



Introduction
Globally, the incidence of obesity is increasing [1, 2]. In general, people tend to become obese when they get older due to being hypokinetic and having decreased rates of metabolism [3]. Population-based studies have identified obesity as a risk factor for an increasing number of chronic diseases, such as cardiovascular disease, diabetes mellitus, chronic renal disease [4], several malignancies [5], and various musculoskeletal disorders [6]. However, once people get become obese, it becomes difficult to lose weight [7, 8].
Methionine is an amino acid and plays an essential role in our body. Methionine is the N-terminal amino acid in nuclear-encoded proteins, and its metabolite S-adenosyl methionine (SAM) is the cell’s main methyl group provider [9]. Methionine restriction was effective to prevent body weight gain in rodents which were fed a methionine-restricted diet starting early in life [10, 11]. Methionine restriction enhances de novo lipogenesis, lipolysis, and fatty acid oxidation, resulting in decreased in fat formation. [12]. In the present study we tested a low-methionine diet to reverse old-age-induced obesity.
All protein sources include methionine, making it impossible to strictly limit methionine by diet alone. Therefore, we have developed recombinant-methioninase (rMETase), an enzyme that degrades methionine [13–15]. We previously reported that oral rMETase prevents obesity in mice fed a high-fat diet [16]. We also reported that oral administration of rMETase-producing E. coli JM109 (E. coli JM109-rMETase) inhibited tumor growth in vivo [17]. Therefore E. coli JM109-rMETase was also tested in the present study to reverse old-age-induced obesity.
Results
Increase of mouse body weight with age
Mouse body weight increased with age. The mean body weight in mice aged 1, 2 months was 25.4 g; 2–6 months was 29.4 g (p = 0.0109 with respect to 1–2 months); 6–12 months was 41.1 g (p < 0.0001 with respect to 2–6 months); and 12–18 months was 45.5 g (p = 0.0248 with respect to 6–12 months). (Figure 1).
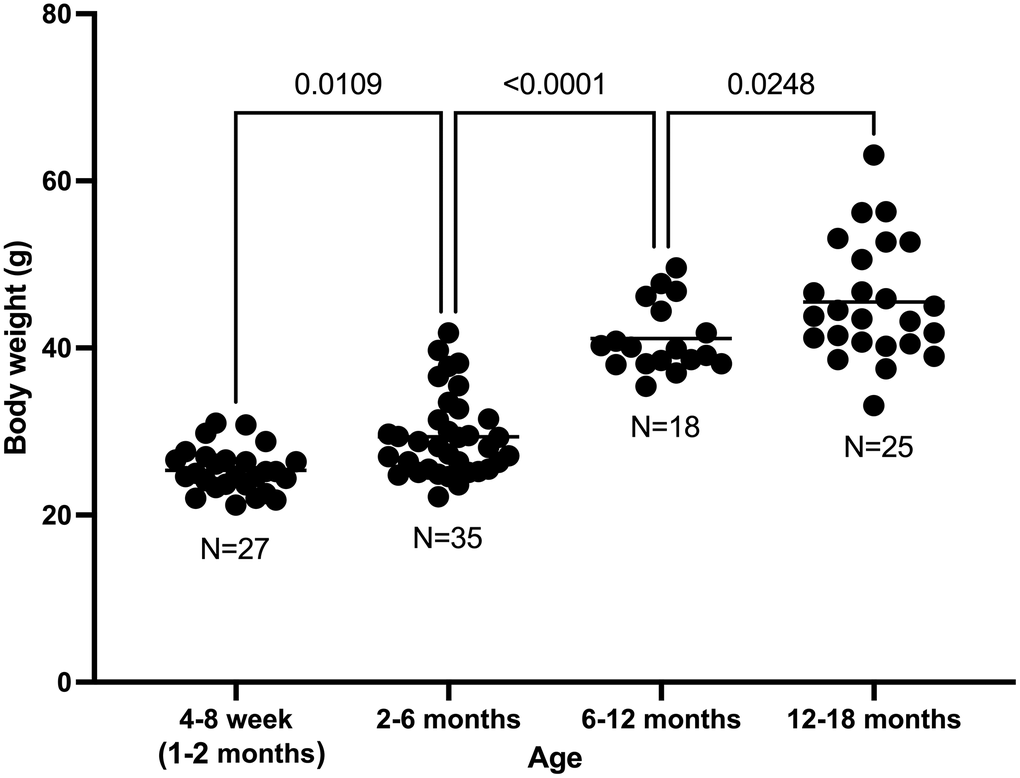
Figure 1. Body weight increase with age in C57BL/6 male mice.
Obesity was reversed by oral administration. rMETase-producing E. coli JM109
Figure 2A shows each group’s percent body weight change for 14 days. In mice fed non-recombinant E. coli JM109 cells (control group), mouse body weight did not change significantly during 14 days. The mice administered recombinant E. coli JM109-rMETase (E. coli JM109-rMETase group) decreased their mean body weight from 43.8 g to 40.7 g by day 15. This 3.1 g body weight loss is significantly higher than that of the control group (p = 0.0325). In the mice fed a methionine- and choline-deficient diet (methionine-deficient diet group), body-weight loss was 5.6 g (p = 0.0001). Mouse body weight continued to decrease until 15 days on the low-methionine diet. (Figure 2B).
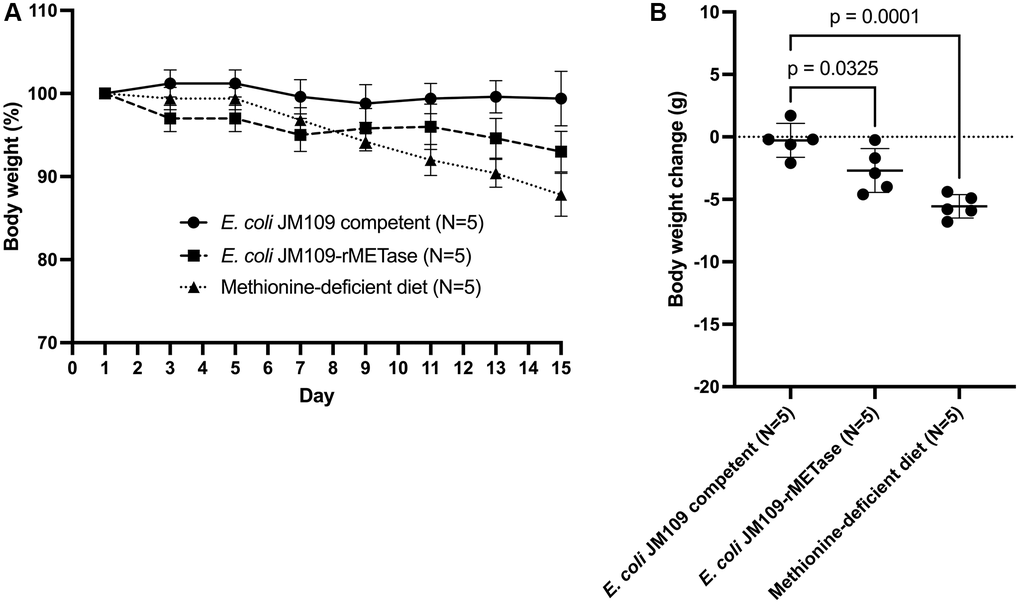
Figure 2. (A) Percent body weight change from study days 1–15. (B) Body weight change from baseline in each group on day 15.
After day 15, the body weight of the methionine-deficient-diet group continued to decrease until week 15 with a total average loss of 26.3 g. (Figure 3) Fifteen weeks after the beginning of the methionine-deficient diet, mouse body weight stabilized at 20 grams.
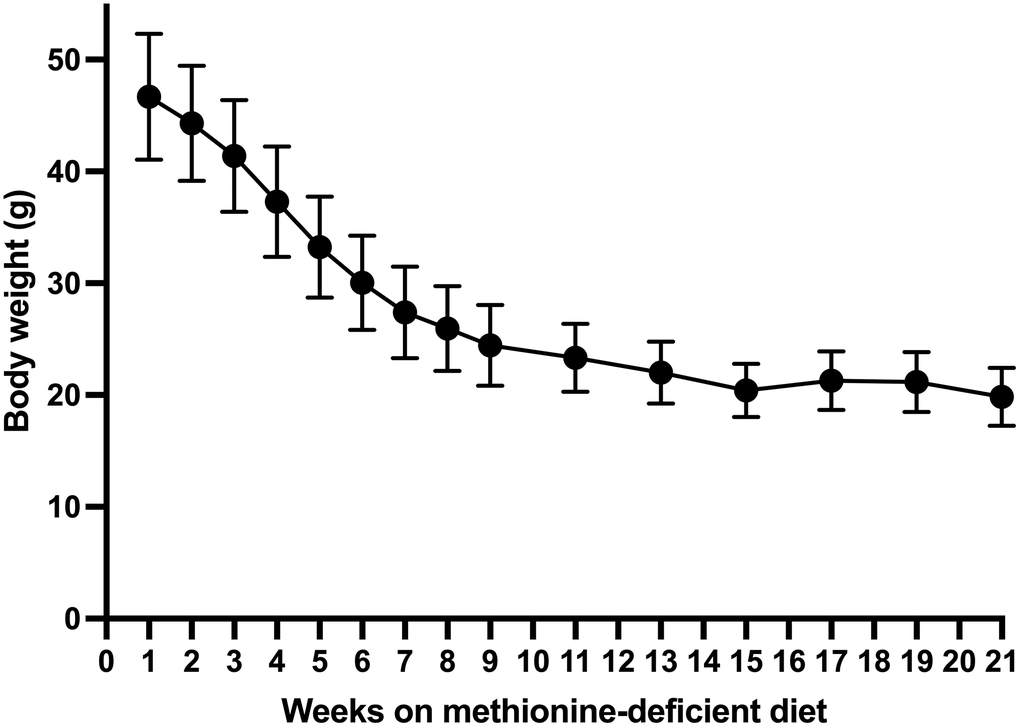
Figure 3. Long-term body weight change in mice on the methionine-deficient diet.
Methionine level
On day 15, the mean methionine level of each group was as follows: control group: 108.4 μM; E. coli JM109-rMETase group: 71.3 μM; methionine-deficient diet group: 34.0 μM. Both the methionine-deficient diet group and the E. coli JM109-rMETase-treated group showed significantly lower levels of methionine compared to the control group (Figure 4A). On day 29, the mean methionine level of each group was 81.2, 59.1, and 15.5 μM, respectively (Figure 4B). There was a negative correlation between the methionine level and negative body-weight change on day 29, R2 = 0.685, (Figure 5).
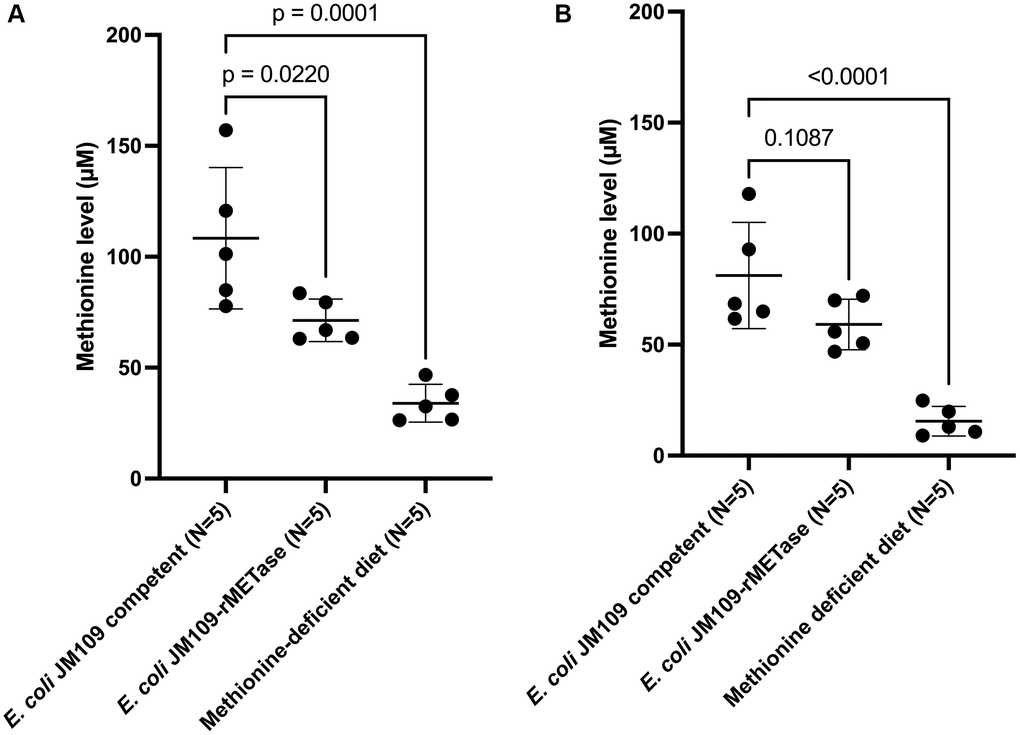
Figure 4. Blood methionine level at day 15 (A) and day 29 (B).
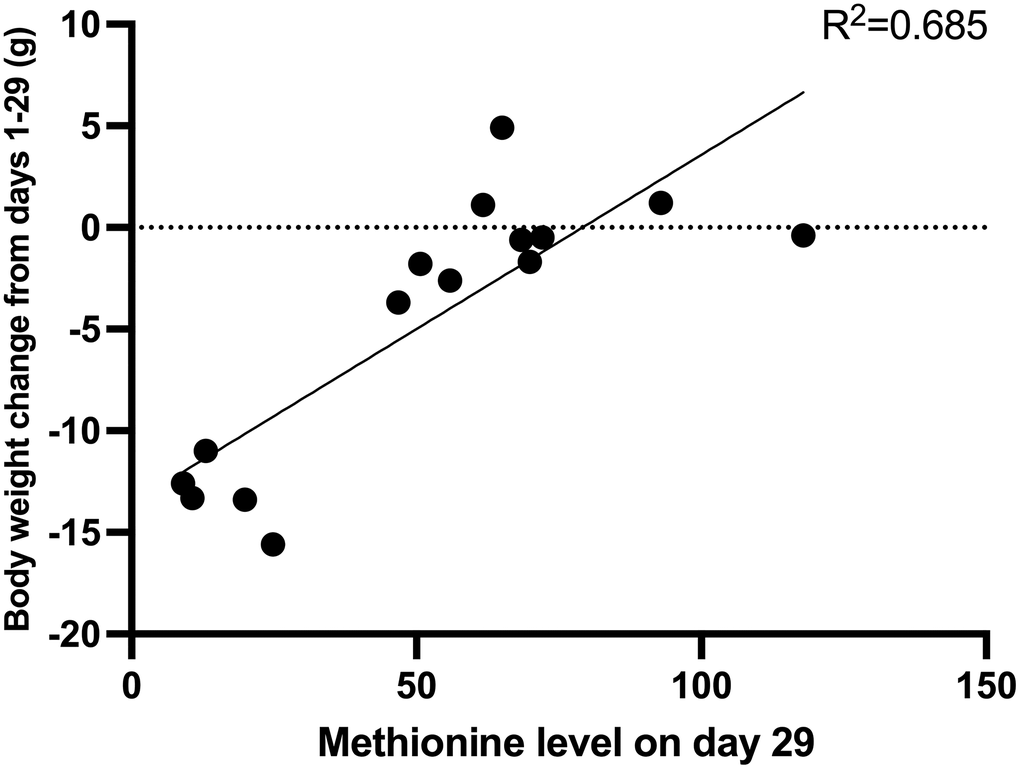
Figure 5. The correlation between methionine level and body weight change on day 29.
Stool culture of E. coli JM109-rMETase group
E. coli JM109-rMETase was detected in the E. coli JM109-rMETase group stool on day 15 and day 22 indicating incorporation in the microbiome.
Discussion
This is the first report that showed the efficacy of methionine restriction to reverse old-age-induced obesity. The correlation between age and body weight shown in the present study is consistent with previous data [11]. Similar to humans, mice become obese as they get older. Both E. coli JM109-rMETase and a methionine deficient diet lowered methionine levels and reversed obesity. The degree of efficacy of lowering methionine and reversal of obesity was superior in the methionine-deficient diet group compared to treatment with E. coli JM109-rMETase. It is notable that the body weight loss of the mice on the low methionine diet stabilized at week 15 at 20 grams. Thus, the low-methionine seems to have decreased adipose tissue and not lean muscle mass.
Different from mice, it is difficult for human beings to restrict methionine strictly only by diet. Therefore, we have developed oral methioninase and E. coli JM109-rMETase. Recently an engineered E. coli Nissle, designed to metabolize methionine via the methionine decarboxylase pathway, showed about 25% blood methionine level reduction for healthy volunteers in a phase I study [18]. Although this probiotic was developed for the treatment of homocystinuria, these results showed that this type of probiotic therapy, similar to our E. coli JM109-rMETase, can decrease the blood methionine level in human patients.
In conclusion, methionine restriction using oral installation of recombinant methioninase-producing Escherichia coli or a methionine-deficient diet is effective to reverse old-age-induced obesity, both of which have clinical potential.
Materials and Methods
Mice
C57BL/6 male mice (AntiCancer Inc., San Diego, CA, USA) aged 1-18 months were used in the present study. The mice were housed in a barrier facility with a HEPA-filtered rack under typical light/dark cycles of 12 hours. Mice were given an autoclaved laboratory rodent meal before the beginning of this study. The AntiCancer Institutional Animal Care and Use Committee’s ethics committee granted approval in accordance with National Institutes of Health Guide Assurance Number 3873-1. All experiments adhered to the Animal Research: Reporting of In Vivo Experiments (ARRIVE) 2.0 guidelines [19].
Culture of E. coli JM109-rMETase
The host strain for the expression of rMETase was E. coli JM109. The P. putida rMETase gene was previously cloned into E. coli JM109 using the plasmid pATG3131, which also contains the tetracycline (TC) resistance gene [14, 15]. The generated E. coli JM109-rMETase was pre-cultured in 5 ml of liquid Luria-Bertani (LB) medium with TC (32 g/ml) for eight hours at 37°C. The pre-culture broth was transferred overnight to 400 ml culture medium containing 32 g/ml TC. To promote the expression of rMETase, isopropyl—D-thiogalactopyranoside (IPTG) was administered at a final concentration of 0.3 mM for 4 hours at 28°C. The concentration of E. coli JM109-rMETase was adjusted to 1011 CFU/mL using phosphate-buffered saline (PBS) and 20% glycerin. E. coli JM109-rMETase was stored at −80°C until administration to mice. Non-recombinant E. coli JM109 competent cells were generated in the same manner as E. coli JM109-rMETase, except for the addition of TC and IPTG to the LB medium.
Study design
Spontaneous body-weight gain was observed in C57BL/6 mice for up to 18 months. Fifteen mice weighing more than 40 g and aged 12 to 18 months were chosen and divided into three groups of five mice each, with similar mean weight in each group: Group 1: normal diet gavaged with non-recombinant E. coli JM109 competent cells (1010/100 μL), treated twice daily (9 AM and 5 PM by oral gavage) for 14 days: Group 2: Normal diet supplemented with E. coli JM109-rMETase cells (1010/100 μL), treated by gavage twice daily (9 AM and 5 PM) for 14 days. Group 3: Mice on a methionine-deficient and choline-diet without treatment for 21 weeks. The normal diet contained 0.5% methionine, while the methionine-deficient diet was depleted of methionine, homocysteine, and choline.
IPTG (10 mM) and TC (0.5 g/L) were added to the drinking water of the mice to induce E. coli JM109-rMETase to produce rMETase and prevent plasmid shedding only in group 2 treated with E. coli JM109-rMETase [15]. Body weight was measured every two days until day 15, then every seven days until 21 weeks.
Blood and stool collection
Mouse blood was obtained through tail hemorrhage at 9 to 10 o’clock in the morning on days 15 and 29. Only the mice that received E. coli JM109-rMETase had their stool collected in the mornings of days 15, 22, and 29.
Mouse stool culture and screening for E. coli JM109-rMETase in the stool
The stool was diluted 1:10 by weight with PBS, and then mechanically disrupted. Large debris was pelletized by brief centrifugation at 200 × g. Subsequently, 100 μL of the supernatant was plated onto LB agar with 32 μg/mL TC and incubated at 37°C overnight. The screening of E. coli JM109-rMETase in the stool was performed using modified M9 agar as described previously [17].
Determination of plasma methionine level
The plasma concentration of methionine was measured using precolumn derivatization, followed by high-performance liquid chromatography separation (HPLC) based on a previously described method [20].
Statistical analysis
GraphPad Prism 9.4.0 was used to perform all statistical analyses (GraphPad Software, Inc., San Diego, CA, USA). Tukey-Kramer was used for the parametric test of group comparison. All data are represented by the mean and standard deviation. p-Values less than 0.05 were considered significant.
Author Contributions
YK performed the experiments, JR bred and maintained the mice. YK and RMH wrote the article. QH, YA, NM, KO, KH, MB, and TT analyzed the data.
Acknowledgments
This paper is dedicated to the memory of A. R. Moossa, MD, Sun Lee, MD, Professor Li Jiaxi, Masaki Kitajima, MD, Shigeo Yagi, PhD, Jack Geller, MD, Joseph R Bertino, MD, and J.A.R. Mead, PhD.
Conflicts of Interest
The authors declare no competing interests regarding this work.
Ethical Statement
The AntiCancer Institutional Animal Care and Use Committee’s ethics committee granted approval in accordance with National Institutes of Health Guide Assurance Number 3873-1.
Funding
The Robert M Hoffman Foundation for Cancer Research provided funds for this study.
References
- 1. Flegal KM, Carroll MD, Ogden CL, Curtin LR. Prevalence and trends in obesity among US adults, 1999-2008. JAMA. 2010; 303:235–41. https://doi.org/10.1001/jama.2009.2014 [PubMed]
- 2. Afshin A, Forouzanfar MH, Reitsma MB, Sur P, Estep K, Lee A, Marczak L, Mokdad AH, Moradi-Lakeh M, Naghavi M, Salama JS, Vos T, Abate KH, et al, and GBD 2015 Obesity Collaborators. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N Engl J Med. 2017; 377:13–27. https://doi.org/10.1056/NEJMoa1614362 [PubMed]
- 3. Roberts SB, Dallal GE. Energy requirements and aging. Public Health Nutr. 2005; 8:1028–36. https://doi.org/10.1079/phn2005794 [PubMed]
- 4. Lavie CJ, Milani RV, Ventura HO. Obesity and Cardiovascular Disease: Risk Factor, Paradox, and Impact of Weight Loss. J Am Coll Cardiol. American College of Cardiology Foundation. 2009; 53:1925–32. http://dx.doi.org/10.1016/j.jacc.2008.12.068.
- 5. Calle EE, Kaaks R. Overweight, obesity and cancer: epidemiological evidence and proposed mechanisms. Nat Rev Cancer. 2004; 4:579–91. https://doi.org/10.1038/nrc1408 [PubMed]
- 6. Wearing SC, Hennig EM, Byrne NM, Steele JR, Hills AP. Musculoskeletal disorders associated with obesity: a biomechanical perspective. Obes Rev. 2006; 7:239–50. https://doi.org/10.1111/j.1467-789X.2006.00251.x [PubMed]
- 7. Hill JO, Wyatt HR, Reed GW, Peters JC. Obesity and the environment: where do we go from here? Science. 2003; 299:853–5. https://doi.org/10.1126/science.1079857 [PubMed]
- 8. Proietto J. Why is treating obesity so difficult? Justification for the role of bariatric surgery. Med J Aust. 2011; 195:144–6. https://doi.org/10.5694/j.1326-5377.2011.tb03242.x [PubMed]
- 9. Sanderson SM, Gao X, Dai Z, Locasale JW. Methionine metabolism in health and cancer: a nexus of diet and precision medicine. Nat Rev Cancer. 2019; 19:625–37. https://doi.org/10.1038/s41568-019-0187-8 [PubMed]
- 10. Orentreich N, Matias JR, DeFelice A, Zimmerman JA. Low methionine ingestion by rats extends life span. J Nutr. 1993; 123:269–74. https://doi.org/10.1093/jn/123.2.269 [PubMed]
- 11. Miller RA, Buehner G, Chang Y, Harper JM, Sigler R, Smith-Wheelock M. Methionine-deficient diet extends mouse lifespan, slows immune and lens aging, alters glucose, T4, IGF-I and insulin levels, and increases hepatocyte MIF levels and stress resistance. Aging Cell. 2005; 4:119–25. https://doi.org/10.1111/j.1474-9726.2005.00152.x [PubMed]
- 12. Zhou X, He L, Wan D, Yang H, Yao K, Wu G, Wu X, Yin Y. Methionine restriction on lipid metabolism and its possible mechanisms. Amino Acids. 2016; 48:1533–40. https://doi.org/10.1007/s00726-016-2247-7 [PubMed]
- 13. Hoffman RM. Development of recombinant methioninase to target the general cancer-specific metabolic defect of methionine dependence: a 40-year odyssey. Expert Opin Biol Ther. 2015; 15:21–31. https://doi.org/10.1517/14712598.2015.963050 [PubMed]
- 14. Tan Y, Xu M, Tan X, Tan X, Wang X, Saikawa Y, Nagahama T, Sun X, Lenz M, Hoffman RM. Overexpression and large-scale production of recombinant L-methionine-alpha-deamino-gamma-mercaptomethane-lyase for novel anticancer therapy. Protein Expr Purif. 1997; 9:233–45. https://doi.org/10.1006/prep.1996.0700 [PubMed]
- 15. Takakura T, Ito T, Yagi S, Notsu Y, Itakura T, Nakamura T, Inagaki K, Esaki N, Hoffman RM, Takimoto A. High-level expression and bulk crystallization of recombinant L-methionine gamma-lyase, an anticancer agent. Appl Microbiol Biotechnol. 2006; 70:183–92. https://doi.org/10.1007/s00253-005-0038-2 [PubMed]
- 16. Tashiro Y, Han Q, Tan Y, Sugisawa N, Yamamoto J, Nishino H, Inubushi S, Higuchi T, Aoki T, Murakami M, Hoffman RM. Oral Recombinant Methioninase Prevents Obesity in Mice on a High-fat Diet. In Vivo. 2020; 34:489–94. https://doi.org/10.21873/invivo.11799 [PubMed]
- 17. Kubota Y, Han Q, Hamada K, Aoki Y, Masaki N, Obara K, Baranov A, Bouvet M, Tsunoda T, Hoffman RM. Oral Installation of Recombinant Methioninase-producing Escherichia coli into the Microbiome Inhibits Colon-cancer Growth in a Syngeneic Mouse Model. Cancer Genomics Proteomics. 2022; 19:683–91. https://doi.org/10.21873/cgp.20351 [PubMed]
- 18. Synlogic Initiates Phase 1 Study of SYNB1353 for the Treatment of Homocystinuria (HCU). 2022; https://investor.synlogictx.com/news-releases/news-release-details/synlogic-initiates-phase-1-study-synb1353-treatment.
- 19. Percie du Sert N, Ahluwalia A, Alam S, Avey MT, Baker M, Browne WJ, Clark A, Cuthill IC, Dirnagl U, Emerson M, Garner P, Holgate ST, Howells DW, et al. Reporting animal research: Explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biol. 2020; 18:e3000411. https://doi.org/10.1371/journal.pbio.3000411 [PubMed]
- 20. Sun X, Tan Y, Yang Z, Li S, Hoffman RM. A rapid HPLC method for the measurement of ultra-low plasma methionine concentrations applicable to methionine depletion therapy. Anticancer Res. 2005; 25:59–62. [PubMed]