



Introduction
Human embryonic stem cells (hESCs) are useful tools for studying human diseases and the genetic functions of healthy organisms [1, 2]. In most current methods, hESCs are cultured on mouse or human feeder cells, which appears to be the most reliable way to maintain hESC propagation [3]. However, feeder cell-dependent cultures are often problematic because animal-derived feeder cell layers pose the risk of zoonotic disease transmission and human-tissue-based feeder layers show a risk of contamination [4]. To overcome these problems, many feeder-free culture systems for supporting hESC propagation, such as mTeSR1 (Stem Cell Technologies, Vancouver, Canada), StemPro hESC SFM (Thermo Fisher, China) and HEScGRO (Millipore, China), have recently been developed [5].
A slightly different composition supporting hESC propagation is used in each feeder-free culture system mentioned above [6]. However, the cells cultured in these systems lack cell attachment, causing cell deaths or leading to overt cell differentiation [7, 8]. In recent years, experimental evidence has indicated that feeder cell layers support the undifferentiated growth of hESCs by releasing growth factors to culture media and promoting hESCs to express a variety of unidentified factors, so as to maintain hESC attachment, differentiation and proliferation [9]. Human-tissue-derived human foreskin fibroblasts (HFFs) are the most widely used feeder cells for hESC propagation because their derivation is more practical and less invasive than other human-tissue-based feeder cells [10]. Moreover, HFFs can be cultured for up to 60 passages before senescence [11]. In the present study, we hypothesized that the transcriptional profiles of hESCs were different in feeder-dependent and feeder-free culture systems. To test this hypothesis, we evaluated the changes of gene expression in three hESC lines, namely NF4, NF5 and P096 cultured in a feeder-free culture system or on HFF feeder layers.
Results
The culture systems maintain the pluripotency of hESCs
Alkaline phosphatase staining (Figure 1A), karyotype analysis (Figure 1B) and immunofluorescence staining with pluripotent markers (OCT4, Nanog, TRA-1-60 and TRA-1-181) (Figure 2) were used to characterize 3 hESC cell lines (NF4, NF5 and P096) derived from human pre-implanted embryos. The above hESC lines were positive in both AP and immunofluorescence staining, indicating that they had the characteristics of hESCs. The results of karyotype analysis showed that these hESCs exhibited normal karyotype patterns. To illustrate the differentiation potential of NF4, NF5 and P096 in feeder-free culture systems or on HFF feeder layers, teratoma formation was performed in vivo, meanwhile H & E staining of teratoma sections indicated that the NF4, NF5 and P096 cells could generate teratoma containing trilineage tissues (Figure 3).
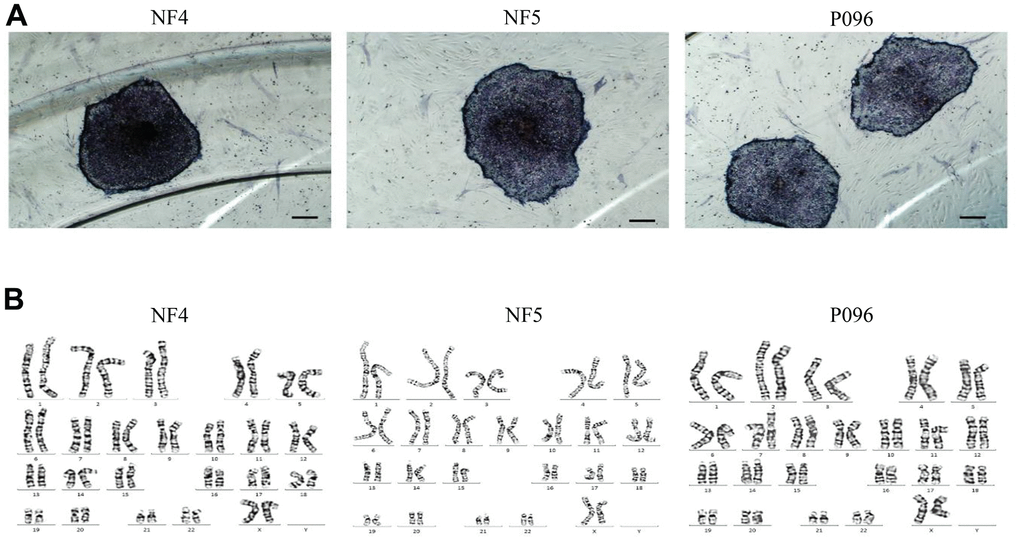
Figure 1. hESCs (NF4, NF5 and P096) grown on human foreskin fibroblast. (A) hESCs are positive for alkaline phosphatase staining. The scale bar was 200 μm. (B) Karyotype analysis revealed that these hESCs had a normal karyotype.
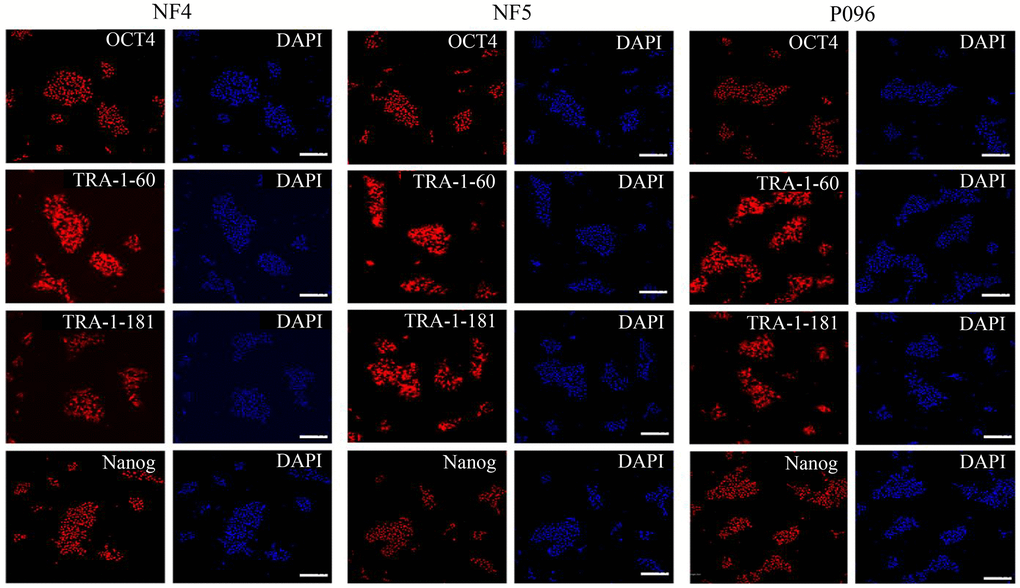
Figure 2. Immunofluorescence staining. The expressions of OCT4, Nanog, TRA-1-60 and TRA-1-181 in hESCs were detected through immunofluorescence staining. The scale bar was 100 μm.
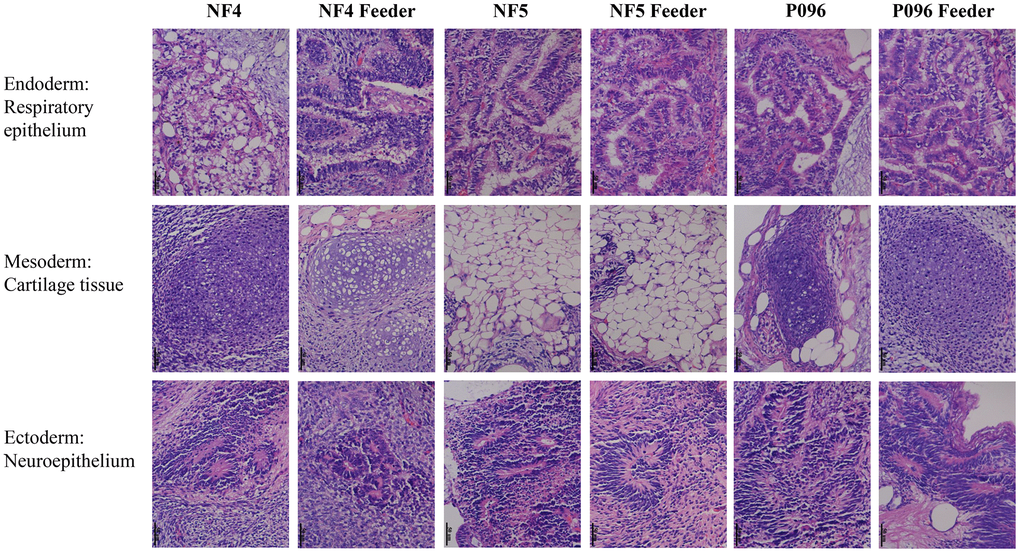
Figure 3. Teratomas containing derivatives of all the 3 germ layers. Histological evidence of germ layer differentiation in NF4, NF5 and P096 in the feeder-free culture system or on HFF feeder layers. A typical teratoma containing endoderm (respiratory epithelium), mesoderm (cartilage tissue) and ectoderm (neuroepithelium). Magnification was 400× in total.
The profiles of mRNA and lncRNA expression
Next, we performed a gene profiling analysis as well as mRNA and lncRNA microarrays for these three hESC lines cultured in a feeder-free culture system or on HFF feeder layers. The changes in the expression were defined significant when the fold change was ≥ 2.0 with a P value ≤0.05. The results are summarized in Table 1. The mRNA and lncRNA expression data was then clustered using Cluster 3.0. Using dendrogram-based methods, the samples were further clustered into two subgroups through hierarchical clustering based on similar expression patterns. The results indicated that the expression of these genes and lncRNA was significantly different in 2 different culture systems. A total of 8 genes (ABI3BP, LEFTY2, LEFTY1, KLF4, FGF4, DUSP10, MFAP5 and CABP7) were upregulated and 7 others (SUPT3H, LRAT, SLC4A5, ADAMTS4, GCNT4, ENST00000418165 and ENST00000503568) were downregulated in the feeder-free culture system compared to those on HFF feeder layers (Figure 4). Also, compared with HFF feeder layers, 6 lncRNA (ENSG00000250337.1, ENSG00000250337.1, uc.332-, XLOC_004323 and XLOC_008995) were upregulated in a feeder-free culture system, while 17 others (ENSG00000224750.2, ENSG00000172965.7, ENSG00000245526.3, ENSG00000268364.1, ENSG00000259116.1, LOC100996319, ENSG00000236373.1, DQ266889, ENSG00000272033.1, ENSG00000255506.1, ENSG00000248329.1, ENSG00000268873.1, BC069804, uc.356-, ASO1967 and ENSG00000237993.1) were downregulated in a feeder-free culture system (Figure 5). Based on the genomic relationship between the lncRNAs and the protein-coding genes, lncRNA was classified as intergenic, sense, antisense, intronic and divergent lncRNA. 23 differently-expressed lncRNA was classified as 10 intergenic, 7 antisense, 3 intronic and 3 unknown lncRNAs.
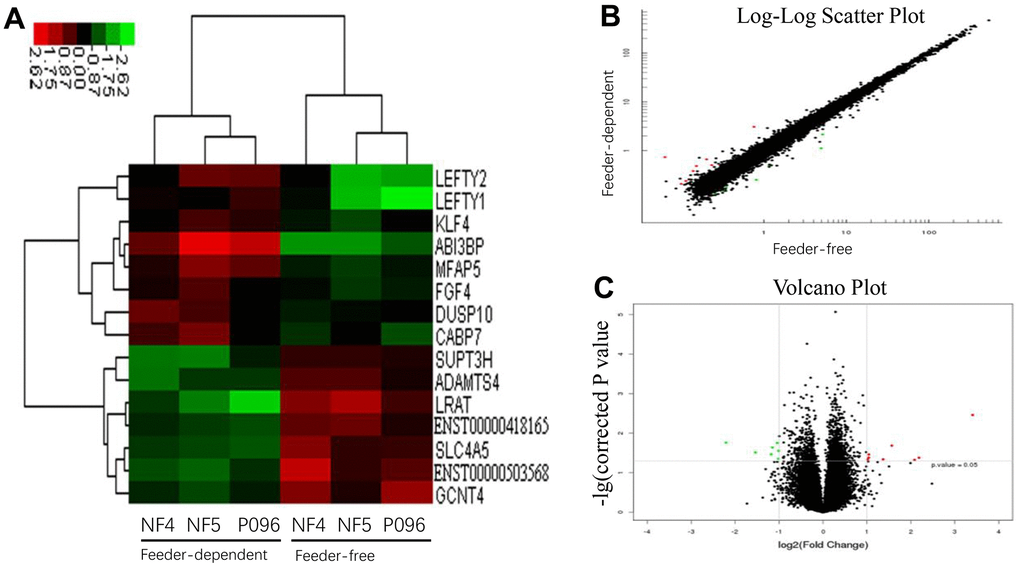
Figure 4. The profiles of the differentially-expressed genes in hESCs. (A) Hierarchical clustered heat maps show the Log2-transformed expression values of differentially-expressed genes in the feeder-free culture system and on human foreskin fibroblast feeder layers. Scatter plots (B) and volcano plots (C) of genes are of significantly different expressions.
Table 1. Summary of differentially expressed genes and lncRNAs in feeder-free culture system and on human foreskin fibroblast feeder layers.
| Up-regulation | Down-regulation | In total | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| mRNA | 8 | 7 | 15 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LncRNA | 6 | 17 | 23 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Significant differential expression was defined as probes with P≤0.05 and absolute fold change ≥2. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
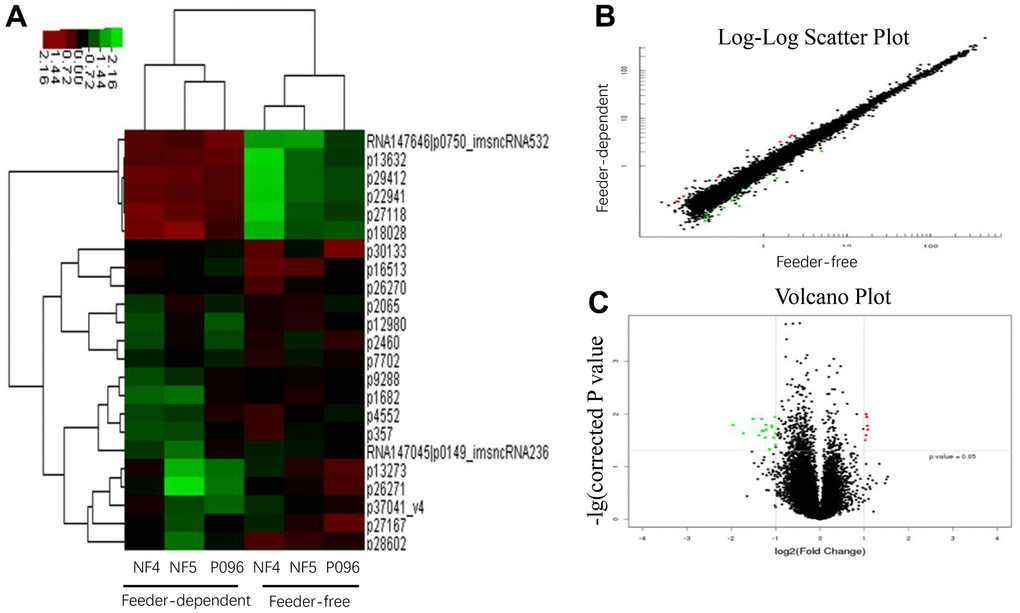
Figure 5. The profiles of the differentially-expressed lncRNA in hESCs. (A) Hierarchical clustered heat maps show the Log2-transformed expression values of differentially-expressed lncRNA in the feeder-free culture system and on human foreskin fibroblast feeder layers. Scatter plots (B) and volcano plots (C) of lncRNA are of significantly different expressions.
Gene ontology and pathway analysis
A GO analysis was performed to investigate the biological processes, cellular components and molecular functions of those differentially-expressed genes. Compared with the hESCs grown on the HFF feeder layers, the differentially-expressed genes in the top 7 GO terms during the biological process were those involved in response to growth factor stimulus (P=6.165E-05), organ morphogenesis (P=0.0004), transforming growth factor-beta (P=0.0007), cell growth (P= 0.0007), stem cell maintenance (P=0.0009), inter-male aggressive behavior (P=0.0006) and the negative regulation of chemokine (C-X-C motif) ligand 2 production (P=0.0006) (Figure 6).
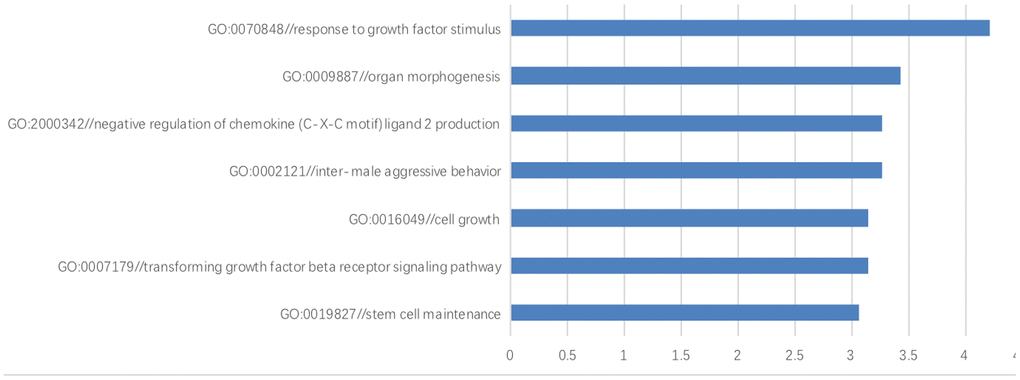
Figure 6. The gene ontology enrichment analysis. The results of gene ontology enrichment analysis revealed that these differentially-expressed genes were involved in several biological processes including growth factor stimulus, organ morphogenesis, negative regulation of chemokine (C-X-C motif) ligand 2 production, inter-male aggressive behavior, cell growth, transforming growth factor beta and stem cell maintenance.
Moreover, a KEGG pathway analysis performed with genes differentially expressed in differently-cultured stem cells showed the enrichment of gene categories. These target genes were significantly enriched in 3 different pathways (Table 2), of which the ‘TGF-beta signaling pathway’ was the most significant (P= 0.002). Next, correlated expression networks were constructed to explore the potential relationship between the differentially-expressed lncRNA and genes (Figure 7A). Uc.356-, ENST00000504246.1, uc003iol.3 and ENST00000607611.1 were identified as the node attributes of the networks, which correlated with the expression of LEFTY2, ABI3BP, MFAP5, KLF4 and FGF4 genes (Figure 7B).
Table 2. KEGG pathway analysis.
| Name | Gene symbol | P-value |
| ko04350 TGF-beta signaling pathway | LEFTY2; LEFTY1 | 0.00244515308437387 |
| ko00512 Mucin type O-Glycan biosynthesis | GCNT4 | 0.0203768610313273 |
| ko04744 Phototransduction | CABP7 | 0.0415987534109394 |
| ko04745 Phototransduction - fly | CABP7 | 0.0487835224034567 |
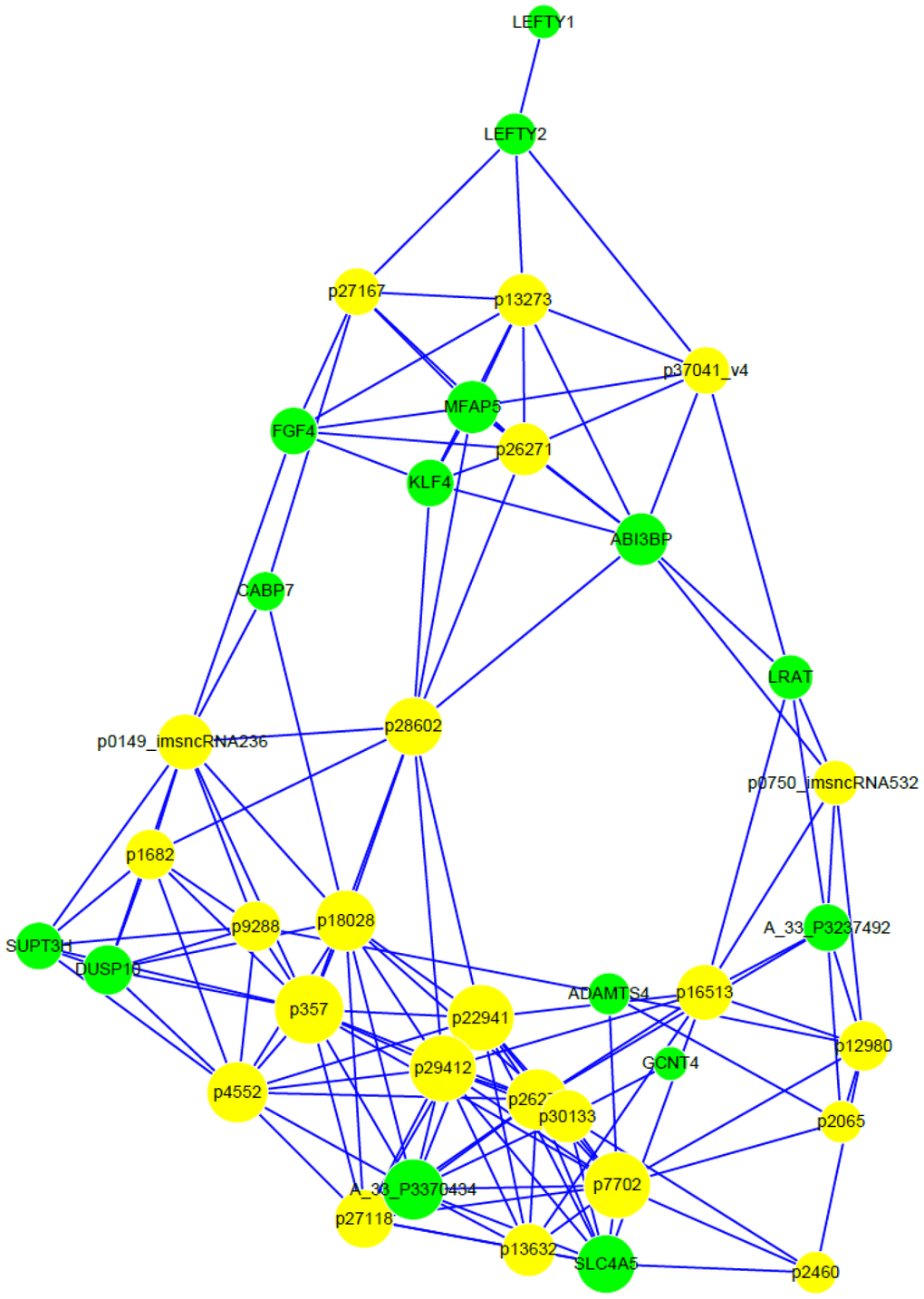
Figure 7. Correlated expression networks of differentially-expressed lncRNA and genes. Co-expression networks were constructed with differentially-expressed lncRNA and genes from 3 paired hESC lines in a feeder-free culture system vs on human foreskin fibroblast feeder layers. Pearson’s correlation > 0.99 or < -0.99, and P < 0.05.
Validation of array-based gene expression profiles through real-time RT-PCR
To validate the changes mentioned above in gene expression obtained through microarray analyses, we selected 3 core pluripotent transcript factors (SOX2, Nanog and OCT4) and 6 differentially-expressed genes (FGF4, LEFTY2, KLF4, MFAP5, LRAT and GCNT4) for qRT-PCR validation. As is shown in Figure 8, 3 core pluripotent transcript factors showed no significant differences in the expression of the feeder-free culture system or the HFF feeder layers. The RT-qPCR confirmed the changes in the expression of those 6 genes (FGF4, LEFTY2, KLF4, MFAP5, LRAT and GCNT4) observed in the microarray analyses.
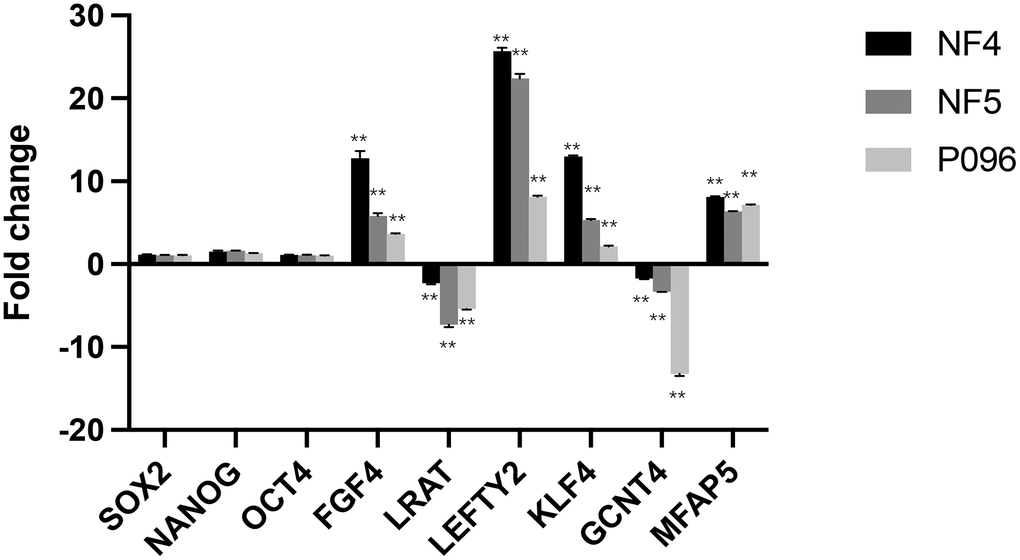
Figure 8. Validation of gene-relative changes through real-time RT-PCR. Fold changes of the gene expression in 3-line hESCs (NF4, NF5, P096) were calculated in a feeder-free culture system vs on human foreskin fibroblast feeder layers. Data was presented as mean values ± SD (n=3). The significant level was determined as P<0.05 using unpaired students’ test. **, P < 0.01.
Discussion
Approaches to developing a feeder-free culture system consisting of fully defined factors will be used to significantly improve the usefulness of hESCs in human therapeutic applications [12]. In our study, a lncRNA+ mRNA human gene expression microarray analysis was used to identify differentially-expressed lncRNA and genes in a feeder-free culture system or on HFF feeder layers, so as to exploit the secreted-factors of hESCs supporting hESC propagation. The expression of 3 pluripotent transcript factors, SOX2, Nanog and OCT4, was no different, indicating that these 2 culture systems could maintain the essential characteristics of hESCs. However, we found that the expression of ABI3BP, LEFTY2, LEFTY1, KLF4, FGF4, MFAP5, SLC4A5, ADAMTS4, DUSP10, CABP7, SUPT3H, LRAT and GCNT4 was significantly different in the feeder-free culture system and on HFF feeder layers. To predict the potential functions of these differential genes identified in this study, a GO analysis was performed using the coding genes associated with the significantly differentially-expressed mRNA. The results revealed that these genes were involved in biological processes including growth factor stimulus, transforming growth factor-beta, cell growth and stem cell maintenance, all of which were important in stem cell growth, proliferation as well as differentiation.
Amongst these genes, LEFTY1 and LEFTY2 controlled the balance between self-renewal and pluripotent differentiation in mouse embryonic stem cells [13]. The differentiation potential of embryonic stem cells increased due to LEFTY1 knockdown, whereas the self-renewal of embryonic stem cells was enhanced by LEFTY2 knockdown [14]. Interestingly, the function of KLF4 was to activate the LEFTY1 promoter cooperating with Oct3/4 and Sox2, suggesting that a network was formed by LEFTY1, LEFTY2 and KLF4 in controlling the hESC propagation [14]. FGF4 has been found to specifically contribute to the differentiation of hESC development at early stages [15]. The inhibition of FGF4 signaling pathway enhanced the derivation and propagation of stem cells at undifferentiated states [16]. ABI3BP is an extracellular matrix protein [17], which was shown as a multifunctional autocrine/paracrine factor regulating mesenchymal stem cells in a previous report [18]. A related function of MFAP5 is that it can remodel extracellular matrixes that may play a role in supporting hESC growth [19]. A protein that transports sodium and bicarbonate across the cell membrane as well as regulates cellular pH is encoded through SLC4A5 [20]. ADAMTS-4 is a metalloprotease that plays a role in aggrecan degradation in an extracellular matrix [21]. These 4 genes, namely ABI3BP, MFAP5, SLC4A5 and ADAMTS-4, express proteins of extracellular matrixes, which have been widely demonstrated to support hESC propagation. The function of remaining genes in hESC growth, namely DUSP10, CABP7, SUPT3H, LRAT and GCNT4, are rarely reported. DUSP10 has been found to negatively regulate the inflammatory response and colorectal cancer cell growth [22]. MFAP5 is a suppressor of oral tongue squamous cell carcinoma and cervical cancer [23, 24]. The value of these factors contributes to maintaining the propagation of hESCs, which needs to be further investigated.
In recent years, accumulating documents have revealed that lncRNA was involved in the differentiation of hPSCs by affecting the level of transcription factors [25, 26]. However, the precise mechanism of how lncRNAs regulate transcription factors in hPSC propagation remains largely unknown. In the present study, 6 and 17 lncRNAs were significantly up- and down- regulated on HFF feeder layers respectively. Of the dysregulated lncRNA, we identified that 10 of them were transcribed from intergenic regions. The remaining 7 and 3 lncRNA was respectively transcribed from the antisense and intronic region. Furthermore, we employed a lncRNA-mRNA network analysis to identify interactions between differentially-expressed genes and lncRNA. Our results showed that the expression of Uc.356-, ENST00000504246.1, uc003iol.3 and ENST00000607611.1 was correlated with LEFTY2, ABI3BP, MFAP5, KLF4 and FGF4. Interestingly, these 5 genes were identified in supporting hPSC propagation in this study, suggesting that a network controlling the hESC propagation was formed by Uc.356-, ENST00000504246.1, uc003iol.3, ENST00000607611.1, LEFTY2, ABI3BP, MFAP5, KLF4 and FGF4.
In conclusion, we identified that 23 lncRNAs and 15 genes in hPSCs were significantly differentially expressed in the feeder-free culture system and on HFF feeder layers. In addition, our data indicated that most of these genes participated in specific biological processes contributing to hPSC propagation. Our findings here provided a piece of newfound information regarding the potential roles of these lncRNAs and genes in supporting hPSC propagation. Further research is required to elucidate the detailed molecular mechanisms underlying the action of these lncRNAs and genes.
Materials and Methods
Derivation and culture of hESCs
The hESCs derived from human pre-implanted embryos and HFF cells were performed in compliance with the protocols approved by the Reproductive Study Ethics Committee of Nanfang Hospital, Southern Medical University (Research License AF/SC-08/03.2). Embryos were obtained from the donor couples undergoing in vitro fertilization (IVF) at the Center of Reproductive Medicine of Nanfang Hospital. Informed consent forms were signed for the participation of those couples in this study.
The blastocysts discarded in a routine IVF program were used to derive the hESC lines. The inner cell mass (ICM) was mechanically isolated with an insulin syringe needle through a stereoscope, which was then cultured on HFF feeder layers that had been mitotically inactivated with 10 μg/ml mitomycin C (M4287, Sigma, Beijing, China). HFFs were isolated from children’s foreskin after circumcision. The procedure was approved by the Reproductive Study Ethics Committee of Southern Medical University Nanfang Hospital (Research License AF/SC-08/03.2). An informed consent from the legal guardians was obtained with signed forms. Briefly, the foreskin was cut into small pieces and digested with 0.25% trypsin (Invitrogen, Beijing, China). After digestion, cells were placed into tissue culture flasks in 90% Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, China) supplied with 10% HS and 1% penicillin–streptomycin (PS; HyClone Laboratories, Inc., Logan, UT, USA). The medium was changed daily. HFFs were passaged through trypsin at a split ratio of 1:6 every 2-3 days.
The primary hESC colonies were sliced into equal-sized pieces and removed from the dish mechanically with a syringe needle through a stereoscope. The cell clusters were then transferred into new dishes containing fresh feeder cells. The hESC colonies were passed to new dishes every 4-5 days. The culture medium of hESCs was mTeSR1 (Stem Cell Technologies, Vancouver, Canada), which was replenished daily. At ten passages, 3 hESC lines (NF4, NF5 and P096) were chosen to be cultured in a feeder-dependent (HFF feeder layer) and a feeder-free culture system (matrigel-coated dish) with mTeSR1 as the culture medium (Stem Cell Technologies, Vancouver, Canada). The cells were maintained in a 37° C incubator with 5% CO2.
Alkaline phosphatase staining, karyotype analysis and immunofluorescence staining
AP staining, karyotype analysis and immunofluorescence staining were performed on hESCs with a passage between 6 and 8. AP staining was carried out using an AP detection kit (Millipore, Beijing, China) according to the manufacturer’s protocol. Briefly, cells were fixed in 4% paraformaldehyde for 20 min at room temperature. After washing thrice with 1×PBS, cells were treated in the ALP buffer for 5 min and stained in BCIP (5-bromo-4-chloro-3-indolyl-phosphate) as well as NBT (nitro-blue-tetrazolium) for another 15 min. The images of AP staining were captured with a Zeiss microscope.
For karyotype analysis, cells were first treated with 0.2 μg/ml colcemid (Gibco Invitrogen, Beijing, China) for 130 min. The hESC colonies were mechanically removed from the feeder layers through a stereoscope after washing twice in DMEM/F12. The cells were incubated in 0.4% kalium chloratum for 20 min at 37° C and then fixed in methanol: acetic acid (3:1, v/v) for 3 min, followed by Giemsa staining. 20 cells per group were randomly selected for the karyotype analysis.
For immunofluorescence staining, cells were washed in 1×PBS (Gibco) twice before and after being fixed in 4% paraformaldehyde for 20 min at room temperature. Cells were then permeabilized in 0.3% triton X-100 (Sigma, Beijing, China) for 1 h, followed by a block in 10% goat serum in PBS. After then, cells were incubated with primary antibodies of interest (anti-OCT4 antibody, CST; anti-Nanog antibody, CST; anti-TRA-1-60 antibody, CST; anti-TRA-1-181 antibody, CST) at 4° C overnight, followed by another incubation with second antibodies (Goat anti-Mouse, Alexa Fluor 568, Abcam) at room temperature for 1 h. DAPI was used for nuclear staining. Immunostaining images were captured with a Leica DMI6000B microscope (Leica Microsystems GmbH, Germany).
Teratoma formation assay
About 3×106 hESCs were suspended in 25% matrigel with DMEM/F12 (1:3), and then injected subcutaneously into immunodeficient nude mice, which were humanely sacrificed 8 weeks after injection. The teratomas were excised and processed for hematoxylin-eosin staining (H & E). Tissues were fixed in 4% paraformaldehyde for 24 h, followed by dehydration in gradient ethanol (70%, 85%, 95% and 100%) and paraffin embedding. For H&E staining, 6 micrometer sections were deparaffinized twice in xylene (each for 5 min), rehydrated in gradient ethanol (100%, 90% and 70%), stained with hematoxylin for 4 min, washed in ddH2O for 5 min, treated with 1.5% hydrochloric acid-75% ethanol for 10 s and washed in ddH2O for 5 min, which was followed by PBS for 2 min, they were then stained with eosin for about 20 s and dehydrated in gradient ethanol as well as xylene, after which they were placed in xylene and neutral resin mounting medium. All animal protocols were approved by the ethics committee at Hainan Medical University.
Sample collection and RNA extraction
The hESCs on a HFF feeder layer with a passage of 3 (totally 13 passages) were mechanically removed from feeder layers through a stereoscope after washing twice in DMEM/F12, after which they were collected concurrently in a trizol (Invitrogen, Beijing, China) and stored at -80° C. Total RNA was extracted from the frozen trizol block and purified using NucleoSpin RNA clean-up (Macherey-Nagel, Duren, Germany). The concentration and purity of RNA were determined by a NanoDrop ND-2000 spectrophotometer (NanoDrop Technologies, Thermo Scientifics, USA), and its integrity was evaluated through formaldehyde denaturing gel electrophoresis. Only the RNA samples with a total RNA quantity ≥ 1 μg and an A260/ 280 ratio ≥ 2.0 showing clear bands of 28 s as well as 18 s rRNA with a brightness ratio of near 2:1 in the formaldehyde denaturing gel were used for the microarray analysis.
Microarray and analysis of gene expression
A total 200 ng of RNA was amplified and reversely transcribed to double-stranded cDNA and subsequently labelled through fluorescent dye (Cy3) according to the manufacturer’s protocol for CapitalBio cDNA Amplification and Labeling Kit (CapitalBio, Beijing, China). The labelled cDNA was then purified and hybridized to Agilent Human lncRNA + mRNA Array V3.0 (Agilent, USA). Images were scanned with an Agilent microarray scanner (G2565CA) and analyzed using Agilent feature extraction (v10.7). The raw data was summarized and normalized using GeneSpring (Agilent V12.0). After completing quantile normalization of the raw data, differentially expressed lncRNA and genes were identified using threshold values of ≥2- and ≤−2-fold changes and a Benjamini-Hochberg corrected P value of ≤0.05. Next, the Molecular Annotation System (CB-MAS) V3.0 (http://bioinfo.capitalbio.com/mas3/) was used to perform gene ontology and pathway analysis. The GO terms consisted of three families: biological process (BP), cellular component (CC) and molecular function (MF).
In addition, signaling pathway analysis of these genes was performed according to KEGG PATHWAY Database (http://www.kegg.jp/kegg/pathway.html), which provided information about the significant enrichment of genes with differential expression. Finally, the differentially-expressed lncRNA and genes with a Pearson correlation coefficient no. < 0.99 were chosen to draw a co-expression network. The target prediction was performed based on their close correlations (the minimum Pearson’s correlation coefficient was -0.7) to a group of differentially-expressed protein-coding genes.
Real-time PCR
The total RNA of these 3 hESC lines was isolated using a RNeasy mini kit (QIAGEN, Beijing, China) and was reversely transcribed using a reverse transcription system kit (Promega, USA) in accordance with the manufacturer’s instructions. Real-time PCR was performed with the oligos for the selected genes with Brilliant III Ultra-Fast qPCR Master Mix (Agilent Technologies, USA). The real-time PCR conditions are shown below: 95° C for 2 min; 30s at 95° C and 1 min at 60° C with a total of 45 cycles; melt curve stage (95° C for 15 s, 55° C for 1 min and 90° C for30 s). GAPDH was used as an internal control, and the 2−ΔΔCT method was used to calculate the mRNA fold changes (feeder-free cultured hESCs/feeder-dependent cultured hESCs). The primers used in this study are listed in Table 3.
Table 3. The primers used in this study.
| Genes | Primer | |
| SOX2 | Forward | GGTTACCTCTTCCTCCCACTCC |
| Reverse | CCCTCCCATTTCCCTCGTTT | |
| NANOG | Forward | ACCTATGCCTGTGATTTGTGG |
| Reverse | AGTGGGTTGTTTGCCTTTGG | |
| OCT4 | Forward | GGCCTGCATGAGGGTTTCT |
| Reverse | CCCCTGAGAAAGGAGACCCA | |
| FGF4 | Forward | CTCGCCCTTCTTCACCGATG |
| Reverse | GTAGGACTCGTAGGCGTTGTA | |
| LEFTY2 | Forward | TGGACCTCAGGGACTATGGAG |
| Reverse | CCGAGGCGATACACTGTCG | |
| KLF4 | Forward | CGGACATCAACGACGTGAG |
| Reverse | GACGCCTTCAGCACGAACT | |
| MFAP5 | Forward | TGAATGATCCCGCTACAGATGA |
| Reverse | TCCTTACAGACAAGACGAGAGC | |
| LRAT | Forward | TGATGCCCGACATCCTGTTG |
| Reverse | ATGTTAGCTCCGTAGGCGAAG | |
| GCNT4 | Forward | TCCCTAAGTACCTCGCCTTTT |
| Reverse | GCTCCTGTTCATAGATACCCGAA | |
| GAPDH | Forward | CATGTTCGTCATGGGTGTGAACCA |
| Reverse | AGTGATGGCATGGACTGTGGTCAT |
Statistical analysis
Results were presented as mean ± SD with at least 3 biological repeats using a GraphPad prism. Significance level was determined by unpaired two-tailed students’ t tests, and P< 0.05 was considered statistically significant.
Author Contributions
YM and LH designed this study. LX, JZ, ZL, BW, XZ, YW, FS, LH, ZW, SQ, QL, JW performed the experiments. LX, ZJ, ZL, and YM analyzed the data. LX, ZJ, and ZL wrote the draft manuscript, drew the figures and tables. YM, LH, and ZL revised the manuscript. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Ethical Statement and Consent
The hESCs derived from human pre-implanted embryos and HFF cells were performed in compliance with the protocols approved by the Reproductive Study Ethics Committee of Nanfang Hospital, Southern Medical University (Research License AF/SC-08/03.2). Embryos were obtained from the donor couples undergoing in vitro fertilization (IVF) at the Center of Reproductive Medicine of Nanfang Hospital. Informed consent forms were signed for the participation of those couples in this study.
Funding
This study was supported by Hainan Provincial Natural Science Foundation of China (No. SQ2020SHFZ0621, 2019CXTD408), International Science and Technology Cooperation Program of China (Beijing, China; No. 2014DFA30180), National Natural Science Foundation of China (Beijing, China; No. 81960283, 82072880, 81471464) and the President Foundation of Nanfang Hospital, Southern Medical University (2018C035).
References
- 1. Saito-Diaz K, Zeltner N. Induced pluripotent stem cells for disease modeling, cell therapy and drug discovery in genetic autonomic disorders: a review. Clin Auton Res. 2019; 29:367–84. https://doi.org/10.1007/s10286-018-00587-4 [PubMed]
- 2. Yabut O, Bernstein HS. The promise of human embryonic stem cells in aging-associated diseases. Aging (Albany NY). 2011; 3:494–508. https://doi.org/10.18632/aging.100328 [PubMed]
- 3. Hynds RE, Bonfanti P, Janes SM. Regenerating human epithelia with cultured stem cells: feeder cells, organoids and beyond. EMBO Mol Med. 2018; 10:139–50. https://doi.org/10.15252/emmm.201708213 [PubMed]
- 4. Tannenbaum SE, Singer O, Gil Y, Berman-Zaken Y, Ilouz N, Khaner H, Haimov M, Reubinoff BE. Hadassah, provider of “Regulatory-Ready” pluripotent clinical-grade stem cell banks. Stem Cell Res. 2020; 42:101670. https://doi.org/10.1016/j.scr.2019.101670 [PubMed]
- 5. Wang L, Zhang R, Ma R, Jia G, Jian S, Zeng X, Xiong Z, Li B, Li C, Lv Z, Bai X. Establishment of a feeder and serum-free culture system for human embryonic stem cells. Zygote. 2020; 28:175–82. https://doi.org/10.1017/S0967199419000625 [PubMed]
- 6. Akopian V, Andrews PW, Beil S, Benvenisty N, Brehm J, Christie M, Ford A, Fox V, Gokhale PJ, Healy L, Holm F, Hovatta O, Knowles BB, et al, and International Stem Cell Initiative Consortium. Comparison of defined culture systems for feeder cell free propagation of human embryonic stem cells. In Vitro Cell Dev Biol Anim. 2010; 46:247–58. https://doi.org/10.1007/s11626-010-9297-z [PubMed]
- 7. Chen YH, Pruett-Miller SM. Improving single-cell cloning workflow for gene editing in human pluripotent stem cells. Stem Cell Res. 2018; 31:186–92. https://doi.org/10.1016/j.scr.2018.08.003 [PubMed]
- 8. Dakhore S, Nayer B, Hasegawa K. Human Pluripotent Stem Cell Culture: Current Status, Challenges, and Advancement. Stem Cells Int. 2018; 2018:7396905. https://doi.org/10.1155/2018/7396905 [PubMed]
- 9. Nava MM, Piuma A, Figliuzzi M, Cattaneo I, Bonandrini B, Zandrini T, Cerullo G, Osellame R, Remuzzi A, Raimondi MT. Two-photon polymerized “nichoid” substrates maintain function of pluripotent stem cells when expanded under feeder-free conditions. Stem Cell Res Ther. 2016; 7:132. https://doi.org/10.1186/s13287-016-0387-z [PubMed]
- 10. Llames S, García-Pérez E, Meana Á, Larcher F, del Río M. Feeder Layer Cell Actions and Applications. Tissue Eng Part B Rev. 2015; 21:345–53. https://doi.org/10.1089/ten.TEB.2014.0547 [PubMed]
- 11. Oliveira T, Costa I, Marinho V, Carvalho V, Uchôa K, Ayres C, Teixeira S, Vasconcelos D. Human foreskin fibroblasts: from waste bag to important biomedical applications. J Clin Urol. 2018; 11:385–94. https://doi.org/10.1177/2051415818761526
- 12. Sams A, Powers MJ. Feeder-free substrates for pluripotent stem cell culture. Methods Mol Biol. 2013; 997:73–89. https://doi.org/10.1007/978-1-62703-348-0_7 [PubMed]
- 13. Kim DK, Cha Y, Ahn HJ, Kim G, Park KS. Lefty1 and lefty2 control the balance between self-renewal and pluripotent differentiation of mouse embryonic stem cells. Stem Cells Dev. 2014; 23:457–66. https://doi.org/10.1089/scd.2013.0220 [PubMed]
- 14. Nakatake Y, Fukui N, Iwamatsu Y, Masui S, Takahashi K, Yagi R, Yagi K, Miyazaki J, Matoba R, Ko MS, Niwa H. Klf4 cooperates with Oct3/4 and Sox2 to activate the Lefty1 core promoter in embryonic stem cells. Mol Cell Biol. 2006; 26:7772–82. https://doi.org/10.1128/MCB.00468-06 [PubMed]
- 15. Johannesson M, Ståhlberg A, Ameri J, Sand FW, Norrman K, Semb H. FGF4 and retinoic acid direct differentiation of hESCs into PDX1-expressing foregut endoderm in a time- and concentration-dependent manner. PLoS One. 2009; 4:e4794. https://doi.org/10.1371/journal.pone.0004794 [PubMed]
- 16. Mossahebi-Mohammadi M, Quan M, Zhang JS, Li X. FGF Signaling Pathway: A Key Regulator of Stem Cell Pluripotency. Front Cell Dev Biol. 2020; 8:79. https://doi.org/10.3389/fcell.2020.00079 [PubMed]
- 17. Delfín DA, DeAguero JL, McKown EN. The Extracellular Matrix Protein ABI3BP in Cardiovascular Health and Disease. Front Cardiovasc Med. 2019; 6:23. https://doi.org/10.3389/fcvm.2019.00023 [PubMed]
- 18. Hodgkinson CP, Naidoo V, Patti KG, Gomez JA, Schmeckpeper J, Zhang Z, Davis B, Pratt RE, Mirotsou M, Dzau VJ. Abi3bp is a multifunctional autocrine/paracrine factor that regulates mesenchymal stem cell biology. Stem Cells. 2013; 31:1669–82. https://doi.org/10.1002/stem.1416 [PubMed]
- 19. Chen AC, Lee YL, Hou DY, Fong SW, Peng Q, Pang RT, Chiu PC, Ho PC, Lee KF, Yeung WS. Study of transforming growth factor alpha for the maintenance of human embryonic stem cells. Cell Tissue Res. 2012; 350:289–303. https://doi.org/10.1007/s00441-012-1476-7 [PubMed]
- 20. Barbuskaite D, Pedersen FD, Christensen HL, Johnsen LØ, Praetorius J, Damkier HH. NBCe2 (Slc4a5) Is Expressed in the Renal Connecting Tubules and Cortical Collecting Ducts and Mediates Base Extrusion. Front Physiol. 2020; 11:560. https://doi.org/10.3389/fphys.2020.00560 [PubMed]
- 21. Wang D, Lin N, Tang Y, Lu H. Inhibition of P2Y11R ameliorated TNF-α-induced degradation of extracellular matrix in human chondrocytic SW1353 cells. Am J Transl Res. 2019; 11:2108–16. [PubMed]
- 22. Jiménez-Martínez M, Ostalé CM, van der Burg LR, Galán-Martínez J, Hardwick JC, López-Pérez R, Hawinkels LJ, Stamatakis K, Fresno M. DUSP10 Is a Regulator of YAP1 Activity Promoting Cell Proliferation and Colorectal Cancer Progression. Cancers (Basel). 2019; 11:1767. https://doi.org/10.3390/cancers11111767 [PubMed]
- 23. Principe S, Mejia-Guerrero S, Ignatchenko V, Sinha A, Ignatchenko A, Shi W, Pereira K, Su S, Huang SH, O’Sullivan B, Xu W, Goldstein DP, Weinreb I, et al. Proteomic Analysis of Cancer-Associated Fibroblasts Reveals a Paracrine Role for MFAP5 in Human Oral Tongue Squamous Cell Carcinoma. J Proteome Res. 2018; 17:2045–59. https://doi.org/10.1021/acs.jproteome.7b00925 [PubMed]
- 24. Yang X, Wu K, Li S, Hu L, Han J, Zhu D, Tian X, Liu W, Tian Z, Zhong L, Yan M, Zhang C, Zhang Z. MFAP5 and TNNC1: Potential markers for predicting occult cervical lymphatic metastasis and prognosis in early stage tongue cancer. Oncotarget. 2017; 8:2525–35. https://doi.org/10.18632/oncotarget.12446 [PubMed]
- 25. Han H, Tian T, Huang G, Li D, Yang S. The lncRNA H19/miR-541-3p/Wnt/β-catenin axis plays a vital role in melatonin-mediated osteogenic differentiation of bone marrow mesenchymal stem cells. Aging (Albany NY). 2021; 13:18257–73. https://doi.org/10.18632/aging.203267 [PubMed]
- 26. Liu J, Liu S, Han L, Sheng Y, Zhang Y, Kim IM, Wan J, Yang L. LncRNA HBL1 is required for genome-wide PRC2 occupancy and function in cardiogenesis from human pluripotent stem cells. Development. 2021; 148:dev199628. https://doi.org/10.1242/dev.199628 [PubMed]