



Introduction
Telomeres served as protective structures located at the ends of chromosomes, and their gradual shortening was intricately linked to the occurrence and progression of cellular senescence, cancer, and various other diseases [1–3]. Recently, there has been extensive research on the role of telomeres in hematologic diseases (HDs). Telomeres have been found to be closely associated with hematopoietic stem cells (HSCs), hematological tumors, and complications arising from hematopoietic stem cell transplantation (HSCT) [4, 5].
Current research has mostly focused on the impact of telomere length (TL) changes on the progression and prognosis of various HDs. For instance, a study revealed that TL of HSCs was significantly reduced in patients with multiple myeloma, which was closely associated with disease progression and prognosis [6]. In patients with acute myeloid leukemia (AML), TL was significantly shortened and associated with disease development and therapeutic efficacy [7, 8]. Studies have shown that HSCT with longer TL has a higher success rate, leading to improved immune system reconstitution, enhanced anti-infection ability, and better anti-tumor effects [9]. In terms of treatment, therapeutic strategies targeting telomeres, such as telomerase inhibitors, telomere-lengthening agents, and the gene therapy, have shown promise in laboratory and clinical trials, providing novel avenues for HDs treatment [10, 11].
However, there is currently no systematic research on the impact of TL changes on the risk of developing various HDs. In recent years, some studies have utilized Mendelian randomization (MR) methods to predict the relationship between TL changes and the risk of developing various diseases. MR can mitigate the problem of reverse causality by using genetic variation as a natural randomization, thus excluding the impact of disease on TL and yielding more reliable conclusions. It can uncover novel causal relationships, such as the causal link between TL and chronic obstructive pulmonary disease [12], Alzheimer’s disease [13], cancer [14], and other diseases, thereby providing new insights for the prevention and treatment of related conditions. Based on this, we aim to leverage the unique advantages of using MR for telomere research to systematically elucidate the impact of genetically predicted TL changes on the risk of developing various HDs.
Materials and Methods
Study design description
The design of the MR study must adhere to three fundamental assumptions: 1) a strong correlation between instrumental variables (IVs) and exposure factors; 2) confounding factors that are independent of the exposure-outcome relationship; 3) genetic variables that only influence outcomes through exposure, rather than through other means [15]. MR analysis was conducted using GWAS data, with TL considered as the exposure and 11 HDs as the outcomes to analyze their association. Figure 1 depicts the overall study design.
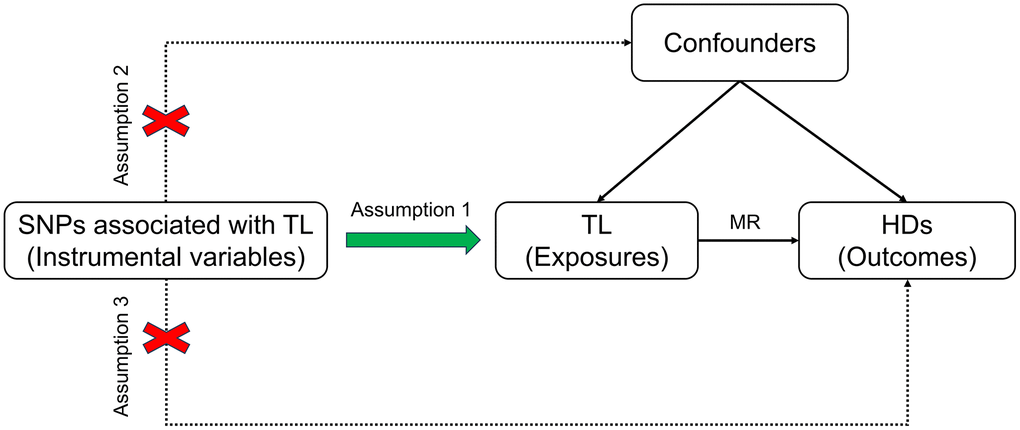
Figure 1. Schematic diagram illustrating the MR. Abbreviations: SNP: single nucleotide polymorphism; TL: telomere length; MR: Mendelian randomization; HDs: hematologic diseases.
Instrumental variables selection
The TL data were obtained from the UK Biobank (download address: https://gwas.mrcieu.ac.uk/datasets/ieu-b-4879/), including 472174 UKB participants [16]. The screening method employed for IVs was based on previous researchers’ approach [17]. Firstly, single nucleotide polymorphisms (SNPs) were selected at the threshold of genome-wide significance (p < 5 × 10−8). Secondly, the parameter r2 threshold was set to 0.001, the kilobase pair (kb) to 10000 and the LD clumping function was used to exclude interference in linkage disequilibrium. Thirdly, we queried all SNPs in PhenoScanner (http://www.phenoscanner.medschl.cam.ac.uk/) to ascertain if there were any SNPs associated with potential confounding factors or outcomes. Subsequently, the F statistic was employed to assess the strength of the IVs, with those having F < 10 being excluded. Lastly, we harmonized exposure and outcome data, excluding palindromic SNPs with intermediate allele frequencies. A total of 152 SNPs were selected for MR analysis (Supplementary File 1).
Hematologic diseases data sources
The GWAS datasets for the 11 HDs (Table 1) were obtained from the FinnGen website (R10 version) (https://www.finngen.fi/en/). The GWAS data related to HDs included acute lymphocytic leukemia (ALL), acute myeloid leukemia (AML), chronic lymphocytic leukemia (CLL), chronic myeloid leukemia (CML), diffuse large b-cell lymphoma (DLBCL), follicular lymphoma (FOLLICULAR), Hodgkin lymphoma (HODGKIN), mantle cell lymphoma (MANTLE), marginal zone b-cell lymphoma (MARGINAL), monocytic leukemia (MONOCYTIC), and mature T/NK-cell lymphomas (TNK). As the aforementioned GWAS datasets were publicly available, ethical approval was not required.
Table 1. Hematologic diseases data sources.
| Diseases | Finngen ID | Population | SNPs | Cases | Controls | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ALL | finngen_R10_C3_ALL_EXALLC | Europeans | 19680787 | 197 | 314192 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AML | finngen_R10_C3_AML_EXALLC | Europeans | 19680799 | 244 | 314192 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CLL | finngen_R10_C3_CLL_EXALLC | Europeans | 19680807 | 668 | 314189 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CML | finngen_R10_C3_CML_EXALLC | Europeans | 19050426 | 115 | 314192 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DLBCL | finngen_R10_C3_DLBCL_EXALLC | Europeans | 19680818 | 1050 | 314193 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FOLLICULAR | finngen_R10_CD2_FOLLICULAR_LYMPHOMA_EXALLC | Europeans | 19681177 | 1181 | 324650 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HODGKIN | finngen_R10_CD2_HODGKIN_LYMPHOMA_EXALLC | Europeans | 19681163 | 846 | 324650 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MANTLE | finngen_R10_C3_MANTLE_CELL_LYMPHOMA_EXALLC | Europeans | 19680787 | 210 | 314193 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MARGINAL | finngen_R10_C3_MARGINAL_ZONE_LYMPHOMA_EXALLC | Europeans | 19680791 | 202 | 314193 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MONOCYTIC | finngen_R10_CD2_MONOCYTIC_LEUKAEMIA_EXALLC | Europeans | 19681137 | 85 | 324650 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TNK | finngen_R10_CD2_TNK_LYMPHOMA_EXALLC | Europeans | 19681156 | 363 | 324650 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: SNP: single nucleotide polymorphism; ALL: acute lymphocytic leukemia; AML: acute myeloid leukemia; CLL: chronic lymphocytic leukemia; CML: chronic myeloid leukemia; DLBCL: diffuse large b-cell lymphoma; FOLLICULAR: follicular lymphoma; HODGKIN: Hodgkin lymphoma; MANTLE: mantle cell lymphoma; MARGINAL: marginal zone b-cell lymphoma; MONOCYTIC: monocytic leukemia; TNK: mature T/NK-cell lymphomas. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Statistical analysis
Six MR methods, namely inverse variance weighted (random effects) (IVW-RE), IVW (fixed effects) (IVW-FE), Weighted median, MR Egger, Simple mode, and Weighted mode were employed to estimate the relationship between TL and 11 HDs. In cases where the causal effect estimates from the six models were inconsistent, the results from the IVW-RE or IVW-FE method were considered as the primary outcome. The odds ratio (OR) and the corresponding 95% confidence interval (CI) were used to estimate the degree of causality, with OR>1 indicating exposure as a risk factor for the outcome, OR<1 indicating exposure as a protective factor for the outcome, and p < 0.05 indicating statistically significant causality. The Cochran’s Q test was utilized to detect heterogeneity between IVs, with p < 0.05 indicating heterogeneity. When there is no heterogeneity, IVW-FE is chosen; when heterogeneity exists, IVW-RE is chosen [18]. Sensitivity analysis was mainly conducted using the leave-one-out method, whereby one SNP was removed at a time to assess the stability and reliability of the causal relationship between the remaining SNPs and the outcome. An MR Egger intercept test was performed to detect the presence of directional pleiotropy in the IVs, with p < 0.05 indicating directional pleiotropy. Funnel plots were generated to assess directional pleiotropy. The MR analysis was implemented in R software (version 4.3.1) using the R package “TwoSampleMR” (version 0.5.6).
Results
The impact of genetically predicted changes in TL on the risk of developing 11 HDs were analyzed, and the findings were presented in Figure 2. The IVW-FE/IVW-RE analysis revealed a significant causal relationship (p < 0.05) between genetically predicted TL changes and the risk of developing ALL (IVW-FE: OR = 3.380, 95% CI: 1.520–7.516, p = 0.003), AML (IVW-FE: OR = 2.260, 95% CI: 1.098–4.652, p = 0.027), CLL (IVW-FE: OR = 3.052, 95% CI: 1.986–4.690, p < 0.001), MANTLE (IVW-FE: OR = 3.133, 95% CI: 1.434–6.843, p = 0.004), and HODGKIN (IVW-FE: OR = 1.854, 95% CI: 1.265–2.717, p = 0.002). And among all the positive results, the results of the six MR methods were consistent (OR > 1), indicating that our analysis was reliable. The scatter plots also showed the consistency of the six MR methods and the reliability of the results (Figure 3).
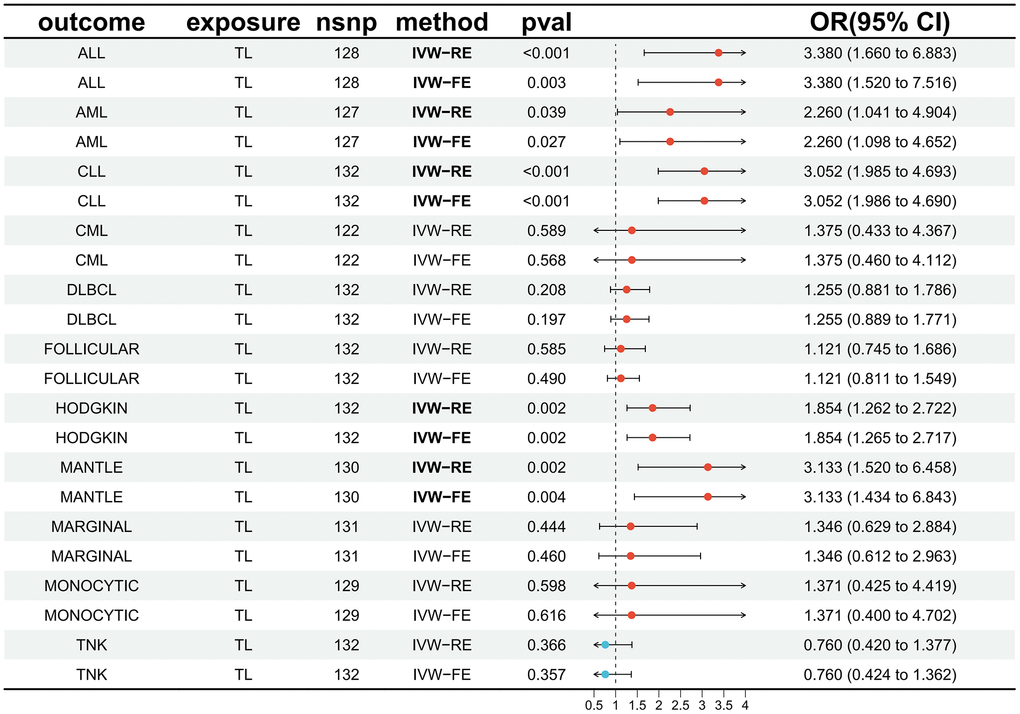
Figure 2. Forest plot illustrating the association between genetically predicted TL and 11 HDs. Abbreviations: TL: telomere length; HDs: hematologic diseases; IVW-RE: inverse variance weighted (random effects); IVW-FE: inverse variance weighted (fixed effects); OR: odds ratio; CI: confidence interval; ALL: acute lymphocytic leukemia; AML: acute myeloid leukemia; CLL: chronic lymphocytic leukemia; CML: chronic myeloid leukemia; DLBCL: diffuse large b-cell lymphoma; FOLLICULAR: follicular lymphoma; HODGKIN: Hodgkin lymphoma; MANTLE: mantle cell lymphoma; MARGINAL: marginal zone b-cell lymphoma; MONOCYTIC: monocytic leukemia; TNK: mature T/NK-cell lymphomas.
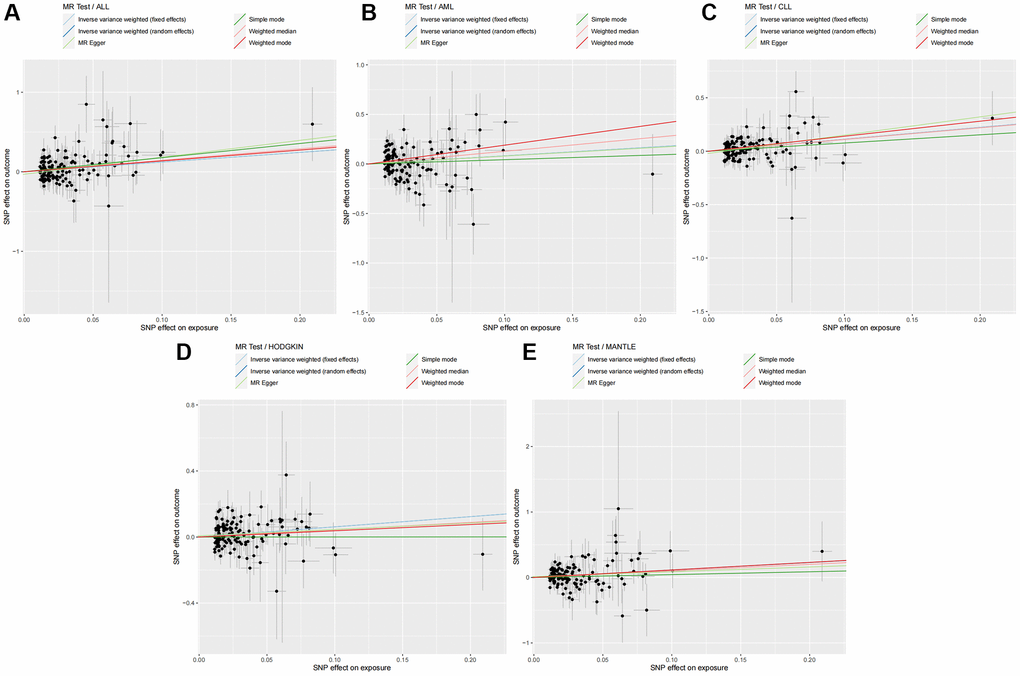
Figure 3. Six MR methods demonstrated the causal effect of TL on HDs. (A) Scatter plot illustrating the association between TL and ALL. (B) Scatter plot illustrating the association between TL and AML. (C) Scatter plot illustrating the association between TL and CLL. (D) Scatter plot illustrating the association between TL and HODGKIN. (E) Scatter plot illustrating the association between TL and MANTLE. Abbreviations: MR: Mendelian randomization; TL: telomere length; HDs: hematologic diseases; ALL: acute lymphocytic leukemia; AML: acute myeloid leukemia; CLL: chronic lymphocytic leukemia; HODGKIN: Hodgkin lymphoma; MANTLE: mantle cell lymphoma.
The IVW-FE/IVW-RE results did not demonstrate a significant causal relationship (p > 0.05) between genetically predicted TL changes and the risk of developing CML (IVW-FE: OR = 1.375, 95% CI: 0.460–4.112, p = 0.568), DLBCL (IVW-FE: OR = 1.255, 95% CI: 0.889–1.771, p = 0.197), MARGINAL (IVW-FE: OR = 1.346, 95% CI: 0.612–2.963, p = 0.460), FOLLICULAR (IVW-RE: OR = 1.121, 95% CI: 0.745–1.686, p = 0.585), MONOCYTIC (IVW-FE: OR = 1.371, 95% CI: 0.400–4.702, p = 0.616), and TNK (IVW-FE: OR = 0.760, 95% CI: 0.424–1.362, p = 0.357). And among all the negative results, the results of other analytical methods also failed to establish a significant causal relationship (p > 0.05). The scatter plots for all negative results were available in the Supplementary File 2.
In the MR analysis, we assessed the heterogeneity and directional pleiotropy, as presented in Table 2. The results of the leave-one-out sensitivity analysis and the funnel plot results can be found in the Supplementary File 3. In the MR analysis using TL as the exposure, we observed a certain degree of heterogeneity only in the TL-FOLLICULAR MR analysis, hence selecting IVW-RE as the primary analytical result. No significant heterogeneity and directional pleiotropy were found in all other MR analyses, indicating a high level of reliability and reproducibility in the results. The results of all MR analyses were provided in Supplementary File 4.
Table 2. Pleiotropy and heterogeneity analyses.
| Outcome | Exposure | Heterogeneity | Pleiotropy | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Q_mr egger | Q_pval_mr egger | Q_IVW | Q_pval_IVW | Egger intercept | SE | p | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ALL | TL | 98.484 | 0.967 | 100.578 | 0.960 | −0.031 | 0.021 | 0.150 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AML | TL | 145.068 | 0.106 | 145.072 | 0.118 | 0.001 | 0.020 | 0.957 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CLL | TL | 128.355 | 0.524 | 131.412 | 0.473 | −0.020 | 0.011 | 0.083 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CML | TL | 134.178 | 0.178 | 134.678 | 0.187 | 0.021 | 0.031 | 0.505 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DLBCL | TL | 137.132 | 0.317 | 137.644 | 0.328 | −0.006 | 0.009 | 0.487 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| FOLLICULAR | TL | 207.599 | <0.001 | 208.750 | <0.001 | 0.009 | 0.011 | 0.397 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HODGKIN | TL | 131.681 | 0.442 | 132.293 | 0.452 | 0.008 | 0.010 | 0.438 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MANTLE | TL | 110.147 | 0.871 | 110.599 | 0.878 | 0.014 | 0.020 | 0.503 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MARGINAL | TL | 120.772 | 0.685 | 121.255 | 0.696 | −0.014 | 0.021 | 0.488 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MONOCYTIC | TL | 115.320 | 0.763 | 115.445 | 0.779 | −0.011 | 0.032 | 0.724 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TNK | TL | 134.146 | 0.384 | 136.102 | 0.362 | −0.021 | 0.016 | 0.171 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: IVW: inverse variance weighted; SE: standard error; ALL: acute lymphocytic leukemia; AML: acute myeloid leukemia; CLL: chronic lymphocytic leukemia; CML: chronic myeloid leukemia; DLBCL: diffuse large b-cell lymphoma; FOLLICULAR: follicular lymphoma; HODGKIN: Hodgkin lymphoma; MANTLE: mantle cell lymphoma; MARGINAL: marginal zone b-cell lymphoma; MONOCYTIC: monocytic leukemia; TNK: mature T/NK-cell lymphomas. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Discussion
In this study, we utilized genetic variations determining TL as a surrogate measure and employed MR analysis to explore the relationship between TL and the risk of 11 HDs. These findings merit our attention and necessitate a reevaluation of the potential diagnostic and therapeutic significance of telomeres in HDs. Additionally, it is noteworthy that these investigations analyzed the association between TL and the risk of HDs, which differs from studies analyzing the relationship between TL and the prognosis of HDs.
Acute lymphoblastic leukemia (ALL) is a malignant and invasive tumor that includes B-cell ALL (B-ALL) and T-cell ALL (T-ALL), caused by abnormal proliferation of lymphocytes in the bone marrow [19]. There have been numerous studies on the relationship between TL and the prognosis of ALL. In B-ALL patients, high telomerase activity, elevated telomere reverse transcriptase (TERT) expression, and telomere shortening were closely associated with poor prognosis [20, 21]. However, some studies have also found that telomere elongation was linked to poor prognosis in ALL [22]. Evidently, there was no consistent pattern in the function of telomeres and telomerase in ALL, especially considering the various potential atypical roles of TERT [23]. Our analysis revealed that genetically predicted longer TL increases the risk of ALL and suggests that longer TL is the primary driver of ALL onset; measuring the difference in TL length between tumor cells before the onset of ALL (before the malignant clone proliferates extensively) and healthy individuals may potentially predict the onset of ALL in advance. In terms of treatment, telomerase inhibitors (such as imetelstat) may enhance standard ALL therapy [23]; therefore, the prospect of combined targeted therapy against telomeres and telomerase in treating ALL is promising.
AML is a highly heterogeneous leukemia, influenced by multiple factors [24]. Recent research has focused on the relationship between TL and the prognosis of AML. Most studies have found that TL is shorter in AML patients compared to the general population [25, 26]. Shortened TL in AML patients was closely associated with prognosis and survival [27]. Additionally, some studies have observed that longer TL was linked to an increased risk of developing AML, which aligned with our analysis. Longer TL promoted cell proliferation and viability, thereby increasing the risk of AML [28]. This suggested that TL may serve as a potential biomarker for assessing the risk of developing AML. Several studies have also explored the possibility of targeting TL for therapeutic purposes. For instance, interfering with telomerase activity inhibited AML cell proliferation and viability [29]. Further elucidation of the regulatory mechanisms of TL and AML, as well as the development of therapeutic strategies targeting TL, could facilitate early prevention and diagnosis of AML, ultimately improving patient prognosis and survival rates.
CLL is a mature B-cell lymphoma, and multiple factors influence its development and prognosis. Several studies have shown that CLL patients tend to have shorter average TL compared to normal controls [30–32]. CLL with shorter TL often exhibited specific cytogenetic abnormalities and cell surface markers associated with poor prognosis and faster disease progression [33]. However, an earlier analysis indicated that genetic variants associated with longer TL were linked to an increased risk of developing CLL [34]. This suggests that a baseline propensity for longer telomeres may provide opportunities for premalignant cells to undergo malignant transformation, while subsequent telomere depletion leads to TL shortening [35]. Although the sample size and the study design may influence the results, they were consistent with our analysis using the latest data. Therefore, TL may play a crucial role in the occurrence, development, and prognosis of CLL. The influence of telomeres on the pathogenesis of CLL was complex, underscoring the importance of unraveling the biological characteristics of telomeres at different stages of CLL.
HODGKIN is a malignant tumor of the lymphoid system, and its pathogenesis is not fully understood. Telomeres may play a role in the occurrence and development of HODGKIN. Studies have found that patients with HODGKIN may have shorter TL at diagnosis compared to healthy individuals [36]. HODGKIN also exhibited telomere dysfunction, activation of telomerase, and alternative lengthening of telomeres (ALT) [37, 38]. Telomere dysfunction in HODGKIN includes extremely short telomeres, altered telomere numbers, telomere aggregation, and changes in 3D spatial structure [39]. Furthermore, previous studies have shown that sequential inhibition of telomerase and ALT promotes the cell death in HODGKIN [40]. Our findings indicated that genetic variants associated with longer TL were associated with an increased risk of HODGKIN. In fact, several scholars have suggested that both shorter and longer telomeres may contribute to carcinogenesis, and that an optimal TL represented a balance between cell proliferation, senescence, and control [41–43]. Cells with longer TL had greater proliferative and mutational potential, leading to the onset of HODGKIN, while shortened TL was often correlated with disease severity after onset [44, 45].
MANTLE lymphoma is a non-Hodgkin lymphoma (NHL) originating from mature B cells, characterized by specific immunophenotype and recurrent genetic abnormalities [46]. Studies have shown that TERT promoter (TERTp) mutations leading to higher TERT expression levels were associated with longer telomeres in MANTLE, particularly in homozygous mutants; TERTp mutations may contribute to a more aggressive clinical behavior of MANTLE and could be associated with poorer prognosis [47]. A study on TL in 73 MANTLE and 20 normal B cell samples found highly variable TL in MANTLE (range, 2.2-13.8 kb; median, 4.3 kb) and significant telomeres dysfunction, but observed no association with any biological or clinical features [48]. Currently, research on telomere biology in MANTLE is limited, and our analysis predictive of genetic variations in TL has revealed a relationship with the risk of MANTLE onset, necessitating large-scale cohort studies to validate these findings.
An increasing body of evidence suggests that longer telomeres may be associated with an increased risk of NHL. A prospective study involving 107 male NHL cases and 107 matched controls found that longer relative TL may be linked to an increased risk of NHL [43]. Nested case-control studies conducted on 464 lymphoma cases and 464 matched controls from the EPIC cohort also revealed an association between longer TL and an increased risk of B-cell lymphoma, particularly in DLBCL and FOLLICULAR subtypes [49]. TL in B lymphocytes infected with Epstein-Barr virus gradually increased and was associated with the accumulation of early granulocyte leukemia bodies [50]. Another study demonstrated a positive correlation between genetically predicted longer TL and the risk of four NHL subtypes (CLL/SLL, DLBCL, FOLLICULAR, and MANTLE) [51]. Currently, there is a lack of research on the relationship between TL and the risk of CML, MONOCYTIC, and TNK. However, in CML patients, telomeres were often shortened [5]. In CML (BCR/ABL-positive), accelerated telomere shortening has been linked to the disease progression, risk score, and treatment response [52]. Even though our analysis has not found an association between genetically predicted TL and the risk of CML, DLBCL, FOLLICULAR, MARGINAL, MONOCYTIC, and TNK, when combined with previous research, there still seems to be an association between TL and the risk of certain diseases.
Despite the effective and stable results of our analysis, there were some limitations. Firstly, the sample size of the outcome factors was small, and the statistical efficacy could be further improved by conducting future studies with a larger sample size. Secondly, due to the limitation in sample size, our MR analysis failed to stratify specific factors such as age and sex, and only obtained summary-level statistics. Lastly, the individuals included in our analysis were of European populations, so caution should be exercised when generalizing these findings to other populations.
Conclusions
In conclusion, our findings indicate that a longer genetically predicted TL is associated with an increased risk of developing ALL, AML, CLL, MANTLE, and HODGKIN. However, there is no significant causal relationship between genetically predicted TL changes and the risk of developing CML, DLBCL, FOLLICULAR, MARGINAL, MONOCYTIC, and TNK. This study further supports the notion that cells with longer TL have greater proliferative and mutational potential, leading to an increased risk of certain HDs. Future research should focus on exploring the early diagnosis and therapeutic value of telomeres in patients with HDs.
Supplementary Materials
Author Contributions
YM W, QL, SL, and YF W were involved in the conception and design of the study. YM W and QL were responsible for visualization and article writing. QL, MY, and HZ were responsible for manuscript modification and discussion of the data analysis. YF W and DH provided scientific supervision. All authors reviewed and approved the final manuscript.
Acknowledgments
We would like to thank the GWAS summary data (https://gwas.mrcieu.ac.uk/) and FinnGen website (https://www.finngen.fi/en/) for the precious data used for free in scientific research.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
This work is supported by the Shandong Province Medicine and Health Science and Technology Development Project (No. 202103010371).
References
- 1. Armanios M. The Role of Telomeres in Human Disease. Annu Rev Genomics Hum Genet. 2022; 23:363–81. https://doi.org/10.1146/annurev-genom-010422-091101 [PubMed]
- 2. Cancer Prevention During Older Adulthood Writing Group. Highlights From an Expert Meeting on Opportunities for Cancer Prevention Among Older Adults. Gerontologist. 2019; 59:S94–101. https://doi.org/10.1093/geront/gnz037 [PubMed]
- 3. Merighi A, Gionchiglia N, Granato A, Lossi L. The Phosphorylated Form of the Histone H2AX (γH2AX) in the Brain from Embryonic Life to Old Age. Molecules. 2021; 26:7198. https://doi.org/10.3390/molecules26237198 [PubMed]
- 4. Peffault de Latour R, Calado RT, Busson M, Abrams J, Adoui N, Robin M, Larghero J, Dhedin N, Xhaard A, Clave E, Charron D, Toubert A, Loiseau P, et al. Age-adjusted recipient pretransplantation telomere length and treatment-related mortality after hematopoietic stem cell transplantation. Blood. 2012; 120:3353–9. https://doi.org/10.1182/blood-2012-01-403337 [PubMed]
- 5. Wang L, Xiao H, Zhang X, Wang C, Huang H. The role of telomeres and telomerase in hematologic malignancies and hematopoietic stem cell transplantation. J Hematol Oncol. 2014; 7:61. https://doi.org/10.1186/s13045-014-0061-9 [PubMed]
- 6. Wu KD, Orme LM, Shaughnessy J
Jr , Jacobson J, Barlogie B, Moore MA. Telomerase and telomere length in multiple myeloma: correlations with disease heterogeneity, cytogenetic status, and overall survival. Blood. 2003; 101:4982–9. https://doi.org/10.1182/blood-2002-11-3451 [PubMed] - 7. Gu R, Cao J, Wei S, Gong X, Wang Y, Mi Y, Zhang J, Qiu S, Rao Q, Wang M, Wei H, Wang J. Evaluation of pretreatment telomere length as a prognostic marker in intermediate-risk acute myeloid leukemia. Int J Lab Hematol. 2021; 43:1510–5. https://doi.org/10.1111/ijlh.13665 [PubMed]
- 8. Ventura Ferreira MS, Crysandt M, Ziegler P, Hummel S, Wilop S, Kirschner M, Schemionek M, Jost E, Wagner W, Brümmendorf TH, Beier F. Evidence for a pre-existing telomere deficit in non-clonal hematopoietic stem cells in patients with acute myeloid leukemia. Ann Hematol. 2017; 96:1457–61. https://doi.org/10.1007/s00277-017-3049-z [PubMed]
- 9. Gadalla SM, Wang T, Haagenson M, Spellman SR, Lee SJ, Williams KM, Wong JY, De Vivo I, Savage SA. Association between donor leukocyte telomere length and survival after unrelated allogeneic hematopoietic cell transplantation for severe aplastic anemia. JAMA. 2015; 313:594–602. https://doi.org/10.1001/jama.2015.7 [PubMed]
- 10. Harley CB. Telomerase and cancer therapeutics. Nat Rev Cancer. 2008; 8:167–79. https://doi.org/10.1038/nrc2275 [PubMed]
- 11. Mocellin S, Pooley KA, Nitti D. Telomerase and the search for the end of cancer. Trends Mol Med. 2013; 19:125–33. https://doi.org/10.1016/j.molmed.2012.11.006 [PubMed]
- 12. Duckworth A, Gibbons MA, Allen RJ, Almond H, Beaumont RN, Wood AR, Lunnon K, Lindsay MA, Wain LV, Tyrrell J, Scotton CJ. Telomere length and risk of idiopathic pulmonary fibrosis and chronic obstructive pulmonary disease: a mendelian randomisation study. Lancet Respir Med. 2021; 9:285–94. https://doi.org/10.1016/S2213-2600(20)30364-7 [PubMed]
- 13. Zhan Y, Song C, Karlsson R, Tillander A, Reynolds CA, Pedersen NL, Hägg S. Telomere Length Shortening and Alzheimer Disease--A Mendelian Randomization Study. JAMA Neurol. 2015; 72:1202–3. https://doi.org/10.1001/jamaneurol.2015.1513 [PubMed]
- 14. Haycock PC, Burgess S, Nounu A, Zheng J, Okoli GN, Bowden J, Wade KH, Timpson NJ, Evans DM, Willeit P, Aviv A, Gaunt TR, Hemani G, et al, and Telomeres Mendelian Randomization Collaboration. Association Between Telomere Length and Risk of Cancer and Non-Neoplastic Diseases: A Mendelian Randomization Study. JAMA Oncol. 2017; 3:636–51. https://doi.org/10.1001/jamaoncol.2016.5945 [PubMed]
- 15. Burgess S, Scott RA, Timpson NJ, Davey Smith G, Thompson SG, and EPIC- InterAct Consortium. Using published data in Mendelian randomization: a blueprint for efficient identification of causal risk factors. Eur J Epidemiol. 2015; 30:543–52. https://doi.org/10.1007/s10654-015-0011-z [PubMed]
- 16. Codd V, Wang Q, Allara E, Musicha C, Kaptoge S, Stoma S, Jiang T, Hamby SE, Braund PS, Bountziouka V, Budgeon CA, Denniff M, Swinfield C, et al. Polygenic basis and biomedical consequences of telomere length variation. Nat Genet. 2021; 53:1425–33. https://doi.org/10.1038/s41588-021-00944-6 [PubMed]
- 17. Huang D, Lin S, He J, Wang Q, Zhan Y. Association between COVID-19 and telomere length: A bidirectional Mendelian randomization study. J Med Virol. 2022; 94:5345–53. https://doi.org/10.1002/jmv.28008 [PubMed]
- 18. Yuan S, Kim JH, Xu P, Wang Z. Causal association between celiac disease and inflammatory bowel disease: A two-sample bidirectional Mendelian randomization study. Front Immunol. 2023; 13:1057253. https://doi.org/10.3389/fimmu.2022.1057253 [PubMed]
- 19. Inaba H, Mullighan CG. Pediatric acute lymphoblastic leukemia. Haematologica. 2020; 105:2524–39. https://doi.org/10.3324/haematol.2020.247031 [PubMed]
- 20. Nogueira BMD, da Costa Pantoja L, da Silva EL, Mello Júnior FAR, Teixeira EB, Wanderley AV, da Silva Maués JH, de Moraes Filho MO, de Moraes MEA, Montenegro RC, Khayat AS, Moreira-Nunes CA. Telomerase (hTERT) Overexpression Reveals a Promising Prognostic Biomarker and Therapeutical Target in Different Clinical Subtypes of Pediatric Acute Lymphoblastic Leukaemia. Genes (Basel). 2021; 12:1632. https://doi.org/10.3390/genes12101632 [PubMed]
- 21. Karow A, Haubitz M, Oppliger Leibundgut E, Helsen I, Preising N, Steiner D, Dantonello TM, Ammann RA, Roessler J, Kartal-Kaess M, Röth A, Baerlocher GM. Targeting Telomere Biology in Acute Lymphoblastic Leukemia. Int J Mol Sci. 2021; 22:6653. https://doi.org/10.3390/ijms22136653 [PubMed]
- 22. Borssén M, Cullman I, Norén-Nyström U, Sundström C, Porwit A, Forestier E, Roos G. hTERT promoter methylation and telomere length in childhood acute lymphoblastic leukemia: associations with immunophenotype and cytogenetic subgroup. Exp Hematol. 2011; 39:1144–51. https://doi.org/10.1016/j.exphem.2011.08.014 [PubMed]
- 23. da Mota THA, Camargo R, Biojone ER, Guimarães AFR, Pittella-Silva F, de Oliveira DM. The Relevance of Telomerase and Telomere-Associated Proteins in B-Acute Lymphoblastic Leukemia. Genes (Basel). 2023; 14:691. https://doi.org/10.3390/genes14030691 [PubMed]
- 24. Ley TJ, Miller C, Ding L, Raphael BJ, Mungall AJ, Robertson A, Hoadley K, Triche TJ
Jr , Laird PW, Baty JD, Fulton LL, Fulton R, Heath SE, et al, and Cancer Genome Atlas Research Network. Genomic and epigenomic landscapes of adult de novo acute myeloid leukemia. N Engl J Med. 2013; 368:2059–74. https://doi.org/10.1056/NEJMoa1301689 [PubMed] - 25. de Oliveira Lisboa M, Brofman PRS, Schmid-Braz AT, Rangel-Pozzo A, Mai S. Chromosomal Instability in Acute Myeloid Leukemia. Cancers (Basel). 2021; 13:2655. https://doi.org/10.3390/cancers13112655 [PubMed]
- 26. Dratwa M, Wysoczańska B, Butrym A, Łacina P, Mazur G, Bogunia-Kubik K. TERT genetic variability and telomere length as factors affecting survival and risk in acute myeloid leukaemia. Sci Rep. 2021; 11:23301. https://doi.org/10.1038/s41598-021-02767-1 [PubMed]
- 27. Ernst P, Heidel FH. Molecular Mechanisms of Senescence and Implications for the Treatment of Myeloid Malignancies. Cancers (Basel). 2021; 13:612. https://doi.org/10.3390/cancers13040612 [PubMed]
- 28. Sullivan SM, Cole B, Lane J, Meredith JJ, Langer E, Hooten AJ, Roesler M, McGraw KL, Pankratz N, Poynter JN. Predicted leukocyte telomere length and risk of myeloid neoplasms. Hum Mol Genet. 2023; 32:2996–3005. https://doi.org/10.1093/hmg/ddad126 [PubMed]
- 29. Kim JJ, Sayed ME, Ahn A, Slusher AL, Ying JY, Ludlow AT. Dynamics of TERT regulation via alternative splicing in stem cells and cancer cells. PLoS One. 2023; 18:e0289327. https://doi.org/10.1371/journal.pone.0289327 [PubMed]
- 30. Hoxha M, Fabris S, Agnelli L, Bollati V, Cutrona G, Matis S, Recchia AG, Gentile M, Cortelezzi A, Morabito F, Bertazzi PA, Ferrarini M, Neri A. Relevance of telomere/telomerase system impairment in early stage chronic lymphocytic leukemia. Genes Chromosomes Cancer. 2014; 53:612–21. https://doi.org/10.1002/gcc.22171 [PubMed]
- 31. Augereau A, T'kint de Roodenbeke C, Simonet T, Bauwens S, Horard B, Callanan M, Leroux D, Jallades L, Salles G, Gilson E, Poncet D. Telomeric damage in early stage of chronic lymphocytic leukemia correlates with shelterin dysregulation. Blood. 2011; 118:1316–22. https://doi.org/10.1182/blood-2010-07-295774 [PubMed]
- 32. Rossi D, Lobetti Bodoni C, Genuardi E, Monitillo L, Drandi D, Cerri M, Deambrogi C, Ricca I, Rocci A, Ferrero S, Bernocco E, Capello D, De Paoli L, et al. Telomere length is an independent predictor of survival, treatment requirement and Richter's syndrome transformation in chronic lymphocytic leukemia. Leukemia. 2009; 23:1062–72. https://doi.org/10.1038/leu.2008.399 [PubMed]
- 33. Gramatges MM, Bertuch AA. Short telomeres: from dyskeratosis congenita to sporadic aplastic anemia and malignancy. Transl Res. 2013; 162:353–63. https://doi.org/10.1016/j.trsl.2013.05.003 [PubMed]
- 34. Ojha J, Codd V, Nelson CP, Samani NJ, Smirnov IV, Madsen NR, Hansen HM, de Smith AJ, Bracci PM, Wiencke JK, Wrensch MR, Wiemels JL, Walsh KM, and ENGAGE Consortium Telomere Group. Genetic Variation Associated with Longer Telomere Length Increases Risk of Chronic Lymphocytic Leukemia. Cancer Epidemiol Biomarkers Prev. 2016; 25:1043–9. https://doi.org/10.1158/1055-9965.EPI-15-1329 [PubMed]
- 35. Sellmann L, de Beer D, Bartels M, Opalka B, Nückel H, Dührsen U, Dürig J, Seifert M, Siemer D, Küppers R, Baerlocher GM, Röth A. Telomeres and prognosis in patients with chronic lymphocytic leukaemia. Int J Hematol. 2011; 93:74–82. https://doi.org/10.1007/s12185-010-0750-2 [PubMed]
- 36. M'kacher R, Bennaceur-Griscelli A, Girinsky T, Koscielny S, Delhommeau F, Dossou J, Violot D, Leclercq E, Courtier MH, Béron-Gaillard N, Assaf E, Ribrag V, Bourhis J, et al. Telomere shortening and associated chromosomal instability in peripheral blood lymphocytes of patients with Hodgkin's lymphoma prior to any treatment are predictive of second cancers. Int J Radiat Oncol Biol Phys. 2007; 68:465–71. https://doi.org/10.1016/j.ijrobp.2007.01.050 [PubMed]
- 37. Knecht H, Sawan B, Lichtensztejn D, Lemieux B, Wellinger RJ, Mai S. The 3D nuclear organization of telomeres marks the transition from Hodgkin to Reed-Sternberg cells. Leukemia. 2009; 23:565–73. https://doi.org/10.1038/leu.2008.314 [PubMed]
- 38. M'kacher R, Cuceu C, Al Jawhari M, Morat L, Frenzel M, Shim G, Lenain A, Hempel WM, Junker S, Girinsky T, Colicchio B, Dieterlen A, Heidingsfelder L, et al. The Transition between Telomerase and ALT Mechanisms in Hodgkin Lymphoma and Its Predictive Value in Clinical Outcomes. Cancers (Basel). 2018; 10:169. https://doi.org/10.3390/cancers10060169 [PubMed]
- 39. Knecht H, Kongruttanachok N, Sawan B, Brossard J, Prévost S, Turcotte E, Lichtensztejn Z, Lichtensztejn D, Mai S. Three-dimensional Telomere Signatures of Hodgkin- and Reed-Sternberg Cells at Diagnosis Identify Patients with Poor Response to Conventional Chemotherapy. Transl Oncol. 2012; 5:269–77. https://doi.org/10.1593/tlo.12142 [PubMed]
- 40. Lima MF, Freitas MO, Hamedani MK, Rangel-Pozzo A, Zhu XD, Mai S. Consecutive Inhibition of Telomerase and Alternative Lengthening Pathway Promotes Hodgkin's Lymphoma Cell Death. Biomedicines. 2022; 10:2299. https://doi.org/10.3390/biomedicines10092299 [PubMed]
- 41. Seow WJ, Cawthon RM, Purdue MP, Hu W, Gao YT, Huang WY, Weinstein SJ, Ji BT, Virtamo J, Hosgood HD
3rd , Bassig BA, Shu XO, Cai Q, et al. Telomere length in white blood cell DNA and lung cancer: a pooled analysis of three prospective cohorts. Cancer Res. 2014; 74:4090–8. https://doi.org/10.1158/0008-5472.CAN-14-0459 [PubMed] - 42. Harris BHL, Macaulay VM, Harris DA, Klenerman P, Karpe F, Lord SR, Harris AL, Buffa FM. Obesity: a perfect storm for carcinogenesis. Cancer Metastasis Rev. 2022; 41:491–515. https://doi.org/10.1007/s10555-022-10046-2 [PubMed]
- 43. Lan Q, Cawthon R, Shen M, Weinstein SJ, Virtamo J, Lim U, Hosgood HD
3rd , Albanes D, Rothman N. A prospective study of telomere length measured by monochrome multiplex quantitative PCR and risk of non-Hodgkin lymphoma. Clin Cancer Res. 2009; 15:7429–33. https://doi.org/10.1158/1078-0432.CCR-09-0845 [PubMed] - 44. Rybicki BA, Sadasivan SM, Chen Y, Loveless I, Gupta NS, Chitale DA, Williamson SR, Rundle AG, Tang DL. Race Differences in Telomere Length in Benign Prostate Biopsies and Subsequent Risk of Prostate Cancer. Cancer Epidemiol Biomarkers Prev. 2022; 31:991–8. https://doi.org/10.1158/1055-9965.EPI-21-1221 [PubMed]
- 45. Holesova Z, Krasnicanova L, Saade R, Pös O, Budis J, Gazdarica J, Repiska V, Szemes T. Telomere Length Changes in Cancer: Insights on Carcinogenesis and Potential for Non-Invasive Diagnostic Strategies. Genes (Basel). 2023; 14:715. https://doi.org/10.3390/genes14030715 [PubMed]
- 46. Armitage JO, Longo DL. Mantle-Cell Lymphoma. N Engl J Med. 2022; 386:2495–506. https://doi.org/10.1056/NEJMra2202672 [PubMed]
- 47. Panero J, Alves-Paiva RM, Roisman A, Santana-Lemos BA, Falcão RP, Oliveira G, Martins D, Stanganelli C, Slavutsky I, Calado RT. Acquired TERT promoter mutations stimulate TERT transcription in mantle cell lymphoma. Am J Hematol. 2016; 91:481–5. https://doi.org/10.1002/ajh.24324 [PubMed]
- 48. Jebaraj BM, Kienle D, Lechel A, Mertens D, Heuberger M, Ott G, Rosenwald A, Barth TF, Möller P, Zenz T, Döhner H, Stilgenbauer S. Telomere length in mantle cell lymphoma. Blood. 2013; 121:1184–7. https://doi.org/10.1182/blood-2012-08-452649 [PubMed]
- 49. Hosnijeh FS, Matullo G, Russo A, Guarrera S, Modica F, Nieters A, Overvad K, Guldberg P, Tjønneland A, Canzian F, Boeing H, Aleksandrova K, Trichopoulou A, et al. Prediagnostic telomere length and risk of B-cell lymphoma-Results from the EPIC cohort study. Int J Cancer. 2014; 135:2910–7. https://doi.org/10.1002/ijc.28934 [PubMed]
- 50. Kamranvar SA, Chen X, Masucci MG. Telomere dysfunction and activation of alternative lengthening of telomeres in B-lymphocytes infected by Epstein-Barr virus. Oncogene. 2013; 32:5522–30. https://doi.org/10.1038/onc.2013.189 [PubMed]
- 51. Machiela MJ, Lan Q, Slager SL, Vermeulen RC, Teras LR, Camp NJ, Cerhan JR, Spinelli JJ, Wang SS, Nieters A, Vijai J, Yeager M, Wang Z, et al. Genetically predicted longer telomere length is associated with increased risk of B-cell lymphoma subtypes. Hum Mol Genet. 2016; 25:1663–76. https://doi.org/10.1093/hmg/ddw027 [PubMed]
- 52. Braig M, Pällmann N, Preukschas M, Steinemann D, Hofmann W, Gompf A, Streichert T, Braunschweig T, Copland M, Rudolph KL, Bokemeyer C, Koschmieder S, Schuppert A, et al. A 'telomere-associated secretory phenotype' cooperates with BCR-ABL to drive malignant proliferation of leukemic cells. Leukemia. 2014; 28:2028–39. https://doi.org/10.1038/leu.2014.95 [PubMed]