



Introduction
Heart failure (HF) is the severe or terminal stage of various heart diseases and is characterized by high morbidity, mortality, and hospitalization rates; poor quality of life; and high medical costs [1]. The data show that the global incidence of HF ranges from 1% to 3%. Due to population aging, the treatment and prognosis of ischemic heart disease have improved, and effective evidence-based treatment has prolonged the survival of patients with HF. However, the incidence of HF has continued to increase, ranging from 1/1000 person-years to 20/1000 person-years, and the prevalence in different regional populations also differs. The incidence of HF in European and American populations ranges from 2/1000 person-years to 3/1000 person-years, and the incidence of HF is positively correlated with age [2]. Heart failure can lead to reduced pumping function of the heart muscle and reduced blood flow throughout the body, resulting in insufficient blood supply to the brain. In fact, abnormal cerebral hemodynamics may lead to a lack of glucose and oxygen in the brain, which in turn leads to a series of adverse biochemical events, ultimately leading to metabolic and tissue damage, which is a major cause of structural changes in the brain [3]. The structural changes in the cerebral cortex caused by this phenomenon are also related mainly to cognitive dysfunction, such as vascular dementia and Alzheimer’s disease. Studies have shown that patients with coronary heart disease exhibit a wide range of gray matter density decreases, while patients with heart failure mainly exhibit a significant decrease in gray matter density in the posterior, middle and precuneus regions of the cingulate gyrus [4]. Moreover, the cortical thickness of the frontal, parietal, temporal and occipital lobes is reduced in patients with heart failure, which mainly controls autonomic, cognitive, emotional, language and visual functions. With the atrophy of these functional cortical areas, patients also exhibit corresponding neurological dysfunction [5]. Brain structure is closely related to heart failure, and the existing research is limited mainly to the study of brain structure changes caused by hemodynamics; however, there is no clear targeted research on whether there is a genetic link between heart failure and cerebral cortex structure.
Randomized controlled trials (RCTs) are not implemented clinically due to various limiting factors, and observational experimental methods can lead to biased study results due to confounding factors and reverse causation, resulting in relatively low credibility. Mendelian randomization (MR) involves an analysis of genetic variables that follow Mendel’s law of inheritance and exploits single nucleotide polymorphisms (SNPs) as instrumental variables (IVs) to infer the causality of an observed association between a modifiable exposure and a clinically relevant outcome [6]. Alleles are randomly separated during meiosis, so MR can reduce the bias caused by confounding factors [7]. In addition, since genetic variation occurs before disease and the order of the two cannot be reversed, MR can also avoid the interference of reverse causality [7].
This study was based on a large sample genome-wide association study (GWAS; GE-Nanowide Association Study). SNP sites published in the GWAS database were used as instrumental variables of genetic variation to explore the causal relationship between HF and cerebral cortex structure through a two-sample MR research method.
Materials and Methods
Study design
In this study, heart failure was used as an exposure factor, and single nucleotide polymorphisms (SNPs) that were significantly correlated with heart failure were used as instrumental variables (IVs). The cerebral cortex structure was selected as the outcome. The TwoSampleMR package in R was used to conduct causal association analysis, and the Cochran Q heterogeneity test, pleiotropy test and sensitivity analysis were performed to verify the reliability of the results. MR analysis relies on three assumptions, as shown in Figure 1 [8]: (1) IVs are strongly correlated with exposure variables, (2) IVs are not associated with confounding factors affecting exposure outcomes, and (3) IVs affect outcome factors only through exposure factors.
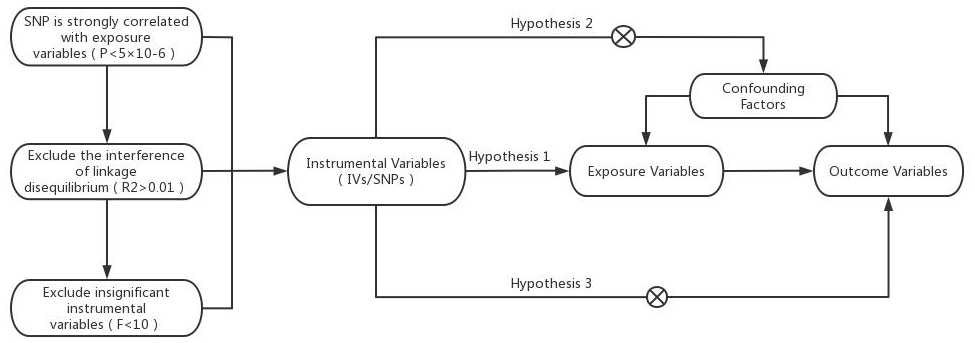
Figure 1. Model of the Mendelian randomization analysis.
Data sources
We searched for GWAS data on HFs and the cerebral cortex on the website ‘https://gwas.mrcieu.ac.uk/datasets’. The data for HF (ebi-a-GCST009541) were derived from the GWAS statistical results published in 2020 and included a sample of 977,323 people, 47,309 cases and 930,014 controls, and 7,773,021 SNPs. In this study, GWAS data related to cortical structure were obtained from the ENIGMA study, a genome-wide association meta-analysis of brain magnetic resonance imaging data from 51,665 people; the surface area, average thickness, and 34 known functional regions of the entire cortex were analyzed. The HF patients with cerebral cortex structure data included in this study were mostly European.
Instrumental variables
We obtained HF-related genetic data from the website ‘https://gwas.mrcieu.ac.uk/datasets’ using R software. To avoid analytical bias caused by linkage disequilibrium (LD) between SNPs, we used all SNPs that were significant (P < 5×10-8) and predicted significant genome-wide exposure. SNPs with an R2 value >0.001 and a base physical distance less than 10,000 kb were removed, and SNPs with the smallest P-values were retained. Candidate SNPs were matched with GWAS data for the outcome variable based on the chromosome and location. To further evaluate weak IV bias, the F statistic was used to calculate the power of the IV. When the IVs with an F value <10 were eliminated, the specific calculation formula was F=β2/SE2, where β2 is the effect value of the β allele and SE is the standard error. Since only 12 significant and independent SNPs with genome-wide significance remained after screening, we used a higher P-value (P < 5×10-6) to obtain SNPs predicting HF as the final IV included in the study.
MR and sensitivity analysis
The analysis methods used in this study were mainly the inverse-variance weighted (IVW) method in the TwoSampleMR package [9, 10], MR-Egger regression [11] and the weighted median (WME) method [12].
The heterogeneity test tests the difference between various IVs. If P was >0.05, there was no heterogeneity. In this study, the P-value of the Cochran Q test was used to assess heterogeneity. A P-value <0.05 indicated heterogeneity. In contrast, P>0.05 indicated no heterogeneity. The pleiotropy test [13] verifies the reliability of MR analysis results and is often performed via the MR-PRESSO test and the intercept term of the MR-Egger regression method. P>0.05 was considered to indicate no horizontal pleiotropy; if pleiotropy was indicated, the MR analysis results were not reliable. The “leave-one-out” method [14] was adopted to test the sensitivity. The principle is to gradually eliminate the results of a single SNP to determine whether the results are outliers and to observe the stability and reliability of the results after the removal of each SNP.
Availability of data and materials
All data are publicly available.
Consent for publication
Written informed consent for publication was obtained from all participants.
Results
Instrumental variables
In this study, HF was taken as an exposure factor, R software was used to screen SNPs with genome-wide significance according to the screening criteria, and a total of 52 SNPs were included as IVs. Table 1 shows the 15 most significant SNPs, and the remaining complete SNP data are shown in Supplementary Table 1.
Table 1. Basic information of the SNPs associated with HF.
| SNPs | CHR | POS | Other allele | Effect allele | β | SE | P | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs55751848 | 1 | 57018257 | C | G | -0.0425 | 0.0089 | 1.79E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs593467 | 1 | 70584460 | G | A | -0.0548 | 0.0118 | 3.42E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs660240 | 1 | 109817838 | T | C | 0.0611 | 0.0097 | 3.00E-10 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs35054810 | 1 | 222722282 | G | A | 0.0725 | 0.0143 | 3.98E-07 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs7559452 | 2 | 3885011 | A | G | 0.0468 | 0.0102 | 4.47E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs17496249 | 2 | 37102249 | A | G | -0.0372 | 0.0079 | 2.49E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs12477245 | 2 | 107584422 | C | T | 0.1192 | 0.0236 | 4.40E-07 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs7369998 | 2 | 125815568 | G | A | -0.059 | 0.0126 | 2.83E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs72844714 | 2 | 133386122 | C | A | 0.0559 | 0.0121 | 3.84E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs80087882 | 2 | 201379864 | G | A | 0.0609 | 0.0125 | 1.10E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs4376020 | 3 | 5397743 | T | A | -0.0612 | 0.0123 | 6.50E-07 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs9815816 | 3 | 85930582 | T | C | 0.0479 | 0.0099 | 1.31E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs10938398 | 4 | 45186139 | G | A | 0.0389 | 0.008 | 1.16E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs11722972 | 4 | 69897984 | T | G | -0.0519 | 0.0114 | 5.30E-06 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| rs2634071 | 4 | 111669220 | T | C | -0.0923 | 0.0101 | 6.33E-20 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SNPs, single nucleotide polymorphisms; CHR, chromosome; POS, location; EA, effector allele; OA, noneffector allele. β, standard error. SE, standard error of beta. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
MR analysis of HF and cortical thickness
In this study, IVW regression, MR-Egger regression and WME analysis in the TwoSampleMR package were used to perform MR analysis of HF and cerebral cortex thickness. Table 2 shows the results of the MR analysis, and the scatter plot is shown in Figure 2. IVW analysis revealed β=-0.0068, SE=0.0033, P=0.0402 and an OR (95% CI)=0.9932 (0.9868-1.00); MR-Egger analysis revealed β=-0.004, SE=0.0114, P=0.7273, and an OR (95% CI)=0.996 (0.974-1.0185). WME analysis revealed β=-0.005, SE=0.0041, P=0.2223, and OR (95% CI)=0.995 (0.9871-1.003). Although the results of the MR-Egger and WME analyses showed that HF had no significant effect on the thickness of the cerebral cortex, the results of the IVW analysis showed P=0.0402. In addition, the β values of the IVW, MR-Egger and WME analyses were in the same direction, and the IVW results prevailed. Moreover, we used the MR-Egger intercept to verify the presence of pleiotropy in this study. The results showed an Egger’s intercept value of -15.6× 10-5, which is infinitely close to 0, SE = 6.04× 10-4, P = 0.7974 and the global test pval=0.078, indicating that horizontal pleiotropy did not exist. There was no multi-effect interference in the MR results. Therefore, these results suggest that HF has a significant effect on the thickness of the cerebral cortex, that there is a causal relationship between the two, and that the thickness of the cerebral cortex is negatively correlated with the incidence of HF.
Table 2. MR results of cortical thickness in HF patients.
| MR method | β | SE | OR (95% CI) | P | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IVW | -0.0068 | 0.0033 | 0.9932 (0.9868-1.00) | 0.0402 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MR–Egger | -0.004 | 0.0114 | 0.9960 (0.974-1.0185) | 0.7273 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Weighted median | -0.005 | 0.0041 | 0.995 (0.9871-1.003) | 0.2223 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MR, Mendelian randomization; β, beta; SE, standard error; OR, odds ratio; IVW, inverse-variance weighted. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
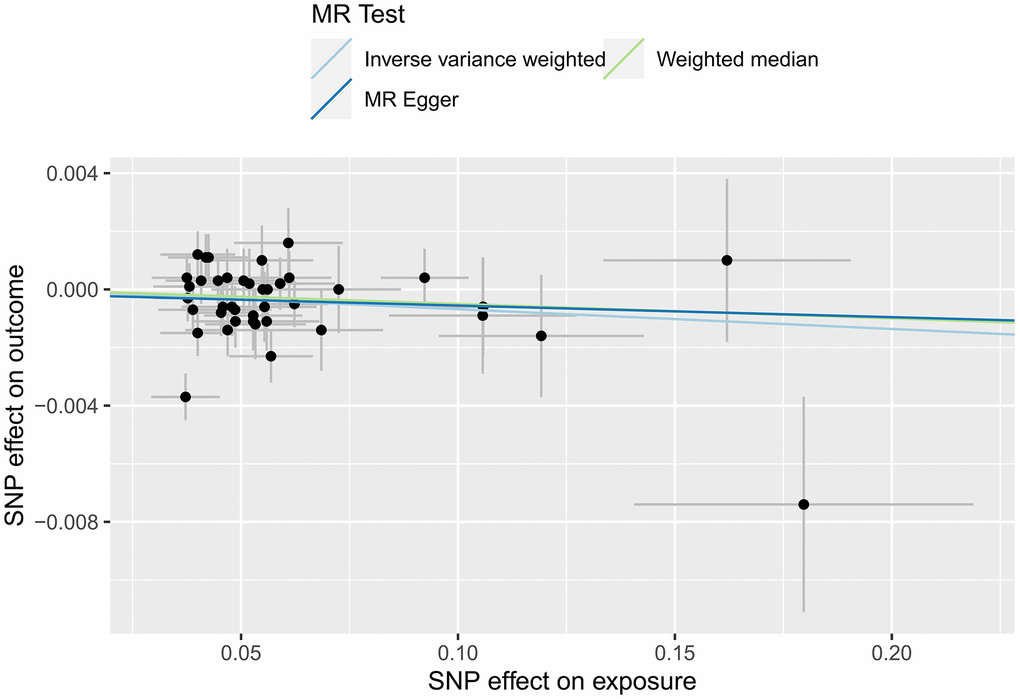
Figure 2. MR scatter plot of cortical thickness in HF patients.
MR analysis of the HF and cortical surface
In this study, IVW, MR-Egger regression and WME analyses in the TwoSampleMR package were used to perform MR analysis of HFs and the surface area of the cerebral cortex. Table 3 shows the results of the MR analysis, and the scatter plot is shown in Figure 3. IVW analysis revealed β=-601.7768, SE=491.1655, P=0.2205, and OR (95% CI)=4.483756E-262 (0-5.497039E +156); MR-Egger analysis revealed β=23.0309, SE=1675.8267, P=0.9891, and OR (95% CI)=1.005105e+10 (0-inf); and WME analysis showed β=-535.222, SE=641.9665, P=0.4044, and OR (95% CI)=3.597802E-233 (0-inf). According to the results of the IVW, MR-Egger and WME analyses, none of the three analysis methods showed statistical significance, suggesting that there was no direct causal relationship between the cerebral cortex surface area and HF incidence. Moreover, we used the MR-Egger intercept to verify the presence of pleiotropy in this study, and the results showed Egger’s intercept=-34.69052, SE =88.86852, P = 0.6984, and a global test pval=0.045, indicating that heart failure and cerebral cortex surface area may be pleiotropic.
Table 3. MR results of the effect of HF on cortical surface area.
| MR method | β | SE | OR (95% CI) | P | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IVW | -601.7768 | 491.1655 | 4.483756E-262 (0-5.497039e+156) | 0.2205 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MR–Egger | 23.0309 | 1675.8267 | 1.005105e+10 (0-inf) | 0.9891 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Weighted median | -535.222 | 641.9665 | 3.597802E-233 (0-inf) | 0.4044 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MR, Mendelian randomization; β, beta; SE, standard error; OR, odds ratio; IVW, inverse-variance weighted. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
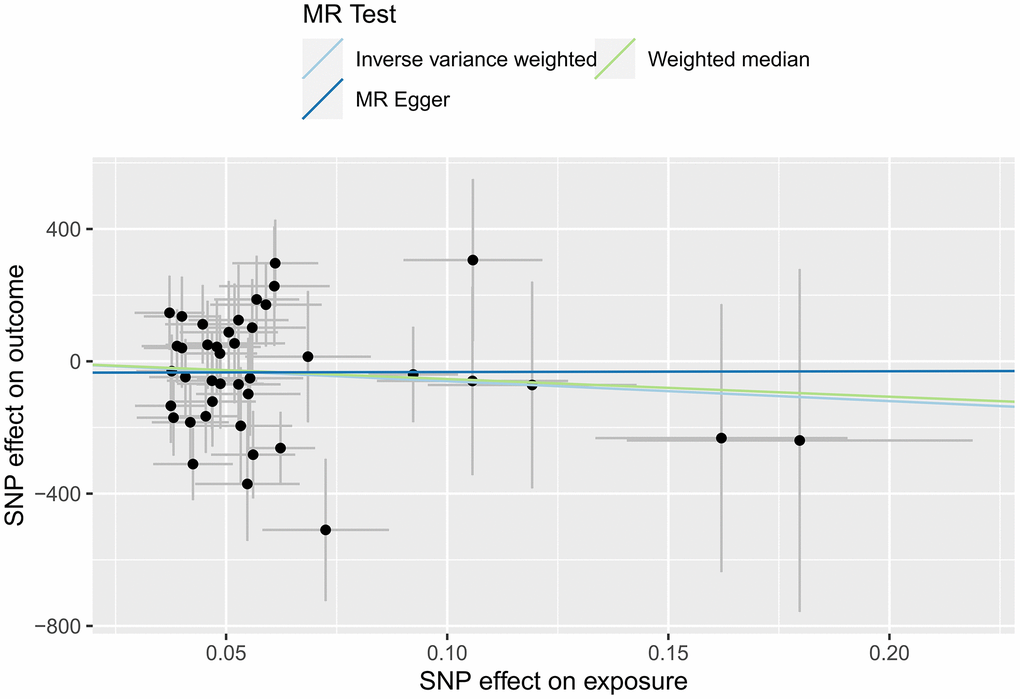
Figure 3. MR scatter plot of the effect of HF on the cerebral cortex surface area.
MR analysis of the HF and cerebral cortex
In this study, according to 34 specific cerebral cortical functional areas with known functions defined by the Desikan-Killiany atlas [15], the IVW analysis method in the TwoSampleMR package was used to conduct MR analysis on the effects of HF on the structure of cerebral cortical functional areas. In this section, we present the MR analysis results for 34 functional brain regions using the global weighted method because the global weighted method may be less affected by neuroanatomical variation between different individuals [16]. Figure 4 and Supplementary Table 2 show the results of this part of the study, suggesting that HF had a significant impact on the surface area of the caudal middle frontal lobule (P=0.009), insula lobule (P=0.01), precuneus lobule (P=0.049) and thickness of the superior parietal lobule (P=0.044). This difference was statistically significant. A scatter diagram of the MR analysis of the structure of various functional areas of the cerebral cortex in HF patients is shown (Supplementary Figures 1–3, 8–10). The pleiotropic analysis of the effects of HF on the structure of various functional areas of the brain is shown in Supplementary Tables 3, 4.
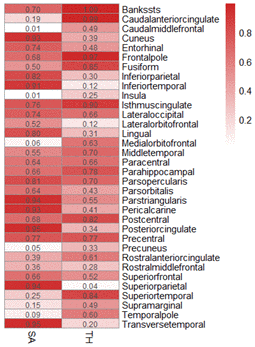
Figure 4. MR heatmap of the effect of HF on the structure of the cerebral cortex.
Sensitivity analysis of the HF and cerebral cortex structure
This study strictly followed the screening criteria for IVs, and most of the included participants were European; therefore, the possibility of false negative results was unlikely, and the results were tested for heterogeneity. The IVW test of the effect of HF on cerebral cortex thickness yielded Cochran’s Q=53.8265 and P=0.071; the MR-Egger test yielded Cochran’s Q=53.7344 and P=0.058. All P-values were >0.05, indicating no heterogeneity. The results are shown in Figure 5.
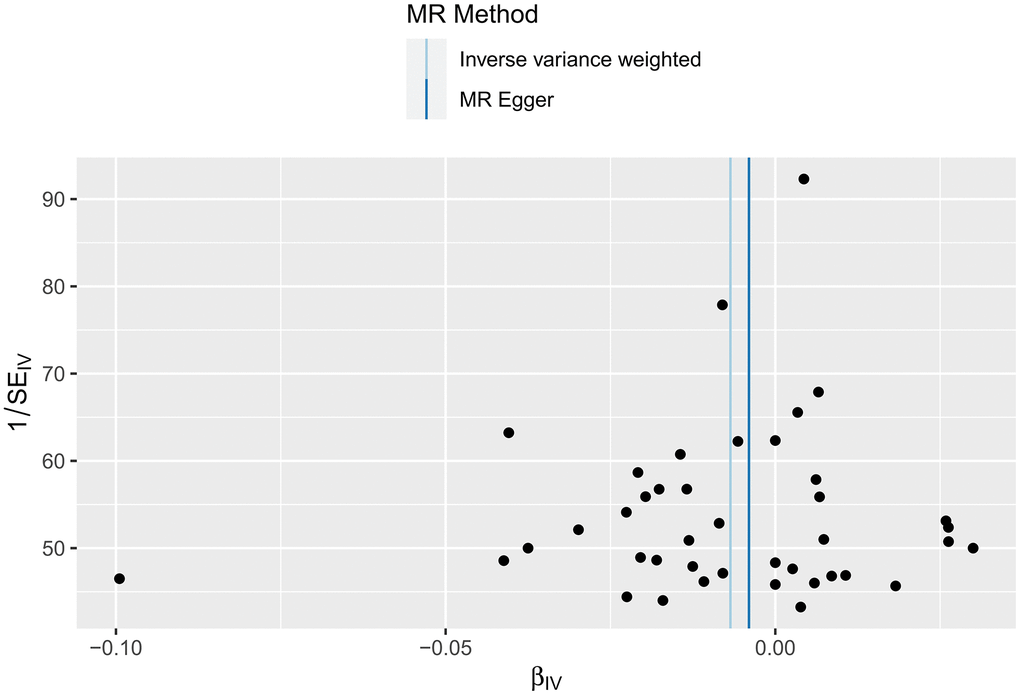
Figure 5. Funnel plot of the heterogeneity test results for the effect of HF on cerebral cortex thickness according to the MR method.
The IVW test of the effect of HF on the cerebral cortex surface area yielded Cochran’s Q=56.6142 and P=0.042; the MR-Egger test yielded Cochran’s Q=56.3939 and P=0.035. All P-values were <0.05, indicating heterogeneity. The results are shown in Figure 6. A funnel plot for the analysis of cortical heterogeneity in the relationship between various functional areas of the brain and HF is shown in Supplementary Figures 4, 5, 11, 12 and Supplementary Tables 5, 6.
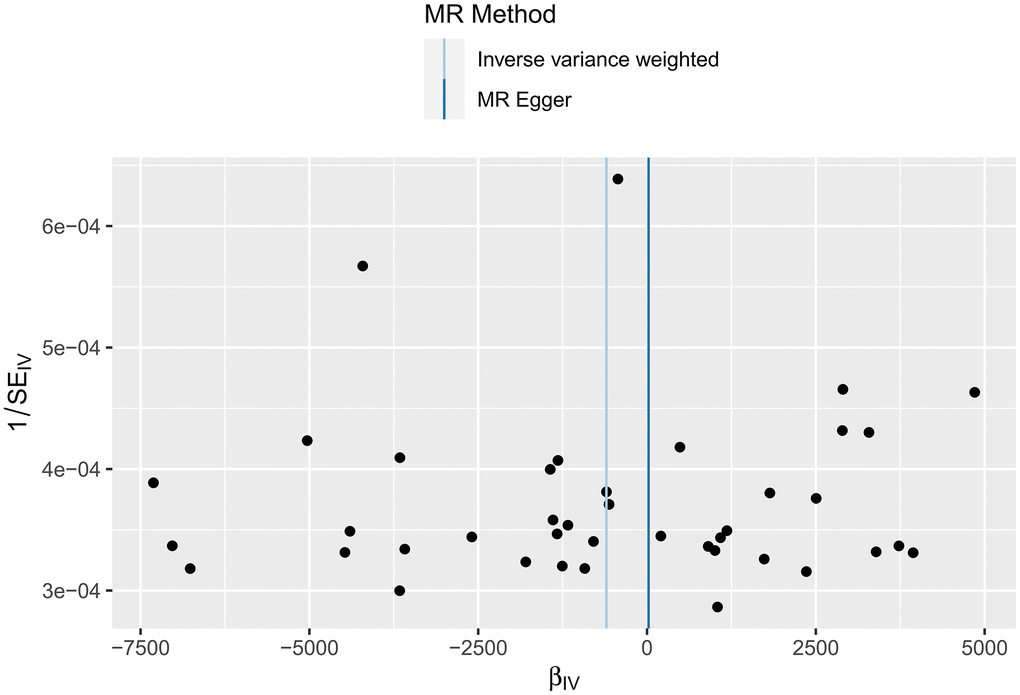
Figure 6. Funnel plot of the heterogeneity test results of the MR analysis of the relationship between HF and the cerebral cortex surface area.
Subsequently, we applied the “leave-one-out” method of sensitivity analysis to visualize the IVW analysis results of the relationship between HF and cerebral cortex structure (Figures 7, 8). After the above significant and independent SNPs were sequentially excluded, the IVW outcome effect values of the remaining SNPs did not significantly fluctuate. All the SNPs were close to the red dot position in the forest plot, and all the P-values were >0.05, indicating that there was no SNP in the IVs that strongly influenced the results; this finding showed that the results obtained by the IVW analysis method were stable and reliable. The sensitivity analysis results of the effects of HF on the various functional areas of the brain are shown in Supplementary Figures 6, 7, 13, 14.
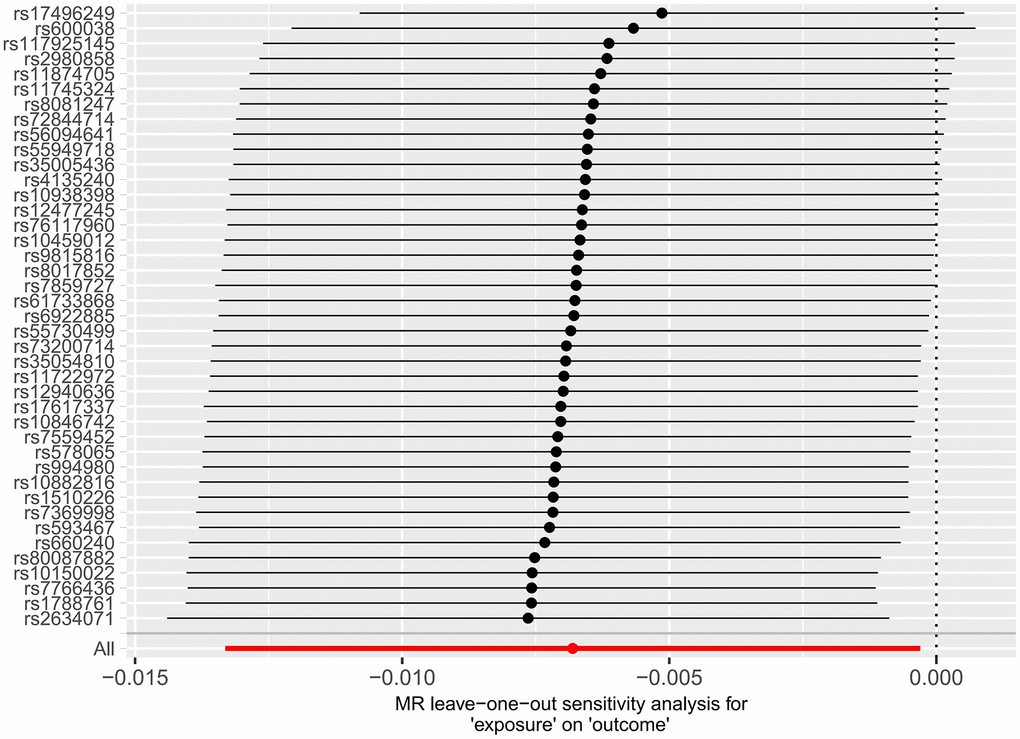
Figure 7. Forest plot of the “leave-one-out” method in the MR analysis of the relationship between HF and cerebral cortex thickness.
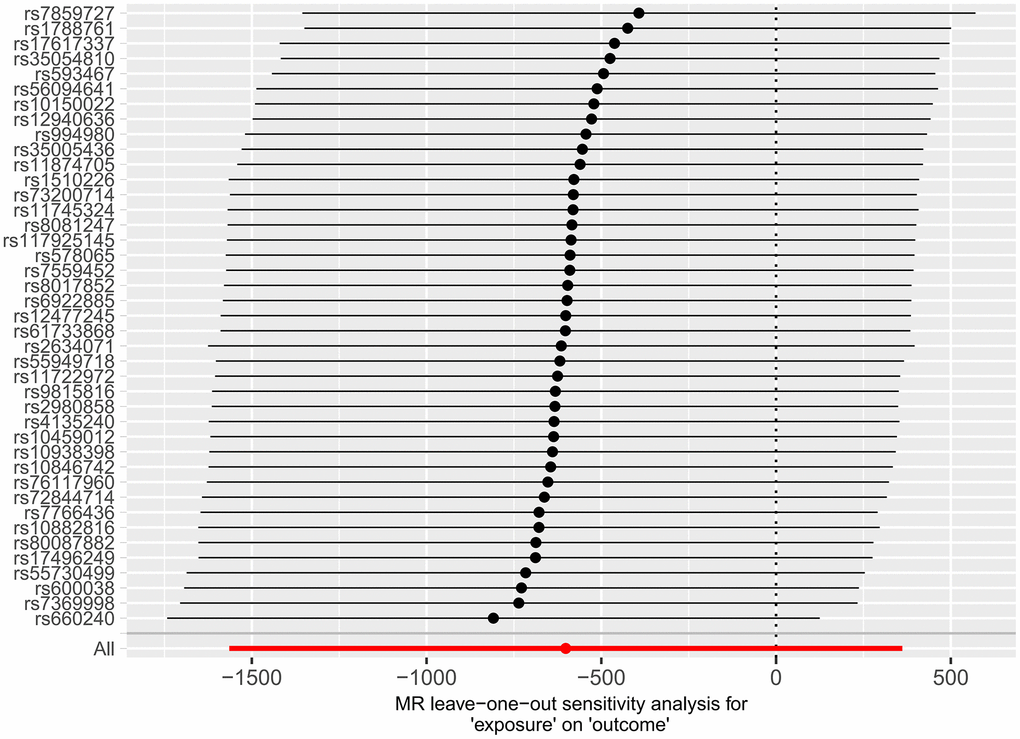
Figure 8. Forest plot of the “leave-one-out” method used in the MR analysis of the relationship between HF and cerebral cortex surface area.
Discussion
At present, numerous observational studies have shown that cerebral cortex structure is related to HF incidence, but there is no evidence showing whether there is a causal relationship between the two conditions. This study systematically identified a causal relationship between HF and cerebral cortex structure using two-sample MR analysis. Our findings suggest that HF affects cortical thickness, specifically the surface area of the caudal middle frontal lobule, insula, precuneus lobule, and superior parietal lobule. Moreover, sensitivity analysis did not reveal pleiotropy, further confirming the stability of the conclusion. This difference may be related to the reduced pumping function of the heart muscle in patients with HF.
Cortical structure, specifically cortical thickness, is considered a neuroimaging biomarker for predicting cognitive decline. Moreover, it is also believed that the surface area of the cerebral cortex is the key to reducing cortical volume in patients with cognitive or mental disorders [17]. Moreover, numerous studies have suggested that cortical surface area may be more sensitive than cortical thickness for the prediction of mental illness [18–20]. The middle frontal gyrus, where the caudal middle frontal lobule is located, is a region of the ventral prefrontal cortex that has been recognized as an important brain region leading to depression and is responsible for many cognitive functions. Examples of emotion processing include decision processing, emotional cognition [21], working memory [22], attention processing [23, 24], and top-down regulation in emotional processing [25]. Moreover, the prefrontal regions (BA8, BA9, BA10, BA46, and BA47), where the medial frontal gyrus is located, are located in front of the motor and premotor areas. These functional areas are related to human personality and determine a person’s social and moral consciousness and emotional depth [26].
The insula, located deep in the lateral sulci of the brain, is also known as the “Island of Reil” and is located deep in the temporal lobe. The insula was originally described as the paralimbic or limbic integration cortex [27]. Analysis of nearly 1,800 functional neuroimaging experiments revealed that the insula is divided into four main functional regions: the sensorimotor, central-olfactogustatory, socioemotional and cognitive anterior-dorsal regions [28]. The anterior insula, as the cortical center for visceral information processing and perception, is believed to play a crucial role in emotional experiences and subjective feelings [29]. Neural function imaging has shown that the expression of negative emotions is also related to the activation of the insula [30]. Moreover, patients with insular lobe injury exhibit various changes in subjective emotions, mainly manifested as anxiety-related indifference [31, 32]. The insula may play an important role in the management of social emotions.
The precuneus is a functional region of the parietal lobe; as one of the core regions of the default network, the precuneus is involved in the human body’s situational memory, self-focused attention, visuospatial intention and self-emotional processing [33]. We found that the functional connectivity of the Broadman region 47/12 of the lateral orbitofrontal cortex was enhanced with the precuneus, angular gyrus and Broadman region 21 of the visual cortex. This enhanced nonreward or punishment system (Brodmann: BA47 and areas 12) with functional connections to the precuneus and angular gyrus is associated with explicit emotional negative self-awareness and self-esteem in individuals with depression [34]. The posterior cingulate cortex and anterior cuneus are considered hubs of the default mode network (DMN) and are involved in social cognition and theory of mind. Moreover, a previous study revealed that the functional connectivity of the posterior cingulate gyrus, anterior cuneus and angular gyrus is related to the severity of depression [35]. In addition, studies have shown that the superior lobular cortex plays an important role in cognitive control and detail attention [36, 37].
Subsequently, we analyzed the SNPs significantly associated with heart failure identified in the present study and observed that the expression of GPR39 corresponding to rs72844714 decreased in the hippocampus and cortex of patients with depression [38]. Zinc, a stimulator of GPR39, can activate the Gαs pathway, leading to an increase in cyclic adenosine monophosphate (cAMP) and activation of protein kinase A (PKA). This pathway can result in phosphorylation of cAMP response element-binding protein (CREB) and an increase in cAMP response element (CRE)-dependent transcription. Consequently, BDNF leads to the upregulation of brain-derived neurotrophic factor (BDNF) and tropomyosin receptor kinase B (TrkB) in neurons [39]. A zinc-deficient diet for 6 weeks can reduce the protein expression of GPR39 and BDNF in the prefrontal cortex [40]. After prolonged antidepressant treatment, GPR39 is upregulated, where it exerts antidepressant effects through the Gαq pathway via the CREB/BDNF/TrkB pathway [41–43]. Simultaneously, zinc can activate postsynaptic GPR39-mediated increases in intracellular calcium [44]. Calcium release, through upregulation of the postsynaptic membrane KCC2 (K+/Cl−) cotransporter protein, increases K-dependent Cl− efflux in postsynaptic cells, thereby enhancing inhibitory tone and preventing excitotoxicity [44–46].
Notably, GPR39 is closely associated with the dopaminergic and 5-HT systems. When mice are treated with tyrosine hydroxylase inhibitor (alphaMT) and tryptophan hydroxylase inhibitor (pCPA) to block dopamine and 5-HT synthesis, GPR39 in the prefrontal cortex is significantly upregulated [47, 48]. Furthermore, the striatin (STRN) gene corresponding to rs17496249 is widely expressed in the striatum and serves as a regulator of striatal neuron development; moreover, this gene is significantly relevant for diagnosing symptomatic depression in patients with subsyndromal syndromes and severe depressive disorders [50, 51]. The FTO gene corresponding to rs56094641 is highly enriched in the cortex and hippocampus. Interaction with CaMKII delays the dephosphorylation of CREB in human neuroblastoma cells [52]. CaMKII-mediated activation of CREB promotes the transcription and translation of the key neuronal plasticity proteins SYN and PSD95 [53, 54], potentially influencing the occurrence of depression. Further research has confirmed the neuroprotective role of hippocampal FTO in depression-like behavior through the activation of the CaMKII/CREB signaling pathway, improving hippocampal synaptic plasticity (dendritic remodeling, PSD95, and SYN expression) [55].
The structural changes in the cerebral cortex caused by HF may lead to a range of neuropsychiatric symptoms in patients. The link between psychosocial factors and CVD incidence has been identified as an important public health problem that mainly includes psychiatric symptoms such as anxiety and depression and can increase the occurrence of major adverse cardiovascular events [56]. Depression and anxiety have been shown to be prevalent in approximately 15%-20% of CVD patients and can coexist for a long time [57]. This, together with the results of this study, also confirms the theory of “psycho-cardiology”. The concept of “psycho-cardiology” began with an article published in the American Journal of Psychosomatics in 1985 titled “Psychocardiology: meeting place of the heart and mind” [58]. Therefore, we suggest that timely and appropriate mental health education and treatment should be given to patients with HF and other heart diseases to reduce the incidence of various psychiatric symptoms and improve the prognosis of patients with CVD.
This study explored the effect of HF on cerebral cortex structure at the genetic level. According to our review results, this study is the first to perform MR analysis on the causal relationship between HF and cerebral cortex structure. Our results showed that although there was a direct effect of HF only on the cortical thickness of the whole-brain structure, there was a direct causal relationship between HF and the surface areas of the caudal middle frontal lobule, insula lobule, and precuneus lobule and the cortical thickness of the superior parietal lobule. This result supports the causal relationship between cardiac injury and neurological dysfunction, providing systematic and strong evidence for the theory of the “heart-brain axis” and “psycho-cardiology” Figure 9.
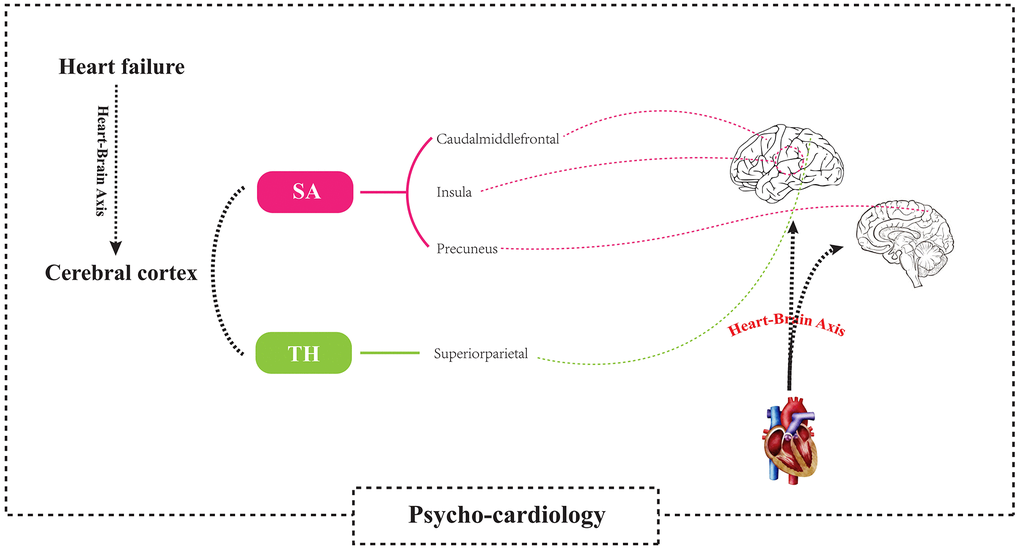
Figure 9. Using a two-sample Mendelian randomization framework, we revealed that heart failure causally influences brain cortical structure alterations, supporting the existence of the “heart-brain axis”. SA, Surficial area; TH, thickness.
This study has several limitations. First, our study was limited to individuals of European descent, so whether our findings are generalizable to other individuals of different ethnicities remains unknown. Second, in this two-sample MR study design, we were unable to determine whether there was sample overlap between the exposure and outcome factor populations of the included GWAS dataset, which could lead to bias in the results. Third, we did not distinguish the phenotypes of patients with HF in more detail, such as those with reserved ejection fraction, those with reduced ejection fraction, or those with intermediate ejection fraction; therefore, we did not explore the effects of these different phenotypes on cerebral cortex structure. Finally, although a series of methods were used to rule out potential confounders and outliers and the sensitivity analyses did not detect any pleiotropy, we still cannot completely rule out all potential pleiotropy. Given these limitations, additional research should be performed to better confirm these possible associations, especially the clinical outcomes reflected in these results.
Conclusions
In summary, this study revealed a direct association between HF and cerebral cortex structure through comprehensive and systematic MR analysis. Our results showed that the surface area of the caudal middle frontal lobule, insula lobule, and precuneus lobule and the cortical thickness of the superior parietal lobule were directly affected by HF. Head MRI may be used for the early diagnosis and prediction of neuropsychiatric diseases in patients with HF. To some extent, this study provides a theoretical basis for theories of the “heart-brain axis” and “psycho-cardiology”. However, due to the limitations of this study, the specific mechanism of the “heart-brain axis” should be further investigated.
Author Contributions
Hu Y.Q. designed the study, wrote the first draft of the manuscript and verified the underlying data. Hu Y.Q., Xu Z.H., Yang J.H. conducted the statistical analysis, Xu Z.H., Zhang M. and Shi T.Y. played a role in the acquisition of the data and analyses, and participated in data interpretation. Liu Z.R. and Wan Q.Q edited the figures and tables. Liu Y.M. directed the study design and funded the study. All authors revised and approved the final manuscript. The guarantor (Liu Y.M.) confirms that all listed authors meet the authorship criteria and that no others meeting the criteria have been omitted.
Acknowledgments
We would like to extend our sincere gratitude to the corresponding author of this study, Yongming Liu, for her instructive advice and useful suggestions on this study. We are deeply grateful of her funding and help in the completion of this study.
Conflicts of Interest
All authors declare the following: no support from any organization for the submitted work; no financial relationships with any organizations that might have an interest in the submitted work in the previous three years; and no other relationships or activities that could appear to have influenced the submitted work.
Funding
This study was supported by the National Natural Science Foundation of China (No. 81973611) and the Traditional Chinese Medicine Advantage Disease Diagnosis and Treatment Capacity Construction Project of Administration of Traditional Chinese Medicine of Anhui Province (340000232428000100010).
References
- 1. Benjamin EJ, Muntner P, Alonso A, Bittencourt MS, Callaway CW, Carson AP, Chamberlain AM, Chang AR, Cheng S, Das SR, Delling FN, Djousse L, Elkind MS, et al, and American Heart Association Council on Epidemiology and Prevention Statistics Committee and Stroke Statistics Subcommittee. Heart Disease and Stroke Statistics-2019 Update: A Report From the American Heart Association. Circulation. 2019; 139:e56–528. https://doi.org/10.1161/CIR.0000000000000659 [PubMed]
- 2. Lampros P, Prabhjot G, Andreas PK, Kalogeropoulos AP, Skopicki HA, Butler J. Heart Failure: An Essential Clinical Guide. st ed. Boca Raton: CRC Press. 2022:244–53.
- 3. Zhao Y, Gong CX. From chronic cerebral hypoperfusion to Alzheimer-like brain pathology and neurodegeneration. Cell Mol Neurobiol. 2015; 35:101–10. https://doi.org/10.1007/s10571-014-0127-9 [PubMed]
- 4. Mueller K, Thiel F, Beutner F, Teren A, Frisch S, Ballarini T, Möller HE, Ihle K, Thiery J, Schuler G, Villringer A, Schroeter ML. Brain Damage With Heart Failure: Cardiac Biomarker Alterations and Gray Matter Decline. Circ Res. 2020; 126:750–64. https://doi.org/10.1161/CIRCRESAHA.119.315813 [PubMed]
- 5. Kumar R, Yadav SK, Palomares JA, Park B, Joshi SH, Ogren JA, Macey PM, Fonarow GC, Harper RM, Woo MA. Reduced regional brain cortical thickness in patients with heart failure. PLoS One. 2015; 10:e0126595. https://doi.org/10.1371/journal.pone.0126595 [PubMed]
- 6. Holmes MV, Ala-Korpela M, Smith GD. Mendelian randomization in cardiometabolic disease: challenges in evaluating causality. Nat Rev Cardiol. 2017; 14:577–90. https://doi.org/10.1038/nrcardio.2017.78 [PubMed]
- 7. Zheng J, Baird D, Borges MC, Bowden J, Hemani G, Haycock P, Evans DM, Smith GD. Recent Developments in Mendelian Randomization Studies. Curr Epidemiol Rep. 2017; 4:330–45. https://doi.org/10.1007/s40471-017-0128-6 [PubMed]
- 8. Hemani G, Zheng J, Elsworth B, Wade KH, Haberland V, Baird D, Laurin C, Burgess S, Bowden J, Langdon R, Tan VY, Yarmolinsky J, Shihab HA, et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018; 7:e34408. https://doi.org/10.7554/eLife.34408 [PubMed]
- 9. Wu F, Huang Y, Hu J, Shao Z. Mendelian randomization study of inflammatory bowel disease and bone mineral density. BMC Med. 2020; 18:312. https://doi.org/10.1186/s12916-020-01778-5 [PubMed]
- 10. Yuan S, Larsson S. Causal associations of iron status with gout and rheumatoid arthritis, but not with inflammatory bowel disease. Clin Nutr. 2020; 39:3119–24. https://doi.org/10.1016/j.clnu.2020.01.019 [PubMed]
- 11. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017; 32:377–89. https://doi.org/10.1007/s10654-017-0255-x [PubMed]
- 12. Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet Epidemiol. 2016; 40:304–14. https://doi.org/10.1002/gepi.21965 [PubMed]
- 13. Carter AR, Sanderson E, Hammerton G, Richmond RC, Davey Smith G, Heron J, Taylor AE, Davies NM, Howe LD. Mendelian randomisation for mediation analysis: current methods and challenges for implementation. Eur J Epidemiol. 2021; 36:465–78. https://doi.org/10.1007/s10654-021-00757-1 [PubMed]
- 14. Shi Y, Feng S, Yan M, Wei S, Yang K, Feng Y. Inflammatory bowel disease and celiac disease: A bidirectional Mendelian randomization study. Front Genet. 2022; 13:928944. https://doi.org/10.3389/fgene.2022.928944 [PubMed]
- 15. Chen CH, Fiecas M, Gutiérrez ED, Panizzon MS, Eyler LT, Vuoksimaa E, Thompson WK, Fennema-Notestine C, Hagler DJ
Jr , Jernigan TL, Neale MC, Franz CE, Lyons MJ, et al. Genetic topography of brain morphology. Proc Natl Acad Sci USA. 2013; 110:17089–94. https://doi.org/10.1073/pnas.1308091110 [PubMed] - 16. Mueller S, Wang D, Fox MD, Yeo BT, Sepulcre J, Sabuncu MR, Shafee R, Lu J, Liu H. Individual variability in functional connectivity architecture of the human brain. Neuron. 2013; 77:586–95. https://doi.org/10.1016/j.neuron.2012.12.028 [PubMed]
- 17. Buechler R, Wotruba D, Michels L, Theodoridou A, Metzler S, Walitza S, Hänggi J, Kollias S, Rössler W, Heekeren K. Cortical Volume Differences in Subjects at Risk for Psychosis Are Driven by Surface Area. Schizophr Bull. 2020; 46:1511–9. https://doi.org/10.1093/schbul/sbaa066 [PubMed]
- 18. Schmaal L, Yücel M, Ellis R, Vijayakumar N, Simmons JG, Allen NB, Whittle S. Brain Structural Signatures of Adolescent Depressive Symptom Trajectories: A Longitudinal Magnetic Resonance Imaging Study. J Am Acad Child Adolesc Psychiatry. 2017; 56:593–601.e9. https://doi.org/10.1016/j.jaac.2017.05.008 [PubMed]
- 19. Yao N, Winkler AM, Barrett J, Book GA, Beetham T, Horseman R, Leach O, Hodgson K, Knowles EE, Mathias S, Stevens MC, Assaf M, van Erp TG, et al. Inferring pathobiology from structural MRI in schizophrenia and bipolar disorder: Modeling head motion and neuroanatomical specificity. Hum Brain Mapp. 2017; 38:3757–70. https://doi.org/10.1002/hbm.23612 [PubMed]
- 20. Madre M, Canales-Rodríguez EJ, Fuentes-Claramonte P, Alonso-Lana S, Salgado-Pineda P, Guerrero-Pedraza A, Moro N, Bosque C, Gomar JJ, Ortíz-Gil J, Goikolea JM, Bonnin CM, Vieta E, et al. Structural abnormality in schizophrenia versus bipolar disorder: A whole brain cortical thickness, surface area, volume and gyrification analyses. Neuroimage Clin. 2020; 25:102131. https://doi.org/10.1016/j.nicl.2019.102131 [PubMed]
- 21. Fleck MS, Daselaar SM, Dobbins IG, Cabeza R. Role of prefrontal and anterior cingulate regions in decision-making processes shared by memory and nonmemory tasks. Cereb Cortex. 2006; 16:1623–30. https://doi.org/10.1093/cercor/bhj097 [PubMed]
- 22. Kikinis Z, Fallon JH, Niznikiewicz M, Nestor P, Davidson C, Bobrow L, Pelavin PE, Fischl B, Yendiki A, McCarley RW, Kikinis R, Kubicki M, Shenton ME. Gray matter volume reduction in rostral middle frontal gyrus in patients with chronic schizophrenia. Schizophr Res. 2010; 123:153–9. https://doi.org/10.1016/j.schres.2010.07.027 [PubMed]
- 23. Beevers CG, Clasen PC, Enock PM, Schnyer DM. Attention bias modification for major depressive disorder: Effects on attention bias, resting state connectivity, and symptom change. J Abnorm Psychol. 2015; 124:463–75. https://doi.org/10.1037/abn0000049 [PubMed]
- 24. Japee S, Holiday K, Satyshur MD, Mukai I, Ungerleider LG. A role of right middle frontal gyrus in reorienting of attention: a case study. Front Syst Neurosci. 2015; 9:23. https://doi.org/10.3389/fnsys.2015.00023 [PubMed]
- 25. Burgess PW, Gilbert SJ, Dumontheil I. Function and localization within rostral prefrontal cortex (area 10). Philos Trans R Soc Lond B Biol Sci. 2007; 362:887–99. https://doi.org/10.1098/rstb.2007.2095 [PubMed]
- 26. Chauhan P, Rathawa A, Jethwa K, Mehra S. The Anatomy of the Cerebral Cortex. In: Pluta R, editor. Cerebral Ischemia [Internet]. Brisbane (AU): Exon Publications; 2021. https://doi.org/10.36255/exonpublications.cerebralischemia.2021.cerebralcortex [PubMed]
- 27. Augustine JR. Circuitry and functional aspects of the insular lobe in primates including humans. Brain Res Brain Res Rev. 1996; 22:229–44. https://doi.org/10.1016/s0165-0173(96)00011-2 [PubMed]
- 28. Kurth F, Zilles K, Fox PT, Laird AR, Eickhoff SB. A link between the systems: functional differentiation and integration within the human insula revealed by meta-analysis. Brain Struct Funct. 2010; 214:519–34. https://doi.org/10.1007/s00429-010-0255-z [PubMed]
- 29. Craig AD. How do you feel? Interoception: the sense of the physiological condition of the body. Nat Rev Neurosci. 2002; 3:655–66. https://doi.org/10.1038/nrn894 [PubMed]
- 30. Craig AD. How do you feel--now? The anterior insula and human awareness. Nat Rev Neurosci. 2009; 10:59–70. https://doi.org/10.1038/nrn2555 [PubMed]
- 31. Knutson KM, Rakowsky ST, Solomon J, Krueger F, Raymont V, Tierney MC, Wassermann EM, Grafman J. Injured brain regions associated with anxiety in Vietnam veterans. Neuropsychologia. 2013; 51:686–94. https://doi.org/10.1016/j.neuropsychologia.2013.01.003 [PubMed]
- 32. Knutson KM, Dal Monte O, Raymont V, Wassermann EM, Krueger F, Grafman J. Neural correlates of apathy revealed by lesion mapping in participants with traumatic brain injuries. Hum Brain Mapp. 2014; 35:943–53. https://doi.org/10.1002/hbm.22225 [PubMed]
- 33. Geng J, Yan R, Shi J, Chen Y, Mo Z, Shao J, Wang X, Yao Z, Lu Q. Altered regional homogeneity in patients with somatic depression: A resting-state fMRI study. J Affect Disord. 2019; 246:498–505. https://doi.org/10.1016/j.jad.2018.12.066 [PubMed]
- 34. Cheng W, Rolls ET, Qiu J, Liu W, Tang Y, Huang CC, Wang X, Zhang J, Lin W, Zheng L, Pu J, Tsai SJ, Yang AC, et al. Medial reward and lateral non-reward orbitofrontal cortex circuits change in opposite directions in depression. Brain. 2016; 139:3296–309. https://doi.org/10.1093/brain/aww255 [PubMed]
- 35. de Coster L, Leentjens AF, Lodder J, Verhey FR. The sensitivity of somatic symptoms in post-stroke depression: a discriminant analytic approach. Int J Geriatr Psychiatry. 2005; 20:358–62. https://doi.org/10.1002/gps.1290 [PubMed]
- 36. Kompus K, Hugdahl K, Ohman A, Marklund P, Nyberg L. Distinct control networks for cognition and emotion in the prefrontal cortex. Neurosci Lett. 2009; 467:76–80. https://doi.org/10.1016/j.neulet.2009.10.005 [PubMed]
- 37. Qiu L, Su J, Ni Y, Bai Y, Zhang X, Li X, Wan X. The neural system of metacognition accompanying decision-making in the prefrontal cortex. PLoS Biol. 2018; 16:e2004037. https://doi.org/10.1371/journal.pbio.2004037 [PubMed]
- 38. Młyniec K, Doboszewska U, Szewczyk B, Sowa-Kućma M, Misztak P, Piekoszewski W, Trela F, Ostachowicz B, Nowak G. The involvement of the GPR39-Zn(2+)-sensing receptor in the pathophysiology of depression. Studies in rodent models and suicide victims. Neuropharmacology. 2014; 79:290–7. https://doi.org/10.1016/j.neuropharm.2013.12.001 [PubMed]
- 39. Holst B, Egerod KL, Schild E, Vickers SP, Cheetham S, Gerlach LO, Storjohann L, Stidsen CE, Jones R, Beck-Sickinger AG, Schwartz TW. GPR39 signaling is stimulated by zinc ions but not by obestatin. Endocrinology. 2007; 148:13–20. https://doi.org/10.1210/en.2006-0933 [PubMed]
- 40. Młyniec K, Budziszewska B, Reczyński W, Sowa-Kućma M, Nowak G. The role of the GPR39 receptor in zinc deficient-animal model of depression. Behav Brain Res. 2013; 238:30–5. https://doi.org/10.1016/j.bbr.2012.10.020 [PubMed]
- 41. Holst B, Holliday ND, Bach A, Elling CE, Cox HM, Schwartz TW. Common structural basis for constitutive activity of the ghrelin receptor family. J Biol Chem. 2004; 279:53806–17. https://doi.org/10.1074/jbc.M407676200 [PubMed]
- 42. Petrilli MA, Kranz TM, Kleinhaus K, Joe P, Getz M, Johnson P, Chao MV, Malaspina D. The Emerging Role for Zinc in Depression and Psychosis. Front Pharmacol. 2017; 8:414. https://doi.org/10.3389/fphar.2017.00414 [PubMed]
- 43. Młyniec K, Nowak G. Up-regulation of the GPR39 Zn2+-sensing receptor and CREB/BDNF/TrkB pathway after chronic but not acute antidepressant treatment in the frontal cortex of zinc-deficient mice. Pharmacol Rep. 2015; 67:1135–40. https://doi.org/10.1016/j.pharep.2015.04.003 [PubMed]
- 44. Gilad D, Shorer S, Ketzef M, Friedman A, Sekler I, Aizenman E, Hershfinkel M. Homeostatic regulation of KCC2 activity by the zinc receptor mZnR/GPR39 during seizures. Neurobiol Dis. 2015; 81:4–13. https://doi.org/10.1016/j.nbd.2014.12.020 [PubMed]
- 45. Besser L, Chorin E, Sekler I, Silverman WF, Atkin S, Russell JT, Hershfinkel M. Synaptically released zinc triggers metabotropic signaling via a zinc-sensing receptor in the hippocampus. J Neurosci. 2009; 29:2890–901. https://doi.org/10.1523/JNEUROSCI.5093-08.2009 [PubMed]
- 46. Saadi RA, He K, Hartnett KA, Kandler K, Hershfinkel M, Aizenman E. SNARE-dependent upregulation of potassium chloride co-transporter 2 activity after metabotropic zinc receptor activation in rat cortical neurons in vitro. Neuroscience. 2012; 210:38–46. https://doi.org/10.1016/j.neuroscience.2012.03.001 [PubMed]
- 47. Rychlik M, Mlyniec K. Zinc-mediated Neurotransmission in Alzheimer’s Disease: A Potential Role of the GPR39 in Dementia. Curr Neuropharmacol. 2020; 18:2–13. https://doi.org/10.2174/1570159X17666190704153807 [PubMed]
- 48. Młyniec K, Gaweł M, Librowski T, Reczyński W, Bystrowska B, Holst B. Investigation of the GPR39 zinc receptor following inhibition of monoaminergic neurotransmission and potentialization of glutamatergic neurotransmission. Brain Res Bull. 2015; 115:23–9. https://doi.org/10.1016/j.brainresbull.2015.04.005 [PubMed]
- 49. Yang C, Hu G, Li Z, Wang Q, Wang X, Yuan C, Wang Z, Hong W, Lu W, Cao L, Chen J, Wang Y, Yu S, et al. Differential gene expression in patients with subsyndromal symptomatic depression and major depressive disorder. PLoS One. 2017; 12:e0172692. https://doi.org/10.1371/journal.pone.0172692 [PubMed]
- 50. Li D, Musante V, Zhou W, Picciotto MR, Nairn AC. Striatin-1 is a B subunit of protein phosphatase PP2A that regulates dendritic arborization and spine development in striatal neurons. J Biol Chem. 2018; 293:11179–94. https://doi.org/10.1074/jbc.RA117.001519 [PubMed]
- 51. Andersen H, Braestrup C, Randrup A. Apomorphine-induced stereotyped biting in the tortoise in relation to dopaminergic mechanisms. Brain Behav Evol. 1975; 11:365–73. https://doi.org/10.1159/000123646 [PubMed]
- 52. Lin L, Hales CM, Garber K, Jin P. Fat mass and obesity-associated (FTO) protein interacts with CaMKII and modulates the activity of CREB signaling pathway. Hum Mol Genet. 2014; 23:3299–306. https://doi.org/10.1093/hmg/ddu043 [PubMed]
- 53. Yan X, Liu J, Ye Z, Huang J, He F, Xiao W, Hu X, Luo Z. CaMKII-Mediated CREB Phosphorylation Is Involved in Ca2+-Induced BDNF mRNA Transcription and Neurite Outgrowth Promoted by Electrical Stimulation. PLoS One. 2016; 11:e0162784. https://doi.org/10.1371/journal.pone.0162784 [PubMed]
- 54. Song SH, Augustine GJ. Synapsin Isoforms and Synaptic Vesicle Trafficking. Mol Cells. 2015; 38:936–40. https://doi.org/10.14348/molcells.2015.0233 [PubMed]
- 55. Shen J, Yang L, Wei W. Role of Fto on CaMKII/CREB signaling pathway of hippocampus in depressive-like behaviors induced by chronic restraint stress mice. Behav Brain Res. 2021; 406:113227. https://doi.org/10.1016/j.bbr.2021.113227 [PubMed]
- 56. Rozanski A. Behavioral cardiology: current advances and future directions. J Am Coll Cardiol. 2014; 64:100–10. https://doi.org/10.1016/j.jacc.2014.03.047 [PubMed]
- 57. Lichtman JH, Froelicher ES, Blumenthal JA, Carney RM, Doering LV, Frasure-Smith N, Freedland KE, Jaffe AS, Leifheit-Limson EC, Sheps DS, Vaccarino V, Wulsin L, and American Heart Association Statistics Committee of the Council on Epidemiology and Prevention and the Council on Cardiovascular and Stroke Nursing. Depression as a risk factor for poor prognosis among patients with acute coronary syndrome: systematic review and recommendations: a scientific statement from the American Heart Association. Circulation. 2014; 129:1350–69. https://doi.org/10.1161/CIR.0000000000000019 [PubMed]
- 58. Jefferson JW. Psychocardiology: meeting place of heart and mind. Psychosomatics. 1985; 26:841–2. https://doi.org/10.1016/S0033-3182(85)72774-0 [PubMed]