



Introduction
Aging is associated with a progressive loss of skeletal muscle mass and function across multiple organisms [1, 2]. This can manifest as an inability to perform activities of daily living or maintain mobility [3]. Animal models have been developed to understand mechanisms underlying this aging-associated loss of muscle mass and identify potential therapeutic targets. Currently available animal models of aging, all of which exhibit loss of muscle and function to varying extent, include zebrafish [4], mouse [5], and rat [6]. Murine models, in particular, have been crucial in identifying potential therapeutic targets for improving skeletal muscle maintenance with aging [7]. However, although animal models provide a useful substrate for medical and scientific research, discordant findings have been commonly described in the existing literature between animals and humans at the transcriptional level [8]. A divergence in the transcriptomics between mouse and human have been described in embryogenesis [9], nephrogenesis [10, 11], neurological and cortical architecture and expression [12], macrophage stress responses [13, 14] and healthy myocardium [15, 16]. This may partially explain a poor rate of successful translation from bench to bedside. Given this, understanding whether the transcriptome diverges with aging in humans versus mice is critical in understanding the appropriateness of using a murine model of aging in skeletal muscle.
Previous evaluation of human muscle samples demonstrated a significant transcriptomic shift with aging in skeletal muscle, particularly in genes associated with cellular senescence and insulin signaling [17]. In contrast, similar studies in aging mouse models have highlighted increases in genes related to mitochondrial function and inflammation [18] and loss of hypoxia pathway signaling [5, 2] in muscle. Other tissue-types have demonstrated varying levels of similarities between the transcriptome with aging [19, 20]. With regards to skeletal muscle, specifically, Borsch et al. suggested that many pathways were conserved among mice, rats, and humans, up to the age of 70 years old with regards to the transcriptomic changes with aging [21]. The present study selected data from healthy human subjects aged 80 years and older, to provide a comparison point at an age where substantial muscle loss has occurred [22]. For comparison, 24- to 25-month-old mice were evaluated, as mice of that age similarly demonstrate a functional loss of mass and function and are similarly aged to 80-year-old humans with regards to lifespan [23].
Results
Mouse and human muscles exhibit divergent transcriptomic phenotypes
NMR analyses of mice demonstrated significantly increased proportion of fat and commensurately decreased proportion of muscle and tetanic strength in the setting of similar total body weights (Supplementary Figure 1).
The transcriptional data were obtained from mouse gastrocnemius muscles through bulk-RNA sequencing performed on harvested muscles and from human vastus lateralis muscles through abstraction from NIH GEO. Raw counts were pre-processed with filters and normalization with resultant gene and count matrix files (Figure 1A). Volcano plots demonstrate genes with at least a 2-fold difference in expression levels (statistically significant) in mouse and human (Figure 1B). In gene ontology enrichment for biological processes, molecular functions, and cellular components, there were 1, 2, and 0 overlapping terms respectively. KEGG pathway database terms yielded 2 overlapping terms. These reflect less than 5% of the total number of statistically significant enriched keywords and pathways (Figure 1C).
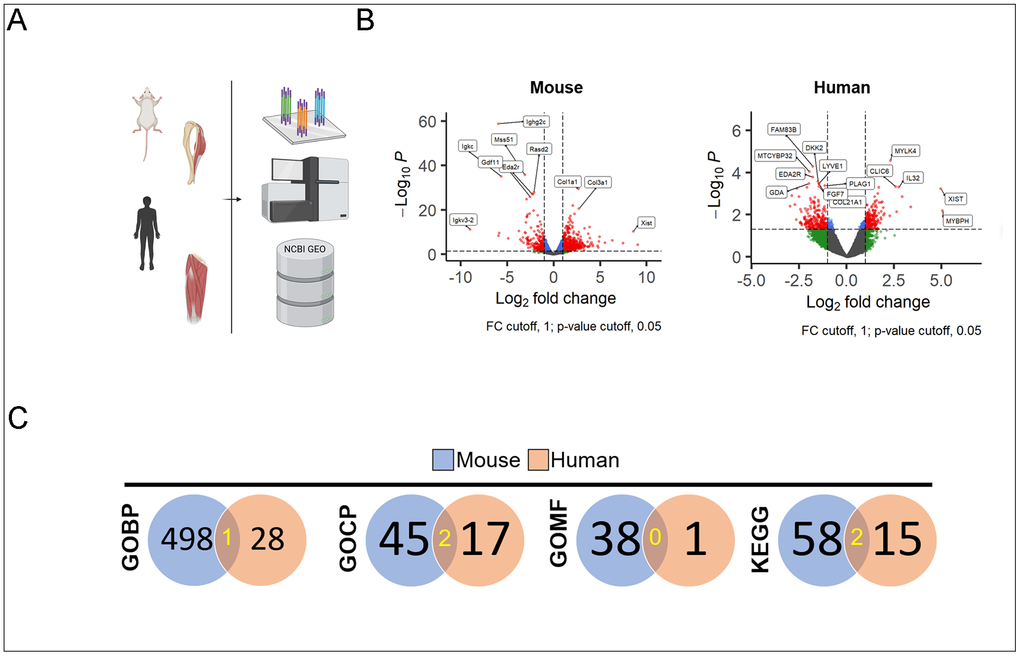
Figure 1. Schematic of mouse and human bulk-seq analysis and processing (A). Volcano plots demonstrate those with statistical significance and absolute value of fold change >1 (red) in mouse (left) and human (right) muscles (B). Counts of statistically significant enrichment terms derived from upregulated DEG in both old vs. young mice (blue circle) and humans (red circle) with the number of overlapping terms highlighted in yellow (C).
Mouse samples yielded an average of 25M reads per sample (n=6, min 23.3M, max 27.6M). 49315 differentially expressed genes in old vs. young comparison with 14472 matched ENSEMBL genes, 2760 genes with adjusted Benjamini–Hochberg FDR statistical significance (p.adj < 0.05). Human samples (n=13) yielded an average of 53M reads per sample (min 7.8M, max 118.8M). 57773 differentially expressed genes in old vs. young comparison with 21638 matched ENSEMBL genes, 613 genes with adjusted (Benjamini-Hochberg FDR) statistical significance (p.adj < 0.05).
Mouse and human muscles exhibit heterogeneity in transcriptome with aging
Head-to-head comparisons of enrichment terms between species, stratified by the up- or down-regulation of transcription in young versus old. GO BP and KEGG terms are shown in Supplementary Table 1.
The 20 genes most upregulated with aging (expression increased in old muscle compared to young muscle) in mice and humans were visualized (Figure 2). Examination demonstrated little overlap between mouse and human. The two with similarities included osteocrin (Ostn; also known as musclin [24], a connective tissue derived humoral factor [25] with protective effects in setting of acute metabolic stressors on kidney [26] and muscle [27, 28] as well as ankyrin repeat domain-containing protein 1 (Ankrd1; also known as cardiac ankyrin repeat protein CARP) Ankrd11p1, a putative pseudogene resembling an ankyrin isotype involved in stress responses within skeletal muscle [29], as well as bony/cardiac development [30–33].
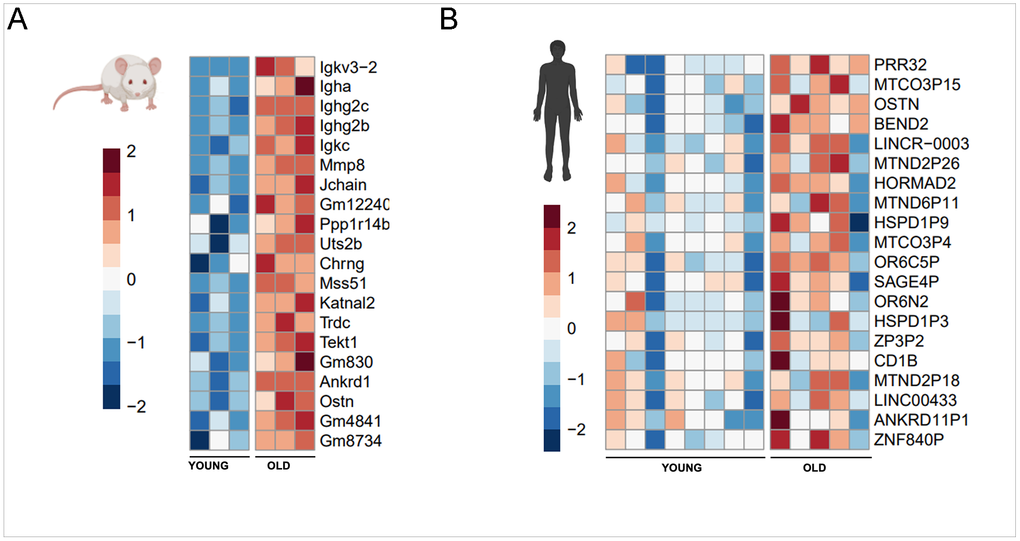
Figure 2. 20 genes with greatest increase in expression with aging in mice (A) and human (B) muscle.
Between species, older mice demonstrated relative upregulation of inflammatory terms including several families of immunoglobulins and regulation of their binding/downstream responses (Igkv3-2 [34], Igha [35], Ighg2c [36], Ighg2b [37], Igkc [38], Jchain [39], Ppp1r14b [40], Trdc [41], and Gm4841 [42]. In the human counterpart, a less unifying motif was found with various terms pertaining to modes of replication and transcriptional regulation via chromatin structuring (BEND2 [43], HORMAD2 [44], long noncoding RNA modulation of Yap1 signaling (LINCR-0003 [45], and zinc finger control of RNA polymerase II (Znf840p [46] along with various pseudogenes pertaining to stress response via heat shock proteins (HSPD1P9, HSPD1P3).
Examination of the genes most differentially expressed in aging with lower expression in old muscle similarly limited overlap across species (Figure 3). Of note, X inactive specific transcript (Xist [47]) is found in the top 20 genes in both mouse and human, a long non-coding RNA that has been shown to have regulatory effects on the Wnt/β-catenin pathway in the proliferation of vascular smooth muscle cells, fibroblasts [47], and upregulation of the larger Hif-1a hypoxia axis in oncologic contexts [48–50].
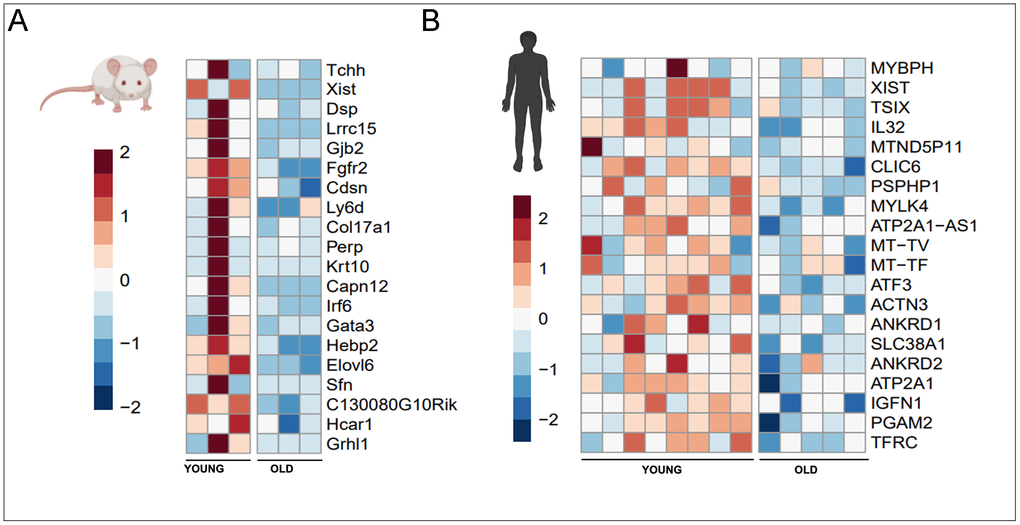
Figure 3. 20 genes with greatest decrease in expression with aging in mice (A) and human (B) muscle.
In the mouse, other genes which exhibit less expression in old muscle include structural genes which govern cytoskeletal homeostasis (Tchh [50], Dsp [51], Hebp2 [52] and cell surface contact and receptor transduction for cellular proliferation/differentiation (Lrrc15 [53], Gjb2 [54], Fgfr2 [55], Col17a1 [56], Perp [57], Krt10 [58], Irf6 [59], Gata3 [60], Sfn [61], Hcar1 [62]. Of note, in humans TSIX [63] was found to be differentially expressed in young human muscle – a non-coding antisense transcript to XIST that results in its silencing in mouse tissues but conversely is found to potentiate human stem cell pluripotency [63]. The other genes specific to human include constituents of muscle (MYBPH [64], MYLK4 [65], ACTN3 [66], IFGN1 [67], various cytokines and regulators of inflammatory axis (IL32 [68], ATF3 [69–71]), and mitochondrial genes (MT-TV [72], MT-TF [73]), TFRC [74]. While Ankrd1 was found upregulated in old mice, this same gene was found differentially upregulated in young humans. Another member of the ankyrin family, Ankrd2 was also found similarly upregulated.
Mice and humans demonstrate variable homology across hypoxia, vascular, neurogenic, and muscular programs
Given demonstrated transcriptome differences across age and species, we subsequently examined accepted gene lists for various pathways implemented into standard gene expression arrays (Qiagen, RT2 profiler PCR arrays) to further delineate the transcriptome patterns across species and age. In animal models, hypoxia signaling [2], VEGFA signaling [5, 75], as well as angiogenic capacity [76, 77] have been demonstrated to downregulate with aging muscle. Given previous works corroborating age related changes in nerve regeneration [78–80] and axonal transport [81], the neurogenesis pathway was also hypothesized to show an inverse relationship between the young and old. Outside of these extra-myogenic pathways regulating nascent muscle growth, aging has been also highlighted as a risk factor for impaired myogenesis at the myoblast and progenitor level [82, 83], implicating a reduced myogenic potential in aged muscle.
Principal component analysis (PCA) of the transcriptomic data highlighted notable differences in inter-sample variance between species. In mice, young (blue) and old (red) samples formed largely distinct clusters along PC1 (39% variance explained), indicating a clear and consistent age-related separation in gene expression (Supplementary Figure 2A). In contrast, the human PCA revealed considerable overlap between young and old samples, with PC1 accounting for 48.2% of variance yet failing to resolve the two age groups (Supplementary Figure 2B). This overlap reflects the inherently high biological variability present in human skeletal muscle transcriptomics, compounded by the disproportionate and modest sample sizes in this cohort (n=8 young, n=5 old). Factors such as sex, baseline fitness, and the broader genetic diversity of the human population likely contribute to this transcriptional heterogeneity, underscoring a fundamental challenge in cross-sectional human aging studies [83, 84]. Consequently, differential expression filtering and downstream heatmap visualization in mice were performed using a stringent FDR threshold of <0.1, while pairwise comparisons were assessed by Student's t-test. In humans, however, given that few genes achieved statistical significance after multiple testing correction, unadjusted p-values <0.05 were used to retain biologically relevant signal for visualization and pathway-level interpretation.
Visualization and pathway scoring revealed relative downregulation of the hypoxia and VEGFA pathways with age in both humans and mice (Figure 4A, 4B). Consistent with previous literature, angiogenesis and neurogenesis were also downregulated in aged mice; however, in humans, angiogenesis showed upregulation while neurogenesis did not reach statistical significance (Figure 4C, 4D). The myogenesis pathway showed no concordance between species (Figure 4E): while aged human patients demonstrated downregulation of the pro-myogenic transcriptome, this pattern was not recapitulated in mice at the ages tested. Finally, aging was associated with increased inflammatory and cytokine signaling in both species (Figure 4F).
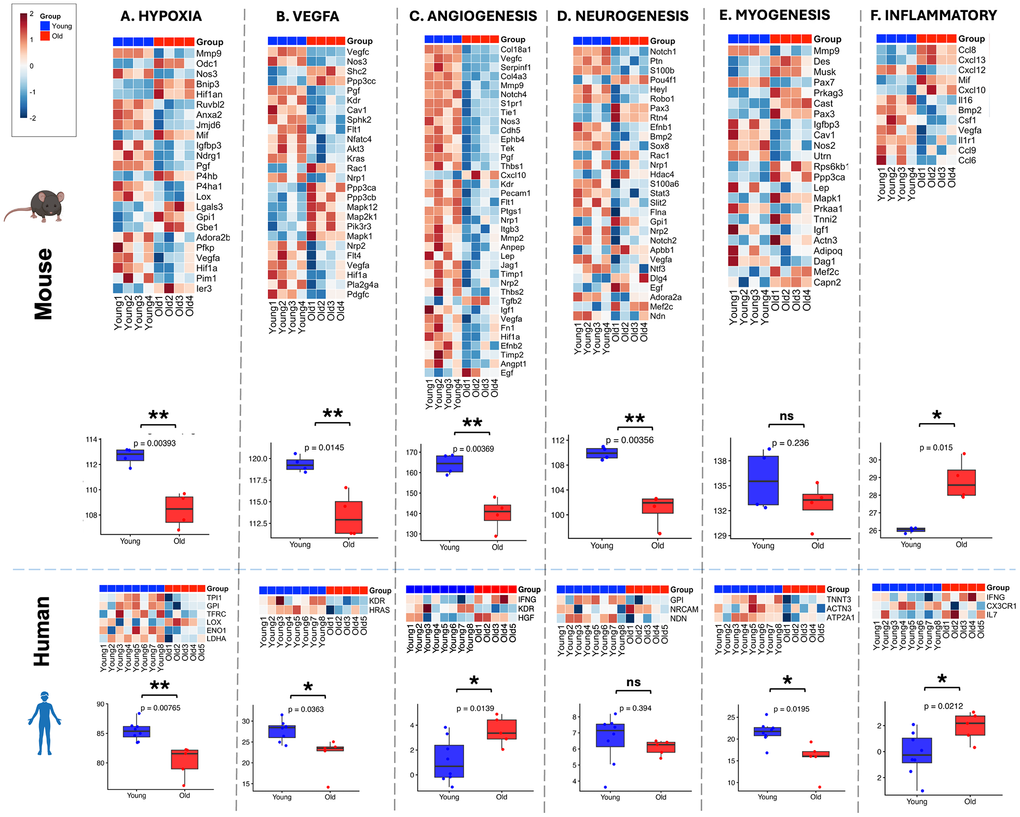
Figure 4. Pathway gene expression during aging in mouse and human tissues. Heatmaps display normalized logCPM expression of pathway genes. Differential expression in mouse samples was determined using edgeR with Benjamini–Hochberg correction (FDR < 0.1). In the human dataset, statistical significance was defined as p < 0.05 (unadjusted) due to high biological heterogeneity and limited sample size.
Discussion
Mice have long been accepted as an important model organism for studying the biological basis of human diseases given the high degrees of gene orthology between human and mouse (80% and 72%, respectively) [85], extensive annotations [86] and flexibility offered by genetic manipulations through Cre-lox transgenic systems and CRISPR Cas9 [87] technologies. However, similarities in genetic blueprint do not necessarily translate into transcriptional or translational concordance, which may impact the validity of biological inferences between mouse models and human disease [88]. In the present study, examination of RNA-Seq datasets from the gastrocnemius muscles of mice and vastus lateralis muscle of humans demonstrated divergent gene expression patterns in dimensional analyses across young and old age groups.
Consistent with prior report [88], human samples exhibited greater variance compared to mice. Volcano plot analysis revealed a larger number of genes with at least a 2-fold statistically significant difference in expression in mice relative to humans. Notably, muscles from young and old mice, which phenotypically exhibit decreased tetanic strength with age, displayed a distinctly binary transcriptional pattern. In contrast, human muscle samples demonstrated a more heterogeneous spectrum of gene expression profiles across age groups. Enrichment analysis of significantly upregulated genes revealed minimal overlap between species, with two or fewer shared terms across Gene Ontology categories and KEGG pathways. While mice on a uniform genetic background demonstrated clear age-dependent clustering, human samples were widely dispersed, suggesting substantial diversity in transcriptional states despite comparable age extremes.
To further investigate these differences, we performed direct cross-species comparisons of enrichment terms stratified by transcriptional directionality. In young mouse muscle, upregulated pathways reflected anabolic and structural processes, including extracellular matrix organization, cell migration, vascular and neural ingrowth, and stemness, relative to aged muscle. In contrast, young human muscle was characterized by enrichment of metabolic pathways, including pyruvate metabolism, glycolysis, oxidative phosphorylation, mitochondrial function, and muscle contraction (GO BP and KEGG terms shown in Supplementary Table 1).
Within human muscle, the most prominent age-associated decline involved metabolic pathways, which were significantly enriched in young compared to old samples. This aligns with established evidence that metabolism declines with aging [88], with skeletal muscle contributing to systemic metabolic dysfunction through impaired mitochondrial activity and insulin sensitivity [89]. These metabolic signatures were not significantly altered in mice, consistent with prior observations that fundamental differences in energy expenditure and metabolic regulation exist between species [90], potentially limiting translational relevance in this domain.
In contrast, murine muscle exhibited significant downregulation of pathways related to cellular proliferation and extracellular matrix (ECM) maintenance with aging [91]. Age-associated decline in muscle regenerative capacity has been extensively characterized in murine models [7, 92, 93], whereas direct evaluation in humans remains limited, though emerging ex vivo studies suggest similar trends [94]. Consistent with our findings, ECM composition and structural integrity decline with age in murine skeletal muscle, with preliminary evidence suggesting comparable changes in humans [95]. Given the role of the ECM as a reservoir for growth factors [96] and a regulator of muscle stem cell function [97], these alterations may directly influence regenerative capacity.
Cross-referencing enrichment analyses with the top differentially expressed genes further highlighted a consistent divergence in age-associated transcriptional programs between species. Notably, prior work comparing aging across mice, rats, and humans has reported greater transcriptomic similarity [21]. Several factors may account for these differences. First, muscle sampling differed between studies, with our analysis comparing gastrocnemius in mice to vastus lateralis in humans, whereas prior work utilized the same muscle group across species. Second, our human cohort included individuals over 80 years of age, in contrast to previous studies limited to individuals ≤70 years, potentially capturing a more advanced stage of transcriptomic remodeling associated with aging [17, 98].
Building on these observations, we examined several key regulatory pathways implicated in muscle aging, including VEGFA signaling, angiogenesis, neurogenesis, hypoxia response, inflammatory signaling, and myogenesis. Despite global transcriptomic divergence, select pathways demonstrated concordant age-related downregulation across species. Notably, while myogenesis did not reach statistical significance at the pathway level in mice, individual genes with established roles in muscle regeneration were differentially expressed, including Pax7 [99] and Igf-1 [100]. These findings suggest that, despite broad species-level differences, critical components of muscle aging biology remain conserved.
While the primary goal of this study was to characterize transcriptomic differences in healthy skeletal muscle aging between species, we acknowledge that therapeutic inference represents a valuable and clinically meaningful extension of these findings. The concordant downregulation of hypoxia and VEGFA signaling across both species supports the potential relevance of pro-angiogenic interventions in aged muscle, with agents targeting the HIF-1α axis and VEGF-mediated pathways warranting further investigation ([101, 102]). Similarly, the shared upregulation of inflammatory signaling across species lends biological rationale to anti-inflammatory strategies targeting the IL-6 and NF-κB pathways in the context of muscle aging [103, 104]. The divergence observed in neurogenic pathway expression between species, however, raises important questions regarding the direct translatability of pro-neurogenic approaches such as BDNF- or NT-3-based therapies from murine models to human aging muscle [105, 106]. These conserved pathways may represent viable cross-species therapeutic targets and should be explored in future pharmacological and interventional studies.
Several limitations should be considered. A fundamental distinction lies in genetic variability: human populations exhibit substantial inter-individual diversity, whereas inbred mouse strains are genetically homogeneous. This discrepancy likely contributes to the greater transcriptional heterogeneity observed in human samples and limits direct cross-species comparisons. The use of outbred mouse strains, such as CD-1 or CFW, may better model human population variability and should be explored in future studies [107]. Additionally, modest sample sizes, particularly in the human cohort (n=8 young, n=5 old), increase susceptibility to sampling bias despite rigorous preprocessing. Statistical thresholds also differed between species, with FDR < 0.1 applied in mice and unadjusted p-values used in humans due to limited significance after multiple testing correction.
Technical considerations further include the use of bulk RNA sequencing, which lacks cellular resolution and precludes identification of the specific cell populations driving observed transcriptional changes. Age-related shifts in cellular composition, including immune cell infiltration, may contribute to these differences. Additionally, variation in muscle type between species may influence transcriptomic profiles [108]. Finally, as a cross-sectional study, these analyses capture a single timepoint and do not reflect longitudinal transcriptional dynamics across aging.
Despite these limitations, the partial convergence of key aging-associated pathways supports the continued utility of murine models in studying skeletal muscle aging. However, our findings emphasize the need for careful interpretation of cross-species transcriptomic data and underscore the importance of integrating human datasets to validate and contextualize preclinical observations.
Materials and Methods
Animals
All animal procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Brigham and Women's Hospital (Protocol number: 2016N000375) and were performed in accordance with the U.S. National Institutes of Health guidelines. 20-22-week-old young (n=4) and 24-25-months old (n=4) C57BL/6 mice were obtained from Jackson Laboratories and the National Institution of Aging, respectively. The animals were housed at the Brigham and Women’s Hospital Animal Care Facility following a 12 light / 12 dark cycle and were given ad libitum access to food and water.
Murine nuclear magnetic resonance imaging
Body composition, whole body fat mass, lean body mass, free water and total body water were measured, prior to sacrifice, by nuclear magnetic resonance (NMR) using the EchoMRI-100™ QMR system (Houston, TX) as previously described [109].
Murine in situ force testing
In situ force tests were performed prior to muscle sacrifice and following NMR testing as previously described [110]. Briefly, the right gastrocnemius (GA) muscle and Achilles tendon were isolated. The tendon was then transected at the distal most point of insertion and then sutured onto the lever arm of the force transducer (Aurora Scientific, Aurora, Ontario, Canada). Direct stimulation of the muscle was performed with needle electrode placement within the muscle. The tetanic force was measured with pulses of 100 Hz with increasing amperage from 10mA to 1A. The maximum tetanic force of the right GA muscle of each animal was normalized to the estimated cross-sectional area (CSA) of the muscle (mm2) calculated as CSA = Muscle weight (mg)/[1.06×Lo(mm)].
RNA Bulk-sequencing analysis
Gastrocnemius muscle was harvested from the animals and immediately snap-frozen using liquid nitrogen for the subsequent analyses. Total RNA was extracted using TRIzole reagent. Following rRNA deletion, the sequencing library was prepared using Illumina HiSeq® at 2x150bp configuration, with the minimal read of 30 million base pairs per sample. After quality assessment and adaptor trimming, the reads were aligned to the reference genome using STAR [111]. The expression level of each gene was then estimated by counting the number of reads that aligned to each exon using FeatureCounts [112] and normalized as fragments per kilobase per million reads (FPKM) and log2 transformation. Resulting matrix files were further filtered for lowly expressed genes and gene inclusion criteria 10/L ratio as described by Chen et al. [113]. Differential expression analysis was performed using edgeR [113] to compare the gene expression in the murine old and young groups (database available in GEO database: GSE155937 [114]. Sample similarity was assessed by dimensional reduction via biological coefficient of variance (BCV) distances.
GEO database comparative analysis and pathway scoring
Data from healthy human vastus lateralis muscles (young, 22-30 years, n=8; old, 80-83 years, n=5) were derived from the GESTALT study (NIA/NIH) as found in the Gene Expression Omnibus (NIH) database (GSE164471, [17]). Differential expression profile was determined via edgeR as described above.
Known QIAGEN microarray genesets were investigated (Qiagen, RT2 profiler PCR arrays): Hypoxia (PAMM-032Z), Vegf (PAMM-091Z, AHS-091Z), Angiogenesis (PAMM-024Z, PAHS-024Z), Neurogenesis (PAMM-404Z, PAHS-404Z), Myogenesis (PAMM-099Z, PAHS-099Z), and Inflammatory Cytokines and Receptors (PAMM-011Z, PAHS-011Z). Normalized counts across all genes were summed for each sample or patient to generate a representative pathway score. Young and old cohorts were compared using Student’s t-test, with significance thresholds set at FDR < 0.10 (Benjamini-Hochberg correction) for mouse and unadjusted p < 0.05 for human samples, the latter reflecting the high inter-individual biological variability inherent to human cohorts.
Gene identification and enrichment/ontology
Gene annotations performed with AnnotationHub (Bioconductor) via mouse (AH98078, Ensembl 105 database for Mus musculus) and human (AH98047, Ensembl 105 database for Homo sapiens) libraries. Agnostic gene enrichment and ontology analyses were performed with Enrichr [115], utilizing Gene Ontology (BP2021, CC2021, MF2021) and KEGG pathway libraries (mouse 2019, human 2021).
Author Contributions
CH and SR contributed equally to this work and share co-first authorship. CH and SR performed transcriptomic and statistical analyses. CH, SR, and IS drafted and revised the manuscript. YE, SC, LW, JM, and SM contributed to study design and execution of animal experiments. All authors reviewed and approved the final manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement and Consent
All animal procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Brigham and Women's Hospital (Protocol No. 2016N000375) and were conducted in accordance with the guidelines set forth by the U.S. National Institutes of Health. Human skeletal muscle transcriptomic data were obtained exclusively from a publicly available repository, the Gene Expression Omnibus (GEO) database (Accession No. GSE164471), derived from the Genotype-Tissue Expression in Aging Long-Term (GESTALT) study conducted by the National Institute on Aging (NIA/NIH). As no new human participants were recruited, examined, or contacted in this study, additional ethical approval and informed consent were not required.
Funding
This study was supported by a grant from the National Institutes of Health (R01AG078307) to IS.
References
- 1. Endo Y, Nourmahnad A, Sinha I. Optimizing Skeletal Muscle Anabolic Response to Resistance Training in Aging. Front Physiol. 2020; 11:874. https://doi.org/10.3389/fphys.2020.00874 [PubMed]
- 2. Endo Y, Baldino K, Li B, Zhang Y, Sakthivel D, MacArthur M, Panayi AC, Kip P, Spencer DJ, Jasuja R, Bagchi D, Bhasin S, Nuutila K, et al. Loss of ARNT in skeletal muscle limits muscle regeneration in aging. FASEB J. 2020; 34:16086–104. https://doi.org/10.1096/fj.202000761RR [PubMed]
- 3. Newman AB, Blackwell TL, Mau T, Cawthon PM, Coen PM, Cummings SR, Toledo FG, Goodpaster BH, Glynn NW, Hepple RT, Kritchevsky SB. Vigor to Frailty As a Continuum-A New Approach in the Study of Muscle, Mobility, and Aging Cohort. J Gerontol A Biol Sci Med Sci. 2024; 79:glad244. https://doi.org/10.1093/gerona/glad244 [PubMed]
- 4. Callegari S, Mirzaei F, Agbaria L, Shariff S, Kantawala B, Moronge D, Ogendi BM. Zebrafish as an Emerging Model for Sarcopenia: Considerations, Current Insights, and Future Directions. Int J Mol Sci. 2023; 24:17018. https://doi.org/10.3390/ijms242317018 [PubMed]
- 5. Endo Y, Hwang CD, Zhang Y, Olumi S, Koh DJ, Zhu C, Neppl RL, Agarwal S, Sinha I. VEGFA Promotes Skeletal Muscle Regeneration in Aging. Adv Biol (Weinh). 2023; 7:e2200320. https://doi.org/10.1002/adbi.202200320 [PubMed]
- 6. Franczak E, Maurer A, Drummond VC, Kugler BA, Wells E, Wenger M, Peelor FF 3r, Crosswhite A, McCoin CS, Koch LG, Britton SL, Miller BF, Thyfault JP. Divergence in aerobic capacity and energy expenditure influence metabolic tissue mitochondrial protein synthesis rates in aged rats. Geroscience. 2024; 46:2207–22. https://doi.org/10.1007/s11357-023-00985-1 [PubMed]
- 7. Conboy IM, Conboy MJ, Wagers AJ, Girma ER, Weissman IL, Rando TA. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. Nature. 2005; 433:760–4. https://doi.org/10.1038/nature03260 [PubMed]
- 8. Su AI, Cooke MP, Ching KA, Hakak Y, Walker JR, Wiltshire T, Orth AP, Vega RG, Sapinoso LM, Moqrich A, Patapoutian A, Hampton GM, Schultz PG, Hogenesch JB. Large-scale analysis of the human and mouse transcriptomes. Proc Natl Acad Sci USA. 2002; 99:4465–70. https://doi.org/10.1073/pnas.012025199 [PubMed]
- 9. Boroviak T, Stirparo GG, Dietmann S, Hernando-Herraez I, Mohammed H, Reik W, Smith A, Sasaki E, Nichols J, Bertone P. Single cell transcriptome analysis of human, marmoset and mouse embryos reveals common and divergent features of preimplantation development. Development. 2018; 145:dev167833. https://doi.org/10.1242/dev.167833 [PubMed]
- 10. Lindström NO, Guo J, Kim AD, Tran T, Guo Q, De Sena Brandine G, Ransick A, Parvez RK, Thornton ME, Baskin L, Grubbs B, McMahon JA, Smith AD, McMahon AP. Conserved and Divergent Features of Mesenchymal Progenitor Cell Types within the Cortical Nephrogenic Niche of the Human and Mouse Kidney. J Am Soc Nephrol. 2018; 29:806–24. https://doi.org/10.1681/ASN.2017080890 [PubMed]
- 11. Lindström NO, McMahon JA, Guo J, Tran T, Guo Q, Rutledge E, Parvez RK, Saribekyan G, Schuler RE, Liao C, Kim AD, Abdelhalim A, Ruffins SW, et al. Conserved and Divergent Features of Human and Mouse Kidney Organogenesis. J Am Soc Nephrol. 2018; 29:785–805. https://doi.org/10.1681/ASN.2017080887 [PubMed]
- 12. Hodge RD, Bakken TE, Miller JA, Smith KA, Barkan ER, Graybuck LT, Close JL, Long B, Johansen N, Penn O, Yao Z, Eggermont J, Höllt T, et al. Conserved cell types with divergent features in human versus mouse cortex. Nature. 2019; 573:61–8. https://doi.org/10.1038/s41586-019-1506-7 [PubMed]
- 13. Vijayan V, Pradhan P, Braud L, Fuchs HR, Gueler F, Motterlini R, Foresti R, Immenschuh S. Human and murine macrophages exhibit differential metabolic responses to lipopolysaccharide - A divergent role for glycolysis. Redox Biol. 2019; 22:101147. https://doi.org/10.1016/j.redox.2019.101147 [PubMed]
- 14. Seok J, Warren HS, Cuenca AG, Mindrinos MN, Baker HV, Xu W, Richards DR, McDonald-Smith GP, Gao H, Hennessy L, Finnerty CC, López CM, Honari S, et al., and Inflammation and Host Response to Injury, Large Scale Collaborative Research Program. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc Natl Acad Sci USA. 2013; 110:3507–12. https://doi.org/10.1073/pnas.1222878110 [PubMed]
- 15. Zhao Y, Sheng Z, Huang J. A systematic analysis of heart transcriptome highlights divergent cardiovascular disease pathways between animal models and humans. Mol Biosyst. 2012; 8:504–10. https://doi.org/10.1039/c1mb05415e [PubMed]
- 16. Anzai T, Yamagata T, Uosaki H. Comparative Transcriptome Landscape of Mouse and Human Hearts. Front Cell Dev Biol. 2020; 8:268. https://doi.org/10.3389/fcell.2020.00268 [PubMed]
- 17. Tumasian RA 3r, Harish A, Kundu G, Yang JH, Ubaida-Mohien C, Gonzalez-Freire M, Kaileh M, Zukley LM, Chia CW, Lyashkov A, Wood WH 3r, Piao Y, Coletta C, et al. Skeletal muscle transcriptome in healthy aging. Nat Commun. 2021; 12:2014. https://doi.org/10.1038/s41467-021-22168-2 [PubMed]
- 18. Kang YK, Min B, Eom J, Park JS. Different phases of aging in mouse old skeletal muscle. Aging (Albany NY). 2022; 14:143–60. https://doi.org/10.18632/aging.203812 [PubMed]
- 19. Zahn JM, Poosala S, Owen AB, Ingram DK, Lustig A, Carter A, Weeraratna AT, Taub DD, Gorospe M, Mazan-Mamczarz K, Lakatta EG, Boheler KR, Xu X, et al. AGEMAP: a gene expression database for aging in mice. PLoS Genet. 2007; 3:e201. https://doi.org/10.1371/journal.pgen.0030201 [PubMed]
- 20. Zhuang J, Zhang L, Dai S, Cui L, Guo C, Sloofman L, Yang J. Comparison of multi-tissue aging between human and mouse. Sci Rep. 2019; 9:6220. https://doi.org/10.1038/s41598-019-42485-3 [PubMed]
- 21. Börsch A, Ham DJ, Mittal N, Tintignac LA, Migliavacca E, Feige JN, Rüegg MA, Zavolan M. Molecular and phenotypic analysis of rodent models reveals conserved and species-specific modulators of human sarcopenia. Commun Biol. 2021; 4:194. https://doi.org/10.1038/s42003-021-01723-z [PubMed]
- 22. Shur NF, Creedon L, Skirrow S, Atherton PJ, MacDonald IA, Lund J, Greenhaff PL. Age-related changes in muscle architecture and metabolism in humans: The likely contribution of physical inactivity to age-related functional decline. Ageing Res Rev. 2021; 68:101344. https://doi.org/10.1016/j.arr.2021.101344 [PubMed]
- 23. Wang S, Lai X, Deng Y, Song Y. Correlation between mouse age and human age in anti-tumor research: Significance and method establishment. Life Sci. 2020; 242:117242. https://doi.org/10.1016/j.lfs.2019.117242 [PubMed]
- 24. Nishizawa H, Matsuda M, Yamada Y, Kawai K, Suzuki E, Makishima M, Kitamura T, Shimomura I. Musclin, a novel skeletal muscle-derived secretory factor. J Biol Chem. 2004; 279:19391–5. https://doi.org/10.1074/jbc.C400066200 [PubMed]
- 25. Harris MP, Zeng S, Zhu Z, Lira VA, Yu L, Hodgson-Zingman DM, Zingman LV. Myokine Musclin Is Critical for Exercise-Induced Cardiac Conditioning. Int J Mol Sci. 2023; 24:6525. https://doi.org/10.3390/ijms24076525 [PubMed]
- 26. Nishiguchi Y, Hata Y, Date R, Fujimoto D, Umemoto S, Kanki T, Yokoi H, Mori KP, Handa T, Watanabe-Takano H, Kanai Y, Yasoda A, Izumi Y, et al. Osteocrin, a bone-derived humoral factor, exerts a renoprotective role in ischemia-reperfusion injury in mice. Nephrol Dial Transplant. 2022; 37:444–53. https://doi.org/10.1093/ndt/gfab286 [PubMed]
- 27. Zhang X, Hu C, Yuan XP, Yuan YP, Song P, Kong CY, Teng T, Hu M, Xu SC, Ma ZG, Tang QZ. Osteocrin, a novel myokine, prevents diabetic cardiomyopathy via restoring proteasomal activity. Cell Death Dis. 2021; 12:624. https://doi.org/10.1038/s41419-021-03922-2 [PubMed]
- 28. Phang RJ, Ritchie RH, Hausenloy DJ, Lees JG, Lim SY. Cellular interplay between cardiomyocytes and non-myocytes in diabetic cardiomyopathy. Cardiovasc Res. 2023; 119:668–90. https://doi.org/10.1093/cvr/cvac049 [PubMed]
- 29. Boskovic S, Marín Juez R, Stamenkovic N, Radojkovic D, Stainier DY, Kojic S. The stress responsive gene ankrd1a is dynamically regulated during skeletal muscle development and upregulated following cardiac injury in border zone cardiomyocytes in adult zebrafish. Gene. 2021; 792:145725. https://doi.org/10.1016/j.gene.2021.145725 [PubMed]
- 30. Roth DM, Baddam P, Lin H, Vidal-García M, Aponte JD, De Souza ST, Godziuk D, Watson AE, Footz T, Schachter NF, Egan SE, Hallgrímsson B, Graf D, Voronova A. The Chromatin Regulator Ankrd11 Controls Palate and Cranial Bone Development. Front Cell Dev Biol. 2021; 9:645386. https://doi.org/10.3389/fcell.2021.645386 [PubMed]
- 31. Zhang Y, Zhou L, Fu Q, Liu Z. ANKRD1 activates the Wnt signaling pathway by modulating CAV3 expression and thus promotes BMSC osteogenic differentiation and bone formation in ovariectomized mice. Biochim Biophys Acta Mol Basis Dis. 2023; 1869:166693. https://doi.org/10.1016/j.bbadis.2023.166693 [PubMed]
- 32. Digilio MC, Calcagni G, Gnazzo M, Versacci P, Dentici ML, Capolino R, Sinibaldi L, Baban A, Putotto C, Alfieri P, Unolt M, Lepri FR, Alesi V, et al. Congenital heart defects in molecularly confirmed KBG syndrome patients. Am J Med Genet A. 2022; 188:1149–59. https://doi.org/10.1002/ajmg.a.62632 [PubMed]
- 33. Murphy NP, Lubbers ER, Mohler PJ. Advancing our understanding of AnkRD1 in cardiac development and disease. Cardiovasc Res. 2020; 116:1402–4. https://doi.org/10.1093/cvr/cvaa063 [PubMed]
- 34. Kirschbaum T, Pourrajabi S, Zocher I, Schwendinger J, Heim V, Röschenthaler F, Kirschbaum V, Zachau HG. The 3' part of the immunoglobulin kappa locus of the mouse. Eur J Immunol. 1998; 28:1458–66. https://doi.org/10.1002/(SICI)1521-4141(199805)28:05<1458::AID-IMMU1458>3.0.CO;2-5 [PubMed]
- 35. Phillips-Quagliata JM. Structural correlates of mouse IgA allotypes. Immunogenetics. 2010; 62:1–13. https://doi.org/10.1007/s00251-009-0414-7 [PubMed]
- 36. Martin RM, Silva A, Lew AM. The Igh-1 sequence of the non-obese diabetic (NOD) mouse assigns it to the IgG2c isotype. Immunogenetics. 1997; 46:167–8. https://doi.org/10.1007/s002510050258 [PubMed]
- 37. Kataoka T, Yamawaki-Kataoka Y, Yamagishi H, Honjo T. Cloning immunoglobulin gamma 2b chain gene of mouse: characterization and partial sequence determination. Proc Natl Acad Sci USA. 1979; 76:4240–4. https://doi.org/10.1073/pnas.76.9.4240 [PubMed]
- 38. Solin ML, Kaartinen M. Immunoglobulin constant kappa gene alleles in twelve strains of mice. Immunogenetics. 1993; 37:401–7. https://doi.org/10.1007/BF00222463 [PubMed]
- 39. Erlandsson L, Akerblad P, Vingsbo-Lundberg C, Kallberg E, Lycke N, Leanderson T. Joining chain-expressing and -nonexpressing B cell populations in the mouse. J Exp Med. 2001; 194:557–70. https://doi.org/10.1084/jem.194.5.557 [PubMed]
- 40. Eto M, Karginov A, Brautigan DL. A novel phosphoprotein inhibitor of protein type-1 phosphatase holoenzymes. Biochemistry. 1999; 38:16952–7. https://doi.org/10.1021/bi992030o [PubMed]
- 41. Lindner C, Thomsen I, Wahl B, Ugur M, Sethi MK, Friedrichsen M, Smoczek A, Ott S, Baumann U, Suerbaum S, Schreiber S, Bleich A, Gaboriau-Routhiau V, et al. Diversification of memory B cells drives the continuous adaptation of secretory antibodies to gut microbiota. Nat Immunol. 2015; 16:880–8. https://doi.org/10.1038/ni.3213 [PubMed]
- 42. Krayem I, Sohrabi Y, Javorková E, Volkova V, Strnad H, Havelková H, Vojtíšková J, Aidarova A, Holáň V, Demant P, Lipoldová M. Genetic Influence on Frequencies of Myeloid-Derived Cell Subpopulations in Mouse. Front Immunol. 2022; 12:760881. https://doi.org/10.3389/fimmu.2021.760881 [PubMed]
- 43. Liu K, Zhang J, Xiao Y, Yang A, Song X, Li Y, Chen Y, Hughes TR, Min J. Structural insights into DNA recognition by the BEN domain of the transcription factor BANP. J Biol Chem. 2023; 299:104734. https://doi.org/10.1016/j.jbc.2023.104734 [PubMed]
- 44. Kogo H, Tsutsumi M, Inagaki H, Ohye T, Kiyonari H, Kurahashi H. HORMAD2 is essential for synapsis surveillance during meiotic prophase via the recruitment of ATR activity. Genes Cells. 2012; 17:897–912. https://doi.org/10.1111/gtc.12005 [PubMed]
- 45. Zhu P, Wang Y, Wu J, Huang G, Liu B, Ye B, Du Y, Gao G, Tian Y, He L, Fan Z. LncBRM initiates YAP1 signalling activation to drive self-renewal of liver cancer stem cells. Nat Commun. 2016; 7:13608. https://doi.org/10.1038/ncomms13608 [PubMed]
- 46. Fasci D, van Ingen H, Scheltema RA, Heck AJ. Histone Interaction Landscapes Visualized by Crosslinking Mass Spectrometry in Intact Cell Nuclei. Mol Cell Proteomics. 2018; 17:2018–33. https://doi.org/10.1074/mcp.RA118.000924 [PubMed]
- 47. Zou L, Xia PF, Chen L, Hou YY. XIST knockdown suppresses vascular smooth muscle cell proliferation and induces apoptosis by regulating miR-1264/WNT5A/β-catenin signaling in aneurysm. Biosci Rep. 2021; 41:BSR20201810. https://doi.org/10.1042/BSR20201810 [PubMed]
- 48. Yang F, Zhang J, Zhao Z, Liu Y, Zhao Z, Fu K, Li B, Jin J. Artemisinin suppresses aerobic glycolysis in thyroid cancer cells by downregulating HIF-1a, which is increased by the XIST/miR-93/HIF-1a pathway. PLoS One. 2023; 18:e0284242. https://doi.org/10.1371/journal.pone.0284242 [PubMed]
- 49. Yang LG, Cao MZ, Zhang J, Li XY, Sun QL. LncRNA XIST modulates HIF-1A/AXL signaling pathway by inhibiting miR-93-5p in colorectal cancer. Mol Genet Genomic Med. 2020; 8:e1112. https://doi.org/10.1002/mgg3.1112 [PubMed]
- 50. Liu H, Deng H, Zhao Y, Li C, Liang Y. LncRNA XIST/miR-34a axis modulates the cell proliferation and tumor growth of thyroid cancer through MET-PI3K-AKT signaling. J Exp Clin Cancer Res. 2018; 37:279. https://doi.org/10.1186/s13046-018-0950-9 [PubMed]
- 51. Norgett EE, Hatsell SJ, Carvajal-Huerta L, Cabezas JC, Common J, Purkis PE, Whittock N, Leigh IM, Stevens HP, Kelsell DP. Recessive mutation in desmoplakin disrupts desmoplakin-intermediate filament interactions and causes dilated cardiomyopathy, woolly hair and keratoderma. Hum Mol Genet. 2000; 9:2761–6. https://doi.org/10.1093/hmg/9.18.2761 [PubMed]
- 52. Liu L, McKeehan WL. Sequence analysis of LRPPRC and its SEC1 domain interaction partners suggests roles in cytoskeletal organization, vesicular trafficking, nucleocytosolic shuttling, and chromosome activity. Genomics. 2002; 79:124–36. https://doi.org/10.1006/geno.2001.6679 [PubMed]
- 53. Wang Y, Liu Y, Zhang M, Lv L, Zhang X, Zhang P, Zhou Y. LRRC15 promotes osteogenic differentiation of mesenchymal stem cells by modulating p65 cytoplasmic/nuclear translocation. Stem Cell Res Ther. 2018; 9:65. https://doi.org/10.1186/s13287-018-0809-1 [PubMed]
- 54. Elias LA, Wang DD, Kriegstein AR. Gap junction adhesion is necessary for radial migration in the neocortex. Nature. 2007; 448:901–7. https://doi.org/10.1038/nature06063 [PubMed]
- 55. Moore KB, Mood K, Daar IO, Moody SA. Morphogenetic movements underlying eye field formation require interactions between the FGF and ephrinB1 signaling pathways. Dev Cell. 2004; 6:55–67. https://doi.org/10.1016/s1534-5807(03)00395-2 [PubMed]
- 56. Matsumura H, Mohri Y, Binh NT, Morinaga H, Fukuda M, Ito M, Kurata S, Hoeijmakers J, Nishimura EK. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science. 2016; 351:aad4395. https://doi.org/10.1126/science.aad4395 [PubMed]
- 57. Ihrie RA, Marques MR, Nguyen BT, Horner JS, Papazoglu C, Bronson RT, Mills AA, Attardi LD. Perp is a p63-regulated gene essential for epithelial integrity. Cell. 2005; 120:843–56. https://doi.org/10.1016/j.cell.2005.01.008 [PubMed]
- 58. Müller FB, Huber M, Kinaciyan T, Hausser I, Schaffrath C, Krieg T, Hohl D, Korge BP, Arin MJ. A human keratin 10 knockout causes recessive epidermolytic hyperkeratosis. Hum Mol Genet. 2006; 15:1133–41. https://doi.org/10.1093/hmg/ddl028 [PubMed]
- 59. Oberbeck N, Pham VC, Webster JD, Reja R, Huang CS, Zhang Y, Roose-Girma M, Warming S, Li Q, Birnberg A, Wong W, Sandoval W, Kőműves LG, et al. The RIPK4-IRF6 signalling axis safeguards epidermal differentiation and barrier function. Nature. 2019; 574:249–53. https://doi.org/10.1038/s41586-019-1615-3 [PubMed]
- 60. Tong Q, Dalgin G, Xu H, Ting CN, Leiden JM, Hotamisligil GS. Function of GATA transcription factors in preadipocyte-adipocyte transition. Science. 2000; 290:134–8. https://doi.org/10.1126/science.290.5489.134 [PubMed]
- 61. Li Y, Lin X, Kilani RT, Jones JC, Ghahary A. 14-3-3 sigma isoform interacts with the cytoplasmic domain of the transmembrane BP180 in keratinocytes. J Cell Physiol. 2007; 212:675–81. https://doi.org/10.1002/jcp.21064 [PubMed]
- 62. Wise A, Foord SM, Fraser NJ, Barnes AA, Elshourbagy N, Eilert M, Ignar DM, Murdock PR, Steplewski K, Green A, Brown AJ, Dowell SJ, Szekeres PG, et al. Molecular identification of high and low affinity receptors for nicotinic acid. J Biol Chem. 2003; 278:9869–74. https://doi.org/10.1074/jbc.M210695200 [PubMed]
- 63. Migeon BR, Lee CH, Chowdhury AK, Carpenter H. Species differences in TSIX/Tsix reveal the roles of these genes in X-chromosome inactivation. Am J Hum Genet. 2002; 71:286–93. https://doi.org/10.1086/341605 [PubMed]
- 64. Vaughan KT, Weber FE, Ried T, Ward DC, Reinach FC, Fischman DA. Human myosin-binding protein H (MyBP-H): complete primary sequence, genomic organization, and chromosomal localization. Genomics. 1993; 16:34–40. https://doi.org/10.1006/geno.1993.1136 [PubMed]
- 65. Herrer I, Roselló-Lletí E, Rivera M, Molina-Navarro MM, Tarazón E, Ortega A, Martínez-Dolz L, Triviño JC, Lago F, González-Juanatey JR, Bertomeu V, Montero JA, Portolés M. RNA-sequencing analysis reveals new alterations in cardiomyocyte cytoskeletal genes in patients with heart failure. Lab Invest. 2014; 94:645–53. https://doi.org/10.1038/labinvest.2014.54 [PubMed]
- 66. North KN, Yang N, Wattanasirichaigoon D, Mills M, Easteal S, Beggs AH. A common nonsense mutation results in alpha-actinin-3 deficiency in the general population. Nat Genet. 1999; 21:353–4. https://doi.org/10.1038/7675 [PubMed]
- 67. Mansilla F, Dominguez CA, Yeadon JE, Corydon TJ, Burden SJ, Knudsen CR. Translation elongation factor eEF1A binds to a novel myosin binding protein-C-like protein. J Cell Biochem. 2008; 105:847–58. https://doi.org/10.1002/jcb.21880 [PubMed]
- 68. Kim SH, Han SY, Azam T, Yoon DY, Dinarello CA. Interleukin-32: a cytokine and inducer of TNFalpha. Immunity. 2005; 22:131–42. https://doi.org/10.1016/j.immuni.2004.12.003 [PubMed]
- 69. Bambouskova M, Gorvel L, Lampropoulou V, Sergushichev A, Loginicheva E, Johnson K, Korenfeld D, Mathyer ME, Kim H, Huang LH, Duncan D, Bregman H, Keskin A, et al. Electrophilic properties of itaconate and derivatives regulate the IκBζ-ATF3 inflammatory axis. Nature. 2018; 556:501–4. https://doi.org/10.1038/s41586-018-0052-z [PubMed]
- 70. Hoetzenecker W, Echtenacher B, Guenova E, Hoetzenecker K, Woelbing F, Brück J, Teske A, Valtcheva N, Fuchs K, Kneilling M, Park JH, Kim KH, Kim KW, et al. ROS-induced ATF3 causes susceptibility to secondary infections during sepsis-associated immunosuppression. Nat Med. 2011; 18:128–34. https://doi.org/10.1038/nm.2557 [PubMed]
- 71. Gilchrist M, Thorsson V, Li B, Rust AG, Korb M, Roach JC, Kennedy K, Hai T, Bolouri H, Aderem A. Systems biology approaches identify ATF3 as a negative regulator of Toll-like receptor 4. Nature. 2006; 441:173–8. https://doi.org/10.1038/nature04768 [PubMed]
- 72. Fay A, Garcia Y, Margeta M, Maharjan S, Jürgensen C, Briceño J, Garcia M, Yin S, Bassaganyas L, McMahon T, Hou YM, Fu YH, Ptáček LJ. A Mitochondrial tRNA Mutation Causes Axonal CMT in a Large Venezuelan Family. Ann Neurol. 2020; 88:830–42. https://doi.org/10.1002/ana.25854 [PubMed]
- 73. Ling J, Roy H, Qin D, Rubio MA, Alfonzo JD, Fredrick K, Ibba M. Pathogenic mechanism of a human mitochondrial tRNAPhe mutation associated with myoclonic epilepsy with ragged red fibers syndrome. Proc Natl Acad Sci USA. 2007; 104:15299–304. https://doi.org/10.1073/pnas.0704441104 [PubMed]
- 74. Ishii KA, Fumoto T, Iwai K, Takeshita S, Ito M, Shimohata N, Aburatani H, Taketani S, Lelliott CJ, Vidal-Puig A, Ikeda K. Coordination of PGC-1beta and iron uptake in mitochondrial biogenesis and osteoclast activation. Nat Med. 2009; 15:259–66. https://doi.org/10.1038/nm.1910 [PubMed]
- 75. Yeo HS, Lim JY, Ahn NY. Effects of Aging on Angiogenic and Muscle Growth-Related Factors in Naturally Aged Rat Skeletal Muscles. Ann Geriatr Med Res. 2020; 24:305–12. https://doi.org/10.4235/agmr.20.0077 [PubMed]
- 76. Rhoads RP, Flann KL, Cardinal TR, Rathbone CR, Liu X, Allen RE. Satellite cells isolated from aged or dystrophic muscle exhibit a reduced capacity to promote angiogenesis in vitro. Biochem Biophys Res Commun. 2013; 440:399–404. https://doi.org/10.1016/j.bbrc.2013.09.085 [PubMed]
- 77. Kubo M, Li TS, Kurazumi H, Takemoto Y, Ohshima M, Murata T, Katsura S, Morikage N, Furutani A, Hamano K. Hypoxic preconditioning enhances angiogenic potential of bone marrow cells with aging-related functional impairment. Circ J. 2012; 76:986–94. https://doi.org/10.1253/circj.cj-11-0605 [PubMed]
- 78. Huang W, Yi S, Zhao L. Genetic Features of Young and Aged Animals After Peripheral Nerve Injury: Implications for Diminished Regeneration Capacity. Cell Mol Neurobiol. 2023; 43:4363–75. https://doi.org/10.1007/s10571-023-01431-8 [PubMed]
- 79. Fuentes-Flores A, Geronimo-Olvera C, Girardi K, Necuñir-Ibarra D, Patel SK, Bons J, Wright MC, Geschwind D, Hoke A, Gomez-Sanchez JA, Schilling B, Rebolledo DL, Campisi J, Court FA. Senescent Schwann cells induced by aging and chronic denervation impair axonal regeneration following peripheral nerve injury. EMBO Mol Med. 2023; 15:e17907. https://doi.org/10.15252/emmm.202317907 [PubMed]
- 80. Maita KC, Garcia JP, Avila FR, Torres-Guzman RA, Ho O, Chini CC, Chini EN, Forte AJ. Evaluation of the Aging Effect on Peripheral Nerve Regeneration: A Systematic Review. J Surg Res. 2023; 288:329–40. https://doi.org/10.1016/j.jss.2023.03.017 [PubMed]
- 81. Frolkis VV, Tanin SA, Gorban YN. Age-related changes in axonal transport. Exp Gerontol. 1997; 32:441–50. https://doi.org/10.1016/s0531-5565(96)00168-4 [PubMed]
- 82. Hong X, Campanario S, Ramírez-Pardo I, Grima-Terrén M, Isern J, Muñoz-Cánoves P. Stem cell aging in the skeletal muscle: The importance of communication. Ageing Res Rev. 2022; 73:101528. https://doi.org/10.1016/j.arr.2021.101528 [PubMed]
- 83. He GJ, Galvis J, Cheung TH, Le Grand F. Single-Cell Transcriptomic Analysis of Mononuclear Cell Populations in Skeletal Muscle. Methods Mol Biol. 2023; 2640:369–95. https://doi.org/10.1007/978-1-0716-3036-5_26 [PubMed]
- 84. Matsuda Y, Uchimura A, Satoh Y, Kato N, Toshishige M, Kajimura J, Hamasaki K, Yoshida K, Hayashi T, Noda A, Tanabe O. Spectra and characteristics of somatic mutations induced by ionizing radiation in hematopoietic stem cells. Proc Natl Acad Sci USA. 2023; 120:e2216550120. https://doi.org/10.1073/pnas.2216550120 [PubMed]
- 85. Yue F, Cheng Y, Breschi A, Vierstra J, Wu W, Ryba T, Sandstrom R, Ma Z, Davis C, Pope BD, Shen Y, Pervouchine DD, Djebali S, et al., and Mouse ENCODE Consortium. A comparative encyclopedia of DNA elements in the mouse genome. Nature. 2014; 515:355–64. https://doi.org/10.1038/nature13992 [PubMed]
- 86. Mudge JM, Harrow J. Creating reference gene annotation for the mouse C57BL6/J genome assembly. Mamm Genome. 2015; 26:366–78. https://doi.org/10.1007/s00335-015-9583-x [PubMed]
- 87. Singh P, Schimenti JC, Bolcun-Filas E. A mouse geneticist's practical guide to CRISPR applications. Genetics. 2015; 199:1–15. https://doi.org/10.1534/genetics.114.169771 [PubMed]
- 88. Breschi A, Gingeras TR, Guigó R. Comparative transcriptomics in human and mouse. Nat Rev Genet. 2017; 18:425–40. https://doi.org/10.1038/nrg.2017.19 [PubMed]
- 89. Grevendonk L, Connell NJ, McCrum C, Fealy CE, Bilet L, Bruls YM, Mevenkamp J, Schrauwen-Hinderling VB, Jörgensen JA, Moonen-Kornips E, Schaart G, Havekes B, de Vogel-van den Bosch J, et al. Impact of aging and exercise on skeletal muscle mitochondrial capacity, energy metabolism, and physical function. Nat Commun. 2021; 12:4773. https://doi.org/10.1038/s41467-021-24956-2 [PubMed]
- 90. Fuller KN, Thyfault JP. Barriers in translating preclinical rodent exercise metabolism findings to human health. J Appl Physiol (1985). 2021; 130:182–92. https://doi.org/10.1152/japplphysiol.00683.2020 [PubMed]
- 91. Lofaro FD, Cisterna B, Lacavalla MA, Boschi F, Malatesta M, Quaglino D, Zancanaro C, Boraldi F. Age-Related Changes in the Matrisome of the Mouse Skeletal Muscle. Int J Mol Sci. 2021; 22:10564. https://doi.org/10.3390/ijms221910564 [PubMed]
- 92. Cosgrove BD, Gilbert PM, Porpiglia E, Mourkioti F, Lee SP, Corbel SY, Llewellyn ME, Delp SL, Blau HM. Rejuvenation of the muscle stem cell population restores strength to injured aged muscles. Nat Med. 2014; 20:255–64. https://doi.org/10.1038/nm.3464 [PubMed]
- 93. Sinha M, Jang YC, Oh J, Khong D, Wu EY, Manohar R, Miller C, Regalado SG, Loffredo FS, Pancoast JR, Hirshman MF, Lebowitz J, Shadrach JL, et al. Restoring systemic GDF11 levels reverses age-related dysfunction in mouse skeletal muscle. Science. 2014; 344:649–52. https://doi.org/10.1126/science.1251152 [PubMed]
- 94. Hicks MR, Saleh KK, Clock B, Gibbs DE, Yang M, Younesi S, Gane L, Gutierrez-Garcia V, Xi H, Pyle AD. Regenerating human skeletal muscle forms an emerging niche in vivo to support PAX7 cells. Nat Cell Biol. 2023; 25:1758–73. https://doi.org/10.1038/s41556-023-01271-0 [PubMed]
- 95. Kragstrup TW, Kjaer M, Mackey AL. Structural, biochemical, cellular, and functional changes in skeletal muscle extracellular matrix with aging. Scand J Med Sci Sports. 2011; 21:749–57. https://doi.org/10.1111/j.1600-0838.2011.01377.x [PubMed]
- 96. Hynes RO. The extracellular matrix: not just pretty fibrils. Science. 2009; 326:1216–9. https://doi.org/10.1126/science.1176009 [PubMed]
- 97. Stearns-Reider KM, D'Amore A, Beezhold K, Rothrauff B, Cavalli L, Wagner WR, Vorp DA, Tsamis A, Shinde S, Zhang C, Barchowsky A, Rando TA, Tuan RS, Ambrosio F. Aging of the skeletal muscle extracellular matrix drives a stem cell fibrogenic conversion. Aging Cell. 2017; 16:518–28. https://doi.org/10.1111/acel.12578 [PubMed]
- 98. Perez K, Ciotlos S, McGirr J, Limbad C, Doi R, Nederveen JP, Nilsson MI, Winer DA, Evans W, Tarnopolsky M, Campisi J, Melov S. Single nuclei profiling identifies cell specific markers of skeletal muscle aging, frailty, and senescence. Aging (Albany NY). 2022; 14:9393–422. https://doi.org/10.18632/aging.204435 [PubMed]
- 99. Bodine SC, Sinha I, Sweeney HL. Mechanisms of Skeletal Muscle Atrophy and Molecular Circuitry of Stem Cell Fate in Skeletal Muscle Regeneration and Aging. J Gerontol A Biol Sci Med Sci. 2023 (Suppl 1); 78:14–8. https://doi.org/10.1093/gerona/glad023 [PubMed]
- 100. Ascenzi F, Barberi L, Dobrowolny G, Villa Nova Bacurau A, Nicoletti C, Rizzuto E, Rosenthal N, Scicchitano BM, Musarò A. Effects of IGF-1 isoforms on muscle growth and sarcopenia. Aging Cell. 2019; 18:e12954. https://doi.org/10.1111/acel.12954 [PubMed]
- 101. Nandeesh BN, Mahadevan A, Santosh V, Yasha TC, Shankar SK. Acute aortic dissection presenting as painful paraplegia. Clin Neurol Neurosurg. 2007; 109:531–4. https://doi.org/10.1016/j.clineuro.2007.03.010 [PubMed]
- 102. Gameiro PA, Yang J, Metelo AM, Pérez-Carro R, Baker R, Wang Z, Arreola A, Rathmell WK, Olumi A, López-Larrubia P, Stephanopoulos G, Iliopoulos O. In vivo HIF-mediated reductive carboxylation is regulated by citrate levels and sensitizes VHL-deficient cells to glutamine deprivation. Cell Metab. 2013; 17:372–85. https://doi.org/10.1016/j.cmet.2013.02.002 [PubMed]
- 103. McClelland RL, Chung H, Detrano R, Post W, Kronmal RA. Distribution of coronary artery calcium by race, gender, and age: results from the Multi-Ethnic Study of Atherosclerosis (MESA). Circulation. 2006; 113:30–7. https://doi.org/10.1161/CIRCULATIONAHA.105.580696 [PubMed]
- 104. Hu MC, Lee DF, Xia W, Golfman LS, Ou-Yang F, Yang JY, Zou Y, Bao S, Hanada N, Saso H, Kobayashi R, Hung MC. IkappaB kinase promotes tumorigenesis through inhibition of forkhead FOXO3a. Cell. 2004; 117:225–37. https://doi.org/10.1016/s0092-8674(04)00302-2 [PubMed]
- 105. Schulz S, Wong RJ, Kalish FS, Zhao H, Jang KY, Vreman HJ, Stevenson DK. Effect of light exposure on metalloporphyrin-treated newborn mice. Pediatr Res. 2012; 72:161–8. https://doi.org/10.1038/pr.2012.62 [PubMed]
- 106. Alisky JM, Chertkova EL, Iczkowski KA. Drug interactions and pharmacogenetic reactions are the basis for chloroquine and mefloquine-induced psychosis. Med Hypotheses. 2006; 67:1090–4. https://doi.org/10.1016/j.mehy.2006.01.059 [PubMed]
- 107. Chia R, Achilli F, Festing MF, Fisher EM. The origins and uses of mouse outbred stocks. Nat Genet. 2005; 37:1181–6. https://doi.org/10.1038/ng1665 [PubMed]
- 108. Kho AT, Kang PB, Kohane IS, Kunkel LM. Transcriptome-scale similarities between mice and human skeletal muscles with normal and myopathic phenotypes. BMC Musculoskelet Disord. 2006; 7:23. https://doi.org/10.1186/1471-2474-7-23 [PubMed]
- 109. Jang H, Bhasin S, Guarneri T, Serra C, Schneider M, Lee MJ, Guo W, Fried SK, Pencina K, Jasuja R. The Effects of a Single Developmentally Entrained Pulse of Testosterone in Female Neonatal Mice on Reproductive and Metabolic Functions in Adult Life. Endocrinology. 2015; 156:3737–46. https://doi.org/10.1210/EN.2015-1117 [PubMed]
- 110. Endo Y, Samandari M, Karvar M, Mostafavi A, Quint J, Rinoldi C, Yazdi IK, Swieszkowski W, Mauney J, Agarwal S, Tamayol A, Sinha I. Aerobic exercise and scaffolds with hierarchical porosity synergistically promote functional recovery post volumetric muscle loss. Biomaterials. 2023; 296:122058. https://doi.org/10.1016/j.biomaterials.2023.122058 [PubMed]
- 111. Dobin A, Davis CA, Schlesinger F, Drenkow J, Zaleski C, Jha S, Batut P, Chaisson M, Gingeras TR. STAR: ultrafast universal RNA-seq aligner. Bioinformatics. 2013; 29:15–21. https://doi.org/10.1093/bioinformatics/bts635 [PubMed]
- 112. Liao Y, Smyth GK, Shi W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics. 2014; 30:923–30. https://doi.org/10.1093/bioinformatics/btt656 [PubMed]
- 113. Chen Y, Lun AT, Smyth GK. From reads to genes to pathways: differential expression analysis of RNA-Seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Res. 2016; 5:1438. https://doi.org/10.12688/f1000research.8987.2 [PubMed]
- 114. Endo Y, Zhang Y, Olumi S, Karvar M, Argawal S, Neppl RL, Sinha I. Exercise-induced gene expression changes in skeletal muscle of old mice. Genomics. 2021; 113:2965–76. https://doi.org/10.1016/j.ygeno.2021.06.035 [PubMed]
- 115. Kuleshov MV, Jones MR, Rouillard AD, Fernandez NF, Duan Q, Wang Z, Koplev S, Jenkins SL, Jagodnik KM, Lachmann A, McDermott MG, Monteiro CD, Gundersen GW, Ma'ayan A. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016; 44:W90–97. https://doi.org/10.1093/nar/gkw377 [PubMed]