



Introduction
Rheumatoid arthritis, one of the most common inflammatory joint diseases in humans, is characterized by inflammation in synovium, destruction of cartilage and bone, generation of autoantibody, and complications of systemic organs [1]. Although RA affects 0.5–1% of the Western populations, the worldwide incidence of RA is increasing with the aging trend of the population [2]. Because of the results of reduced physical function, declined work capacity, decreased quality of life, and increased comorbid risk, RA carries heavy socioeconomic burden [3]. RA is believed to be a consequence of both genetic factors and environmental factors though main etiology has not yet been clearly clarified. In twin studies 50–65% of the risk for developing RA is ascribed to its heritability [4], indicating genetic factors have a strong effect on RA. So far more than one hundred gene loci associated with RA risk have been identified by single nucleotide polymorphisms (SNPs) [5, 6]. Apart from the human leukocyte antigen (HLA) locus, a well-known genetic risk factor for RA, numbers of other susceptibility genes and loci have been characterized [6]. Recently, a growing body of non-HLA genetic predisposition studies have been conducted on the association with the risk of RA [7–9].
Cytotoxic T lymphocyte-associated protein 4 (CTLA-4), one of widely studied non-HLA susceptibility gene of RA, is mainly expressed on the surface of Treg cells and conventional T cells and suppresses self-reactive T cell responses via downregulating ligand availability for the costimulatory receptor CD28 to elicit inhibitory signals [10, 11]. Besides, the polymorphisms of CTLA-4 have already been proved to be candidates of the risk of the common autoimmune diseases at the genetic level [12–15]. As RA is a T cell mediated autoimmune disorder and CTLA-4 plays a vital role in regulating T cell function [11, 12, 16], it suggests that CTLA-4 expression or function is most likely associated with the pathogenesis of RA. Single nucleotide polymorphisms in the CTLA-4 gene may contribute to abnormal levels of CTLA-4, and subsequently play a leading part in the susceptibility to RA [12, 17, 18].
Among the identified SNPs in this gene, these three loci of CTLA-4, +49A/G (rs231775), -318C/T (rs5742909) and CT60 G/A(rs3087243), are most-often studied for the association with the predisposition of RA [18–20]. However, the conclusions which previous reports drew are inconsistent and incomprehensive. Although the association of CTLA-4 genetic polymorphisms and the risk of RA has been assessed in several meta-analyses [21–23], some recent studies also described this association in different populations in the past several years [9, 15, 24–27]. Hence these studies should be included to increase statistical power and obtain the reliable conclusion. On the other hand, all the three common loci should be included to embody the association comprehensively while the previous meta-analysis only researched one or two of the above loci. In view of these, it is necessary to incorporate the latest research into investigating the association of the three polymorphisms of CTLA-4 with susceptibility to RA. Here we use the latest case-control data to carry out an updated and comprehensive meta-analysis and obtain a more accurate estimation of the effect of the 3 SNPs (+49A/G (rs231775), CT60 G/A(rs3087243) and -318 C/T (rs5742909)) on RA risk.
Results
Characteristics of the studies
Based on the predetermined inclusion criteria, 66 eligible case–control studies with 42 articles were enrolled ultimately in the current analysis [8, 9, 13–15, 17–20, 24–56]. These publications had a high methodological quality whose NOS stars were more than 6 in general. There were 22 studies with 16394 patients and 17453 controls for rs3087243 SNP [8, 9, 13–15, 18, 19, 26, 40, 41, 43, 46–49, 52, 53, 56], 34 studies with 11452 patients and 12444 controls for rs231775 SNP [9, 14, 17, 19, 20, 24, 25, 28–39, 41–45, 49–51, 54], and 10 studies with 2477 patients and 2941 controls for rs5742909 SNP [14, 20, 27, 29, 34, 37–39, 44, 56]. The references of all enrolled articles were subject to scrutiny and no more ones were available. The process of study selection according to the PRISMA principle was generalized in Figure 1. Quality assessment of included studies was shown in Supplementary Table 1. Details of included studies were listed in Table 1. Allele/genotype frequencies were displayed in Table 2.
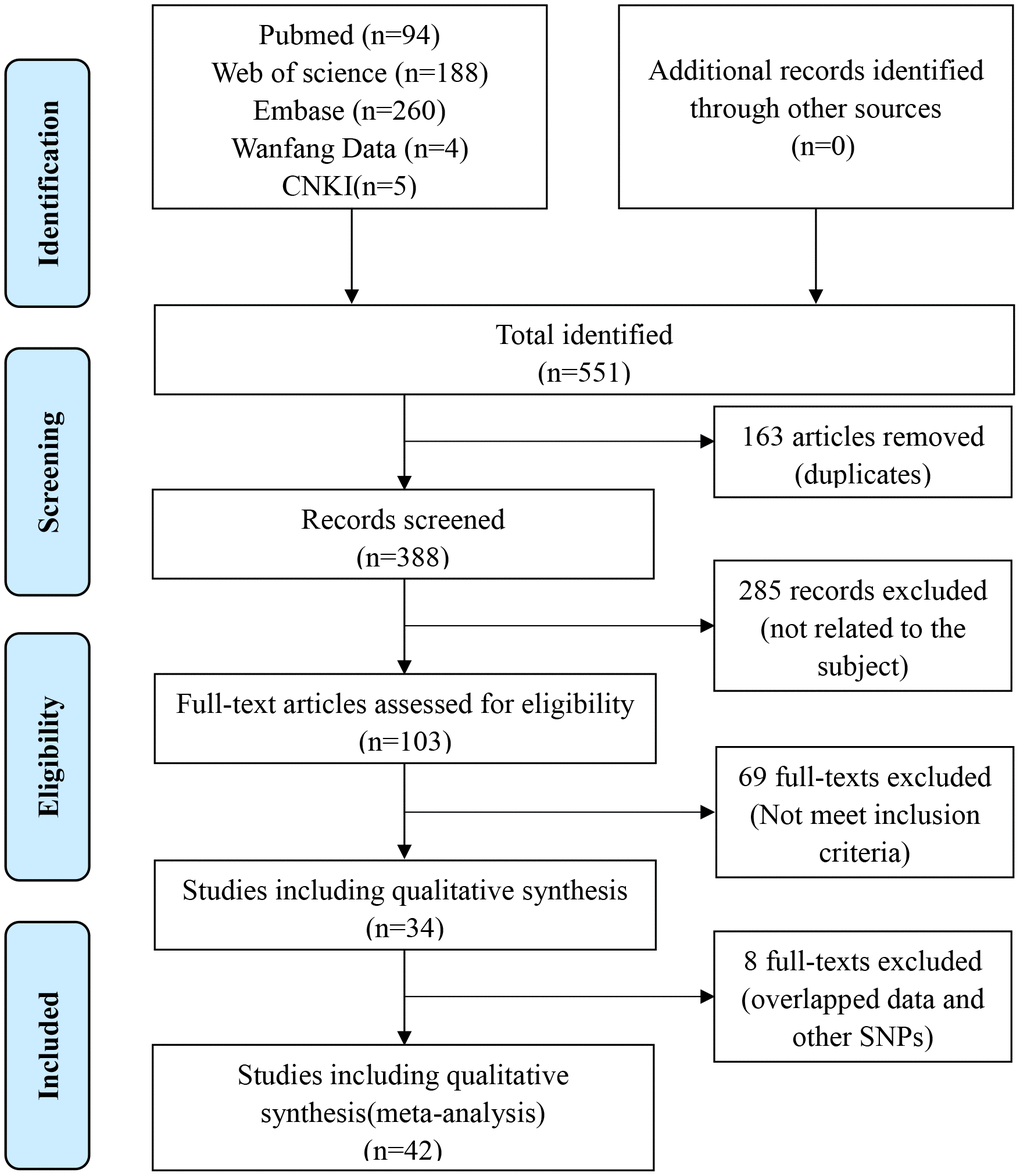
Figure 1. Flow diagram of the literature retrieval and screen.
Table 1. Main characteristics of included studies.
| Study | Year | Country | Ethnicity | Numbers | Genotype method | Diagnostic criteria | Quality score | |
| RA | Con | |||||||
| Rs3087243(CT60) | ||||||||
| Orozco | 2004 | Spain | Caucasian | 433 | 398 | TaqMan | ACR1987 | 7 |
| Lei | 2005 | China | Asian | 326 | 250 | DGGE | ACR1987 | 8 |
| Plenge (EIRA) | 2005 | Sweden | European | 1505 | 878 | MALDI-TOF | ACR1987 | 8 |
| Plenge (NARAC) | 2005 | Sweden | European | 828 | 845 | MALDI-TOF | ACR1987 | 8 |
| Zhernakova | 2005 | Dutch | Caucasian | 153 | 900 | PCR-RFLP | ACR1987 | 6 |
| Suppiah | 2006 | Northern Ireland | Caucasian | 289 | 168 | PCR-RFLP | ACR1987 | 7 |
| Costenbader | 2008 | USA | Caucasian | 423 | 420 | TaqMan | ACR1987 | 7 |
| Tsukahara | 2008 | Japan | Asian | 1498 | 441 | TaqMan | ACR1987 | 8 |
| Kelley | 2009 | USA | African | 505 | 712 | TaqMan | ACR1987 | 7 |
| Daha | 2009 | Dutch | Caucasian | 867 | 863 | Sequenom | ACR1987 | 7 |
| Barton | 2009 | UK | European | 3669 | 3049 | TaqMan | ACR1987 | 8 |
| Walker | 2009 | Canada | Caucasian | 1140 | 1248 | Sequenom | ACR1987 | 8 |
| Plant (1) | 2010 | France | Caucasian | 671 | 177 | Sequenom | ACR1987 | 8 |
| Plant (2) | 2010 | Germany | Caucasian | 218 | 209 | Sequenom | ACR1987 | 8 |
| Plant (3) | 2010 | Greece | Caucasian | 268 | 290 | Sequenom | ACR1987 | 8 |
| Plant (4) | 2010 | UK | Caucasian | 1002 | 2725 | Sequenom | ACR1987 | 8 |
| Danoy | 2011 | China | Asian | 1035 | 1702 | Sequenom | ACR1987 | 7 |
| Torres-Carrillo | 2013 | Mexico | Latin American | 200 | 200 | PCR–RFLP | ACR1987 | 8 |
| Luterek-Puszyńska | 2016 | Poland | Caucasian | 422 | 338 | TaqMan | ACR1987 | 7 |
| Schulz | 2020 | Germany | Caucasian | 111 | 256 | PCR–RFLP | ACR2010 | 6 |
| El-Gabalawy | 2011 | Canada | Caucasian | 332 | 490 | Sequenom | ACR1987 | 6 |
| Vernerova | 2016 | Slovakia | Caucasian | 499 | 894 | TaqMan | ACR2010 | 9 |
| Rs231775(49G/A) | ||||||||
| AIFadhli | 2013 | Kuwait | Asian | 114 | 282 | PCR–RFLP | ACR1987 | 6 |
| Barton (I) | 2000 | Spain | Caucasian | 136 | 144 | PCR–RFLP | ACR1987 | 7 |
| Barton (II) | 2000 | UK | Caucasian | 192 | 96 | PCR–RFLP | ACR1987 | 7 |
| Benhatchi | 2011 | Slovakia | Caucasian | 57 | 51 | PCR–RFLP | ACR1987 | 6 |
| Elshazli | 2015 | Egypt | Caucasian | 112 | 122 | PCR–RFLP | ACR1987 | 6 |
| Feng | 2005 | China | Asian | 50 | 60 | PCR–RFLP | ACR1987 | 6 |
| Gonzalez-Escribano | 1999 | Spain | Caucasian | 138 | 305 | PCR-ARMS | ACR1987 | 6 |
| Hadj | 2001 | Tunisia | African | 60 | 150 | PCR–RFLP | ACR1987 | 7 |
| Lee 2002 | 2002 | Korea | Asian | 86 | 86 | PCR–RFLP | ACR1987 | 6 |
| Lee 2003 | 2003 | China | Asian | 186 | 203 | PCR–RFLP | ACR1987 | 6 |
| Lei | 2005 | China | Asian | 326 | 250 | DGGE | ACR1987 | 8 |
| Liu 2004 | 2004 | Taiwan | Asian | 65 | 81 | PCR–RFLP | ACR1987 | 6 |
| Barton | 2004 | UK | European | 132 | 156 | TaqMan | ACR1987 | 7 |
| Liu 2013 | 2013 | China | Asian | 213 | 303 | PCR–RFLP | ACR1987 | 7 |
| Luterek-Puszyńska | 2016 | Poland | Caucasian | 422 | 338 | TaqMan | ACR2010 | 7 |
| Matsushita | 1999 | Japan | Asian | 461 | 150 | PCR-SSCP | ACR1987 | 7 |
| Milicic | 2001 | UK | Caucasian | 421 | 452 | PCR–RFLP | ACR1987 | 8 |
| Miterski | 2004 | Germany | Caucasian | 284 | 362 | PCR–RFLP | ACR1987 | 7 |
| Munoz-Valle | 2010 | Mexico | Mexican | 199 | 199 | PCR–RFLP | ACR1987 | 6 |
| Plant (1) | 2010 | France | Caucasian | 684 | 162 | Sequenom | ACR1987 | 8 |
| Plant (2) | 2010 | Germany | European | 220 | 209 | Sequenom | ACR1987 | 8 |
| Plant (3) | 2010 | Greece | European | 272 | 287 | Sequenom | ACR1987 | 8 |
| Plant (4) | 2010 | UK | European | 1004 | 2659 | Sequenom | ACR1987 | 6 |
| Seidl | 1998 | Germany | Caucasian | 258 | 456 | RFLP-SSCP | ACR1987 | 8 |
| Suppiah | 2006 | UK | European | 289 | 475 | PCR–RFLP | ACR1987 | 7 |
| Takeuchi | 2006 | Japan | Asian | 100 | 104 | PCR–RFLP | ACR1987 | 6 |
| Tang | 2013 | China | Asian | 1489 | 1200 | TaqMan | ACR1987 | 8 |
| Tsukahara | 2008 | Japan | Asian | 1490 | 448 | TaqMan | ACR1987 | 8 |
| Kelley | 2009 | USA | African | 505 | 712 | TaqMan | ACR1987 | 7 |
| Vaidya | 2002 | UK | Caucasian | 123 | 349 | PCR–RFLP | ACR1987 | 6 |
| Walker | 2009 | Canada | Caucasian | 1140 | 1248 | Sequenom | ACR1987 | 8 |
| Yanagawa | 2000 | Japan | Asian | 85 | 200 | PCR–RFLP | ACR1987 | 6 |
| Zhou | 2007 | China | Asian | 39 | 44 | PCR–RFLP | ACR1987 | 6 |
| Sameem | 2015 | Pakistani | Asian | 100 | 100 | PCR–RFLP | RF test | 6 |
| Rs5742909 (318C/T) | ||||||||
| Gonzalez-Escribano | 1999 | Spain | Caucasian | 138 | 305 | PCR-ARMS | ACR1987 | 6 |
| Lee 2002 | 2002 | Korea | Asian | 86 | 86 | PCR–RFLP | ACR1987 | 6 |
| Barton | 2004 | UK | European | 151 | 152 | TaqMan | ACR1987 | 7 |
| Liu 2004 | 2004 | Tainan | Asian | 65 | 81 | PCR–RFLP | ACR1987 | 6 |
| Miterski | 2004 | Germany | Caucasian | 284 | 362 | PCR–RFLP | ACR1987 | 7 |
| Takeuchi | 2006 | Japan | Asian | 100 | 104 | PCR–RFLP | ACR1987 | 6 |
| Walker | 2009 | Canada | Caucasian | 1140 | 1248 | Sequenom | ACR1987 | 8 |
| Liu 2013 | 2013 | China | Asian | 213 | 303 | PCR–RFLP | ACR1987 | 7 |
| Torres-Carrillo | 2013 | Mexico | Latin American | 200 | 200 | PCR–RFLP | ACR1987 | 7 |
| Fattah | 2017 | Egypt | Caucasian | 100 | 100 | PCR–RFLP | ACR2010 | 6 |
Table 2. Distribution of genotype and allele among RA patients and controls.
| Study | Cases | Controls | HEW | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| MM | Mm | mm | M | m | MM | Mm | mm | M | m | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs3087243(CT60) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Orozco | 118 | 198 | 117 | 434 | 432 | 98 | 199 | 101 | 395 | 401 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lei | 33 | 137 | 156 | 203 | 449 | 32 | 131 | 87 | 195 | 305 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plenge (EIRA) | 230 | 680 | 595 | 1140 | 1870 | 145 | 396 | 337 | 686 | 1070 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plenge (NARAC) | 133 | 387 | 308 | 653 | 1003 | 165 | 426 | 254 | 756 | 934 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Zhernakova | NA | NA | NA | 133 | 173 | NA | NA | NA | 841 | 959 | NA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Suppiah | NA | NA | NA | 234 | 344 | NA | NA | NA | 145 | 191 | NA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Costenbader | 82 | 201 | 140 | 365 | 481 | 87 | 195 | 138 | 369 | 471 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tsukahara | 87 | 538 | 873 | 712 | 2284 | 33 | 163 | 245 | 229 | 653 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Kelley | NA | NA | NA | NA | 505 | NA | NA | NA | NA | 712 | NA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Daha | NA | NA | NA | 729 | 1005 | NA | NA | NA | 785 | 941 | NA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Barton | 677 | 1760 | 1232 | 3114 | 4224 | 634 | 1523 | 892 | 2791 | 3307 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Walker | 207 | 518 | 415 | 932 | 1348 | 273 | 613 | 362 | 1159 | 1337 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plant (1) | 131 | 332 | 208 | 594 | 748 | 45 | 91 | 41 | 181 | 173 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plant (2) | 35 | 105 | 78 | 175 | 261 | 35 | 101 | 73 | 171 | 247 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plant (3) | 55 | 135 | 78 | 245 | 291 | 70 | 145 | 75 | 285 | 295 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plant (4) | 204 | 487 | 311 | 895 | 1109 | 542 | 1344 | 839 | 2428 | 3022 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Danoy | NA | NA | NA | 310 | 1760 | NA | NA | NA | 681 | 2723 | NA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Torres-Carrillo | 31 | 86 | 83 | 148 | 252 | 32 | 106 | 62 | 170 | 230 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Luterek-Puszyńska | 53 | 193 | 176 | 299 | 545 | 45 | 174 | 119 | 264 | 412 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Schulz | 13 | 49 | 49 | 75 | 147 | 42 | 124 | 90 | 208 | 304 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| El-Gabalawy | 45 | 161 | 126 | 251 | 413 | 66 | 226 | 198 | 358 | 622 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Vernerova | NA | NA | NA | 616 | 382 | NA | NA | NA | 1064 | 1064 | NA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs231775(49G/A) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AIFadhli | 10 | 30 | 74 | 50 | 178 | 14 | 86 | 182 | 114 | 450 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Barton (I) | 14 | 57 | 65 | 85 | 187 | 12 | 70 | 62 | 94 | 194 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Barton (II) | 38 | 86 | 68 | 162 | 222 | 19 | 51 | 26 | 89 | 103 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Benhatchi | 6 | 33 | 18 | 45 | 69 | 5 | 25 | 21 | 35 | 67 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Elshazli | 14 | 55 | 43 | 83 | 141 | 6 | 45 | 71 | 57 | 187 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Feng | 20 | 21 | 9 | 61 | 39 | 9 | 32 | 19 | 50 | 70 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gonzalez-Escribano | 10 | 63 | 65 | 83 | 193 | 30 | 103 | 172 | 163 | 447 | NO | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Hadj | 23 | 27 | 10 | 73 | 47 | 68 | 62 | 20 | 198 | 102 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lee 2002 | 41 | 35 | 10 | 117 | 55 | 49 | 29 | 8 | 127 | 45 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lee 2003 | 103 | 67 | 16 | 273 | 99 | 85 | 100 | 18 | 270 | 136 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lei | 148 | 138 | 40 | 434 | 218 | 86 | 125 | 39 | 297 | 203 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liu 2004 | 14 | 42 | 9 | 70 | 60 | 21 | 50 | 10 | 92 | 70 | NO | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Barton | 34 | 55 | 43 | 123 | 141 | 29 | 68 | 59 | 126 | 186 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liu 2013 | 77 | 111 | 25 | 265 | 161 | 130 | 125 | 48 | 385 | 221 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Luterek-Puszyńska | 79 | 210 | 133 | 368 | 476 | 63 | 160 | 115 | 286 | 390 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Matsushita | 200 | 199 | 62 | 599 | 323 | 56 | 72 | 22 | 184 | 116 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Milicic | 63 | 223 | 135 | 349 | 493 | 73 | 213 | 166 | 359 | 545 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Miterski | NA | NA | NA | 222 | 346 | NA | NA | NA | 269 | 455 | NA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Munoz-Valle | 42 | 102 | 55 | 186 | 212 | 34 | 82 | 83 | 150 | 248 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plant (1) | 96 | 315 | 273 | 507 | 861 | 15 | 75 | 72 | 105 | 219 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plant (2) | 37 | 111 | 72 | 185 | 255 | 32 | 94 | 83 | 158 | 260 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plant (3) | 26 | 133 | 113 | 185 | 359 | 33 | 107 | 147 | 173 | 401 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Plant (4) | 146 | 451 | 407 | 743 | 1265 | 410 | 1255 | 994 | 2075 | 3243 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Seidl | 37 | 138 | 83 | 212 | 304 | 68 | 210 | 179 | 346 | 568 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Suppiah | 40 | 144 | 105 | 224 | 354 | 92 | 241 | 142 | 425 | 525 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Takeuchi | 49 | 39 | 12 | 137 | 63 | 44 | 49 | 11 | 137 | 71 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tang | 652 | 642 | 195 | 1946 | 1032 | 474 | 535 | 191 | 1483 | 917 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tsukahara | 636 | 668 | 186 | 1940 | 1040 | 181 | 194 | 73 | 556 | 340 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Kelley | NA | NA | NA | NA | 505 | NA | NA | NA | NA | 712 | NA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Vaidya | 20 | 65 | 38 | 105 | 141 | 45 | 158 | 146 | 248 | 450 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Walker | 177 | 554 | 409 | 908 | 1372 | 178 | 577 | 493 | 933 | 1563 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Yanagawa | 29 | 50 | 6 | 108 | 62 | 78 | 88 | 34 | 244 | 156 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Zhou | 22 | 9 | 8 | 53 | 25 | 8 | 14 | 22 | 30 | 58 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Sameem | 54 | 26 | 20 | 134 | 66 | 28 | 31 | 41 | 87 | 113 | NO | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs5742909 (318C/T) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Gonzalez-Escribano | 1 | 29 | 108 | 31 | 245 | 2 | 60 | 243 | 64 | 546 | NO | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lee 2002 | 2 | 19 | 65 | 23 | 149 | 4 | 14 | 68 | 22 | 150 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Barton | 1 | 18 | 132 | 20 | 282 | 3 | 27 | 122 | 33 | 271 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liu 2004 | 0 | 15 | 50 | 15 | 115 | 0 | 23 | 58 | 23 | 139 | NO | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Miterski | NA | NA | NA | 64 | 504 | NA | NA | NA | 50 | 674 | NA | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Takeuchi | 0 | 13 | 87 | 13 | 187 | 0 | 22 | 82 | 22 | 186 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Walker | 13 | 219 | 908 | 245 | 2035 | 10 | 183 | 1055 | 203 | 2293 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Liu 2013 | 14 | 97 | 102 | 125 | 301 | 13 | 77 | 213 | 103 | 503 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Torres-Carrillo | 2 | 16 | 182 | 20 | 380 | 0 | 20 | 180 | 20 | 380 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Fattah | 7 | 52 | 41 | 66 | 134 | 2 | 32 | 66 | 36 | 164 | YES | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| M, minor allele; m, major allele; NA, not available; HWE, Hardy-Weinberg Equilibrium. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Efficiency analysis
Meta-analysis of CTLA-4 CT60(rs3087243) SNP and RA susceptibility
By analyzing quantitatively allele or genotype distribution of 16394 patients and 17453 controls, a significant association between RA and CTLA-4 CT60(rs3087243) SNP was observed in all genetic comparisons (A vs. G: OR = 0.87, 95% CI = 0.83-0.91, P<0.00001; AA vs. GG: OR = 0.80, 95% CI =0.74-0.87, P<0.00001; AG vs. AA: OR = 0.85, 95% CI =0.80-0.90, P<0.0001; AA + AG vs. GG: OR =0.83, 95% CI=0.77-0.90, P<0.0001, and AA vs. AG+ GG: OR =0.88, 95% CI=0.83-0.94, P=0.0003) (Table 3 and Figure 2). Among the 22 included studies, 17 studies were performed in Caucasians, 3 were in Asians, 1 was African and 1 was in Latin Americans. Likewise, we carried out a stratified analysis by race to evaluate the ethnicity effects. In Caucasians, a protective role of rs3087243 SNP on RA was detected in all the five genetic comparisons. Similarly, a decreased risk of RA was found among Asians in the allelic comparison (OR = 0.77, 95% CI =0.65-0.90, P=0.001) and the homozygote comparison (OR = 0.67, 95% CI = 0.48-0.94, P=0.02). The heterozygote model and dominant model detected also this correlation in Latin Americans and the allelic comparison detected this correlation in Africans, but both the two populations needed more enrolled studies to elevate statistical power because this analysis currently included individually only one study. The outcomes were shown in Table 3. Collectively, Subgroup analyses revealed a significant protective association in Caucasians and Asians. When the I2 > 50% and P>0.1, the Fix-effect model was used for the synthesis; otherwise, the Random-effect model was used.
Table 3. Results of different comparative genetic models on the association of CTLA-4 SNPs with RA.
| Genetic model | Population | Cases | Controls | Association | Heterogeneity | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OR | 95%CI | P-value | Model | I2 | P-value | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs308724 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A vs. G | Total | 16394 | 17453 | 0.87 | 0.83-0.91 | <0.00001 | REM | 39 | 0.003 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 12830 | 14148 | 0.89 | 0.86-0.93 | <0.00001 | FEM | 25 | 0.17 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 2859 | 2393 | 0.77 | 0.65-0.90 | 0.001 | REM | 56 | 0.10 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 0.79 | 0.60-1.06 | 0.11 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| African | 505 | 712 | 0.83 | 0.67-1.02 | 0.08 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AA vs. GG | Total | 13046 | 12214 | 0.80 | 0.74-0.87 | <0.00001 | FEM | 22 | 0.20 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 11022 | 11323 | 0.81 | 0.75-0.88 | <0.00001 | FEM | 32 | 0.13 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 1824 | 691 | 0.67 | 0.48-0.94 | 0.02 | FEM | 0 | 0.48 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 0.72 | 0.40-1.31 | 0.29 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AG vs. GG | Total | 13046 | 12214 | 0.85 | 0.80-0.90 | <0.0001 | FEM | 28 | 0.14 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 11022 | 11323 | 0.86 | 0.81-0.92 | <0.0001 | FEM | 11 | 0.33 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 1824 | 691 | 0.75 | 0.48-1.18 | 0.21 | REM | 78 | 0.03 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 0.61 | 0.39-0.94 | 0.02 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AA+GA vs. GG | Total | 13046 | 12214 | 0.83 | 0.77-0.90 | <0.0001 | REM | 46 | 0.02 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 11022 | 11323 | 0.85 | 0.78-0.93 | <0.0002 | REM | 40 | 0.07 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 1824 | 691 | 0.74 | 0.48-1.12 | 0.15 | REM | 77 | 0.04 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 0.60 | 0.40-0.90 | 0.01 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| AA vs. GA+GG | Total | 13046 | 12214 | 0.88 | 0.83-0.94 | 0.0003 | FEM | 0 | 0.75 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 11022 | 11323 | 0.89 | 0.83-0.95 | 0.0008 | FEM | 0 | 0.60 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 1824 | 691 | 0.76 | 0.55-1.06 | 0.10 | FEM | 0 | 0.98 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 0.96 | 0.56-1.65 | 0.89 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs231775 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| G vs. A | Total | 11452 | 12444 | 1.16 | 1.08-1.25 | <0.0001 | REM | 66 | 0.00001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 5884 | 7872 | 1.09 | 1.01-1.19 | 0.04 | REM | 38 | 0.004 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 4804 | 3511 | 1.27 | 1.10-1.47 | 0.001 | REM | 71 | <0.0001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| African | 565 | 862 | 1.06 | 0.68-1.65 | 0.81 | REM | 73 | 0.05 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 199 | 199 | 1.45 | 1.09-1.92 | 0.010 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GG vs. AA | Total | 10663 | 11370 | 1.29 | 1.12-1.50 | 0.0006 | REM | 54 | 0.0002 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 5600 | 7510 | 1.11 | 0.94-1.31 | 0.21 | FEM | 25 | 0.17 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 4804 | 3511 | 1.58 | 1.24-2.01 | 0.0002 | REM | 51 | 0.01 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| African | 60 | 150 | 0.68 | 0.28-1.65 | 0.39 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 199 | 199 | 1.24 | 1.09-1.42 | 0.03 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GA vs. AA | Total | 10663 | 11370 | 1.19 | 1.07-1.32 | 0.001 | REM | 46 | 0.003 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 5600 | 7510 | 1.18 | 1.02-1.35 | 0.02 | REM | 59 | 0.001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 4804 | 3511 | 1.20 | 1.05-1.38 | 0.08 | FEM | 3 | 0.42 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| African | 60 | 150 | 0.87 | 0.36-2.11 | 0.76 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 199 | 199 | 1.88 | 1.20-2.94 | 0.006 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GG+GA vs. AA | Total | 10663 | 11370 | 1.24 | 1.11-1.39 | 0.0001 | FEM | 56 | 0.001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 5600 | 7510 | 1.17 | 1.02-1.34 | 0.02 | REM | 62 | 0.0006 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 4804 | 3511 | 1.33 | 1.17-1.51 | <0.0001 | FEM | 31 | 0.12 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| African | 60 | 150 | 0.77 | 0.34-1.76 | 0.53 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 199 | 199 | 1.87 | 1.23-2.85 | 0.003 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| GG vs. GA+AA | Total | 10663 | 11370 | 1.15 | 1.02-1.30 | 0.02 | REM | 57 | <0.0001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 5600 | 7510 | 1.01 | 0.91-1.12 | 0.80 | FEM | 10 | 0.34 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 4804 | 3511 | 1.34 | 1.08-1.65 | 0.008 | REM | 72 | <0.0001 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| African | 60 | 150 | 0.75 | 0.41-1.38 | 0.36 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 199 | 199 | 1.30 | 0.79-2.15 | 0.31 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Rs5742909 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T vs. C | Total | 2477 | 2941 | 1.21 | 0.93-1.57 | 0.15 | REM | 71 | 0.0003 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 1813 | 2167 | 1.31 | 0.94-1.84 | 0.11 | REM | 73 | 0.005 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 464 | 574 | 1.05 | 0.56-1.96 | 0.88 | REM | 80 | 0.002 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 1.00 | 0.53-1.89 | 1.00 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TT vs. CC | Total | 2193 | 2579 | 1.71 | 1.08-2.73 | 0.08 | FEM | 17 | 0.30 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 1529 | 1805 | 1.58 | 0.60-4.17 | 0.35 | REM | 32 | 0.22 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 464 | 574 | 1.34 | 0.34-5.28 | 0.68 | REM | 56 | 0.13 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 4.95 | 0.24-103.73 | 0.30 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TC vs. CC | Total | 2193 | 2579 | 1.19 | 0.84-1.69 | 0.33 | FEM | 76 | <0.0001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 1529 | 1805 | 1.27 | 0.81-1.99 | 0.29 | REM | 74 | 0.01 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 464 | 574 | 1.16 | 0.53-2.56 | 0.70 | REM | 83 | 0.0004 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 0.79 | 0.40-1.58 | 0.51 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TT+TC vs. CC | Total | 2193 | 2579 | 1.19 | 0.84-1.69 | 0.33 | FEM | 77 | <0.0001 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 1529 | 1805 | 1.28 | 0.79-2.07 | 0.32 | REM | 78 | 0.003 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 464 | 574 | 1.12 | 0.52-2.43 | 0.77 | REM | 84 | 0.0003 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 0.89 | 0.46-1.74 | 0.73 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| TT vs. TC+CC | Total | 2193 | 2579 | 1.43 | 0.90-2.27 | 0.13 | FEM | 0 | 0.52 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Caucasian | 1529 | 1805 | 1.46 | 0.77-2.78 | 0.25 | FEM | 0 | 0.39 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Asian | 464 | 574 | 1.27 | 0.63-2.54 | 0.51 | FEM | 32 | 0.23 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Latin | 200 | 200 | 5.05 | 0.24-105.86 | 0.30 | ¯ | ¯ | ¯ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| OR, odds ratio; CI, confidence interval; FEM, fix-effect model; REM, random-effect model. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
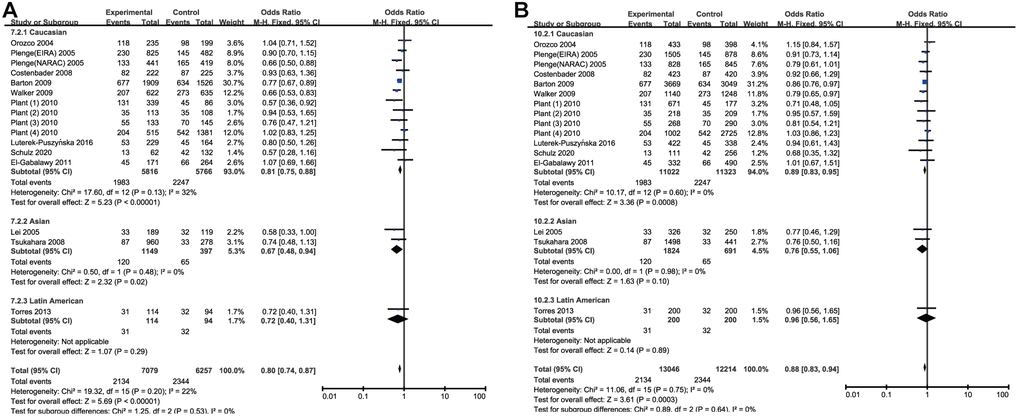
Figure 2. Forest plot of the association between rs308724 polymorphism and RA risk under the homozygous (A) and recessive model (B).
Meta-analysis of CTLA-4 +49A/G (rs231775) SNP and RA susceptibility
By quantitative analysis of allele or genotype distribution of 11452 patients and 12444 controls, there was a significant risk association between RA and CTLA-4 +49A/G (rs231775) SNP. The overall pooled ORs of all the populations were as follows: G vs. A: OR =1.16, 95% CI =1.08-1.25, P<0.0001; GG vs. AA: OR =1.29, 95% CI =1.12-1.50, P=0.0006; GA vs. AA: OR =1.19, 95% CI =1.07-1.32, P=0.001; GG + GA vs. AA: OR =1.24, 95% CI=1.11-1.39, P=0.0001 and GG vs. GA+AA: OR =1.15, 95% CI=1.02-1.30, P=0.02. The main results of overall analyses were shown in Table 3. 17 studies were conducted on Caucasians, 14 on Asians, 2 on Africans and 1 on Latin Americans. Subsequently, stratified analysis by ethnicity was conducted to get more clarifications. In the subgroup analysis, a significantly increased risk of RA was observed among the Asian population in all genetic comparisons except heterozygote comparison (G vs. A: OR =1.27, 95% CI =1.10-1.47, P=0.001; GG vs. AA: OR =1.58, 95% CI =1.24-2.01, P=0.0002; GG + GA vs. AA: OR =1.33, 95% CI=1.17-1.51, P<0.0001; GG vs. GA+AA: OR = 1.15, 95% CI =1.02-1.30, P=0.02). In Latin American population, rs231775 SNP was a significant risk factor of RA, but it only included single study and the result might be incredible. Besides, no association of the rs231775 SNP with RA risk was found among the Caucasian population in all genetic comparisons when the Elshazli’s study [24] was excluded because of its heterogeneity (G vs. A: OR =1.07, 95%CI =0.99-1.15, P =0.08; GG vs. AA: OR = 1.07, 95% CI = 0.92–1.23, P=0.37; GA vs. AA: OR = 1.15, 95% CI =1.00-1.31, P=0.05; GG + GA vs. AA: OR =1.14, 95% CI=1.00-1.29, P=0.05 and GG vs. GA+AA: OR =1.00, 95% CI=0.90–1.11, P=0.98) (Table 3 and Figure 3). There was no remarkable association between rs231775 SNP and RA in Africans. The results were summarized in Table 3 and Figure 3. These data with moderate heterogeneity employed the random-effect model for the synthesis.
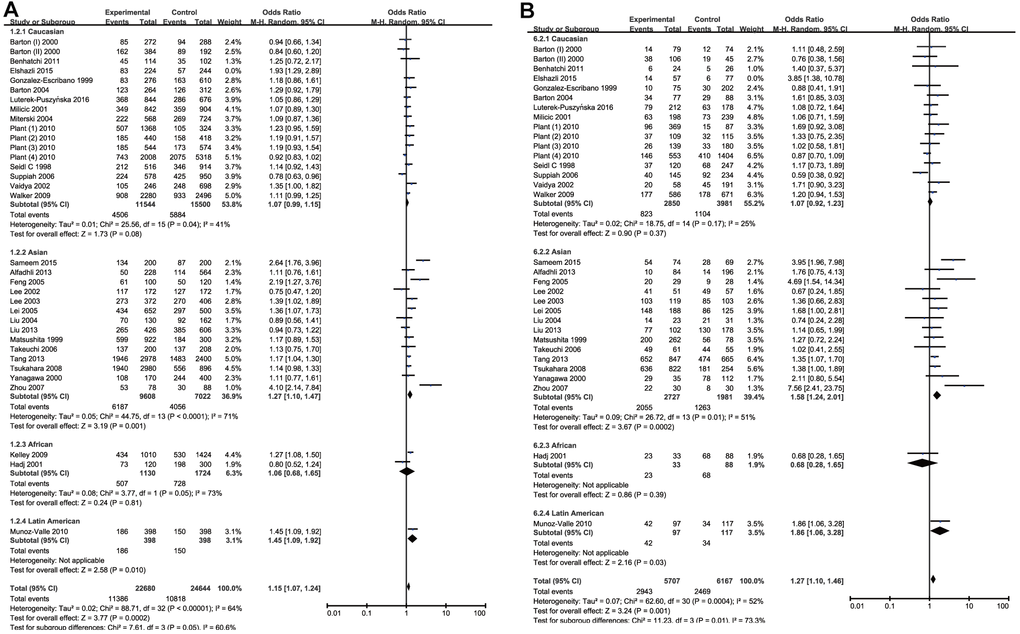
Figure 3. Forest plot of the association between rs231775 polymorphism and RA risk under the allelic model with Elshazli R et al.’s study excluded (A) and homozygous model (B).
Meta-analysis of CTLA-4 318C/T (rs5742909) SNP and RA susceptibility
Through the pooled analysis of genetic data of 2477 patients and 2941 controls in a total of 10 studies, of which 5 were conduct on Caucasians, 4 on Asians, and 1 on Latin Americans, no significant associations between rs5742909 SNP and RA in the overall pooled results were found among all populations for the allelic and genotypic comparisons (T vs. C: OR =1.21, 95% CI =0.93-1.57, P=0.15; TT vs. CC: OR =1.71, 95% CI =1.08-2.73, P=0.08; TC vs. CC: OR =1.19, 95% CI =0.84-1.69, P=0.33; TT+TC vs. CC: OR =1.19, 95% CI=0.84-1.69, P=0.33 and TT vs. TC+CC: OR =1.43, 95% CI=0.90-2.27, P=0.13) (Table 3 and Figure 4). Meanwhile, the subgroup analysis by ethnicity did not indicate any remarkable associations in all genetic models (Table 3). As the heterogeneity of genetic model existed, random effect model in this part was used to make a reliable result.
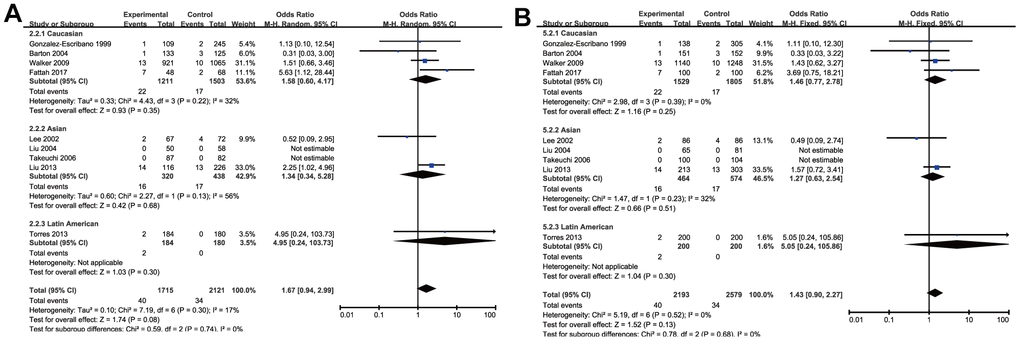
Figure 4. Forest plot of the association between rs574299 polymorphism and RA risk under the homozygous (A) and recessive model (B).
Heterogeneity analysis and publication bias
To ensure the reliability of the results, we first evaluate the heterogeneity (by I2) and found that heterogeneity existed in some genetic models of rs231775 SNP and rs5742909 SNP (Table 3). In order to minimize heterogeneity, the following methods were carried out in this meta-analysis. On the one hand, the random-effect models were exploited in the genetic models with moderate heterogeneity(I2>50%). On the other hand, sensitivity analysis was adopted to evaluate the effect of a single study on the pooled ORs by removing each study in turn from the pooled analysis. Although the heterogeneity had not changed obviously, the P values for pooled ORs under allelic comparison, heterozygous comparison and dominant comparison were reversed when the study [24] led by Elshazli R was removed. Therefore, we deleted this study and recalculated the relevant ORs and 95%CIs to harvest a stable and credible outcome (Figure 3). The funnel plots were used to investigate publication bias and the outlines of the funnel plots appear to be symmetrical (Figure 5). For rs231775 SNP, the asymmetry of the funnel plot was attributed to Zhou et al.’s study [45] which was published in Chinese. HWE estimation indicated that allele or genotype frequencies were deviant from HWE in control group in the Liu et al.’s, Gonzalez-Escribano et al.’s and Sameem et al.’s studies [25, 29, 38], but the results of synthesis analysis were not substantially inversed. Hence, we didn't remove these studies from the meta-analysis.
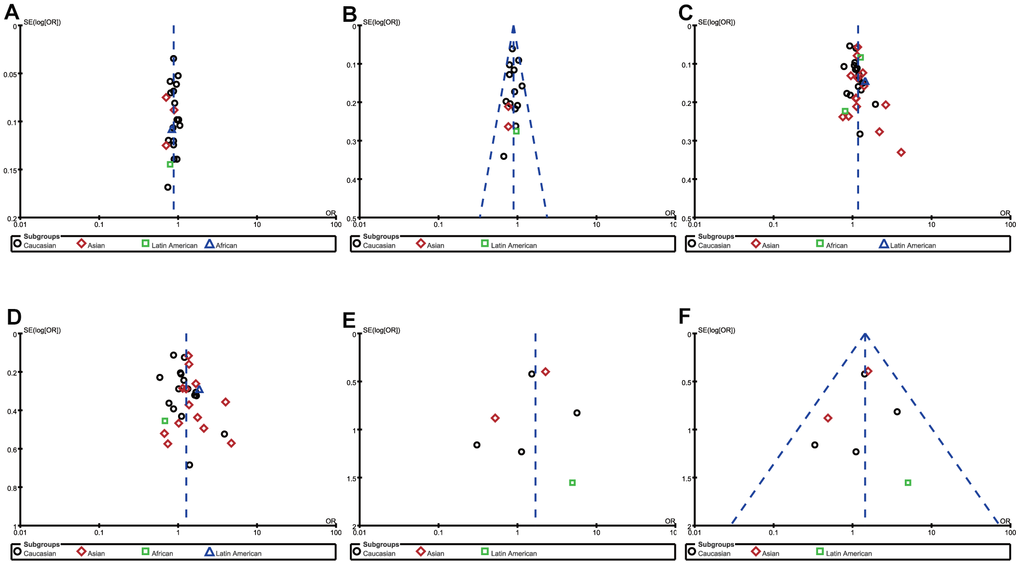
Figure 5. Funnel plot of the association between RA risk and rs308724 polymorphism under the allelic (A) and recessive model (B), rs231775 polymorphism under the allelic (C) and homozygous model (D), and rs574299 polymorphism under the homozygous (E) and recessive model (F).
Discussion
To our knowledge, this was the first meta-analysis to investigate the association between the three most-often SNPs of CTLA-4 and RA susceptibility. From the data integration of 66 studies in 21681 cases and 23457 controls, we found that the rs3087243 SNP decreased the risk of RA risk in Caucasians and Asians, the rs231775 SNP of CTLA-4 increased RA risk in Asians but not in Caucasians and Africans, and the rs5742909 SNP was not significantly associated with RA risk in both Caucasians and Africans.
The CTLA-4 gene, located on chromosome 2q33, encodes a 223 amino acid receptor protein on T cell surface which is responsible for T cell immune regulation. As an antagonist of the costimulatory receptor CD28 which binds the same ligand B7 as CTLA-4, CTLA-4 with higher affinity transmits an inhibitory signal and subsequently plays a suppressive role in regulating T-cell activation [57], which suggests it is involved in the pathological processes of many autoimmune disorders [12–15]. It is widely believed that RA is a T cell-mediated autoimmune disease [58], of which the chronic inflammation and damage of the joints are typical [1]. Although a great many genes whose protein products are critical to T cell function don’t have genetic associations with RA, the effect of CTLA-4 on RA pathogenesis has attracted growing attentions.
Previous research had found that serum levels of soluble CTLA-4 were increased in RA patients and had a positive correlation with Disease Activity Score in RA patients and even proposed that serum levels of CTLA-4 could serve as a new marker of RA disease activity [59, 60]. Besides, function experiments in vivo indicated that gene delivery of CTLA4 by intra-articular injection could alleviate experimental arthritis [61]. Furthermore, CTLA-4Ig administration on RA synovial macrophages and T helper cells downregulated the production of proinflammatory cytokines, and these evidences suggested that CTLA-4 could be a treatment target for RA [62, 63]. In fact, blockade of CTLA-4 by CTLA-4Ig had been successfully applied to treatment for RA [64].
As we all know, the protein level, structure and function are determined in large part by gene. Apart from these function research, numerous studies on correlation between CTLA-4 and RA risk from gene level also had been conducted to investigate genetic factors [8, 9, 13–15, 17–20, 24–56]. However, the results were inconsistent or contrary likely due to the various ethnic background, disparate geographic environment, limited sample size, insufficient data and so on. Thus, it was urgently necessary to perform a comprehensive up-to-date meta-analysis as an effective methodology to draw an overall objective appraisal on the association between CTLA-4 polymorphism and RA susceptibility.
In the present meta-analysis, we extracted 66 studies with 21681 cases and 23457 controls to inspect the correlation between three most-often SNPs in the CTLA-4 gene and the risk of RA. There were 22 studies with 16394 cases and 17453 controls for rs3087243 SNP, 34 studies with 11452 cases and 12444 controls for rs231775 SNP, and 10 studies with 2477 cases and 2941 controls for rs5742909 SNP. For rs3087243 polymorphism, our findings demonstrated a decreased susceptibility of RA both in total and in Caucasians in any gene mode. In total, carriers with allele A reduced an approximate 13% risk of RA than ones with allele G and genotype AA reduced 20% or so than genotype GG. Moreover, a decreased susceptibility of RA was respectively also found among Asians in the allele and homozygote comparison and among Latin Americans in the heterozygote and dominant comparison. However, only one study was included in Latin Americans and Asians so it needed to enlarge sample size to further research. For rs231775 polymorphism, significant association did exist among the whole population in all genetic models except recessive model: compared with allele A and genotype AA, allele G and genotype GG and GA respectively was associated with an increased risk of RA. The same association was observed in Asians and Latin Americans in the subgroup analysis. On the contrary, no significant association between rs231775 SNP and RA risk could be detected in Caucasians and Africans using any gene model after excluding the Elshazli R’s study [24] with the apparent heterogeneity. Here, it should be noted that only one or two case–control study was included in Africans and Latin Americans, so the conclusions were not particularly convincing. For rs5742909 polymorphism, no significant association between this locus polymorphism and RA risk was observed among any population in any model. Although the heterogeneity existed in some genetic model, but no obvious change had happened in heterogeneity and P value for the pooled ORs when each study was individually removed by sensitivity analysis.
With regard to the diverse results of the same SNP on different populations, it might be attributed to clinical and genetic real heterogeneity of RA, interaction of genetic background and region environment, and even lack of vigorous statistical power. Besides, it was noteworthy that one important factor for the diverse and disparate results was linkage disequilibrium (LD). These CTLA-4 SNPs might be not definitely the causative alleles, but they were likely to be in LD with the causative alleles which were yet unidentified. And, LD was different between ethnic and racial groups.
It should be pointed out that previous several meta-analyses have summarized the effect of CTLA-4 polymorphism on RA risk [21–23, 65]. But a few points need to be taken notice. On one hand, the previous conclusions were discordant as the following: the conclusion of Li’s (2014) study [65] on the association of rs231775 SNP of CTLA-4 with RA was contrary to the others; the genetic models which indicated significant association were diverse in these analyses. These differences were mainly originated from divergent diagnostic criteria, limited number of studies and sample sizes. On the other hand, all these meta-analyses focused on only one of the three well-studied loci except Li’s study [23] on two. As we all know, the expression and function of the protein are determined by the whole gene. Therefore, it is of great necessity to investigate simultaneously the effect of all the 3 SNPs on RA risk to obtain an overall evaluation. Besides, the number of included studies in previous meta-analyses was small. Some original association studies [9, 15, 24–27] have emerged in the past few years and they can be incorporated. Taking these points into considerations, we updated the meta-analysis to achieve a more valid and comprehensive estimation on the association of CTLA-4 gene and RA susceptibility.
However, some limitations of our study should be acknowledged. Firstly, the small sample size in some studies and the limited studies for some stratified analysis were not sufficient enough to detect the relationship. Especially, the results of populations including only one study should be interpreted with caution. Secondly, we only investigated the role of three loci polymorphisms. As CTLA-4 gene had various SNPs, the function of protein CTLA-4 depended on the whole gene and RA was a multigene susceptibility disease, more SNPs of CTLA-4 should be included. Thirdly, certain degree of heterogeneity still existed in rs5742909 polymorphism and some genetic models. Although the elimination of each single study did not distinctly alter the P value, the results must still be treated cautiously. Fourthly, inadequate raw data in some studies result in the inability to calculate the number of the genotypes and perform stratified analysis by age, gender and autoantibody status such as RF etc. As a consequence, any potential gene-environment and gene-gene interactions could not be accessed.
In conclusion, this meta-analysis suggested that rs3087243 polymorphisms were corelated with a reduced RA risk in both Asian and Caucasian populations, rs231775 polymorphisms was associated with an increased risk of RA in Asians, and rs5742909 polymorphism had no significant association with RA risk. Larger-scale studies of populations with different ethnicities are encouraged to validate the role CTLA-4 played in the pathogenesis of RA.
Materials and Methods
This meta-analysis was performed according to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines [66].
Search strategy
From the databases PubMed, EMBASE, Web of Science and, the China National Knowledge Infrastructure (CNKI) and Wan Fang data, a comprehensive systematic literature retrieval was conducted to derive all relevant studies published before 10 October, 2020 (the search was constantly updated to submission). The following terms as Medical Subject Heading and free words were applied: “CTLA-4 or cytotoxic T lymphocyte antigen-4” and “single nucleotide polymorphism or polymorphism or variant or variation” and “rheumatoid arthritis or RA”. The bibliographic lists of included studies were also browsed for potential related studies. There were no restrictions on language and publication date in this study.
Inclusion and exclusion criteria
The current meta-analysis used the following inclusion criteria to screen available literatures: 1) case-control study; 2) evaluation of the associations between CTLA-4 (rs3087243, rs231775 and rs5742909)polymorphism and RA risk; 3) with sufficient data for extract odds ratios (ORs) and 95% confidence intervals(CIs); (4) with reported allele or genotype numbers or frequencies in cases and control group; 5) with a clear diagnostic criteria. Accordingly, we excluded meaninglessness literatures if they had the following trait: 1) case report, comment, animal studies and conference abstracts; 2) with no detailed allele or genotype data; 3) duplications or no controls.
Data extraction and assess of quality
Two independent investigators respectively conducted a literature search according to the above search strategy, screened each article based on the predesigned inclusion and exclusion criteria, and extracted data from these eligible studies. It would be settled by discussion with the third party when the disagreement between investigators occurred. The following information was collected from every paper: 1) first author's surname, 2) the year of publication, 3) country or region of origin, 4) ethnicity, 5) total numbers of cases and controls, 6) genotype method, 7) diagnostic criteria, 8) polymorphism locus, 9) allele distribution or/and genotype distribution.
The methodological quality of included studies was accessed in light of the Newcastle–Ottawa Scale (NOS) for the evaluation of observational studies [67]. In brief, three broad perspectives were evaluated using the Star system (http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp). Any divergence between two investigators was solved by discussion until agreement was reached.
Statistical analysis
The strength of association of rs231775, rs5742909 and rs3087243 SNPs with RA risk was appraised via estimating ORs with their corresponding 95% CIs. For each SNP, the pooled ORs were calculated individually for five gene models (allele model, homozygote model, heterozygote model, dominant model and recessive model). The Z test was used to evaluate the significance of the pooled ORs. p<0.05 was judged as statistically significant difference. Statistical Heterogeneity between studies was assessed by Chi square and I2 values which range from 0% to 100%. 25%, 50%, and 75% were regarded as respectively low, moderate, and high level [68, 69]. The random -effect model was employed when the value of I2 was more than 50%. If not, the fixed effect model was employed. Hardy–Weinberg equilibrium (HWE) was tested in the control group for all studies by Chi-square test to judge whether the selection bias existed. Potential publication bias was examined by funnel plots. Besides, the current meta-analysis had carried out subgroup analyses by the racial descent to assess the effects of ethnic background.
The above statistical analyses were performed using Review Manager 5.3 software (Nordic Cochrane Centre, Cochrane Collaboration, Copenhagen). All the P values were 2-sided and P<0.05 signified statistically significance.
Supplementary Materials
Author Contributions
J.X. and H. L. conceived and designed this study. C. Z., S.G, X. Y., Z. S. and S. L. performed the experiments. C. Z., S.G analyzed the data. C. Z. and H. L. draft the manuscript. X.S. and J.X. revised the paper. All authors have contributed to the final version and approved the final manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Funding
This study is supported by National Natural Science Foundation of China, Nos.81772396.
References
- 1. Smolen JS, Aletaha D, McInnes IB. Rheumatoid arthritis. Lancet. 2016; 388:2023–38. https://doi.org/10.1016/S0140-6736(16)30173-8 [PubMed]
- 2. Minichiello E, Semerano L, Boissier MC. Time trends in the incidence, prevalence, and severity of rheumatoid arthritis: A systematic literature review. Joint Bone Spine. 2016; 83:625–30. https://doi.org/10.1016/j.jbspin.2016.07.007 [PubMed]
- 3. Cross M, Smith E, Hoy D, Carmona L, Wolfe F, Vos T, Williams B, Gabriel S, Lassere M, Johns N, Buchbinder R, Woolf A, March L. The global burden of rheumatoid arthritis: estimates from the global burden of disease 2010 study. Ann Rheum Dis. 2014; 73:1316–22. https://doi.org/10.1136/annrheumdis-2013-204627 [PubMed]
- 4. MacGregor AJ, Snieder H, Rigby AS, Koskenvuo M, Kaprio J, Aho K, Silman AJ. Characterizing the quantitative genetic contribution to rheumatoid arthritis using data from twins. Arthritis Rheum. 2000; 43:30–37. https://doi.org/10.1002/1529-0131(200001)43:1<30::AID-ANR5>3.0.CO;2-B [PubMed]
- 5. Okada Y, Wu D, Trynka G, Raj T, Terao C, Ikari K, Kochi Y, Ohmura K, Suzuki A, Yoshida S, Graham RR, Manoharan A, Ortmann W, et al, and RACI consortium, and GARNET consortium. Genetics of rheumatoid arthritis contributes to biology and drug discovery. Nature. 2014; 506:376–81. https://doi.org/10.1038/nature12873 [PubMed]
- 6. Kim K, Bang SY, Lee HS, Bae SC. Update on the genetic architecture of rheumatoid arthritis. Nat Rev Rheumatol. 2017; 13:13–24. https://doi.org/10.1038/nrrheum.2016.176 [PubMed]
- 7. Leng RX, Di DS, Ni J, Wu XX, Zhang LL, Wang XF, Liu RS, Huang Q, Fan YG, Pan HF, Wang B, Ye DQ. Identification of new susceptibility loci associated with rheumatoid arthritis. Ann Rheum Dis. 2020; 79:1565–71. https://doi.org/10.1136/annrheumdis-2020-217351 [PubMed]
- 8. Costenbader KH, Chang SC, De Vivo I, Plenge R, Karlson EW. Genetic polymorphisms in PTPN22, PADI-4, and CTLA-4 and risk for rheumatoid arthritis in two longitudinal cohort studies: evidence of gene-environment interactions with heavy cigarette smoking. Arthritis Res Ther. 2008; 10:R52. https://doi.org/10.1186/ar2421 [PubMed]
- 9. Luterek-Puszyńska K, Malinowski D, Paradowska-Gorycka A, Safranow K, Pawlik A. CD28, CTLA-4 and CCL5 gene polymorphisms in patients with rheumatoid arthritis. Clin Rheumatol. 2017; 36:1129–35. https://doi.org/10.1007/s10067-016-3496-2 [PubMed]
- 10. Klocke K, Sakaguchi S, Holmdahl R, Wing K. Induction of autoimmune disease by deletion of CTLA-4 in mice in adulthood. Proc Natl Acad Sci USA. 2016; 113:E2383–92. https://doi.org/10.1073/pnas.1603892113 [PubMed]
- 11. Sansom DM. IMMUNOLOGY. Moving CTLA-4 from the trash to recycling. Science. 2015; 349:377–78. https://doi.org/10.1126/science.aac7888 [PubMed]
- 12. Ueda H, Howson JM, Esposito L, Heward J, Snook H, Chamberlain G, Rainbow DB, Hunter KM, Smith AN, Di Genova G, Herr MH, Dahlman I, Payne F, et al. Association of the T-cell regulatory gene CTLA4 with susceptibility to autoimmune disease. Nature. 2003; 423:506–11. https://doi.org/10.1038/nature01621 [PubMed]
- 13. Zhernakova A, Eerligh P, Barrera P, Wesoly JZ, Huizinga TW, Roep BO, Wijmenga C, Koeleman BP. CTLA4 is differentially associated with autoimmune diseases in the Dutch population. Hum Genet. 2005; 118:58–66. https://doi.org/10.1007/s00439-005-0006-z [PubMed]
- 14. Walker EJ, Hirschfield GM, Xu C, Lu Y, Liu X, Lu Y, Coltescu C, Wang K, Newman WG, Bykerk V, Keystone EC, Mosher D, Amos CI, et al. CTLA4/ICOS gene variants and haplotypes are associated with rheumatoid arthritis and primary biliary cirrhosis in the Canadian population. Arthritis Rheum. 2009; 60:931–37. https://doi.org/10.1002/art.24412 [PubMed]
- 15. Schulz S, Zimmer P, Pütz N, Jurianz E, Schaller HG, Reichert S. rs2476601 in PTPN22 gene in rheumatoid arthritis and periodontitis-a possible interface? J Transl Med. 2020; 18:389. https://doi.org/10.1186/s12967-020-02548-w [PubMed]
- 16. Yang J, McGovern A, Martin P, Duffus K, Ge X, Zarrineh P, Morris AP, Adamson A, Fraser P, Rattray M, Eyre S. Analysis of chromatin organization and gene expression in T cells identifies functional genes for rheumatoid arthritis. Nat Commun. 2020; 11:4402. https://doi.org/10.1038/s41467-020-18180-7 [PubMed]
- 17. Barton A, Myerscough A, John S, Gonzalez-Gay M, Ollier W, Worthington J. A single nucleotide polymorphism in exon 1 of cytotoxic T-lymphocyte-associated-4 (CTLA-4) is not associated with rheumatoid arthritis. Rheumatology (Oxford). 2000; 39:63–66. https://doi.org/10.1093/rheumatology/39.1.63 [PubMed]
- 18. Plenge RM, Padyukov L, Remmers EF, Purcell S, Lee AT, Karlson EW, Wolfe F, Kastner DL, Alfredsson L, Altshuler D, Gregersen PK, Klareskog L, Rioux JD. Replication of putative candidate-gene associations with rheumatoid arthritis in >4,000 samples from North America and Sweden: association of susceptibility with PTPN22, CTLA4, and PADI4. Am J Hum Genet. 2005; 77:1044–60. https://doi.org/10.1086/498651 [PubMed]
- 19. Plant D, Flynn E, Mbarek H, Dieudé P, Cornelis F, Arlestig L, Dahlqvist SR, Goulielmos G, Boumpas DT, Sidiropoulos P, Johansen JS, Ørnbjerg LM, Hetland ML, et al. Investigation of potential non-HLA rheumatoid arthritis susceptibility loci in a European cohort increases the evidence for nine markers. Ann Rheum Dis. 2010; 69:1548–53. https://doi.org/10.1136/ard.2009.121020 [PubMed]
- 20. Liu CP, Jiang JA, Wang T, Liu XM, Gao L, Zhu RR, Shen Y, Wu M, Xu T, Zhang XG. CTLA-4 and CD86 genetic variants and haplotypes in patients with rheumatoid arthritis in southeastern China. Genet Mol Res. 2013; 12:1373–82. https://doi.org/10.4238/2013.April.25.8 [PubMed]
- 21. Han S, Li Y, Mao Y, Xie Y. Meta-analysis of the association of CTLA-4 exon-1 +49A/G polymorphism with rheumatoid arthritis. Hum Genet. 2005; 118:123–32. https://doi.org/10.1007/s00439-005-0033-9 [PubMed]
- 22. Lee YH, Bae SC, Choi SJ, Ji JD, Song GG. Association between the CTLA-4 +49 A/G polymorphism and susceptibility to rheumatoid arthritis: a meta-analysis. Mol Biol Rep. 2012; 39:5599–605. https://doi.org/10.1007/s11033-011-1364-3 [PubMed]
- 23. Li X, Zhang C, Zhang J, Zhang Y, Wu Z, Yang L, Xiang Z, Qi Z, Zhang X, Xiao X. Polymorphisms in the CTLA-4 gene and rheumatoid arthritis susceptibility: a meta-analysis. J Clin Immunol. 2012; 32:530–39. https://doi.org/10.1007/s10875-012-9650-y [PubMed]
- 24. Elshazli R, Settin A, Salama A. Cytotoxic T lymphocyte associated antigen-4 (CTLA-4) +49 A>G gene polymorphism in Egyptian cases with rheumatoid arthritis. Gene. 2015; 558:103–07. https://doi.org/10.1016/j.gene.2014.12.046 [PubMed]
- 25. Sameem M, Rani A, Bashir R, Riaz N, Batool SA, Irfan S, Arshad M, Nawaz SK. CTLA-4+49 Polymorphism and Susceptibility to Rheumatoid Arthritis in Pakistani Population. Pak J Zool. 2015; 47:1731–37.
- 26. Vernerova L, Spoutil F, Vlcek M, Krskova K, Penesova A, Meskova M, Marko A, Raslova K, Vohnout B, Rovensky J, Killinger Z, Jochmanova I, Lazurova I, et al. A Combination of CD28 (rs1980422) and IRF5 (rs10488631) Polymorphisms Is Associated with Seropositivity in Rheumatoid Arthritis: A Case Control Study. PLoS One. 2016; 11:e0153316. https://doi.org/10.1371/journal.pone.0153316 [PubMed]
- 27. Fattah SA, Ghattas MH, Saleh SM, Abo-Elmatty DM. Cytotoxic T-lymphocyte-associated protein 4 gene polymorphism is related to rheumatoid arthritis in Egyptian population. Arch Physiol Biochem. 2017; 123:50–53. https://doi.org/10.1080/13813455.2016.1230135 [PubMed]
- 28. Seidl C, Donner H, Fischer B, Usadel KH, Seifried E, Kaltwasser JP, Badenhoop K. CTLA4 codon 17 dimorphism in patients with rheumatoid arthritis. Tissue Antigens. 1998; 51:62–66. https://doi.org/10.1111/j.1399-0039.1998.tb02947.x [PubMed]
- 29. Gonzalez-Escribano MF, Rodriguez R, Valenzuela A, Garcia A, Garcia-Lozano JR, Nuñez-Roldan A. CTLA4 polymorphisms in Spanish patients with rheumatoid arthritis. Tissue Antigens. 1999; 53:296–300. https://doi.org/10.1034/j.1399-0039.1999.530311.x [PubMed]
- 30. Matsushita M, Tsuchiya N, Shiota M, Komata T, Matsuta K, Zama K, Oka T, Juji T, Yamane A, Tokunaga K. Lack of a strong association of CTLA-4 exon 1 polymorphism with the susceptibility to rheumatoid arthritis and systemic lupus erythematosus in Japanese: an association study using a novel variation screening method. Tissue Antigens. 1999; 54:578–84. https://doi.org/10.1034/j.1399-0039.1999.540607.x [PubMed]
- 31. Yanagawa T, Gomi K, Nakao EI, Inada S. CTLA-4 gene polymorphism in Japanese patients with rheumatoid arthritis. J Rheumatol. 2000; 27:2740–42. [PubMed]
- 32. Hadj Kacem H, Kaddour N, Adyel FZ, Bahloul Z, Ayadi H. HLA-DQB1 CAR1/CAR2, TNFa IR2/IR4 and CTLA-4 polymorphisms in Tunisian patients with rheumatoid arthritis and Sjögren’s syndrome. Rheumatology (Oxford). 2001; 40:1370–74. https://doi.org/10.1093/rheumatology/40.12.1370 [PubMed]
- 33. Milicic A, Brown MA, Wordsworth BP. Polymorphism in codon 17 of the CTLA-4 gene (+49 A/G) is not associated with susceptibility to rheumatoid arthritis in British Caucasians. Tissue Antigens. 2001; 58:50–54. https://doi.org/10.1034/j.1399-0039.2001.580110.x [PubMed]
- 34. Lee YH, Choi SJ, Ji JD, Song GG. No association of polymorphisms of the CTLA-4 exon 1(+49) and promoter(-318) genes with rheumatoid arthritis in the Korean population. Scand J Rheumatol. 2002; 31:266–70. https://doi.org/10.1080/030097402760375142 [PubMed]
- 35. Vaidya B, Pearce SH, Charlton S, Marshall N, Rowan AD, Griffiths ID, Kendall-Taylor P, Cawston TE, Young-Min S. An association between the CTLA4 exon 1 polymorphism and early rheumatoid arthritis with autoimmune endocrinopathies. Rheumatology (Oxford). 2002; 41:180–83. https://doi.org/10.1093/rheumatology/41.2.180 [PubMed]
- 36. Lee CS, Lee YJ, Liu HF, Su CH, Chang SC, Wang BR, Chen TL, Liu TL. Association of CTLA4 gene A-G polymorphism with rheumatoid arthritis in Chinese. Clin Rheumatol. 2003; 22:221–24. https://doi.org/10.1007/s10067-003-0720-7 [PubMed]
- 37. Barton A, Jury F, Eyre S, Bowes J, Hinks A, Ward D, Worthington J. Haplotype analysis in simplex families and novel analytic approaches in a case-control cohort reveal no evidence of association of the CTLA-4 gene with rheumatoid arthritis. Arthritis Rheum. 2004; 50:748–52. https://doi.org/10.1002/art.20118 [PubMed]
- 38. Liu MF, Wang CR, Chen PC, Lin TL. CTLA-4 gene polymorphism in promoter and exon-1 regions is not associated with Chinese patients with rheumatoid arthritis. Clin Rheumatol. 2004; 23:180–81. https://doi.org/10.1007/s10067-003-0776-4 [PubMed]
- 39. Miterski B, Drynda S, Böschow G, Klein W, Oppermann J, Kekow J, Epplen JT. Complex genetic predisposition in adult and juvenile rheumatoid arthritis. BMC Genet. 2004; 5:2. https://doi.org/10.1186/1471-2156-5-2 [PubMed]
- 40. Orozco G, Torres B, Núñez-Roldán A, González-Escribano MF, Martín J. Cytotoxic T-lymphocyte antigen-4-CT60 polymorphism in rheumatoid arthritis. Tissue Antigens. 2004; 64:667–70. https://doi.org/10.1111/j.1399-0039.2004.00318.x [PubMed]
- 41. Lei C, Dongqing Z, Yeqing S, Oaks MK, Lishan C, Jianzhong J, Jie Q, Fang D, Ningli L, Xinghai H, Daming R. Association of the CTLA-4 gene with rheumatoid arthritis in Chinese Han population. Eur J Hum Genet. 2005; 13:823–28. https://doi.org/10.1038/sj.ejhg.5201423 [PubMed]
- 42. Feng ZL. [CTLA-4 gene A/G(49) polymorphism association with SLE and RA and expression of soluble CTLA-4 in patients with SLE and with RA]. Shandong University. 2005.
- 43. Suppiah V, O’Doherty C, Heggarty S, Patterson CC, Rooney M, Vandenbroeck K. The CTLA4+49A/G and CT60 polymorphisms and chronic inflammatory arthropathies in Northern Ireland. Exp Mol Pathol. 2006; 80:141–46. https://doi.org/10.1016/j.yexmp.2005.09.004 [PubMed]
- 44. Takeuchi F, Kawasugi K, Mori M, Nakaue N, Kobayashi N, Kuwata S, Murayama T, Matsuta K. The genetic contribution of CTLA-4 dimorphisms in promoter and exon 1 regions in Japanese patients with rheumatoid arthritis. Scand J Rheumatol. 2006; 35:154–55. https://doi.org/10.1080/03009740500407651 [PubMed]
- 45. Zhou Y, Xiao LS. [Association of the polymorphism of CTLA-4 gene with systemic lupus erythematosus or rheumatoid arthitis in the Chinese population]. Immunol J. 2007; 3:349–50.
- 46. Tsukahara S, Iwamoto T, Ikari K, Inoue E, Tomatsu T, Hara M, Yamanaka H, Kamatani N, Momohara S. CTLA-4 CT60 polymorphism is not an independent genetic risk marker of rheumatoid arthritis in a Japanese population. Ann Rheum Dis. 2008; 67:428–29. https://doi.org/10.1136/ard.2007.079186 [PubMed]
- 47. Barton A, Eyre S, Ke X, Hinks A, Bowes J, Flynn E, Martin P, Wilson AG, Morgan AW, Emery P, Steer S, Hocking LJ, Reid DM, et al, and YEAR Consortium, and BIRAC Consortium. Identification of AF4/FMR2 family, member 3 (AFF3) as a novel rheumatoid arthritis susceptibility locus and confirmation of two further pan-autoimmune susceptibility genes. Hum Mol Genet. 2009; 18:2518–22. https://doi.org/10.1093/hmg/ddp177 [PubMed]
- 48. Daha NA, Kurreeman FA, Marques RB, Stoeken-Rijsbergen G, Verduijn W, Huizinga TW, Toes RE. Confirmation of STAT4, IL2/IL21, and CTLA4 polymorphisms in rheumatoid arthritis. Arthritis Rheum. 2009; 60:1255–60. https://doi.org/10.1002/art.24503 [PubMed]
- 49. Kelley JM, Hughes LB, Faggard JD, Danila MI, Crawford MH, Edberg Y, Padilla MA, Tiwari HK, Westfall AO, Alarcón GS, Conn DL, Jonas BL, Callahan LF, et al. An African ancestry-specific allele of CTLA4 confers protection against rheumatoid arthritis in African Americans. PLoS Genet. 2009; 5:e1000424. https://doi.org/10.1371/journal.pgen.1000424 [PubMed]. Retraction in: PLoS Genet. 2009; 5. https://doi.org/10.1371/annotation/80bd7285-9d2d-403a-8e6f-9c375bf977ca [PubMed]
- 50. Muñoz-Valle JF, Valle Y, Padilla-Gutiérrez JR, Parra-Rojas I, Rangel-Villalobos H, Vázquez del Mercado M, Ledezma-Lozano IY, Villafan-Bernal JR, Armendáriz-Borunda J, Pereira-Suárez AL. The +49A>G CTLA-4 polymorphism is associated with rheumatoid arthritis in Mexican population. Clin Chim Acta. 2010; 411:725–28. https://doi.org/10.1016/j.cca.2010.02.001 [PubMed]
- 51. Benhatchi K, Jochmanová I, Habalová V, Wagnerová H, Lazúrová I. CTLA4 exon1 A49G polymorphism in Slovak patients with rheumatoid arthritis and Hashimoto thyroiditis-results and the review of the literature. Clin Rheumatol. 2011; 30:1319–24. https://doi.org/10.1007/s10067-011-1752-z [PubMed]
- 52. Danoy P, Wei M, Johanna H, Jiang L, He D, Sun L, Zeng X, Visscher PM, Brown MA, Xu H. Association of variants in MMEL1 and CTLA4 with rheumatoid arthritis in the Han Chinese population. Ann Rheum Dis. 2011; 70:1793–97. https://doi.org/10.1136/ard.2010.144576 [PubMed]
- 53. El-Gabalawy HS, Robinson DB, Daha NA, Oen KG, Smolik I, Elias B, Hart D, Bernstein CN, Sun Y, Lu Y, Houwing-Duistermaat JJ, Siminovitch KA. Non-HLA genes modulate the risk of rheumatoid arthritis associated with HLA-DRB1 in a susceptible North American Native population. Genes Immun. 2011; 12:568–74. https://doi.org/10.1038/gene.2011.30 [PubMed]
- 54. AlFadhli S. Overexpression and secretion of the soluble CTLA-4 splice variant in various autoimmune diseases and in cases with overlapping autoimmunity. Genet Test Mol Biomarkers. 2013; 17:336–41. https://doi.org/10.1089/gtmb.2012.0391 [PubMed]
- 55. Tang MJ, Zhou ZB. Association of the CTLA-4 +49A/G polymorphism with rheumatoid arthritis in Chinese Han population. Mol Biol Rep. 2013; 40:2627–31. https://doi.org/10.1007/s11033-012-2349-6 [PubMed]
- 56. Torres-Carrillo N, Ontiveros-Mercado H, Torres-Carrillo NM, Parra-Rojas I, Rangel-Villalobos H, Ramírez-Dueñas MG, Gutiérrez-Ureña SR, Valle Y, Muñoz-Valle JF. The -319C/+49G/CT60G haplotype of CTLA-4 gene confers susceptibility to rheumatoid arthritis in Mexican population. Cell Biochem Biophys. 2013; 67:1217–28. https://doi.org/10.1007/s12013-013-9640-6 [PubMed]
- 57. Walker LS, Sansom DM. Confusing signals: recent progress in CTLA-4 biology. Trends Immunol. 2015; 36:63–70. https://doi.org/10.1016/j.it.2014.12.001 [PubMed]
- 58. Cope AP, Schulze-Koops H, Aringer M. The central role of T cells in rheumatoid arthritis. Clin Exp Rheumatol. 2007 (Suppl 46); 25:S4–11. [PubMed]
- 59. Cao J, Zou L, Luo P, Chen P, Zhang L. Increased production of circulating soluble co-stimulatory molecules CTLA-4, CD28 and CD80 in patients with rheumatoid arthritis. Int Immunopharmacol. 2012; 14:585–92. https://doi.org/10.1016/j.intimp.2012.08.004 [PubMed]
- 60. Choi IS, Yoo HS, Collisson EW. Evaluation of expression patterns of feline CD28 and CTLA-4 in feline immunodeficiency virus (FIV)-infected and FIV antigen-induced PBMC. J Vet Sci. 2000; 1:97–103. https://doi.org/10.4142/jvs.2000.1.2.97 [PubMed]
- 61. Zhang W, Wang F, Wang B, Zhang J, Yu JY. Intraarticular gene delivery of CTLA4-FasL suppresses experimental arthritis. Int Immunol. 2012; 24:379–88. https://doi.org/10.1093/intimm/dxs041 [PubMed]
- 62. Cutolo M, Soldano S, Montagna P, Sulli A, Seriolo B, Villaggio B, Triolo P, Clerico P, Felli L, Brizzolara R. CTLA4-Ig interacts with cultured synovial macrophages from rheumatoid arthritis patients and downregulates cytokine production. Arthritis Res Ther. 2009; 11:R176. https://doi.org/10.1186/ar2865 [PubMed]
- 63. Körmendy D, Hoff H, Hoff P, Bröker BM, Burmester GR, Brunner-Weinzierl MC. Impact of the CTLA-4/CD28 axis on the processes of joint inflammation in rheumatoid arthritis. Arthritis Rheum. 2013; 65:81–87. https://doi.org/10.1002/art.37714 [PubMed]
- 64. Kremer JM, Westhovens R, Leon M, Di Giorgio E, Alten R, Steinfeld S, Russell A, Dougados M, Emery P, Nuamah IF, Williams GR, Becker JC, Hagerty DT, Moreland LW. Treatment of rheumatoid arthritis by selective inhibition of T-cell activation with fusion protein CTLA4Ig. N Engl J Med. 2003; 349:1907–15. https://doi.org/10.1056/NEJMoa035075 [PubMed]
- 65. Li G, Shi F, Liu J, Li Y. The effect of CTLA-4 A49G polymorphism on rheumatoid arthritis risk: a meta-analysis. Diagn Pathol. 2014; 9:157. https://doi.org/10.1186/s13000-014-0157-0 [PubMed]
- 66. Moher D, Liberati A, Tetzlaff J, Altman DG, and PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. BMJ. 2009; 339:b2535. https://doi.org/10.1136/bmj.b2535 [PubMed]
- 67. Stang A. Critical evaluation of the Newcastle-Ottawa scale for the assessment of the quality of nonrandomized studies in meta-analyses. Eur J Epidemiol. 2010; 25:603–05. https://doi.org/10.1007/s10654-010-9491-z [PubMed]
- 68. Higgins JP, Thompson SG. Quantifying heterogeneity in a meta-analysis. Stat Med. 2002; 21:1539–58. https://doi.org/10.1002/sim.1186 [PubMed]
- 69. Higgins JP, Thompson SG, Deeks JJ, Altman DG. Measuring inconsistency in meta-analyses. BMJ. 2003; 327:557–60. https://doi.org/10.1136/bmj.327.7414.557 [PubMed]