



Introduction
Statins are commonly used lipid-lowering drugs that target 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGCR) [1]. As first-line agents to lower plasma LDL cholesterol (LDL-C), statins have shown consistent benefits in both primary and secondary prevention for atherosclerotic cardiovascular disease (ASCVD) as supported by numerous clinical trials [2, 3]. However, even with optimal statin therapy, there are still notable residual ASCVD risks [4]. In addition, statin adherence is not clinically sufficient due to a potential risk of newly diagnosed type 2 diabetes [5] and intolerance of adverse events such as myopathy and hepatopathy [6]. Therefore, to improve lipid-lowering effects, new nonstatin lipid-lowering pharmaceutical agents have emerged.
In recent years, several gene-target drugs have been developed due to their favorable lipid-lowering effect. Cholesterol absorption inhibitors, proprotein convertase subtilisin/kexin type 9 (PCSK9) inhibitors, and bempedoic acid (BA) are several newer nonstatin medications with cholesterol-lowering effects [7, 8], while angiopoietin-like protein 3 (ANGPTL3) inhibitors and antisense oligonucleotides targeting the mRNA of apoprotein C-III (APOC3) have effects on lowering serum triglyceride levels [9]. As clinical trials investigating the effectiveness and safety of several newer nonstatin drugs are still currently in progress [10], the impact of long-term use of these drugs on morbidity and mortality remains unclear. Therefore, in our study, we aim to investigate the genetic impact of different lipid-lowering drugs on cardiometabolic diseases, the risk of cancers, and age-related outcomes by examining gene targets.
Mendelian randomization (MR) is an analytical approach that has been widely applied to investigate the causal relations between exposures and outcomes [11]. A drug-target MR analysis belongs to the MR study but only retains genetic variants in or near the target gene of the drug substance [12]. The effects of genetic variants within the encoding gene of a drug target can illustrate the potential causal effect of controlling a drug target on modulating exposures and outcomes. Therefore, we performed a two-sample MR study with drug-target MR analysis, mimicking the long-term administration of different lipid-lowering agents in randomized clinical trials, to provide genetic insights into the safety of different lipid-lowering drugs on age-related traits.
Materials and Methods
Study design and data sources
Our MR study followed the Strengthening the Reporting of Observational Studies in Epidemiology using Mendelian Randomization (STROBE-MR) guidelines (Supplementary Table 1). A two-sample MR analysis with drug-target analysis was designed. The exposures comprised LDL-cholesterol- or triglyceride-lowering genetic variants in or near various drug-target genes. The classification of lipid-lowering drugs and their target genes was based on the latest expert consensus and guidelines regarding lipid-lowering therapies [10, 13], which are summarized in Table 1. To enhance the credibility of the causal effects of gene variants, we performed positive control analyses, given the recognized benefits of lipid-lowering drugs in coronary artery disease.
Table 1. Summary of genetically proxied lipid-lowering drug targets.
| Drug effect | Drug class | Drug target | Encoding genes | Gene location (GRCh37 from ensembl) | Drug substance | Eligible IVs | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ↓LDL-C | Key Modulator | LDL Receptor | LDLR | CHR:19:11,200,038-11,244,492 | - | yes | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HMGCR inhibitors | HMG-CoA reductase | HMGCR | CHR:5:74,632,154-74,657,929 | Atorvastatin Rosuvastatin etc. | yes | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ACLY inhibitors | ATP-citrate synthase | ACLY | CHR:17:40,023,161-40,086,795 | Bempedoic acid | no | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| PCSK9 inhibitors | Proprotein Convertase Subtilisin/Kexin Type 9 | PCSK9 | CHR:1:55,505,221-55,530,525 | Evolocumab Alirocumab | yes | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TC absorption inhibitors | Niemann-Pick C1-like 1 | NPC1L1 | CHR:7:44,552,134-44,580,914 | Ezetimibe | yes | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ASO targeting ApoB mRNA | Apo B100 | APOB | CHR:2:21,224,301-21,266,945 | Mipomersen | yes | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ASO targeting CETP mRNA | Cholesteryl Ester Transfer Protein | CETP | CHR:16:56,995,762-57,017,757 | Torcetrapib | yes | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| BA sequestrants | Bile acids | - | - | Cholestyramine Colestipol | no | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ↓TG | Key Modulator | Lipoprotein Lipase | LPL | CHR:8:19,759,228-19,824,769 | - | yes | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Fibrates | Peroxisome Proliferator-Activated Receptor-alpha | PPARA | CHR:22:46,546,424-46,639,653 | Fenofibrate Gemfibrozil | no | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ANGPTL3 inhibitors | Angiopoietin-related protein 3 | ANGPTL3 | CHR:1:63,063,158-63,071,830 | Evinacumab | yes | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ASO targeting ApoC-III mRNA | Apo C-III | APOC3 | CHR:11:116,700,422-116,703,788 | Volanesorsen | yes | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Abbreviations: LDL-C, Low-Density Lipoprotein Cholesterol; TG, Total triglyceride; LDLR, Low-Density Lipoprotein Receptor; HMGCR, 3-hydroxy-3-methylglutaryl coenzyme A reductase; PCKS9, Proprotein Convertase Subtilisin/Kexin Type 9; NPC1L1, Niemann-Pick C1-like 1; APOB, Apoprotein B-100; CETP, Cholesteryl Ester Transfer Protein; LPL, Lipoprotein Lipase; ANGPTL3, Angiopoietin-related protein 3; APOC3, Apoprotein C-III. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The outcome data included multiple GWAS summary data of cardiometabolic diseases (coronary atherosclerosis, major coronary heart disease events, hypertension, type 2 diabetes, nonalcoholic fatty liver disease (NAFLD)), risk of cancers (colorectal cancer, gastric cancer, esophageal cancer, hepatocellular carcinoma, pancreatic cancer, lung cancer, thyroid cancer, bladder cancer, and cerebral tumors), and age-related outcomes (parental lifespan and longevity, telomere length, chronic obstructive pulmonary disease (COPD), Alzheimer's disease/dementia, and osteoporosis). All the data sources used in this study were derived from publicly accessible GWAS summary data of European populations, and detailed information is presented in Table 2.
Table 2. Data resources of the exposures and outcomes used in this study.
| GWAS traits | GWAS consortium | First author | Year | PMID | Population | Data type | Sample size | Case/control | Unit |
| Circulating Lipids | |||||||||
| LDL cholesterol | GLGC | Willer CJ | 2013 | 24097068 | 96% European | Continuous | 173,082 | - | SD (mg/dL) |
| Total cholesterol | GLGC | Willer CJ | 2013 | 24097068 | 96% European | Continuous | 187,365 | - | SD (mg/dL) |
| Total Triglycerides | GLGC | Willer CJ | 2013 | 24097068 | 96% European | Continuous | 177,861 | - | SD (mg/dL) |
| Age-related Outcomes | |||||||||
| Parental lifespan | UKBiobank | Timmers PR | 2019 | 30642433 | European | Continuous | 500,193 | - | SD |
| Telomere length | UKBiobank | Codd V | 2021 | 34611362 | European | Continuous | 472,174 | - | SD |
| Chronic obstructive pulmonary disease | MRC-IEU | Ben Elsworth | 2018 | - | European | Binary | 462,933 | 1,605/461,328 | LogOR |
| Alzheimer's disease/dementia | MRC-IEU | Ben Elsworth | 2018 | - | European | Binary | 399,793 | 19,255/380,538 | LogOR |
| Osteoporosis | MRC-IEU | Ben Elsworth | 2018 | - | European | Binary | 462,933 | 7,547/455,386 | LogOR |
| Cardiometabolic Diseases | |||||||||
| Coronary atherosclerosis | FinnGen | - | 2022 | - | European | Binary | 328,042 | 42,421 / 285,621 | LogOR |
| Major coronary heart disease events | FinnGen | - | 2022 | - | European | Binary | LogOR | ||
| Hypertension | MRC-IEU | Ben Elsworth | 2018 | - | European | Binary | 463010 | 54,358/408,652 | LogOR |
| Type 2 Diabetes | DIAMANTE | Mahajan A | 2018 | 30297969 | European | Binary | 898,130 | 74,124 / 824,006 | LogOR |
| Nonalcoholic fatty liver disease | Namjou B | 2019 | 31311600 | European | Binary | 9677 | 1,106/8,571 | LogOR | |
| Maligant Tumors | |||||||||
| Colorectal cancer | GECCO | Fernandez-Rozadilla C | 2023 | 36539618 | European | Binary | 185,616 | 78,473/107,143 | LogOR |
| Gastric cancer | FinnGen | - | 2022 | - | European | Binary | 260810 | 1,227/259,583 | LogOR |
| Esophageal cancer | FinnGen | - | 2022 | - | European | Binary | 260086 | 503/259,583 | LogOR |
| Pancreatic cancer | FinnGen | - | 2022 | - | European | Binary | 260832 | 1,249/259,583 | LogOR |
| Hepatocelullar cancer | FinnGen | - | 2022 | - | European | Binary | LogOR | ||
| Lung cancer | UKBiobank | Burrows K | 2021 | - | European | Binary | 374687 | 2,761/372,016 | LogOR |
| Thyroid cancer | FinnGen | - | 2022 | - | European | Binary | 261108 | 1,525/259,583 | LogOR |
| Bladder cancer | FinnGen | - | 2022 | - | European | Binary | 259667 | 84/259,583 | LogOR |
| Brain tumors | FinnGen | - | 2022 | - | European | Binary | 260357 | 774/259,583 | LogOR |
Figure 1 presents the main MR assumptions. As the main effect of lipid-lowering drugs is to reduce LDL-C or triglyceride levels, we used the associations of these selected genetic instruments with circulating lipid concentrations to proxy the pharmacological modulation of the drug-target protein (relevance assumption). The assumption is that genetic variants are not associated with confounders (independence assumption) and affect human lifespan through other pathways (exclusion restriction assumption) [14]. This study employed publicly available summary statistics for analysis, and no ethical approval was needed.
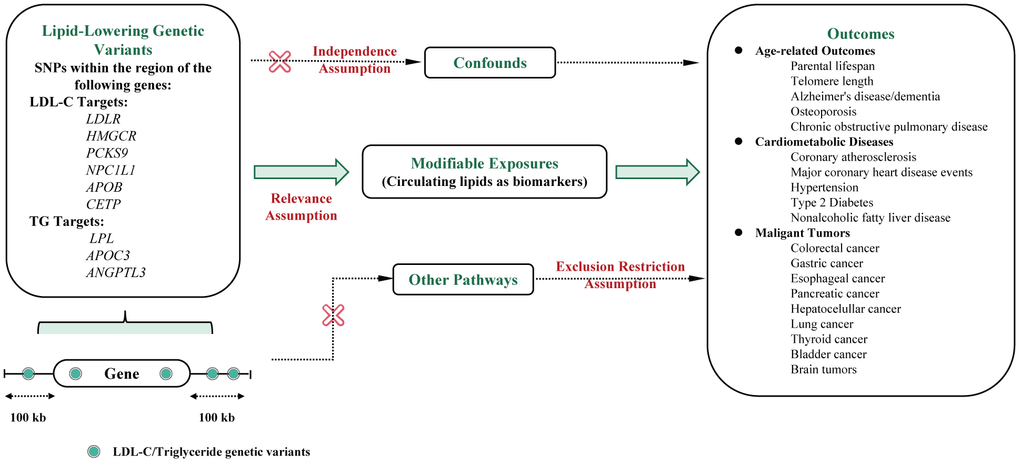
Figure 1. Flowchart of the study design and MR assumptions. Assumptions of the Mendelian randomization study: in this study, genetic instruments were selected to represent the pharmacological modulation of drug target proteins based on their associations with circulating lipid concentrations (relevance assumption). Additionally, it was assumed that the selected genetic variants are not associated with confounding factors (independence assumption). The third assumption was that genetic variants should not affect human lifespan through other pathways (exclusion restriction assumption). Abbreviations: LDL-C, Low-Density Lipoprotein Cholesterol; TC, Total cholesterol; TG, Total triglyceride; LDLR, Low-Density Lipoprotein Receptor; HMGCR, 3-hydroxy-3-methylglutaryl coenzyme A reductase; PCKS9, Proprotein Convertase Subtilisin/Kexin Type 9; NPC1L1, Niemann-Pick C1-like 1; APOB, Apoprotein B-100; CETP, Cholesteryl Ester Transfer Protein; LPL, Lipoprotein Lipase; ANGPTL3, Angiopoietin-related protein 3; APOC3, Apoprotein C-III; CHD, Major coronary heart disease; CAS, Coronary atherosclerosis; T2D, Type 2 diabetes.
Selection of genetic variants
For drug-target MR, information on pharmacologically active protein targets and their encoding genes was extracted from the DrugBank (https://go.drugbank.com/) and NCBI Gene Database (https://www.ncbi.nlm.nih.gov/gene/). A total of 11 target genes were identified, including low-Density Lipoprotein Receptor (LDLR), 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGCR), ATP-citrate synthase (ACLY), proprotein Convertase Subtilisin/Kexin Type 9 (PCKS9), Niemann-Pick C1-like 1 (NPC1L1), apoprotein B-100 (APOB), cholesteryl ester transfer protein (CETP), lipoprotein lipase (LPL), peroxisome proliferator activated receptor alpha (PPARA), angiopoietin-related protein 3 (ANGPTL3), and apoprotein C-III (APOC3). Genetic variants within these genes that encode protein targets of lipid-lowering drugs (cis-variants) were extracted from the GWAS summary data from the Global Lipids Genetics Consortium [15]. Cis-variants are defined as genetic variants located on the same DNA molecule as the target gene [16]. The serum levels of LDL-C and TG were used as proxies for LDL-C-lowering and TG-lowering targets, respectively. Drug-target SNPs, clumped to an LD threshold of r2<0.3 with a 100 kb window distance, were identified at a genome-wide level of significance (p≤5×10−8) within ±100 kb regions of the corresponding genes. PPARA and ACLY were excluded from further analysis due to insufficient numbers of SNPs identified as drug proxies. Steiger filtering [17] was also used to identify the bidirectional effects, and variants with reverse causal effects were removed accordingly. SNPs with inconsistent alleles (i.e., A/G vs. A/C) were strictly excluded.
Statistical analysis
The inverse variance weighting (IVW) method [18] was primarily used to estimate the causal effects. This approach estimates the causality of a 1 standard deviation increase in exposure to genetic predictors of outcome. Beta estimates were utilized to evaluate GWAS data with continuous outcomes, while odds ratios were used to estimate the GAWS data with binary outcomes. The association estimates of the same trait were combined using a meta-analysis of the fixed or random effects model based on heterogeneity [19].
To test the MR assumptions in the study design, we first calculated the F statistic for each instrument using the following formula: F=R2(n−1−k)/(1−R2)k, (R2 stands for the proportion of variation explained, k stands for the number of eligible SNPs, and n stands for the sample size) [20]. No significant weak instrumental bias was defined as eligible SNPs with F-statistics greater than 10. Statistical power was estimated using the mRnd website (https://shiny.cnsgenomics.com/mRnd/). To validate the results from the IVW method, we conducted sensitivity tests using MR-Egger regression, weighted median, maximum likelihood, and weighted mode methods [21]. We estimated heterogeneity and horizontal pleiotropy using the Cochran Q test and MR Egger’s intercept test. Additionally, we utilized “leave-one-out” analysis to identify heterogeneous SNPs by omitting each instrumental SNP in turn. P-values were adjusted from multiple testing using the false discovery rate (FDR, q-value) with the Benjamin-Hochberg method.
All analyses were implemented in R software version 4.1.0 using the TwosampleMR (github.com/MRCIEU/TwpSampleMR), MendelianRandomiszation, and coloc R packages. Forest plots were derived from the ggplot and ggplot2 R packages, and heatmaps were derived from the pheatmap R packages. The figure illustrating the pharmacological mechanisms of lipid-lowering drugs was drawn using the FigDraw platform (https://www.figdraw.com/).
Availability of data and materials
All data generated or analysed during this study are included in this published article and its Supplementary Information Files. The availability of all the data used in the study was summarized in Supplementary Table 2.
Consent for publication
This manuscript has not been previously published. All authors have consented to the publication of the manuscript in this journal.
Results
Effects of genetic variation in lipid-lowering drug targets on human lifespan
We used the largest GWAS dataset that contains the largest-scale lifespan-associated GWAS summary data among 500,193 European individuals [22]. Figure 2 presents the causal effects of 9 genetically proxied lipid-lowering gene targets on human lifespan or longevity-related traits. After Steiger filtration, 13 variants were selected to proxy LDL lowering through LDLR modulator, 7 for HMGCR, 12 for PCKS9, 3 for NPC1L1, 21 for LPL, 18 for APOB, 4 for CETP, 4 for ANGPTL3, and 10 for APOC3, with all F statistics greater than 10 (Supplementary Table 2).
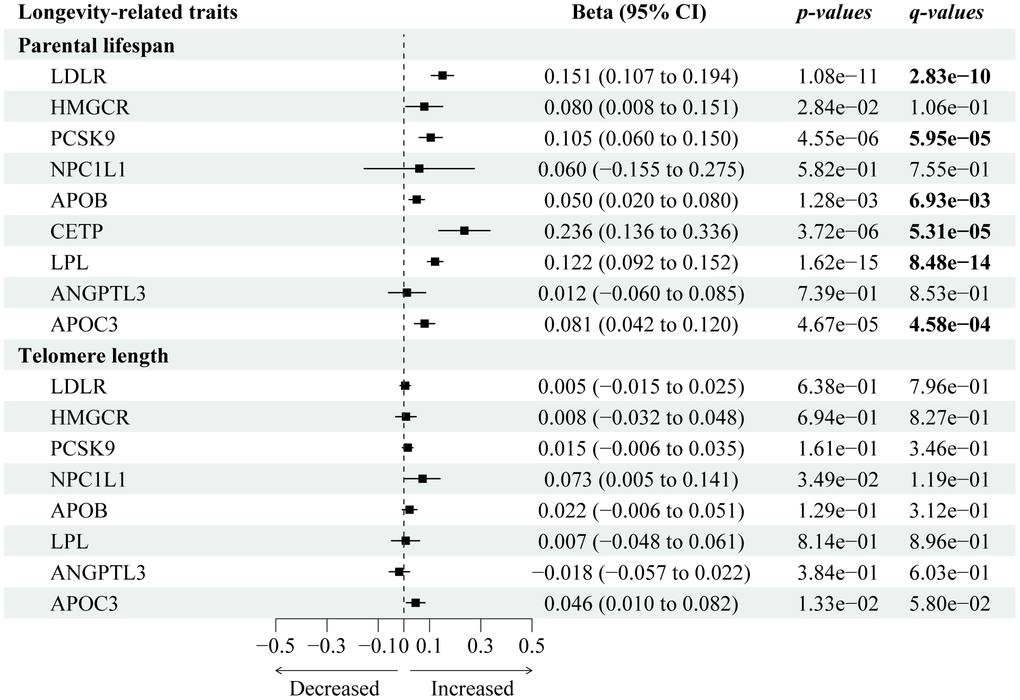
Figure 2. Forest plot visualizing the causal effects of the genetically proxied lipid-lowering drug targets on longevity-related traits. The forest plot showed the estimated effects of 1 mmol/L lower LDL-C or TG concentration by target-specific variants in each drug target gene on longevity-related traits, using the IVW method. Beta and 95% CI were used in quantitative outcomes.
We identified six lipid-lowering variants that were associated with increased lifespan, including PCKS9 (Beta 0.11; 95% CI: 0.06 to 0.15; p-IVW=4.55×10-6, FDR=5.95×10-5), CETP (Beta 0.24; 95% CI: 0.14 to 0.34; p-IVW=3.72×10-06, FDR=5.31×10-5), APOC3 (Beta 0.08; 95% CI: 0.04 to 0.12; p-IVW=4.67×10-5, FDR=4.58×10-4), LDLR (Beta 0.15; 95% CI: 0.11 to 0.19; p-IVW=1.08×10−11, FDR=2.83×10-10), and LPL (Beta 0.12; 95% CI: 0.09 to 0.195; p-IVW=1.62×10-15, FDR=8.48×10-14). There was little statistical evidence of longevity-associated effects among NPC1L1 (Beta 0.06; 95% CI: -0.16 to 0.28; p-IVW=0.58, FDR=0.76) and HMGCR (Beta 0.08; 95% CI: 0.01 to 0.15; p-IVW=0.03, FDR=0.11). Sensitivity tests (Supplementary Table 3) showed consistent trends in the estimates, with no statistical evidence of bias from horizontal pleiotropy and heterogeneity (Supplementary Table 4).
Results of positive control analysis
In the positive control analysis (Figure 3), we identified significant associations between most lipid-lowering gene targets (HMGCR, LDLR, PCSK9, NPC1L1, and LPL) and a decreased risk of both coronary atherosclerosis and major coronary heart disease (CHD) events, except for APOB, CETP, and APOC3 (only associated with reduced coronary atherosclerosis risks) and ANGPTL3 (no association with either CHD or coronary atherosclerosis).
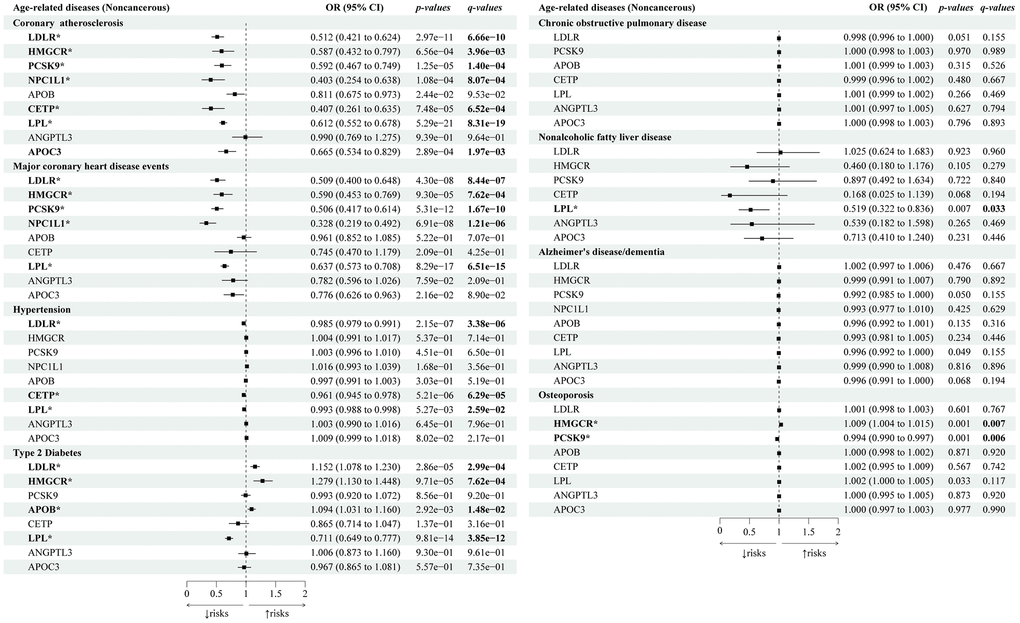
Figure 3. Causal effects of the genetically proxied lipid-lowering drug targets on age-related noncancerous diseases. The forest plot showed the estimated effects of 1 mmol/L lower LDL-C or TG concentration by target-specific variants in each drug target gene on age-related noncancerous diseases, using the IVW method. OR and 95% CI indicated the effect estimates of a 1mmol/L change of circulating lipids on outcomes.
Lipid-lowering drug targets and risk of cancers
Next, we explored the genetic associations between lipid-lowering drugs and cancers (Figure 4). We identified that lipid-lowering variants of ANGPTL3 (OR 0.83; 95% CI: 0.72 to 0.95; p-IVW=7.65×10-3, FDR=3.53×10-2) and LDLR (OR 0.83; 95% CI: 0.82 to 0.93; p-IVW=7.39×10-5, FDR=6.52×10-4) were associated with decreased risks of colorectal cancers, and ANGPTL3 (OR 0.13; 95% CI: 0.04 to 0.41; p-IVW=4.64×10-4, FDR=2.91×10-3) was also associated with lower risks of gastric cancers. Lipid-lowering LPL variants were associated with decreased risks of bladder cancers (OR 0.56; 95% CI: 0.39 to 0.79; p-IVW=1.06×10-3, FDR=5.94×10-3). Lipid-lowering APOB variants were associated with a decreased risk of thyroid cancer (OR 0.52; 95% CI: 0.36 to 0.75; p-IVW=4.29×10-4, FDR=2.81×10-3). None of the nine drug-target genes were genetically associated with esophageal cancers, pancreatic cancers, hepatocellular cancers, lung cancers, or brain cancers (with all q-values>0.05). Sensitivity tests showed consistent trends in the estimates, with no statistical evidence of bias from horizontal pleiotropy and heterogeneity (Supplementary Tables 7, 8).
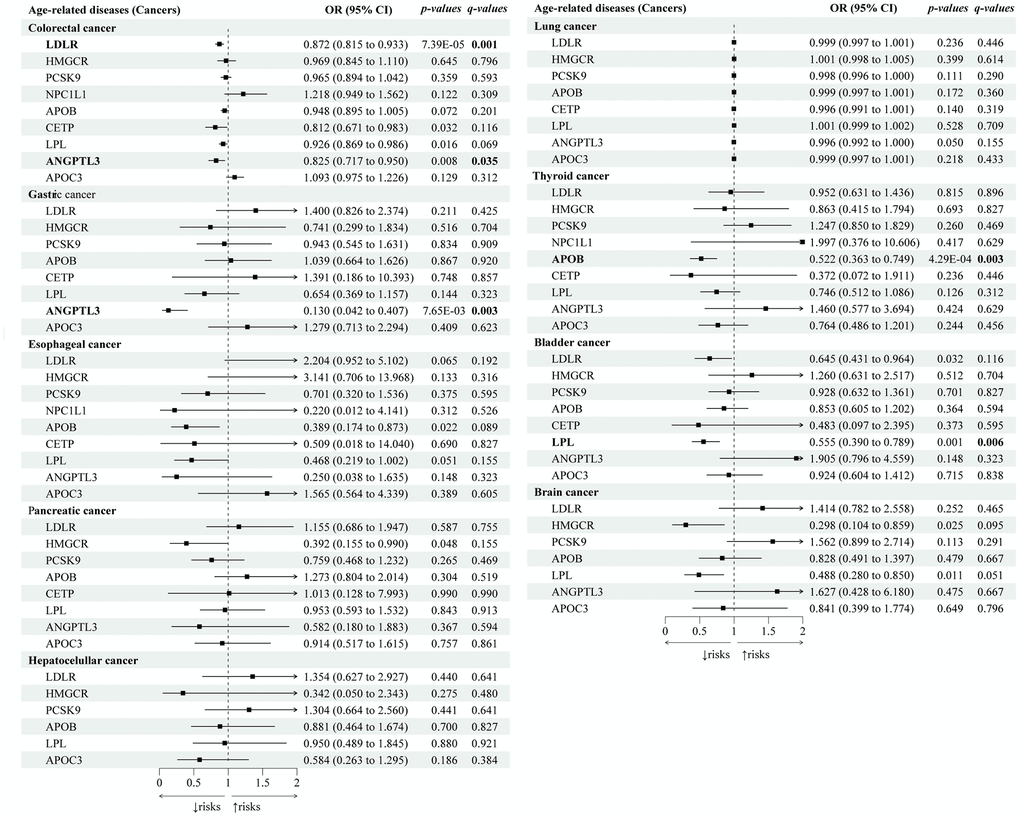
Figure 4. Causal effects of the genetically proxied lipid-lowering drug targets on cancers. The forest plot showed the estimated effects of 1 mmol/L lower LDL-C or TG concentration by target-specific variants in each drug target gene on cancers, using the IVW method. OR and 95% CI indicated the effect estimates of a 1mmol/L change of circulating lipids on outcomes.
All the statistical powers of the MR results are presented in Supplementary Table 9. The main results of the study are summarized in Figures 5, 6. The leave-one-out analyses were presented in Supplementary Figure 1.
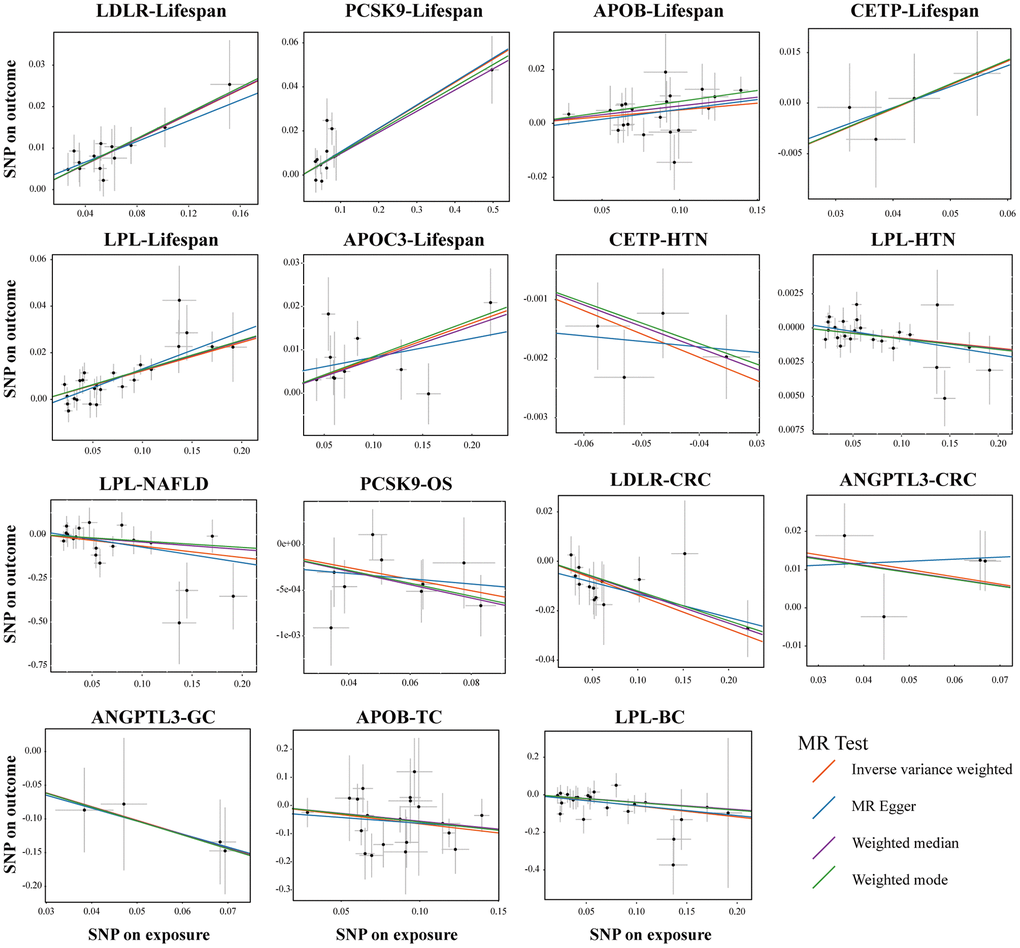
Figure 5. Sensitivity test in drug-target MR analyses. Scatter plots of four statistical tests showing representative lipid-lowering drug target genes that had a causal relationship on the different outcomes. Each black dot represents an SNP significantly associated with lipid-lowering effects. The gray lines around the dot represent the 95% confidence intervals of each SNP. Four lines generated by different MR tests were colored as red (Inverse Variance Weighted, IVW), blue (MR Egger), purple (Weighted Median), and green (Weighted mode). the X-axis represents the SNPs effects of certain lipid-lowering genes, and the Y-axis represents the SNPs effects of different outcomes. Abbreviations: HTN, hypertension; NAFLD, non-alcoholic fatty liver disease; OS, osteoporosis; CRC, colorectal cancer; GC, gastric cancer; TC, thyroid cancer, BC, bladder cancer.
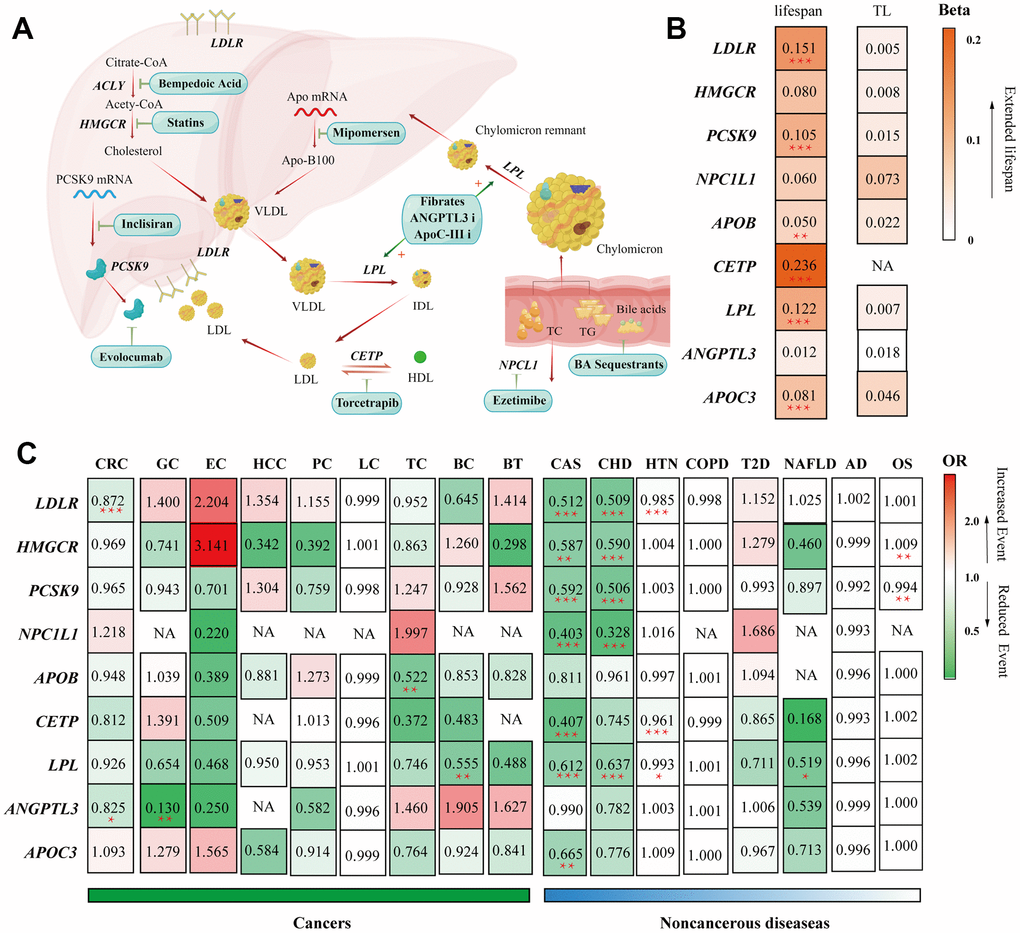
Figure 6. Summary of the study. (A) Summary of the mechanisms of action of lipid-lowering pharmaceutical agents included in our study. (B) Heatmap visualization of the Beta or OR estimates of lipid-lowering drug targets on different outcomes. The figure displays a matrix with rows representing gene targets of lipid-lowering agents and columns representing outcomes from different GWAS consortiums. The values in each square indicate the Beta or OR estimates and are color-coded based on their specific values. (B) Heatmap applied the gradually deepening orange, indicating the increasing Beta values. (C) Heatmap applied the deepening red indicating the increasing OR values and the deepening green representing the decreasing ORs. Abbreviations: TL, telomere length; CHD, Major coronary heart disease; CAS, Coronary atherosclerosis; T2D, Type 2 diabetes; HTN, hypertension; NAFLD, non-alcoholic fatty liver disease; OS, osteoporosis; CRC, colorectal cancer; GC, gastric cancer; EC, esophageal cancer; HCC, hepatocellular carcinoma; LC, lung cancer; PC, pancreatic cancer; TC, thyroid cancer, BC, bladder cancer; BT, brain tumors; COPD, chronic obstructive pulmonary disease; AD, Alzheimer's disease/dementia, OS, osteoporosis.
Discussion
Lipid metabolism has been reported to play an important role in the human lifespan and aging process [23]. However, findings were mostly derived from the data of animal models such as shorter-lived yeast, flies, and rodents, as lifespan research involving human subjects requires large amounts of time and cost [24]. By utilizing MR analysis, we were able to directly examine the genetic links between circulating lipids and longevity in humans. In this study, we used drug-targeted MR and identified several newer nonstatin lipid-lowering agents, such as those targeting LPL, ANGPTL3, and LDLR, which were associated with increased human lifespans and decreased risks of several common cancers and cardiometabolic disorders.
LDLR and LPL are two key modulators in lipid metabolism. In our study, both genes had causal effects on human lifespans. The LDLR gene can be affected by statin therapies, because interfering with the hepatic cholesterol synthesis could compensate for the increase in the de novo synthesis of LDLR and cause transport of more LDLR to the hepatocellular membranes [25]. We identified two other genes, PCSK9 and CETP, which were also associated with prolonged lifespans, and these two genes could also affect LDLR expression in lipid metabolism. The PCSK9 inhibitor prevents LDLR degradation, increases LDLR expression, and ultimately aids in the elimination of circulating LDL-C [26]. The CETP inhibitor works by increasing LDLR expression and thus lowering LDL-C levels [27]. As the discovery of new drugs such as PCSK9 or CETP inhibitors is still ongoing, their impact on cardiovascular morbidity and mortality, as well as lifespan, remains uncertain. Several clinical trials are currently in progress to determine their efficacy [10, 28]. Therefore, our findings provide promising evidence for developing these novel LDL-C-lowering agents.
In addition to LDL-C-lowering drugs through the LDLR pathway, we found that variants in the genes that encode the targets of TG through the LPL pathway were associated with lower risks of several cardiometabolic diseases. We observed that statin use was associated with increased risks of T2D, which is consistent with previous clinical trials reporting potential T2D risks with long-term statin use [29, 30]. In contrast to HMGCR inhibitors, LPL targets were associated with reduced risks of cardiovascular events, T2D, hypertension, and NAFLD, indicating the promising roles of developing newer nonstatin therapies through the LPL pathway. To gain a better understanding, it is imperative to focus on large-scale epidemiological and long-term randomized trials to specifically investigate the magnitude of the expected clinical benefit of newer nonstatin therapies.
Another finding in our MR study is the potential association between lipid-lowering genes and colorectal cancer (CRC). Although we did not find a direct causal effect of statin-targeted HMGCR on CRC risks, we confirmed that lipid-lowering LDLR variants, which are the key downstream genes in cholesterol metabolism, were associated with reduced CRC risks. Previous in vivo and in vitro studies have demonstrated a potential correlation between lipid metabolism and CRC [31, 32]. These studies have suggested that LDLR could be an important target for CRC chemoprevention. Further investigation is needed to study the role of the LDLR gene in preventing CRC. In addition, our findings indicate that targeting the lipid-lowering ANGPTL3 gene may decrease the risk of CRC. ANGPTL3 plays a role in lipid metabolism via the LPL pathway. Previous research has shown a correlation between ANGPTL3 and liver metastasis in CRC [33, 34]. Our results suggest that the TG pathways may also contribute to the development of CRC, and ANGPTL3 could be considered an additional chemopreventive target for CRC.
As a promising approach, this MR study provided genetic associations regarding statin use and human longevity traits. However, these MR results must be interpreted with caution. First, although statistical powers in most MR results exceed 90%, a few results still lack sufficient statistical powers. Thus, these results still need further validation in larger-scale GWAS data that may be available in the future. Third, our study employed multiple genetic proxies of lipid-lowering drugs, which suggests that the observed causal effects on lifespan may be attributed to genetically proxied drug-target genes rather than the whole impact of a specific drug in the real world. While drug-target MR analysis can provide insight into causal effects, it is important to note that it cannot accurately quantify the clinical benefits. Hence, there is a need for additional high-quality randomized trials or large-scale epidemiological studies.
Our study has several limitations. First, we were unable to investigate the causal effects of bempedoic acid or fibrates on human longevity due to the insufficient numbers of target-specific SNPs in the Global Lipids Genetics Consortium. Second, it is worth mentioning that our findings are limited to individuals of European ancestry and should be verified in other populations. To gain a comprehensive understanding, it is crucial to conduct large-scale epidemiological studies and long-term randomized trials to specifically examine the extent of the anticipated clinical benefits of lipid-lowering drugs on age-related outcomes.
Conclusions
Our study provides genetic evidence that newer nonstatin lipid-lowering agents have causal effects on decreased risks of several common cancers and cardiometabolic diseases. These data provide genetic insights into the potential benefits of newer nonstatin therapies. Long-term and large-scale clinical trials should also focus on the efficacy of these newer lipid-lowering drugs on age-related outcomes.
Supplementary Materials
Author Contributions
Han Chen: study concept and design, analysis and interpretation of data; drafting of the manuscript; Xinyu Tang and Wei Su: study concept and design, analysis and interpretation of data. Wei Su, Shuo Li: interpretation of data, data extraction, design and order the figures and tables. Ruoyun Yang and Hong Cheng: design and order the figures and tables, assessment of study quality. Xiaoying Zhou and Guoxin Zhang: critical revision of the manuscript for important intellectual content; obtain funding; study supervision.
Acknowledgments
The authors thank all investigators and participants from the public databases for sharing these data.
Conflicts of Interest
The authors declare that the research was conducted without any conflicts of interest.
Ethical Statement
This study does not require ethical approval as all data sources are based on publicly available summary-level data. All of these studies were approved by the relevant ethical institutions.
Funding
This work was supported by China Postdoctoral Science Foundation (No. 2023M731417), Spring Sunshine Program from the Ministry of Education of China (No.202201552) and Jiangsu province Hospital (the First Affiliated Hospital with Nanjing Medical University) Clinical Capacity Enhancement Project (JSPH-MC-2022-29).
References
- 1. Istvan ES, Deisenhofer J. Structural mechanism for statin inhibition of HMG-CoA reductase. Science. 2001; 292:1160–4. https://doi.org/10.1126/science.1059344 [PubMed]
- 2. Cholesterol Treatment Trialists’ Collaboration. Efficacy and safety of statin therapy in older people: a meta-analysis of individual participant data from 28 randomised controlled trials. Lancet. 2019; 393:407–15. https://doi.org/10.1016/S0140-6736(18)31942-1 [PubMed]
- 3. Fulcher J, O’Connell R, Voysey M, Emberson J, Blackwell L, Mihaylova B, Simes J, Collins R, Kirby A, Colhoun H, Braunwald E, La Rosa J, Pedersen TR, et al, and Cholesterol Treatment Trialists’ (CTT) Collaboration. Efficacy and safety of LDL-lowering therapy among men and women: meta-analysis of individual data from 174,000 participants in 27 randomised trials. Lancet. 2015; 385:1397–405. https://doi.org/10.1016/S0140-6736(14)61368-4 [PubMed]
- 4. Raposeiras-Roubin S, Rosselló X, Oliva B, Fernández-Friera L, Mendiguren JM, Andrés V, Bueno H, Sanz J, Martínez de Vega V, Abu-Assi E, Iñiguez A, Fernández-Ortiz A, Ibáñez B, Fuster V. Triglycerides and Residual Atherosclerotic Risk. J Am Coll Cardiol. 2021; 77:3031–41. https://doi.org/10.1016/j.jacc.2021.04.059 [PubMed]
- 5. Ridker PM, Pradhan A, MacFadyen JG, Libby P, Glynn RJ. Cardiovascular benefits and diabetes risks of statin therapy in primary prevention: an analysis from the JUPITER trial. Lancet. 2012; 380:565–71. https://doi.org/10.1016/S0140-6736(12)61190-8 [PubMed]
- 6. Stroes ES, Thompson PD, Corsini A, Vladutiu GD, Raal FJ, Ray KK, Roden M, Stein E, Tokgözoğlu L, Nordestgaard BG, Bruckert E, De Backer G, Krauss RM, et al, and European Atherosclerosis Society Consensus Panel. Statin-associated muscle symptoms: impact on statin therapy-European Atherosclerosis Society Consensus Panel Statement on Assessment, Aetiology and Management. Eur Heart J. 2015; 36:1012–22. https://doi.org/10.1093/eurheartj/ehv043 [PubMed]
- 7. Raal FJ, Rosenson RS, Reeskamp LF, Hovingh GK, Kastelein JJ, Rubba P, Ali S, Banerjee P, Chan KC, Gipe DA, Khilla N, Pordy R, Weinreich DM, et al, and ELIPSE HoFH Investigators. Evinacumab for Homozygous Familial Hypercholesterolemia. N Engl J Med. 2020; 383:711–20. https://doi.org/10.1056/NEJMoa2004215 [PubMed]
- 8. Thompson PD, Rubino J, Janik MJ, MacDougall DE, McBride SJ, Margulies JR, Newton RS. Use of ETC-1002 to treat hypercholesterolemia in patients with statin intolerance. J Clin Lipidol. 2015; 9:295–304. https://doi.org/10.1016/j.jacl.2015.03.003 [PubMed]
- 9. Akoumianakis I, Zvintzou E, Kypreos K, Filippatos TD. ANGPTL3 and Apolipoprotein C-III as Novel Lipid-Lowering Targets. Curr Atheroscler Rep. 2021; 23:20. https://doi.org/10.1007/s11883-021-00914-7 [PubMed]
- 10. Lloyd-Jones DM, Morris PB, Ballantyne CM, Birtcher KK, Covington AM, DePalma SM, Minissian MB, Orringer CE, Smith SC
Jr , Waring AA, Wilkins JT, and Writing Committee. 2022 ACC Expert Consensus Decision Pathway on the Role of Nonstatin Therapies for LDL-Cholesterol Lowering in the Management of Atherosclerotic Cardiovascular Disease Risk: A Report of the American College of Cardiology Solution Set Oversight Committee. J Am Coll Cardiol. 2022; 80:1366–418. https://doi.org/10.1016/j.jacc.2022.07.006 [PubMed] - 11. Davies NM, Holmes MV, Davey Smith G. Reading Mendelian randomisation studies: a guide, glossary, and checklist for clinicians. BMJ. 2018; 362:k601. https://doi.org/10.1136/bmj.k601 [PubMed]
- 12. Evans DS. Target Discovery for Drug Development Using Mendelian Randomization. Methods Mol Biol. 2022; 2547:1–20. https://doi.org/10.1007/978-1-0716-2573-6_1 [PubMed]
- 13. Authors/Task Force Members, ESC Committee for Practice Guidelines (CPG), and ESC National Cardiac Societies. 2019 ESC/EAS guidelines for the management of dyslipidaemias: Lipid modification to reduce cardiovascular risk. Atherosclerosis. 2019; 290:140–205. https://doi.org/10.1016/j.atherosclerosis.2019.08.014 [PubMed]
- 14. Schmidt AF, Finan C, Gordillo-Marañón M, Asselbergs FW, Freitag DF, Patel RS, Tyl B, Chopade S, Faraway R, Zwierzyna M, Hingorani AD. Genetic drug target validation using Mendelian randomisation. Nat Commun. 2020; 11:3255. https://doi.org/10.1038/s41467-020-16969-0 [PubMed]
- 15. Willer CJ, Schmidt EM, Sengupta S, Peloso GM, Gustafsson S, Kanoni S, Ganna A, Chen J, Buchkovich ML, Mora S, Beckmann JS, Bragg-Gresham JL, Chang HY, et al, and Global Lipids Genetics Consortium. Discovery and refinement of loci associated with lipid levels. Nat Genet. 2013; 45:1274–83. https://doi.org/10.1038/ng.2797 [PubMed]
- 16. Mattioli K, Oliveros W, Gerhardinger C, Andergassen D, Maass PG, Rinn JL, Melé M. Cis and trans effects differentially contribute to the evolution of promoters and enhancers. Genome Biol. 2020; 21:210. https://doi.org/10.1186/s13059-020-02110-3 [PubMed]
- 17. Hemani G, Tilling K, Davey Smith G. Orienting the causal relationship between imprecisely measured traits using GWAS summary data. PLoS Genet. 2017; 13:e1007081. https://doi.org/10.1371/journal.pgen.1007081 [PubMed]
- 18. Hemani G, Zheng J, Elsworth B, Wade KH, Haberland V, Baird D, Laurin C, Burgess S, Bowden J, Langdon R, Tan VY, Yarmolinsky J, Shihab HA, et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018; 7:e34408. https://doi.org/10.7554/eLife.34408 [PubMed]
- 19. Andrade C. Understanding the Basics of Meta-Analysis and How to Read a Forest Plot: As Simple as It Gets. J Clin Psychiatry. 2020; 81:20f13698. https://doi.org/10.4088/JCP.20f13698 [PubMed]
- 20. Staiger D, Stock JH. Instrumental variables regression with weak instruments. Econometrica. 1997; 65:557–86. https://doi.org/10.2307/2171753
- 21. Burgess S, Dudbridge F, Thompson SG. Combining information on multiple instrumental variables in Mendelian randomization: comparison of allele score and summarized data methods. Stat Med. 2016; 35:1880–906. https://doi.org/10.1002/sim.6835 [PubMed]
- 22. Timmers PR, Mounier N, Lall K, Fischer K, Ning Z, Feng X, Bretherick AD, Clark DW, Shen X, Esko T, Kutalik Z, Wilson JF, Joshi PK, and eQTLGen Consortium. Genomics of 1 million parent lifespans implicates novel pathways and common diseases and distinguishes survival chances. Elife. 2019; 8:e39856. https://doi.org/10.7554/eLife.39856 [PubMed]
- 23. Mutlu AS, Duffy J, Wang MC. Lipid metabolism and lipid signals in aging and longevity. Dev Cell. 2021; 56:1394–407. https://doi.org/10.1016/j.devcel.2021.03.034 [PubMed]
- 24. Johnson AA, Stolzing A. The role of lipid metabolism in aging, lifespan regulation, and age-related disease. Aging Cell. 2019; 18:e13048. https://doi.org/10.1111/acel.13048 [PubMed]
- 25. Goldstein JL, Brown MS. The LDL receptor. Arterioscler Thromb Vasc Biol. 2009; 29:431–8. https://doi.org/10.1161/ATVBAHA.108.179564 [PubMed]
- 26. Fitzgerald K, Frank-Kamenetsky M, Shulga-Morskaya S, Liebow A, Bettencourt BR, Sutherland JE, Hutabarat RM, Clausen VA, Karsten V, Cehelsky J, Nochur SV, Kotelianski V, Horton J, et al. Effect of an RNA interference drug on the synthesis of proprotein convertase subtilisin/kexin type 9 (PCSK9) and the concentration of serum LDL cholesterol in healthy volunteers: a randomised, single-blind, placebo-controlled, phase 1 trial. Lancet. 2014; 383:60–8. https://doi.org/10.1016/S0140-6736(13)61914-5 [PubMed]
- 27. Dong B, Singh AB, Fung C, Kan K, Liu J. CETP inhibitors downregulate hepatic LDL receptor and PCSK9 expression in vitro and in vivo through a SREBP2 dependent mechanism. Atherosclerosis. 2014; 235:449–62. https://doi.org/10.1016/j.atherosclerosis.2014.05.931 [PubMed]
- 28. Nicholls SJ, Ditmarsch M, Kastelein JJ, Rigby SP, Kling D, Curcio DL, Alp NJ, Davidson MH. Lipid lowering effects of the CETP inhibitor obicetrapib in combination with high-intensity statins: a randomized phase 2 trial. Nat Med. 2022; 28:1672–8. https://doi.org/10.1038/s41591-022-01936-7 [PubMed]
- 29. Sattar N, Preiss D, Murray HM, Welsh P, Buckley BM, de Craen AJ, Seshasai SR, McMurray JJ, Freeman DJ, Jukema JW, Macfarlane PW, Packard CJ, Stott DJ, et al. Statins and risk of incident diabetes: a collaborative meta-analysis of randomised statin trials. Lancet. 2010; 375:735–42. https://doi.org/10.1016/S0140-6736(09)61965-6 [PubMed]
- 30. Galicia-Garcia U, Jebari S, Larrea-Sebal A, Uribe KB, Siddiqi H, Ostolaza H, Benito-Vicente A, Martín C. Statin Treatment-Induced Development of Type 2 Diabetes: From Clinical Evidence to Mechanistic Insights. Int J Mol Sci. 2020; 21:4725. https://doi.org/10.3390/ijms21134725 [PubMed]
- 31. Pakiet A, Kobiela J, Stepnowski P, Sledzinski T, Mika A. Changes in lipids composition and metabolism in colorectal cancer: a review. Lipids Health Dis. 2019; 18:29. https://doi.org/10.1186/s12944-019-0977-8 [PubMed]
- 32. Tanaka T, Oyama T, Sugie S, Shimizu M. Different Susceptibilities between Apoe- and Ldlr-Deficient Mice to Inflammation-Associated Colorectal Carcinogenesis. Int J Mol Sci. 2016; 17:1806. https://doi.org/10.3390/ijms17111806 [PubMed]
- 33. Wang Y, Yi Y, Pan S, Zhang Y, Fu J, Wu X, Qin X. Angiopoietin-like protein 3 promotes colorectal cancer progression and liver metastasis partly via the mitogen-activated protein kinase 14 pathway. Mol Carcinog. 2023; 62:546–60. https://doi.org/10.1002/mc.23506 [PubMed]
- 34. Cheng J, Song X, Ao L, Chen R, Chi M, Guo Y, Zhang J, Li H, Zhao W, Guo Z, Wang X. Shared liver-like transcriptional characteristics in liver metastases and corresponding primary colorectal tumors. J Cancer. 2018; 9:1500–5. https://doi.org/10.7150/jca.23017 [PubMed]