ford RJ, Bradley A. Effects of genetic background on tumorigenesis in p53-deficient mice. Mol Carcinog. 1995; 14: 16 -22. [PubMed] .
- 97. Venkatachalam S, Shi YP, Jones SN, Vogel H, Bradley A, Pinkel D, Donehower LA. Retention of wild-type p53 in tumors from p53 heterozygous mice: reduction of p53 dosage can promote cancer formation. Embo J. 1998; 17: 4657 -4667. [PubMed] .
- 98. Hinkal G, Parikh N, Donehower LA. Timed somatic deletion of p53 in mice reveals age-associated differences in tumor progression. PLoS One. 2009; 4: e6654 [PubMed] .
- 99. Guevara NV, Kim HS, Antonova EI, Chan L. The absence of p53 accelerates atherosclerosis by increasing cell proliferation in vivo. Nat Med. 1999; 5: 335 -339. [PubMed] .
- 100. Mercer J, Figg N, Stoneman V, Braganza D, Bennett MR. Endogenous p53 protects vascular smooth muscle cells from apoptosis and reduces atherosclerosis in ApoE knockout mice. Circ Res. 2005; 96: 667 -674. [PubMed] .
- 101. Mercer J and Bennett M. The role of p53 in atherosclerosis. Cell Cycle. 2006; 5: 1907 -1909. [PubMed] .
- 102. van Vlijmen BJ, Gerritsen G, Franken AL, Boesten LS, Kockx MM, Gijbels MJ, Vierboom MP, van Eck M, van De Water B, van Berkel TJ, Havekes LM. Macrophage p53 deficiency leads to enhanced atherosclerosis in APOE*3-Leiden transgenic mice. Circ Res. 2001; 88: 780 -786. [PubMed] .
- 103. dinger AL, Linardic CM, Chiang GG, Thompson CB, Abraham RT. Differential effects of rapamycin on mammalian target of rapamycin signaling functions in mammalian cells. Cancer Res. 2003; 63: 8451 -8460. [PubMed] .
- 104. Sipula IJ, Brown NF, Perdomo G. Rapamycin-mediated inhibition of mammalian target of rapamycin in skeletal muscle cells reduces glucose utilization and increases fatty acid oxidation. Metabolism. 2006; 55: 1637 -1644. [PubMed] .
- 105. Brown NF, Stefanovic-Racic M, Sipula IJ, Perdomo G. The mammalian target of rapamycin regulates lipid metabolism in primary cultures of rat hepatocytes. Metabolism. 2007; 56: 1500 -1507. [PubMed] .
- 106. Hu W, Feng Z, Teresky AK, Levine AJ. p53 regulates maternal reproduction through LIF. Nature. 2007; 450: 721 -724. [PubMed] .
- 107. Hu W, Feng Z, Atwal GS, Levine AJ. p53: a new player in reproduction. Cell Cycle. 2008; 7: 848 -852. [PubMed] .
- 108. Roemer K. Are the conspicuous interdependences of fecundity, longevity and cognitive abilities in humans caused in part by p53? Cell Cycle. 2010; 9: 3438 -3441. [PubMed] .
- 109. Levine AJ, Tomasini R, McKeon FD, Mak TW, Melino G. The p53 family: guardians of maternal reproduction. Nat Rev Mol Cell Biol. 2011; 12: 259 -265. [PubMed] .
- 110. Mantovani R. More on the pro-fertility activity of p53: the blastocyst side. Cell Cycle. 2011; 10: 4205 [PubMed] .
- 111. Chen D, Zheng W, Lin A, Uyhazi K, Zhao H, Lin H. Pumilio 1 suppresses multiple activators of p53 to safeguard spermatogenesis. Curr Biol. 2012; 22: 420 -425. [PubMed] .
- 112. McGee MD, Day N, Graham J, Melov S. cep-1/p53-dependent dysplastic pathology of the aging C. elegans gonad. Aging. 2012; 4: 256 -269. [PubMed] .
- 113. Kang HJ, Feng Z, Sun Y, Atwal G, Murphy ME, Rebbeck TR, Rosenwaks Z, Levine AJ, Hu W. Single-nucleotide polymorphisms in the p53 pathway regulate fertility in humans. Proc Natl Acad Sci U S A. 2009; 106: 9761 -9766. [PubMed] .