


Introduction
Breast cancer poses a significant global health challenge [1] and continues to be one of the most prevalent cancers worldwide, with 2.26 million reported cases in 2020. In addition, it is the main factor affecting female cancer-related fatalities [2]. Invasive breast cancer (IBC) is the most prevalent type of breast cancer-related mortality. Although the prognosis has improved for most patients with IBC, inter-individual heterogeneity can still result in a poor prognosis for some patients [3]. Therefore, it is imperative to identify novel biomarkers as soon as possible to improve IBC patient diagnosis and therapy.
When cells separate from the extracellular matrix, a type of programmed cell death known as anoikis occurs. [4]. In normal cells, this process functions as a protective mechanism that prevents the survival and proliferation of cells detached from their normal tissue architecture. However, in cancer cells, resistance to anoikis is a crucial step in acquiring metastatic potential [5]. Cancer cells typically separate from the original tumor and infiltrate the neighboring tissues as the tumor grows. These cells must resist anoikis and survive in the absence of attachment to the extracellular matrix [4]. Anoikis resistance can be mediated by various mechanisms, including alterations in cell-surface receptor expression, activation of survival pathways, and changes in the expression of pro- and anti-apoptotic proteins [6]. According to current findings, anoikis is critical for the development of gastric cancer [7], esophageal squamous cell carcinoma [8], and breast cancer [9].
Long noncoding RNAs (lncRNAs) are involved in various biological activities and play crucial roles in the regulation of gene expression [10]. Dysregulated expression of lncRNAs has been observed in cancer, where they function as oncogenes or tumor suppressors [11]. The mechanisms by which lncRNAs regulate cancer development are diverse and complex and involve interactions with DNA, RNA, and proteins [12]. An enhanced understanding of lncRNA functions in tumor biology may pave the way for innovative cancer therapies. However, the precise roles of anoikis-related lncRNAs (ARLs) in invasive breast cancer have not yet been fully elucidated.
In the present study, we systematically explored the role of ARLs in IBC. We identified IBC-related ARLs and created a prognostic model that can direct prognostic predictions and clinical treatment decisions. Furthermore, we thoroughly examined a key ARL prognostic gene, AL133467.1, to validate its association with migration, proliferative capacity, and anoikis in breast cancer cells. By uncovering the molecular mechanisms underlying IBC progression, our findings offer valuable insights and present a potential novel strategy for the diagnosis and treatment of this disease.
Results
Identification of anoikis-related differentially expressed LncRNAs
We identified 1002 differentially expressed lncRNAs (DELs) between normal breast and IBC samples. In a previous study, 434 anoikis-related genes were identified [13]. We then performed co-expression analysis to identify 1951 lncRNAs that are co-expressed with anoikis-related genes. The 1002 DELs intersected with the 1951 co-expressed lncRNAs, resulting in the selection of 110 anoikis-related DELs (Figure 1A). The co-expression network of anoikis-related DELs and genes is shown in Figure 1B.
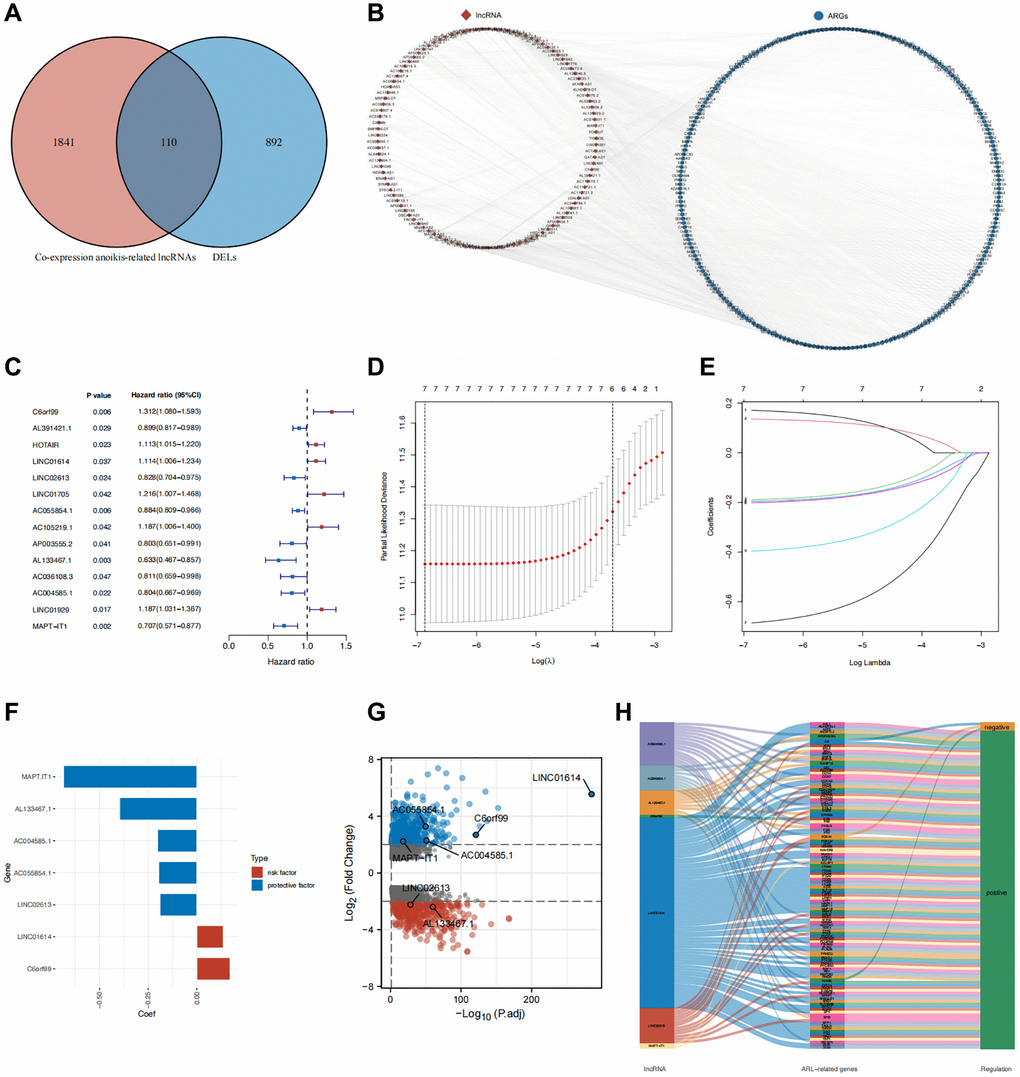
Application and validation of the prognostic model
Patients in both sets were further classified into high and low ARL score groups based on the median ARL score. The ARL score distribution, survival status, and expression levels of the seven core lncRNAs in the low- and high ARL score groups are shown in Figure 2A–2C. These results illustrate the intuitive differences between the two groups in the training, testing, and entire sets, which were supported by Principal Component Analysis (PCA) (Figure 2D–2F).
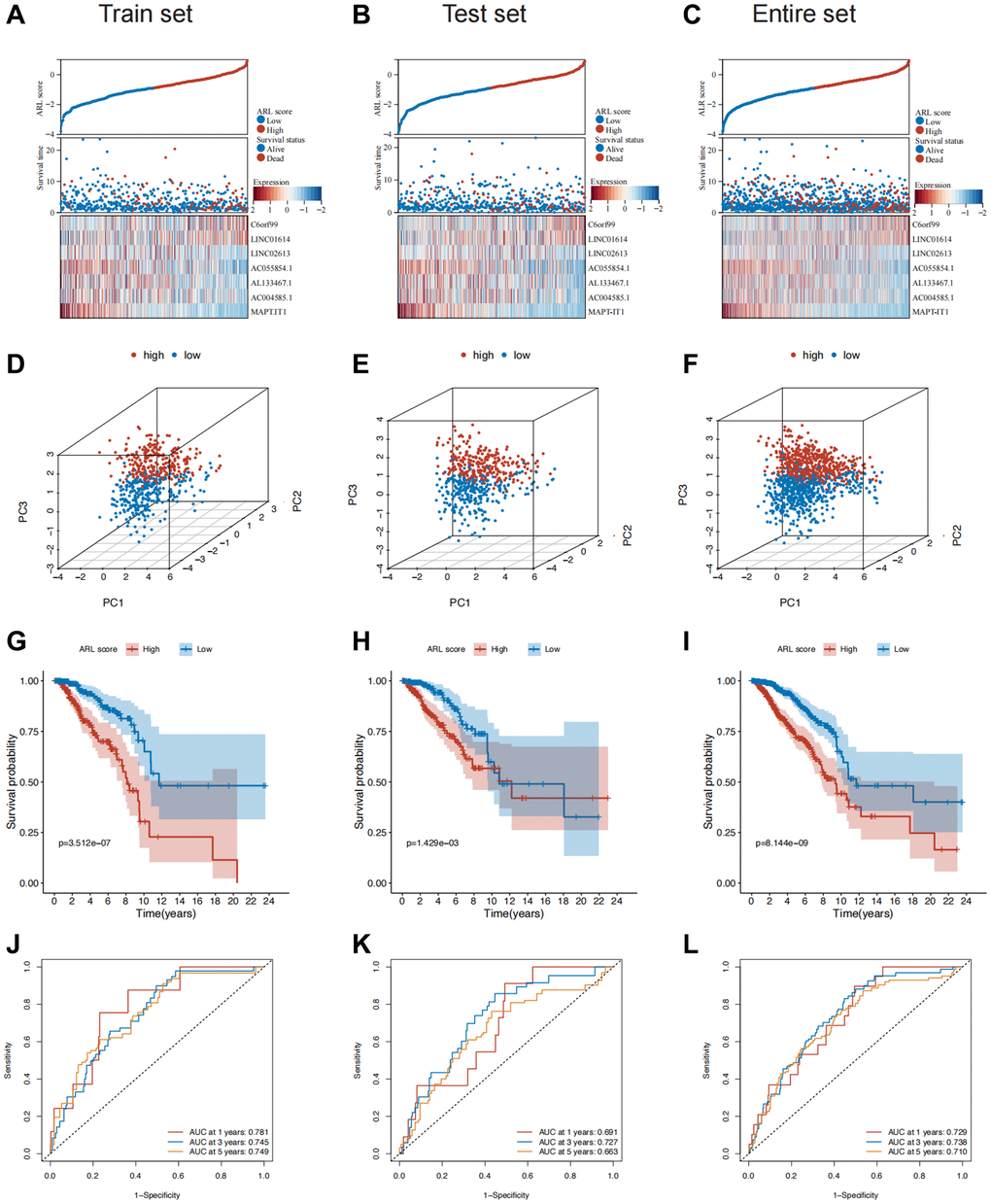
Moreover, the low ARL score group exhibited significantly better overall survival (OS) compared with the high ARL score group (Figure 2G–2I). This trend was also observed for other clinical characteristics such as TNM stage, grade, and age (Supplementary Figure 2A). The 1-, 3-, and 5-year area under the curve (AUC) values of the prognostic model were 0.729, 0.738, and 0.710, respectively (Figure 2J–2L). In addition, univariate Cox regression analysis showed that the N stage, M stage, pathological stage, and ARL score were predictors of survival in the training set, test set, and entire set, whereas multivariate analysis indicated that the ARL score was the only common independent predictive factor for breast cancer patients across all sets (Supplementary Figure 3). These findings suggest that the model has a relatively good predictive ability for prognosis.
Correlation between clinical characteristics and the prognostic model
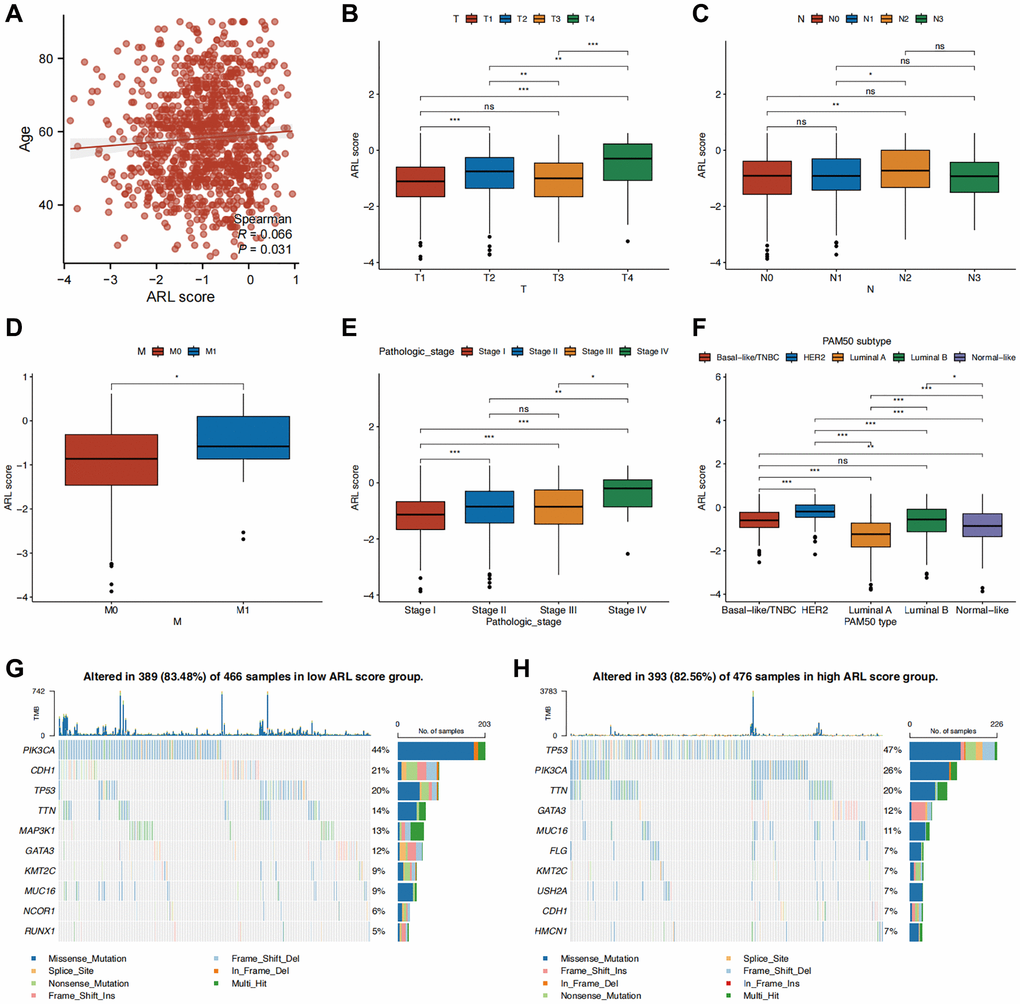
Additionally, we investigated the clinical significance of a prognostic model utilizing ARLs by assessing their association with clinical features. As shown in Figure 3A, a significant correlation was observed between age and the ARL score (P < 0.05). Individuals diagnosed with T4, N2, M1, and stage IV conditions demonstrated the most elevated ARL scores, signifying that elevated ARL scores are associated with advanced TNM and pathological stages. (Figure 3B–3E) (P < 0.05). Among the PAM50 breast cancer subtypes, patients with HER2-enriched breast cancer had the highest ARL scores (P < 0.001), whereas those with luminal A breast cancer had the lowest scores (Figure 3F) (P < 0.05). Interestingly, the frequency of PIK3CA gene mutation was highest (44%) in the group with a low ARL score, in contrast to a mutation rate of 26% in the group with a high ARL score. In the high ARL score group, TP53 emerged as the gene with the highest occurrence of mutations, with a frequency of 47%, whereas the low ARL score group had a mutation rate of only 20% (Figure 3G, 3H). Furthermore, to confirm that the ARL score could differentiate patients with varying prognoses across different breast cancer subtypes, we conducted survival analyses stratified by the ARL score within each breast cancer subtype. The results of our study suggest that patients with low ARL scores consistently had better prognoses (P < 0.05) in all four breast cancer subtypes, namely, basal-like/TNBC, luminal A, luminal B, and normal-like. Although a comparable pattern was observed in patients with HER2-amplified breast cancer, the difference was not statistically significant (P > 0.05), possibly because of the small number of participants in this specific patient cohort (Supplementary Figure 2B). These findings suggest the potential utility of the ARL-based prognostic model in aiding the diagnosis, prognostication, and formulation of treatment strategies.
Developing the nomogram to predict the outcome of IBC
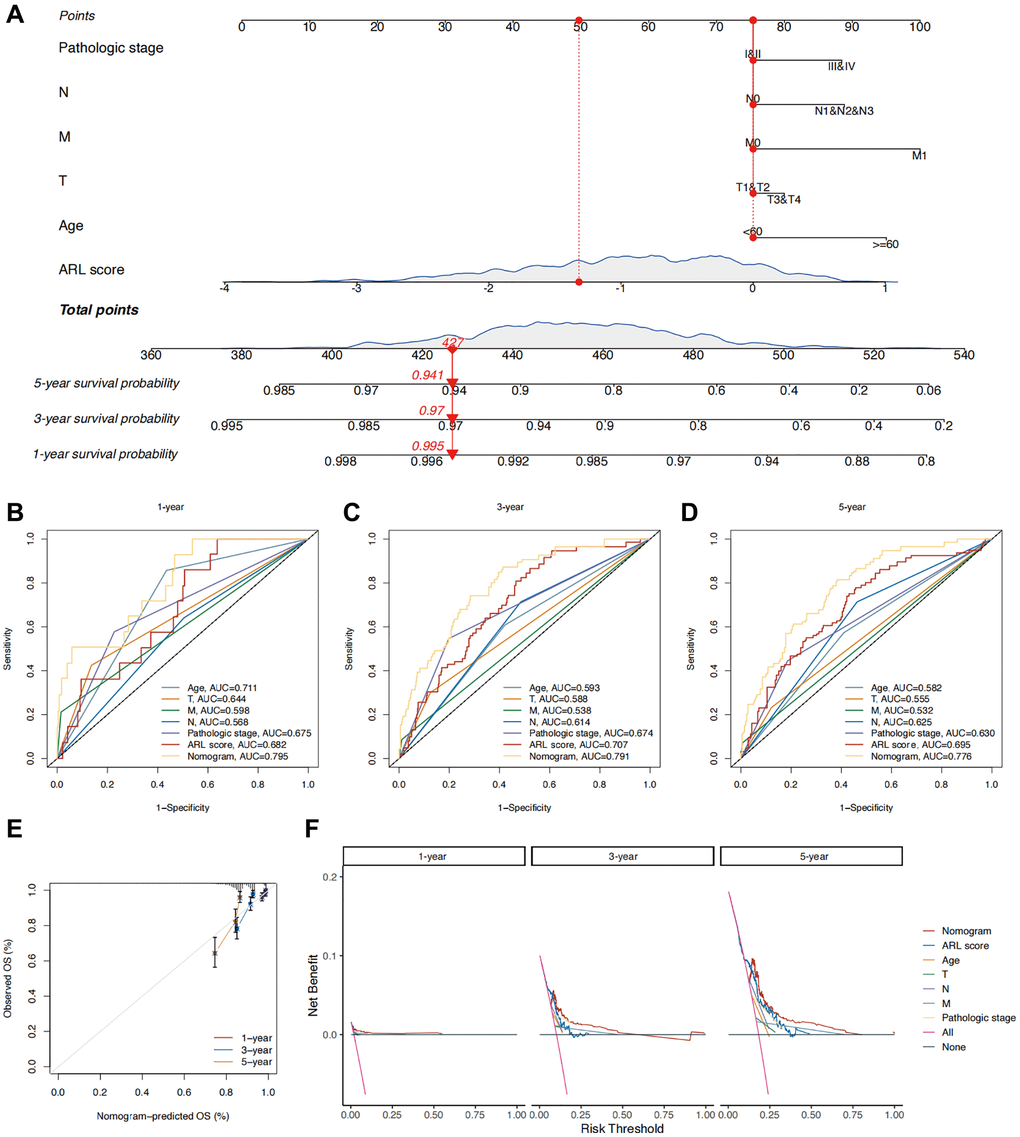
To improve the practicality of the prognostic model, the ARL scores were integrated with prognostic clinical characteristics to develop a nomogram that predicts the overall survival (OS) of patients with IBC (Figure 4A). To assess the superiority of the nomogram in predicting outcomes, various parameters, such as age, T stage, N stage, M stage, pathological stage, and ARL score, were evaluated using a Receiver Operating Characteristic (ROC) curve. As shown in Figure 4B–4D, the AUC values for predicting the 1-, 3-, and 5-year outcomes using the nomogram (0.795, 0.791, and 0.776, respectively) were higher compared to age (0.711, 0.593, and 0.582, respectively), T stage (0.644, 0.588, and 0.555, respectively), N stage (0.568, 0.614, and 0.625, respectively), M stage (0.598, 0.538, and 0.532, respectively), pathological stage (0.675, 0.674, and 0.630, respectively), and ARL scores (0.682, 0.707, and 0.695, respectively). This indicates the strong ability of the nomogram to predict the prognosis of IBC. Figure 4E shows a calibration plot, indicating that the nomogram exhibited a favorable capacity for predicting patient prognosis. Based on the decision curve analysis (DCA) results for 1-, 3-, and 5-year periods, it can be inferred from Figure 4F that the nomogram exhibited superior clinical usefulness compared to alternative factors. The nomogram prediction model, which is based on ARL scores and clinicopathological features, has been thoroughly validated using various approaches and has demonstrated robust predictive capacity and clinical usefulness.
Function enrichment analysis
To clarify the possible biological roles and pathways linked to the ARL signature, we performed various enrichment analyses such as GSEA, GSVA, GO, and KEGG. The results of the GO and KEGG enrichment analyses in Figure 5A show a significant correlation between differentially expressed genes (DEGs) in the low and high ARL score groups and signal transduction-related biological functions, including channel activity, passive transmembrane transporter activity, anion transmembrane transporter activity, hormone activity, and receptor ligand activity.
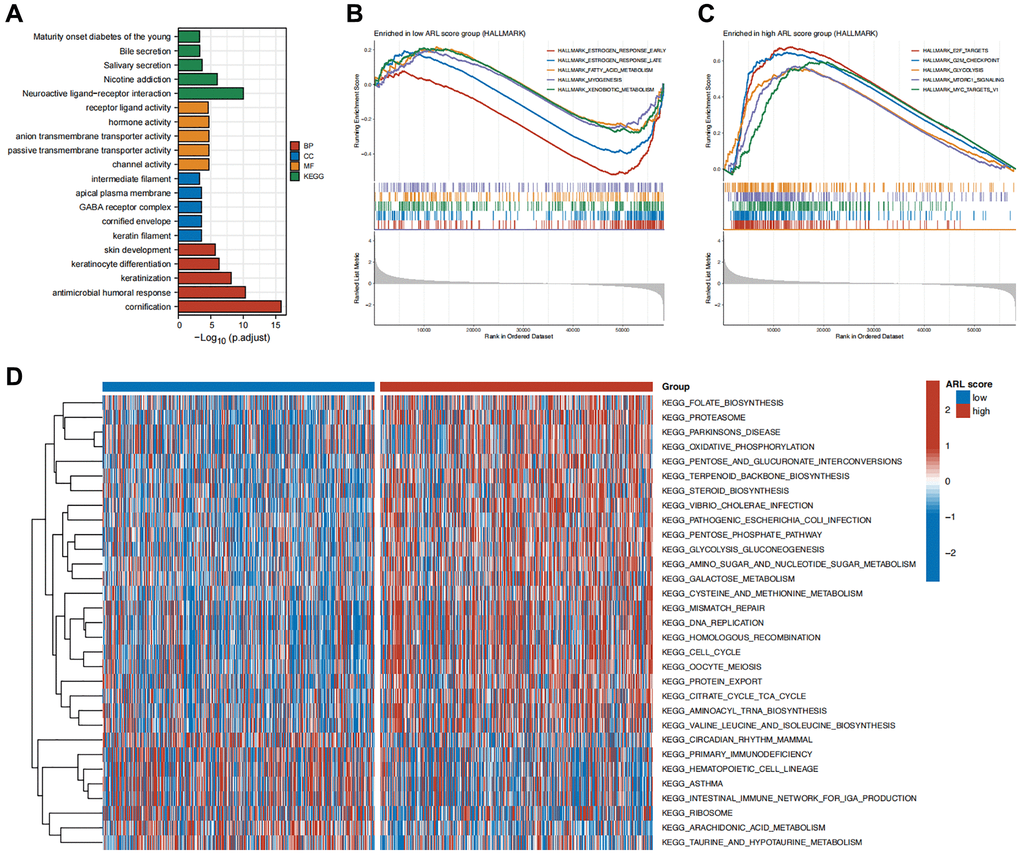
Additionally, we conducted GSEA enrichment analysis to compare the groups with low and high ARL scores and identify distinct biological functions and pathways between the two groups. Figure 5B, 5C illustrate that the group with low ARL scores had a notable enhancement in biological processes, including early estrogen response, late estrogen response, fatty acid metabolism, muscle formation, and xenobiotic metabolism. In contrast, the group with high ARL scores exhibited notable enrichment in pathways including E2F targets, G2/M checkpoint, glycolysis, MTORC1 signaling, and MYC targets.
Furthermore, GSVA revealed 31 distinct pathways that differed between IBC patients with low and high ARL scores. Patients with high ARL scores exhibited positive associations with pathways such as mismatch repair, DNA replication, homologous recombination, and the cell cycle, as shown in Figure 5D, whereas the low ARL score group had negative associations. These results indicated that these physiological processes may be linked to poor outcomes in patients with IBC.
Immune infiltration characteristics of the TME
In light of mounting evidence that immune infiltration characteristics have a major impact on the progression and development of breast cancer, we accessed the relationship between immune infiltration characteristics and ARL scores. As shown in Figure 6A, the low ARL score group exhibited significantly higher infiltration fractions of immune cells (P < 0.05). Furthermore, Figure 6B demonstrates that immune-related pathways, including cytolytic activity, human leukocyte antigen (HLA), T cell co-stimulation, and type II IFN response, were highly enriched in the low ARL score group, whereas only the type I IFN response was highly enriched in the high ARL score group (P < 0.05). Additionally, the low ARL score group exhibited significantly higher immune, stromal, and estimated scores than the high ARL score group (P < 0.05), as depicted in Figure 6C.
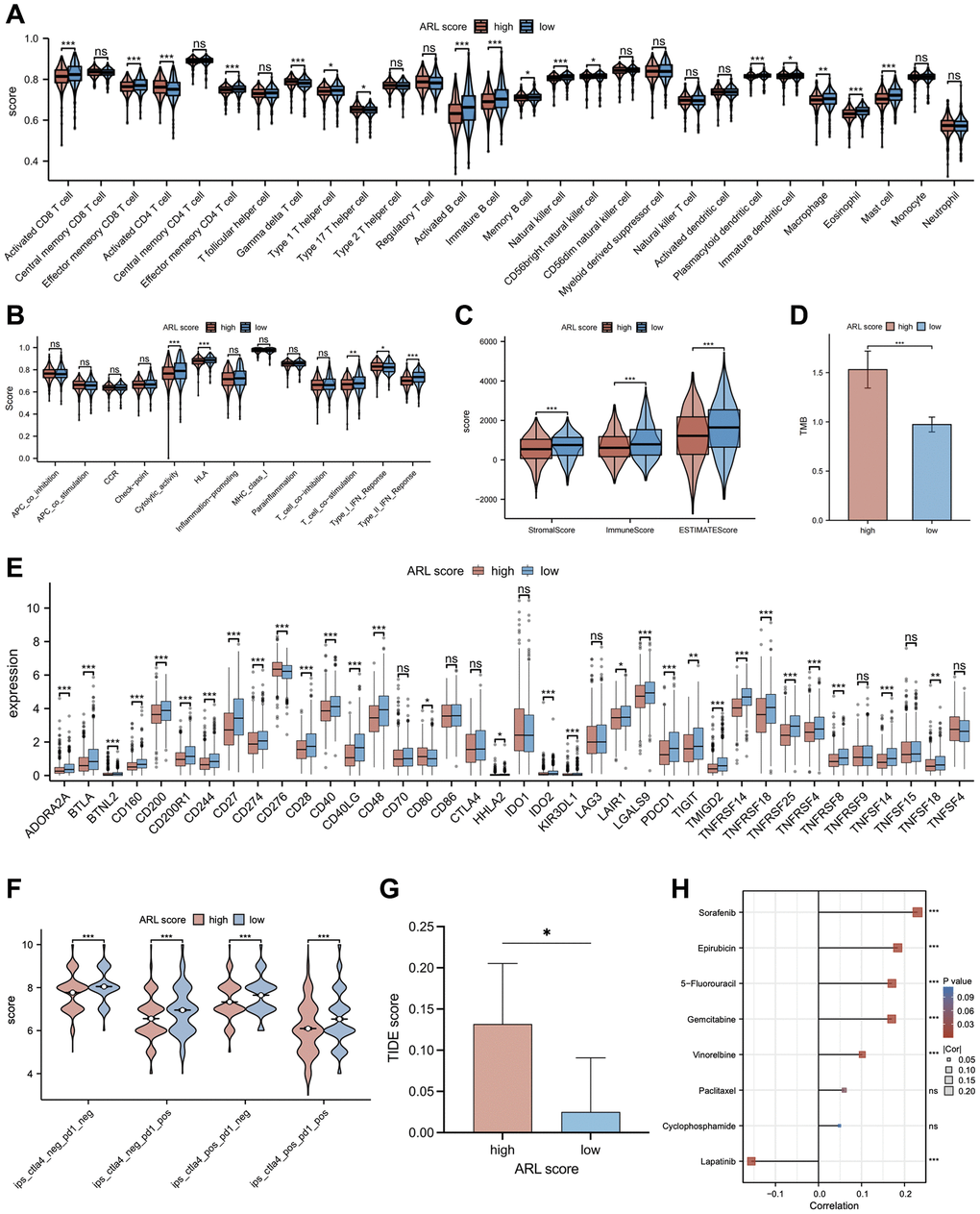
To examine the relationship between ARL scores and the degree of immune cell infiltration in various breast cancer subtypes, we extended our analysis to include immune cell infiltration across five subtypes: basal-like/TNBC, HER2-enriched, luminal A, luminal B, and normal-like (Supplementary Figure 4). Despite the distinct immune cell infiltration characteristics among the four breast subtypes (basal-like/TNBC, HER2-enriched, luminal A, and normal-like), our data demonstrated that patients with low ARL scores invariably exhibited higher levels of immune cell infiltration (P < 0.05). Interestingly, patients with luminal B breast cancer deviated from this trend. An increase in the infiltration levels of certain immune cells, including central memory CD8 T cells, gamma delta T cells, type 17 T helper cells, CD56dim natural killer cells, and neutrophils, was significantly correlated with high ARL scores (P < 0.05). These findings underscore the fact that ARL scores can differentiate immune cell infiltration profiles among different breast cancer subtypes, which may aid the development of targeted treatments in the future.
Immunotherapy response and chemotherapy sensitivity
Recently, immunotherapy and targeted therapy have played important roles in the clinical treatment of breast cancer and have become popular research topics. Therefore, a comparison was made between individuals with IBC who had low versus high ARL scores to determine the differences in responses to immune-targeted therapy. As shown in Figure 6E, most of the immune checkpoint candidate genes, such as PD-L1, were expressed at higher levels in the low ARL score group (P < 0.05). In addition, the results of immunogenomic analyses from The Cancer Immunome Atlas (TCIA) demonstrated that patients in the low ARL score group exhibited better responses to immunotherapy than patients in the high ARL score group, particularly when using CTLA4 targeted therapy alone, PD1 targeted therapy alone, or combined PD1 and CTLA4 targeted therapy, as well as targeted therapy excluding PD1 and CTLA4 (Figure 6F) (P < 0.05). Moreover, Tumor Immune Dysfunction and Exclusion (TIDE) analysis showed that patients in the low ARL score group had lower TIDE scores than patients in the high ARL score group, implying that those with low ARL scores may had better immunotherapy responses at PD-L1/CTLA4 immune checkpoint inhibitors (Figure 6G) (P < 0.05). Moreover, individuals with low ARL scores had lower tumor mutation burdens (TMBs) than patients with high ARL scores (Figure 6D) (P < 0.05). As chemotherapy plays an important role in the pre- and post-surgical treatment of patients with breast cancer, the role of the ARL score in chemotherapy for patients with IBC was explored in this study. Figure 6H illustrates the correlation between ARL scores and the half-maximal inhibitory concentration (IC50) of specific breast cancer chemotherapeutics. The IC50 values of chemotherapeutic drugs, including sorafenib, epirubicin, 5-fluorouracil, gemcitabine, and vinorelbine, were positively correlated with the ARL score (P < 0.05), whereas that of lapatinib was negatively correlated with the ARL score (P < 0.05). These findings suggest that individuals with different ARL scores may benefit from appropriate chemotherapeutic drugs tailored to their ARL, thereby potentially improving the prognosis of patients with IBC.
Validation of the expression levels of the 7 prognostic LncRNAs
To analyze the expression profiles in clinical samples, we investigated the expression levels of the seven core prognostic lncRNAs in both TCGA cohort and clinical samples. As depicted in Supplementary Figure 5A, 5B, C60rf99, LINC01614, AC004585, MAPT-IT1, and AC004585.1 exhibited significantly higher expression levels in breast tumor tissues than in paired or unpaired normal breast tissues (P < 0.05). In contrast, LINC02613 and AL133467.1 displayed significantly lower expression levels in breast tumor tissues than in paired or unpaired normal breast tissues (P < 0.05). Additionally, quantitative reverse transcription-polymerase chain reaction (qRT-PCR) analysis revealed that the expression levels of C60rf99, LINC02613, AL133467.1, MAPT-IT1, and AC004585.1 were downregulated in breast tumor tissues relative to normal breast tissues (P < 0.05) (Supplementary Figure 5C). However, no significant difference was observed in the expression levels of LINC01614 and AC055854.1 between tumor and normal clinical samples (P > 0.05) (Supplementary Figure 5C).
Effect of AL133467.1 expression in pan-cancer
The expression levels of AL133467.1 were investigated across a range of cancers by contrasting tumorous and corresponding normal tissues. Notably, AL133467.1 demonstrated elevated expression in lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC) (P < 0.05). In stark contrast, AL133467.1 displayed significantly reduced expression in colon adenocarcinoma (COAD), colon adenocarcinoma/rectum adenocarcinoma esophageal carcinoma (COADREAD), invasive breast carcinoma (BRCA), kidney renal papillary cell carcinoma (KIRP), liver hepatocellular carcinoma (LIHC), thyroid carcinoma (THCA), rectum adenocarcinoma (READ), bladder urothelial carcinoma (BLCA), kidney chromophobe (KICH), and cholangiocarcinoma (CHOL) (P < 0.05) as depicted in Supplementary Figure 6A). The expression of AL133467.1 did not differ significantly between normal and tumorous tissues in the other 14 cancer types.
Survival analyses were conducted for each cancer type to assess the role of AL133467.1 in the prognosis of patients with different cancer types. A univariate Cox analysis revealed a significant association between AL133467.1 levels and overall survival (OS) outcomes in pan-kidney (KIPAN; P = 3.8e-4), glioblastoma multiforme (GBM; P = 0.05), kidney renal clear cell carcinoma (KIRC; P = 0.03), COADREAD (P = 0.03), KICH (P = 6.8e-5), BRCA (P = 0.01), head and neck squamous cell carcinoma (HNSC; P = 0.02), skin cutaneous melanoma (SKCM; P = 0.02), SKCM-M (P = 0.02), and ovarian serous cystadenocarcinoma (OV; P = 2.3e-3), as shown in Supplementary Figure 6B. These findings suggest that AL133467.1 could be considered to be a high-risk gene in KIPAN, GBM, KIRC, COADREAD, and KICH but a low-risk gene in BRCA, HNSC, SKCM-M, SKCM, and OV.
We employed an additional correlation analysis to elucidate the relationship between AL133467.1 expression levels and immune-related markers across diverse types of cancer. As depicted in Supplementary Figure 6C, AL133467.1 manifested positive correlation with both immune inhibitory and stimulatory markers across most cancer types. The correlation between AL133467.1 expression and various cell infiltration levels within the tumor microenvironment was subsequently investigated. Our findings demonstrate that an increase in AL133467.1 expression considerably amplified the infiltration levels of numerous cells within the microenvironment across most tumor types. As indicated in Supplementary Figure 6D, cells such as aDC, B cells, CD4+ memory T cells, CD4+ naive T cells, CD8+ T cells, CD8+ Tcm, CD8+ Tem, cDC, chondrocytes, class-switched memory B cells, DC, hematopoietic stem cells (HSCs), iDC, M2 macrophages, M1 macrophages, mast cells, megakaryocytes, melanocytes, memory B cells, monocytes, multipotent progenitor (MPP) cells, endothelial cells, naive B cells, pDCs, plasma cells, and Tgd cells demonstrated a positive correlation with heightened AL133467.1 expression in most cancer types (P < 0.05). In contrast, common lymphoid progenitor (CLP) cells, osteoblasts, epithelial cells, and smooth muscle cells were negatively associated with AL133467.1 expression in most cancer types (P < 0.05). In addition, scores pertaining to the immune response, stromal components, and overall tumor microenvironment were positively correlated with AL133467.1 expression. These results underscore the strong association between AL133467.1 and the tumor microenvironment across an array of cancer types.
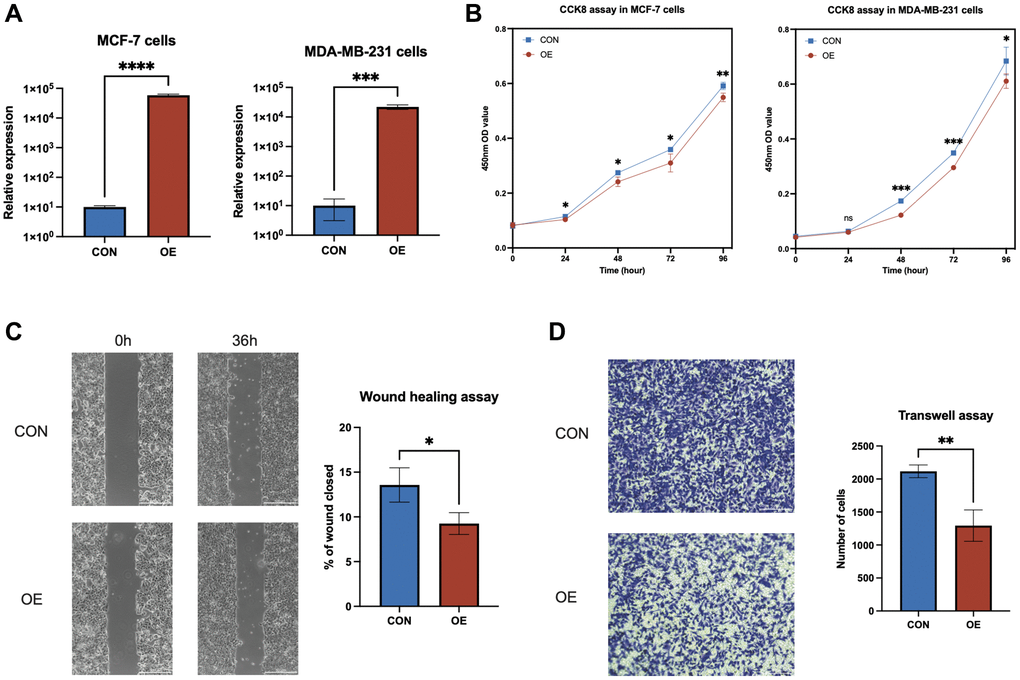
Elevated expression of AL133467.1 notably reduced the proliferation and migration of breast cancer cells
To decipher the molecular roles of anoikis-related genes (ARGs) in breast cancer, a comprehensive study on AL133467.1, based on findings from qRT-PCR and survival analyses, aimed to clarify its distinct function in IBC. Transient transfection was carried out for AL133467.1, to attain its overexpression in both MCF-7 and MDA-MB-231 cell lines (Figure 7A). CCK-8 assay confirmed that enhanced expression of AL133467.1 greatly reduced the proliferation of both MCF-7 and MDA-MB-231 cells (Figure 7B). Moreover, a wound-healing assay showed that enhanced expression of AL133467.1 significantly reduced the migration potential of MCF-7 cells (Figure 7C). Lastly, Transwell assay demonstrated that enhanced expression of AL133467.1 significantly reduced the migration of MDA-MB-231 cells (Figure 7D).
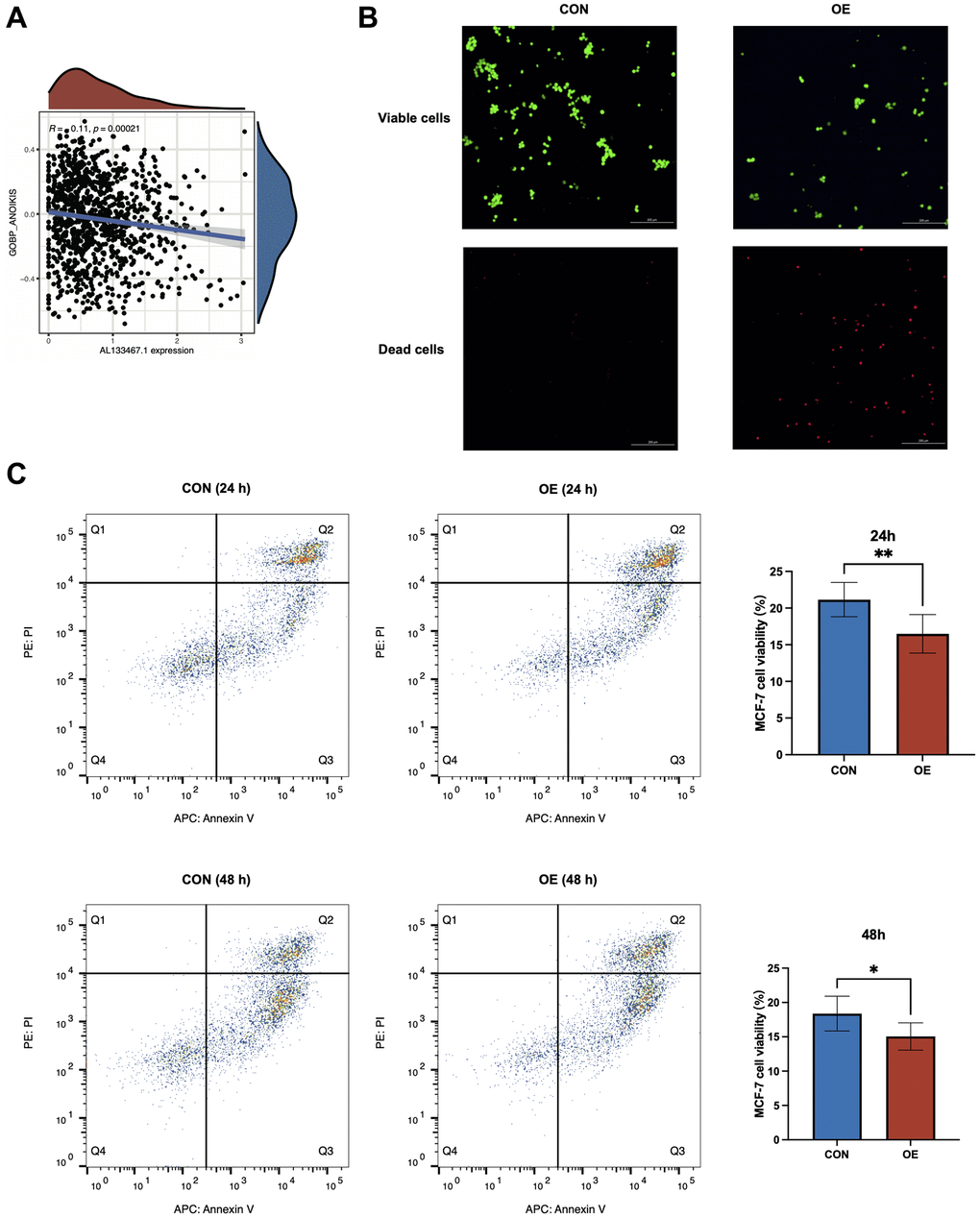
Over-expression of AL133467.1 impaired anoikis in breast cancer cells
To elucidate the effect of AL133467.1 expression on anoikis in breast cancer, we studied the association between AL133467.1 expression and anoikis. As depicted in Figure 8A, AL133467.1 expression was negatively correlated with anoikis, suggesting that AL133467.1 can potentially function as a negative regulator of anoikis. To confirm these findings, we induced anoikis in MCF-7 cells by preventing cell adhesion to the culture dish. As depicted in Figure 8B, following a 24-hour incubation, the cells overexpressing AL133467.1 exhibited lower viability than the control cells. To quantitatively assess differences between the two groups, flow cytometry was used to measure cell viability. After a 24-hour or 48-hour incubation in an anti-adhesion environment, the overexpression group displayed significantly reduced cell viability relative to the control group (Figure 8C). These findings suggest that overexpression of AL133467.1 may enhance anoikis in MCF-7 cells.
Discussion
Anoikis is a unique variant of apoptosis that has significant physiological relevance in organisms as it circumvents unwarranted cell adherence and subsequent proliferation [14]. Recent research has highlighted the pivotal role of anoikis in breast cancer ontogeny and metastasis. Many studies have emphasized that the counter-anoikis signal is a critical phase for breast cancer cells to obtain the capability of invasion and metastasis [15]. Breast cancer cells survive within the circulatory system by impeding anoikis, thereby increasing the likelihood of establishing colonies in distal tissues. Consequently, exploring the mechanisms inhibiting anoikis and methods to obstruct these processes has become a key area for future breast cancer research [4]. Moreover, anoikis could potentially serve as a novel target for drug discovery. For instance, medications designed to counteract anoikis resistance mechanisms may bolster treatment efficacies in patients with breast cancer [16]. To the best of our knowledge, previous studies have not systematically evaluated the prognostic value of anoikis-associated lncRNAs in breast cancer. Thus, this study can be used as the main reference for subsequent research.
LncRNAs are important for the progression of breast cancer. Specific lncRNAs have been reported to either promote or hinder breast cancer progression through key processes that influence tumor proliferation, invasion, and metastasis [17]. For instance, the lncRNA HOTAIR, which is overexpressed in breast cancer, can change gene expression through alterations in chromosome structure [18]. In contrast, expression of the lncRNA GAS5 is diminished in many forms of breast cancer, and its loss is associated with the incidence and progression of breast cancer [19]. These findings indicate that lncRNAs may exert a significant effect on the pathogenesis of breast cancer, thus providing a potential novel approach for breast cancer treatment.
We established and validated a signature of seven ARLs to predict the prognosis of breast cancer patients. The OS of patients in the low and high ARL score groups was distinctly stratified using ARL signatures. Five ARLs (MAPT.IT1, AL133467.1, AC004585.1, AC055854.1, and LINC02613) were identified as protective factors, whereas two (LINC01614 and C6orf99) were identified as risk factors for breast cancer. In vitro functional experiments indicated a significant role of AL133467.1 in the proliferation and migration of breast cancer cells, as well as a certain degree of impact on the resistance of breast cancer cells to anoikis. Previous studies have not reported the role of AL133467.1 in breast cancer; thus, exploring the relationship between AL133467.1 and anoikis could potentially aid in understanding AL133467.1’s role in breast cancer.
In addition, we investigated the correlation between ARL scores and clinicopathological features. Among the parameters of age, tumor stage (T), node stage (N), metastatic stage (M), pathological stage, and ARL score, only age and the ARL score were identified as independent prognostic factors for IBC in the complete training and test sets. This underscores the ability of the ARL score to independently predict the prognosis of IBC. To enhance its clinical applicability, we integrated the ARL score with other clinical prognostic factors to construct a prognostic nomogram. The ROC curve, calibration curve, and DCA confirmed that the nomogram exhibited considerable advantages in predicting survival and guiding clinical decision-making compared with traditional clinicopathological features.
In this study, the results of GSVA enrichment analysis revealed that patients with elevated ARL scores demonstrated significant activation of cell cycle, energy metabolism, and biosynthesis-related pathways, including the proteasome, cell cycle, DNA replication, homologous recombination, and mismatch repair. Increased activation of these pathways may facilitate uncontrolled cellular proliferation and tumorigenesis [20–23]. Concurrently, the amplified expression of energy metabolism pathways such as glycolysis, the citrate cycle (TCA cycle), oxidative phosphorylation, and the pentose phosphate pathway may satisfy the energy requirements of tumor cells [24, 25]. Furthermore, an increase in the expression of amino sugars and nucleotide sugar metabolism, folate biosynthesis, terpenoid backbone biosynthesis, and steroid biosynthesis can promote the growth of tumor cells [26–28]. In contrast, patients with lower ARL scores exhibit increased expression of immune response-related pathways such as primary immunity, hematopoietic cell lineage, and an intentional immune network for IgA production, reflecting the potential protective role of a robust immune system in averting tumorigenesis [29, 30]. In addition, upregulation of circadian rhythms in mammals may underscore the tumor-suppressive role of a well-functioning biological clock [31]. Although these preliminary analyses contributed to a comprehensive understanding of the prognostic implications of ARL scores in patients with breast cancer, there is a compelling need for further research to corroborate and expand these observations.
Gene mutations, particularly in pivotal tumor suppressor genes and oncogenes, can induce unregulated cell growth and division, thereby promoting tumor formation. These alterations can modulate sensitivity to specific treatments and provide critical information for clinical decision-making and prognostic prediction [32]. In our analysis, several key genes, such as TP53 and PIK3CA, exhibited diverse mutation frequencies in breast cancer patients with high and low ARL scores. Intriguingly, we discovered a significantly higher mutation frequency in TP53 among patients with poor prognoses (47%) than among those with good prognoses (20%). Given the crucial role of TP53 as a tumor suppressor, its mutation may precipitate unregulated cell growth and division, thereby increasing tumor risk [33]. Hence, the elevated TP53 mutation rate may suggest enhanced tumorigenic potential in breast cancer patients with a high ARL score. Concurrently, we observed a higher mutation frequency in PIK3CA in patients with a low ARL score (44%) than in those with a high ARL score (26%). The protein encoded by PIK3CA plays a pivotal role in various biological processes including cell growth, survival, and metabolism [34]. While the reason for the higher PIK3CA mutation frequency in patients with a better prognosis remains elusive, it may suggest novel therapeutic avenues, such as PIK3CA-specific mutation inhibitors. Additionally, we detected high mutation frequencies in TTN and GATA3 across both the high and low ARL score groups, potentially reflecting common biological characteristics of breast cancer and underscoring their importance in breast cancer pathogenesis [35, 36]. Although our research has provided interesting insights, various factors warrant further exploration, including the type and location of mutations and the impact of other genetic and environmental factors on breast cancer prognosis.
To delve deeper into the function of the prognostic model based on the ARL score in IBC, we explored the correlation between chemotherapy and ARL scores. Lapatinib, vinorelbine, gemcitabine, 5-fluorouracil, epirubicin, and sorafenib are used for breast cancer chemotherapy. Our results demonstrated a positive association between the IC50 values of vinorelbine, gemcitabine, 5-fluorouracil, epirubicin, and sorafenib and the ARL score, whereas a negative relationship was found between the IC50 value of lapatinib, indicating a strong link between the ARL score and chemotherapy. This discovery further highlights the potential of the prognostic model and may offer clinical guidance for the selection of chemotherapeutic drugs for breast cancer patients with different ARL scores.
Immune cell dynamics within the tumor microenvironment play a pivotal role in determining breast cancer prognoses [37]. Our results revealed that patients with low ARL scores exhibited significantly increased infiltration of activated CD8 T cells, effector memory CD8 T cells, effector memory CD4 T cells, and type 1 T helper cells. These cells are integral to tumor inhibition, especially in facilitating cytotoxic reactions to eliminate tumor cells [38]. Additionally, we observed substantial infiltration of natural killer cells into the tumor microenvironment of patients with low ARL scores. As natural antitumor immune cells, natural killer cells play a critical role in directly eliminating tumor cells and orchestrating adaptive immune responses [39]. In contrast, patients with high ARL scores had elevated levels of activated CD4 T cells, gamma-delta T cells, and type 17 T helper cells. This may reflect an immunosuppressive phenomenon within the tumor microenvironment, and these cells can contribute to the regulation of inflammation and the survival of tumor cells during tumorigenesis [40–42]. Our findings underscore the importance of in-depth examination of the roles of these immune cell subsets in breast cancer prognosis, which has profound implications for devising more effective antitumor immunotherapies.
Despite these insightful findings, this study has several limitations. First, prospective multicenter studies with extensive BRCA cohorts are warranted to corroborate the reliability of the ARL signature and the accompanying results. Secondly, although we explored the functionality of one ARL in vitro, additional experimental investigations are necessary to elucidate its regulatory mechanisms and functions.
Materials and Methods
Data collection
The main information utilized for this research was obtained from TCGA repository, comprising 1,109 IBC samples and 113 healthy breast tissue samples. Patients with unknown clinical information and an overall survival time of less than 30 days were excluded from the study. Gene sets associated with anoikis were obtained from a prior investigation [43]. Figure 9 shows the flowchart of our study.
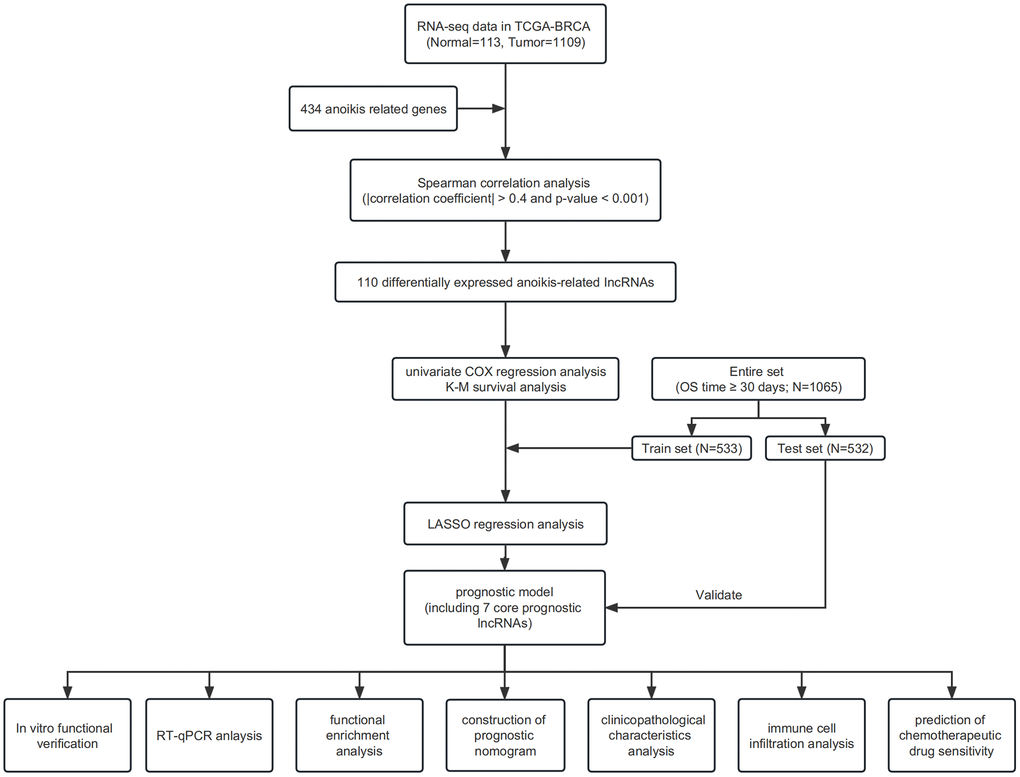
Prognostic model construction and verification
The RNA data of anoikis-related DELs and clinical information on IBC from TCGA were combined based on the sample ID. Only patients with an OS time of 30 days or more were included in further survival analyses. Univariate Cox regression analysis was performed using the Kaplan–Meier “survival” package [47]. Subsequently, for further identification of the core genes related to anoikis and the construction of an ARL score-based prognostic model, we employed the “glmnet” and “survival” packages to conduct LASSO logistic regression analysis [48]. The following formula was used to calculate the ARL score of IBC patients: C6orf99 expression value × corresponding coef + LINC01614 expression value × corresponding coef + LINC02613 expression value × corresponding coef + AC055854.1 expression value × corresponding coef + AL133467.1 expression value × corresponding coef + AC004585.1 expression value × corresponding coef + MAPT.IT1 expression value × corresponding coef. The Shapiro–Wilk normality test and Wilcoxon test were used to perform a difference analysis between the two groups. The Spearman method was used to perform correlation analysis. Kaplan–Meier survival analysis was performed using the “survminer” and “survival” packages [47]. PCA was conducted using the “factoextra” and “FactoMineR” packages [49]. ROC analysis was performed using the “timeROC” package [47].
Correlation between clinical characteristics and the prognostic model
We used the Wilcoxon test to evaluate the differences in ARL scores among patients with different clinicopathological features. Correlation analysis was performed using the Spearman method. The “maftools” package was utilized to explore the somatic variants between individuals with low and high ARL scores [50].
Construction of the nomogram for predicting the prognosis of IBC
Univariate Cox analysis was carried out using the “survival” package [47]. The calibration analysis and nomogram construction were carried out using the “rms” and “survival” packages [47]. The “timeROC” package was employed for ROC analysis [47]. The decision curve analysis (DCA) was carried out using the “survival” and “survminer” packages [51].
Function enrichment analysis
We conducted Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses using the “clusterProfiler” package [52]. Gene set enrichment analysis (GSEA) was performed using the GSEA software available on the GSEA website (http://software.broadinstitute.org/gsea/index.jsp) and the “h.all.v2022.1. Hs.symbols.gmt” gene sets were sourced from the Molecular Signatures Database. Furthermore, Gene Set Variation Analysis (GSVA) was carried out using the “GSEABase” and “GSVA” packages, along with the “c2.cp.kegg.v2022.1. Hs.symbols.gmt” gene set [53, 54]. A false discovery rate (FDR) of less than 0.25 and a P-value cutoff of less than 0.05 were used to evaluate statistical significance.
Immune infiltration characteristics of the TME
The analysis of immune cell infiltration and immune-related pathway enrichment was conducted using the “GSVA” and “GSEABase” packages [53, 54]. Marker genes of immune cells identified by Bindea et al. [54] were used in this analysis. Moreover, we employed the “estimate” package to calculate immune and stromal scores in both the low ARL score group and the high ARL score group. [55]. The Shapiro–Wilk normality test and Wilcoxon test were used to examine the differences between groups.
Immunotherapy response and chemotherapy sensitivity
We retrieved immunogenomic examination data from TCIA website (https://tcia.at/home) [56]. We used TIDE (http://tide.dfci.harvard.edu/) to evaluate the predictive effect of anoikis-related lncRNAs signatures on immunotherapy response. We used the Shapiro–Wilk normality test and Wilcoxon test to assess the disparity between the two clusters. The evaluation of drug susceptibility was executed by applying the “oncopredict” package [57]. We employed the Spearman approach for correlation analysis.
qRT-PCR
The study utilized tissue samples from the biobank of Shanghai First Maternity and Infant Hospital and was conducted with the exemption of informed consent by the Ethics Committee of the same hospital. Total RNA was obtained from 15 breast cancer samples and 12 nearby normal breast tissues using Total RNA Extraction Reagent (TRIzol®) (RK30129, ABclonal, USA), according to the guidelines provided by the manufacturer. With the assistance of ABScript III RT Master Mix (RK20428, ABclonal), we performed the reverse transcription using 1 μg of total RNA. qRT-PCR analysis was conducted utilizing Genlous 2× SYBR™ Green PCR Fast qPCR Mix (Low ROX Premixed) (RK21206, ABclonal). The evaluation of the comparative expression of the genes of concern was carried out utilizing the 2−ΔΔCT methodology, with beta-actin serving as the reference gene. The primer sequences used are listed in Supplementary Table 1.
Pan-cancer analysis
To conduct a pan-cancer differential analysis of AL133467.1, we obtained a standardized and consolidated dataset called TCGA Pan-Cancer (Pancan, N = 10535, G = 60499) from the UCSC database (https://xenabrowser.net/). From this dataset, we specifically extracted the expression data of AL133467.1 for every individual sample. Samples were selected specifically from healthy solid tissues and cancerous peripheral blood derived from primary tumors. Samples with no expression levels were excluded, and all expression values were converted using the log2(x+1) transformation. The expression data for 26 cancer types were obtained after excluding those with fewer than three samples. The Wilcoxon test was used to determine the differences between two groups.
For the survival analysis, in addition to obtaining survival data from the UCSC, we procured a high-quality prognostic dataset from TCGA using an integrated TCGA pan-cancer clinical data resource [47]. Samples with zero expression levels were excluded, as were samples with a follow-up time of less than 30 days. All expression values were transformed by log2(x+1). Cancers with fewer than 10 samples were excluded, and 39 cancer types were included (Supplementary Figure 6B). We employed the coxph function of the R software survival package to perform the Cox regression analysis [58]. The significance of the prognosis was determined through a statistical test with the Logrank test.
To analyze immune checkpoint markers, we obtained an identical comprehensive pan-cancer dataset from the UCSC database. From each sample, we extracted the expression of 60 genes related to immune checkpoint pathways, consisting of 24 inhibitory and 36 stimulatory genes [59]. The association between AL133467.1 and immune checkpoint marker genes was investigated by Pearson correlation analysis.
For immune infiltration analysis, we extracted the gene expression profiles of each cancer type from the UCSC pan-cancer dataset, mapped the expression profiles to the gene symbol, and employed the R software package “IOBR” [60] and deconvo_xCell method [61] to estimate the infiltration scores of 67 tumor microenvironment cell types in 39 cancer types, based on gene expression data. The “psych” package was used to examine the relationship between AL133467.1 and levels of immune cell infiltration in various tumor types.
Cell culture and plasmid transfection
The human breast cancer cell lines MCF-7 and MDA-MB-231 were sourced from Wuhan Procell Life Science and Technology Co., Ltd., (Wuhan, China). We propagated MCF-7 cells in minimum Eagle’s medium (MEM), supplemented with 10% fetal bovine serum (Gibco, USA), 0.01 mg/mL insulin, 100 U/mL penicillin G, and 100 μg/mL streptomycin. MDA-MB-231 cells were grown in Dulbecco’s modified Eagle’s medium (DMEM), supplemented with 10% fetal bovine serum (Gibco, USA), 100 U/mL penicillin G, and 100 μg/mL streptomycin. Both cell lines were incubated in a 5% CO2 atmosphere at 37°C. To assess the effect of AL133467.1, Lipofectamine™ 3000 (Invitrogen, USA) was used to transfect pcDNA-AL133467.1/pcDNA (negative control) into MCF-7 and MDA-MB-231 cells according to the Lipofectamine™ 3000 reagent protocol. Functional assays were performed, and RNA was collected 48 h after transfection.
Cell phenotype assays
The proliferative behavior of breast cancer cells was investigated by CCK-8 assay. Following digestion, cells from different treatment sets were seeded into 96-well plates (Corning, USA) at a cell density of 2,000 cells/well. The CCK-8 assay was performed as per the manufacturer’s guidelines, and the absorbance was measured at 450 nm using a microplate reader.
To examine the migratory capacity of breast cancer cells, wound-healing and Transwell assays were performed. For the wound-healing assay, digested cells from various treatment groups were placed in a Culture-Insert 2 Well (Ibidi, Germany) in 24-well plates (Corning, USA). Following a 24-hour incubation period, the Culture-Insert 2 Well was meticulously removed, generating a cell-free zone of roughly 500 μm. An inverted microscope (Nikon Ts2R, Japan) at 40× magnification was used to capture images of the gap area at 0 and 36 h.
For the Transwell assay, 800 μL of medium supplemented with 10% fetal bovine serum (Gibco) was deposited in the lower chamber of a Transwell™ insert, and the upper chamber was filled with 200 μL of serum-free medium containing 50,000 cells. After a 48-hour incubation phase, cells that had crossed the membrane were fixed using 4% paraformaldehyde, rinsed with phosphate-buffered saline, stained with 0.3% crystal violet, and imaged with an inverted microscope at 100× magnification.
Anoikis assay
Cells from different treatment groups were digested and seeded in anchorage-resistant 96-well plates of an Anoikis Assay Kit (Abcam, UK) at a density of 2,000 cells per well to simulate the state of anoikis. After 24 h of incubation, the culture medium was supplemented with calcein AM and ethidium homodimer from the Anoikis Assay Kit (Abcam, UK), followed by an additional incubation period of 30–60 min. After observing the cells under a fluorescence microscope, fluorometric signals were detected. For flow cytometric analysis, cells from the various treatment groups were digested and seeded in ultra-low-attachment 6-well plates (Corning, USA) at a density of 100,000 cells/well. Following incubation for 24 or 48 h, the cells from the respective treatment groups were collected and subjected to flow cytometry to detect apoptosis. This analysis was performed using an Annexin V-Alexa Fluor 647/PI Apoptosis Detection Kit (Yeasen, China) following the manufacturer’s instructions.
Statistical analysis
The process of data analysis and visual representation was executed using R software (versions 3.6.3 and 4.2.1) and GraphPad Prism software (version 9.0). Unless otherwise indicated, the Wilcoxon test was utilized to discern differences between two groups, the Spearman test was employed to probe correlations between two groups, and Kaplan–Meier survival analysis was applied to evaluate survival differences between two sets. Statistical significance was set at p < 0.05.
Conclusion
Our study constructed and validated a suite of lncRNA models based on ARL scoring, thereby offering a powerful prognostic tool for breast cancer. Furthermore, a comprehensive nomogram was created employing both clinicopathological parameters and the ARL score to predict overall survival in a clinical setting. We demonstrated a significant inverse relationship between TP53 and PIK3CA mutation frequencies relative to the ARL score. Intriguingly, pathways associated with the cell cycle, energy metabolism, and biosynthesis demonstrated increased activation in the high ARL score group. We found a robust correlation between the ARL score and the extent of immune cell infiltration, TMB, expression of immune checkpoints, and predictive IC50 values of chemotherapy agents. Moreover, lncRNA AL133467.1 affected both breast cancer cell proliferation and migration, as well as possibly resistance to anoikis. In conclusion, the ARL signature, as a holistic prognostic tool, facilitates the assessment of risk stratification and informs the clinical decision-making processes for patients with breast cancer. Further investigation is required to elucidate the broader clinical implications of these findings.
Author Contributions
WD: Conceptualization, Methodology, Writing-Original draft preparation. JL: Formal analysis, Data curation, Writing-Reviewing and Editing. ZZ: Investigation, Validation, Supervision. All authors read and approved the final manuscript.
Acknowledgments
The authors would like to express their sincere gratitude to Dr. Tao Wang for his valuable guidance on the in vitro experiment component of this study.
Conflicts of Interest
The authors declare no conflicts of interest related to this study.
Ethical Statement and Consent
The tissue samples employed in this research were obtained from the Biobank of Shanghai First Maternity and Infant Hospital. The study received approval from the Ethics Committee of Shanghai First Maternity and Infant Hospital (KS22361) and an exemption from obtaining informed consent was granted.
Funding
This work was supported by the Natural Science Foundation of Shanghai, China (Grant No. 21ZR1451000).
References
- 1. Loibl S, Poortmans P, Morrow M, Denkert C, Curigliano G. Breast cancer. Lancet. 2021; 397:1750–69. https://doi.org/10.1016/S0140-6736(20)32381-3 [PubMed]
- 2. Wilkinson L, Gathani T. Understanding breast cancer as a global health concern. Br J Radiol. 2022; 95:20211033. https://doi.org/10.1259/bjr.20211033 [PubMed]
- 3. Waks AG, Winer EP. Breast Cancer Treatment. JAMA. 2019; 321:316. https://doi.org/10.1001/jama.2018.20751 [PubMed]
- 4. Paoli P, Giannoni E, Chiarugi P. Anoikis molecular pathways and its role in cancer progression. Biochim Biophys Acta. 2013; 1833:3481–98. https://doi.org/10.1016/j.bbamcr.2013.06.026 [PubMed]
- 5. Raeisi M, Zehtabi M, Velaei K, Fayyazpour P, Aghaei N, Mehdizadeh A. Anoikis in cancer: The role of lipid signaling. Cell Biol Int. 2022; 46:1717–28. https://doi.org/10.1002/cbin.11896 [PubMed]
- 6. Adeshakin FO, Adeshakin AO, Afolabi LO, Yan D, Zhang G, Wan X. Mechanisms for Modulating Anoikis Resistance in Cancer and the Relevance of Metabolic Reprogramming. Front Oncol. 2021; 11:626577. https://doi.org/10.3389/fonc.2021.626577 [PubMed]
- 7. Ye G, Yang Q, Lei X, Zhu X, Li F, He J, Chen H, Ling R, Zhang H, Lin T, Liang Z, Liang Y, Huang H, et al. Nuclear MYH9-induced CTNNB1 transcription, targeted by staurosporin, promotes gastric cancer cell anoikis resistance and metastasis. Theranostics. 2020; 10:7545–60. https://doi.org/10.7150/thno.46001 [PubMed]
- 8. Tian T, Lu Y, Lin J, Chen M, Qiu H, Zhu W, Sun H, Huang J, Yang H, Deng W. CPT1A promotes anoikis resistance in esophageal squamous cell carcinoma via redox homeostasis. Redox Biol. 2022; 58:102544. https://doi.org/10.1016/j.redox.2022.102544 [PubMed]
- 9. Reddig PJ, Juliano RL. Clinging to life: cell to matrix adhesion and cell survival. Cancer Metastasis Rev. 2005; 24:425–39. https://doi.org/10.1007/s10555-005-5134-3 [PubMed]
- 10. Statello L, Guo CJ, Chen LL, Huarte M. Author Correction: Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021; 22:159. https://doi.org/10.1038/s41580-021-00330-4 [PubMed]
- 11. Grillone K, Riillo C, Scionti F, Rocca R, Tradigo G, Guzzi PH, Alcaro S, Di Martino MT, Tagliaferri P, Tassone P. Non-coding RNAs in cancer: platforms and strategies for investigating the genomic "dark matter". J Exp Clin Cancer Res. 2020; 39:117. https://doi.org/10.1186/s13046-020-01622-x [PubMed]
- 12. Gao N, Li Y, Li J, Gao Z, Yang Z, Li Y, Liu H, Fan T. Long Non-Coding RNAs: The Regulatory Mechanisms, Research Strategies, and Future Directions in Cancers. Front Oncol. 2020; 10:598817. https://doi.org/10.3389/fonc.2020.598817 [PubMed]
- 13. Zhang YY, Li XW, Li XD, Zhou TT, Chen C, Liu JW, Wang L, Jiang X, Wang L, Liu M, Zhao YG, Li SD. Comprehensive analysis of anoikis-related long non-coding RNA immune infiltration in patients with bladder cancer and immunotherapy. Front Immunol. 2022; 13:1055304. https://doi.org/10.3389/fimmu.2022.1055304 [PubMed]
- 14. Frisch SM, Screaton RA. Anoikis mechanisms. Curr Opin Cell Biol. 2001; 13:555–62. https://doi.org/10.1016/s0955-0674(00)00251-9 [PubMed]
- 15. Simpson CD, Anyiwe K, Schimmer AD. Anoikis resistance and tumor metastasis. Cancer Lett. 2008; 272:177–85. https://doi.org/10.1016/j.canlet.2008.05.029 [PubMed]
- 16. Douma S, Van Laar T, Zevenhoven J, Meuwissen R, Van Garderen E, Peeper DS. Suppression of anoikis and induction of metastasis by the neurotrophic receptor TrkB. Nature. 2004; 430:1034–9. https://doi.org/10.1038/nature02765 [PubMed]
- 17. Schmitt AM, Chang HY. Long Noncoding RNAs in Cancer Pathways. Cancer Cell. 2016; 29:452–63. https://doi.org/10.1016/j.ccell.2016.03.010 [PubMed]
- 18. Gupta RA, Shah N, Wang KC, Kim J, Horlings HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, Wang Y, Brzoska P, Kong B, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010; 464:1071–6. https://doi.org/10.1038/nature08975 [PubMed]
- 19. Mourtada-Maarabouni M, Pickard MR, Hedge VL, Farzaneh F, Williams GT. GAS5, a non-protein-coding RNA, controls apoptosis and is downregulated in breast cancer. Oncogene. 2009; 28:195–208. https://doi.org/10.1038/onc.2008.373 [PubMed]
- 20. Goldberg AL. Protein degradation and protection against misfolded or damaged proteins. Nature. 2003; 426:895–9. https://doi.org/10.1038/nature02263 [PubMed]
- 21. Hartwell LH, Weinert TA. Checkpoints: controls that ensure the order of cell cycle events. Science. 1989; 246:629–34. https://doi.org/10.1126/science.2683079 [PubMed]
- 22. Li GM. Mechanisms and functions of DNA mismatch repair. Cell Res. 2008; 18:85–98. https://doi.org/10.1038/cr.2007.115 [PubMed]
- 23. Moynahan ME, Jasin M. Mitotic homologous recombination maintains genomic stability and suppresses tumorigenesis. Nat Rev Mol Cell Biol. 2010; 11:196–207. https://doi.org/10.1038/nrm2851 [PubMed]
- 24. Berg JM, Tymoczko JL, Stryer L, Gatto GJ
Jr . Biochemistry. New York: W.H. Freeman. 2012. - 25. Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009; 324:1029–33. https://doi.org/10.1126/science.1160809 [PubMed]
- 26. Locasale JW. Serine, glycine and one-carbon units: cancer metabolism in full circle. Nat Rev Cancer. 2013; 13:572–83. https://doi.org/10.1038/nrc3557 [PubMed]
- 27. Tholl D. Biosynthesis and biological functions of terpenoids in plants. Adv Biochem Eng Biotechnol. 2015; 148:63–106. https://doi.org/10.1007/10_2014_295 [PubMed]
- 28. Cox MM, Nelson DL. Lehninger Principles of Biochemistry. Wh Freeman. 2000; 5.
- 29. MacLennan I, Vinuesa C. Dendritic cells, BAFF, and APRIL: innate players in adaptive antibody responses. Immunity. 2002; 17:235–8. https://doi.org/10.1016/s1074-7613(02)00398-9 [PubMed]
- 30. Murphy K, Travers P, Walport M, Janeway CA
Jr . Janeway's immunobiology. New York: Garland Science. 2012. - 31. Sahar S, Sassone-Corsi P. Metabolism and cancer: the circadian clock connection. Nat Rev Cancer. 2009; 9:886–96. https://doi.org/10.1038/nrc2747 [PubMed]
- 32. Vogelstein B, Papadopoulos N, Velculescu VE, Zhou S, Diaz LA
Jr , Kinzler KW. Cancer genome landscapes. Science. 2013; 339:1546–58. https://doi.org/10.1126/science.1235122 [PubMed] - 33. Muller PA, Vousden KH. Mutant p53 in cancer: new functions and therapeutic opportunities. Cancer Cell. 2014; 25:304–17. https://doi.org/10.1016/j.ccr.2014.01.021 [PubMed]
- 34. Thorpe LM, Yuzugullu H, Zhao JJ. PI3K in cancer: divergent roles of isoforms, modes of activation and therapeutic targeting. Nat Rev Cancer. 2015; 15:7–24. https://doi.org/10.1038/nrc3860 [PubMed]
- 35. Schafer S, de Marvao A, Adami E, Fiedler LR, Ng B, Khin E, Rackham OJ, van Heesch S, Pua CJ, Kui M, Walsh R, Tayal U, Prasad SK, et al. Titin-truncating variants affect heart function in disease cohorts and the general population. Nat Genet. 2017; 49:46–53. https://doi.org/10.1038/ng.3719 [PubMed]
- 36. Asselin-Labat ML, Sutherland KD, Barker H, Thomas R, Shackleton M, Forrest NC, Hartley L, Robb L, Grosveld FG, van der Wees J, Lindeman GJ, Visvader JE. Gata-3 is an essential regulator of mammary-gland morphogenesis and luminal-cell differentiation. Nat Cell Biol. 2007; 9:201–9. https://doi.org/10.1038/ncb1530 [PubMed]
- 37. Mittal S, Brown NJ, Holen I. The breast tumor microenvironment: role in cancer development, progression and response to therapy. Expert Rev Mol Diagn. 2018; 18:227–43. https://doi.org/10.1080/14737159.2018.1439382 [PubMed]
- 38. Schreiber RD, Old LJ, Smyth MJ. Cancer immunoediting: integrating immunity's roles in cancer suppression and promotion. Science. 2011; 331:1565–70. https://doi.org/10.1126/science.1203486 [PubMed]
- 39. Vivier E, Raulet DH, Moretta A, Caligiuri MA, Zitvogel L, Lanier LL, Yokoyama WM, Ugolini S. Innate or adaptive immunity? The example of natural killer cells. Science. 2011; 331:44–9. https://doi.org/10.1126/science.1198687 [PubMed]
- 40. Bailey SR, Nelson MH, Himes RA, Li Z, Mehrotra S, Paulos CM. Th17 cells in cancer: the ultimate identity crisis. Front Immunol. 2014; 5:276. https://doi.org/10.3389/fimmu.2014.00276 [PubMed]
- 41. Palucka AK, Coussens LM. The Basis of Oncoimmunology. Cell. 2016; 164:1233–47. https://doi.org/10.1016/j.cell.2016.01.049 [PubMed]
- 42. Chien YH, Meyer C, Bonneville M. γδ T cells: first line of defense and beyond. Annu Rev Immunol. 2014; 32:121–55. https://doi.org/10.1146/annurev-immunol-032713-120216 [PubMed]
- 43. Chen S, Gu J, Zhang Q, Hu Y, Ge Y. Development of Biomarker Signatures Associated with Anoikis to Predict Prognosis in Endometrial Carcinoma Patients. J Oncol. 2021; 2021:3375297. https://doi.org/10.1155/2021/3375297 [PubMed]
- 44. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014; 15:550. https://doi.org/10.1186/s13059-014-0550-8 [PubMed]
- 45. Xu Z, Peng B, Liang Q, Chen X, Cai Y, Zeng S, Gao K, Wang X, Yi Q, Gong Z, Yan Y. Construction of a Ferroptosis-Related Nine-lncRNA Signature for Predicting Prognosis and Immune Response in Hepatocellular Carcinoma. Front Immunol. 2021; 12:719175. https://doi.org/10.3389/fimmu.2021.719175 [PubMed]
- 46. Lu H, Wu J, Liang L, Wang X, Cai H. Identifying a Novel Defined Pyroptosis-Associated Long Noncoding RNA Signature Contributes to Predicting Prognosis and Tumor Microenvironment of Bladder Cancer. Front Immunol. 2022; 13:803355. https://doi.org/10.3389/fimmu.2022.803355 [PubMed]
- 47. Liu J, Lichtenberg T, Hoadley KA, Poisson LM, Lazar AJ, Cherniack AD, Kovatich AJ, Benz CC, Levine DA, Lee AV, Omberg L, Wolf DM, Shriver CD, et al, and Cancer Genome Atlas Research Network. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell. 2018; 173:400–16.e11. https://doi.org/10.1016/j.cell.2018.02.052 [PubMed]
- 48. Engebretsen S, Bohlin J. Statistical predictions with glmnet. Clin Epigenetics. 2019; 11:123. https://doi.org/10.1186/s13148-019-0730-1 [PubMed]
- 49. Ringnér M. What is principal component analysis? Nat Biotechnol. 2008; 26:303–4. https://doi.org/10.1038/nbt0308-303 [PubMed]
- 50. Mayakonda A, Lin DC, Assenov Y, Plass C, Koeffler HP. Maftools: efficient and comprehensive analysis of somatic variants in cancer. Genome Res. 2018; 28:1747–56. https://doi.org/10.1101/gr.239244.118 [PubMed]
- 51. Van Calster B, Wynants L, Verbeek JFM, Verbakel JY, Christodoulou E, Vickers AJ, Roobol MJ, Steyerberg EW. Reporting and Interpreting Decision Curve Analysis: A Guide for Investigators. Eur Urol. 2018; 74:796–804. https://doi.org/10.1016/j.eururo.2018.08.038 [PubMed]
- 52. Yu G, Wang LG, Han Y, He QY. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS. 2012; 16:284–7. https://doi.org/10.1089/omi.2011.0118 [PubMed]
- 53. Hänzelmann S, Castelo R, Guinney J. GSVA: gene set variation analysis for microarray and RNA-seq data. BMC Bioinformatics. 2013; 14:7. https://doi.org/10.1186/1471-2105-14-7 [PubMed]
- 54. Bindea G, Mlecnik B, Tosolini M, Kirilovsky A, Waldner M, Obenauf AC, Angell H, Fredriksen T, Lafontaine L, Berger A, Bruneval P, Fridman WH, Becker C, et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity. 2013; 39:782–95. https://doi.org/10.1016/j.immuni.2013.10.003 [PubMed]
- 55. Yoshihara K, Shahmoradgoli M, Martínez E, Vegesna R, Kim H, Torres-Garcia W, Treviño V, Shen H, Laird PW, Levine DA, Carter SL, Getz G, Stemke-Hale K, et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat Commun. 2013; 4:2612. https://doi.org/10.1038/ncomms3612 [PubMed]
- 56. Charoentong P, Finotello F, Angelova M, Mayer C, Efremova M, Rieder D, Hackl H, Trajanoski Z. Pan-cancer Immunogenomic Analyses Reveal Genotype-Immunophenotype Relationships and Predictors of Response to Checkpoint Blockade. Cell Rep. 2017; 18:248–62. https://doi.org/10.1016/j.celrep.2016.12.019 [PubMed]
- 57. Maeser D, Gruener RF, Huang RS. oncoPredict: an R package for predicting in vivo or cancer patient drug response and biomarkers from cell line screening data. Brief Bioinform. 2021; 22:bbab260. https://doi.org/10.1093/bib/bbab260 [PubMed]
- 58. Andersen PK, Gill RD. Cox's Regression Model for Counting Processes: A Large Sample Study. Ann Stat. 1982; 10:1100–20. https://www.jstor.org/stable/2240714.
- 59. Thorsson V, Gibbs DL, Brown SD, Wolf D, Bortone DS, Ou Yang TH, Porta-Pardo E, Gao GF, Plaisier CL, Eddy JA, Ziv E, Culhane AC, Paull EO, et al, and Cancer Genome Atlas Research Network. The Immune Landscape of Cancer. Immunity. 2018; 48:812–30.e14. https://doi.org/10.1016/j.immuni.2018.03.023 [PubMed]
- 60. Zeng D, Ye Z, Shen R, Yu G, Wu J, Xiong Y, Zhou R, Qiu W, Huang N, Sun L, Li X, Bin J, Liao Y, et al. IOBR: Multi-Omics Immuno-Oncology Biological Research to Decode Tumor Microenvironment and Signatures. Front Immunol. 2021; 12:687975. https://doi.org/10.3389/fimmu.2021.687975 [PubMed]
- 61. Aran D, Hu Z, Butte AJ. xCell: digitally portraying the tissue cellular heterogeneity landscape. Genome Biol. 2017; 18:220. https://doi.org/10.1186/s13059-017-1349-1 [PubMed]