



Introduction
Gastric cancer (GC) is the 3rd most common cause of tumor-associated death and the 3rd most commonly diagnosed tumor worldwide [1–4]. The cause and pathogenesis of this disease are complex and associated with many factors [5–8]. Despite the great achievements that have been made in GC therapeutics, the survival rate of GC patients remains unsatisfactory [9–12]. The major challenge in the treatment of advanced GC is the manifestation of peritoneal, distal organ and lymphatic metastases [13–15]. Therefore, a detailed and improved understanding of the molecular mechanisms underlying GC progression and development is greatly needed.
As one type of ncRNA, lncRNAs are more than 200 nt long [16–18]. Growing evidence has revealed that lncRNAs are involved in several cellular pathways and processes, including cell apoptosis, angiogenesis, differentiation, immune responses, proliferation and metabolism [19–21]. Compelling studies have shown that lncRNAs play critical roles in the development and initiation of tumors and that lncRNAs act as tumor suppressor genes or oncogenes in tumors [22–25]. Recently, a new lncRNA, MSC-AS1, was identified as a crucial modulator in the development of tumors [26–30]. Cao et al [31]. found that MSC-AS1 promoted hepatocellular carcinoma progression by enhancing PGK1 expression. Yao et al [30]. showed that |MSC-AS1 increased nasopharyngeal carcinoma development by regulating miR-524-5p/NR4A2. However, the functional role of MSC-AS1 in GC remains unknown.
Results
MSC-AS1 and DDX5 were overexpressed and miR-142-5p was downregulated in GC cells
MSC-AS1 was overexpressed in GC cells (HGC-27, MKN-45, SGC-7901 and MGC-803 cells) compared to GES cells (Figure 1A). miR-142-5p was downregulated in GC cells (HGC-27, MKN-45, SGC-7901 and MGC-803 cells) compared to GES cells (Figure 1B).
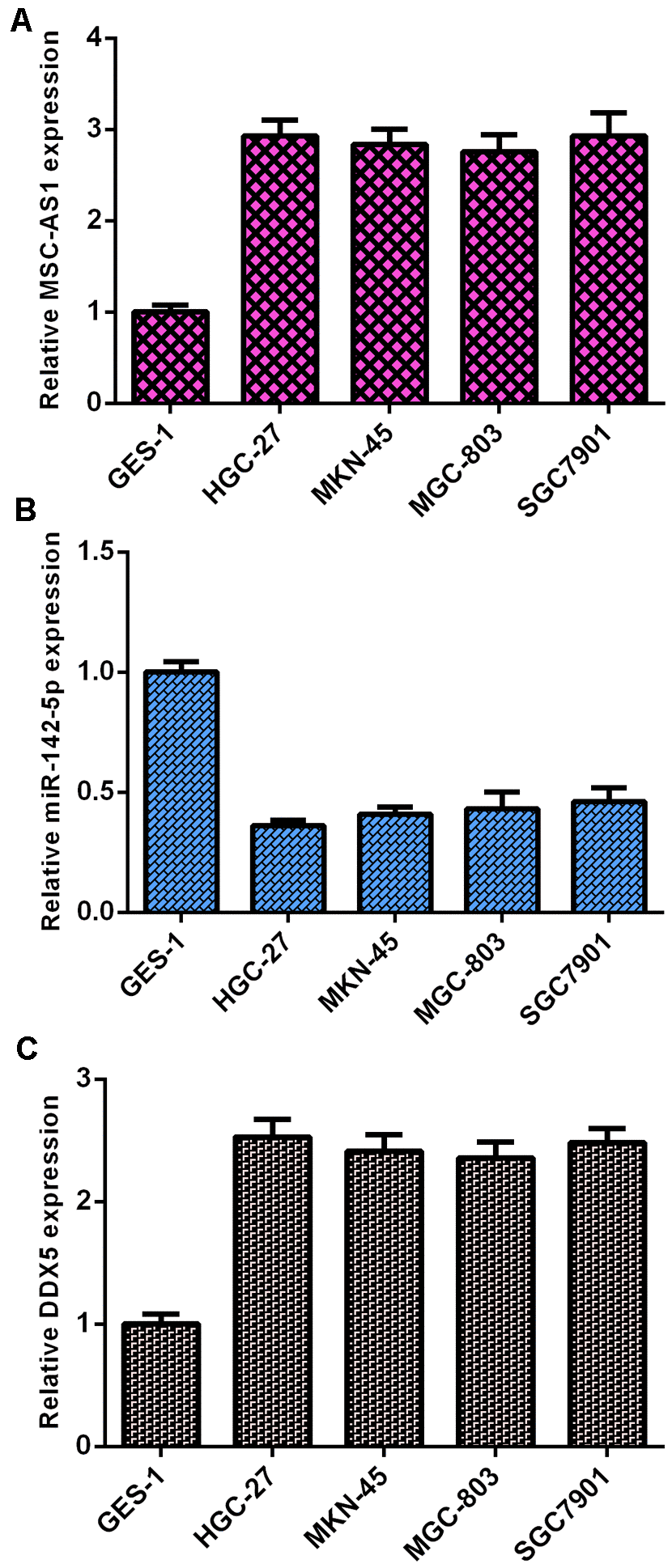
Figure 1. MSC-AS1 and DDX5 were overexpressed and miR-142-5p was downregulated in GC cells. (A) The expression of MSC-AS1 was detected by qRT-PCR analysis. GAPDH was used as the internal control. (B) The expression of miR-142-5p was detected by qRT-PCR analysis. U6 was used as the internal control. (C) DDX5 was upregulated in GC cells (HGC-27, MKN-45, SGC-7901 and MGC-803 cells) compared to GES cells. GAPDH was used as the internal control.
MSC-AS1 was upregulated in GC specimens
We observed that MSC-AS1 was upregulated in GC specimens compared with paired normal specimens (Figure 2A). The level of MSC-AS1 was upregulated in 29 GC specimens (72.5%, 29/40) compared to their paired normal specimens (Figure 2B).
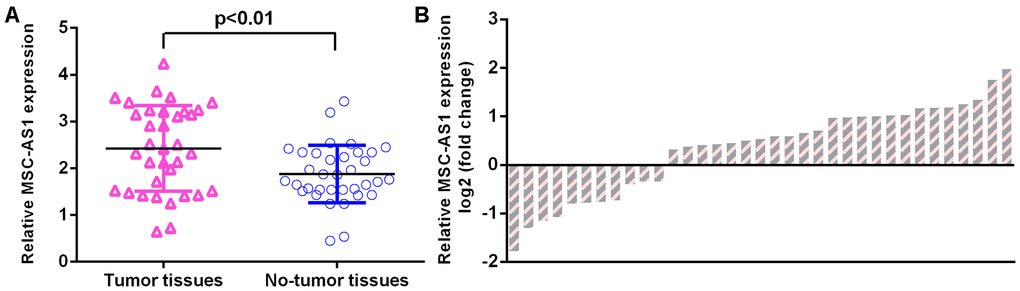
Figure 2. MSC-AS1 was upregulated in GC specimens. (A) MSC-AS1 was upregulated in GC specimens compared with paired normal specimens. (B) The level of MSC-AS1 was upregulated in 29 GC specimens (72.5%, 29/40) compared to their paired normal specimens. GAPDH was used as the internal control.
miR-142-5p was downregulated in GC specimens
We showed that miR-142-5p was downregulated in GC specimens compared to paired normal specimens (Figure 3A). The level of miR-142-5p was decreased in 28 GC specimens (70.0%, 28/40) compared to their paired normal specimens (Figure 3B). MSC-AS1 expression was inversely correlated with miR-142-5p expression in GC specimens (Figure 3C).
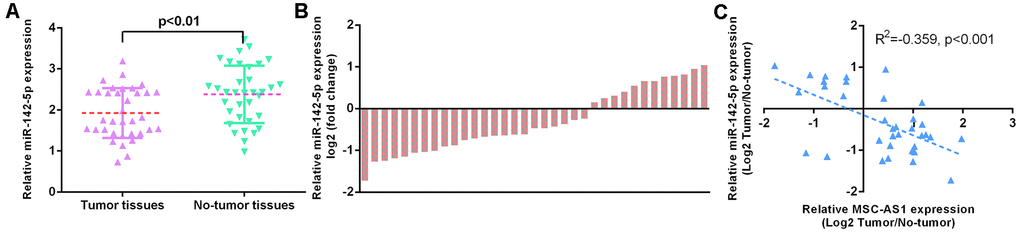
Figure 3. miR-142-5p was downregulated in GC specimens. (A) miR-142-5p was downregulated in GC specimens compared to paired normal specimens. (B) The level of miR-142-5p was decreased in 28 GC specimens (70.0%, 28/40) compared to their paired normal specimens. (C) MSC-AS1 expression was inversely correlated with miR-142-5p expression in GC specimens. U6 was used as the internal control.
MSC-AS1 increased cell growth, cell cycle progression and inflammatory mediator secretion
The level of MSC-AS1 was significantly overexpressed in HGC-27 cells after treatment with the pcDNA-MSC-AS1 plasmid (Figure 4A). Elevated expression of MSC-AS1 enhanced the expression of Ki-67 (Figure 4B) and CKD2 (Figure 4C) in HGC-27 cells. MSC-AS1 overexpression induced cell cycle progression in HGC-27 cells (Figure 4D). Overexpression of MSC-AS1 increased HGC-27 cell growth, according to the CCK-8 analysis (Figure 4E). Overexpression of MSC-AS1 enhanced the secretion of the inflammatory mediators IL-1β, IL-6 and TNF-α (Figure 4F–4H).
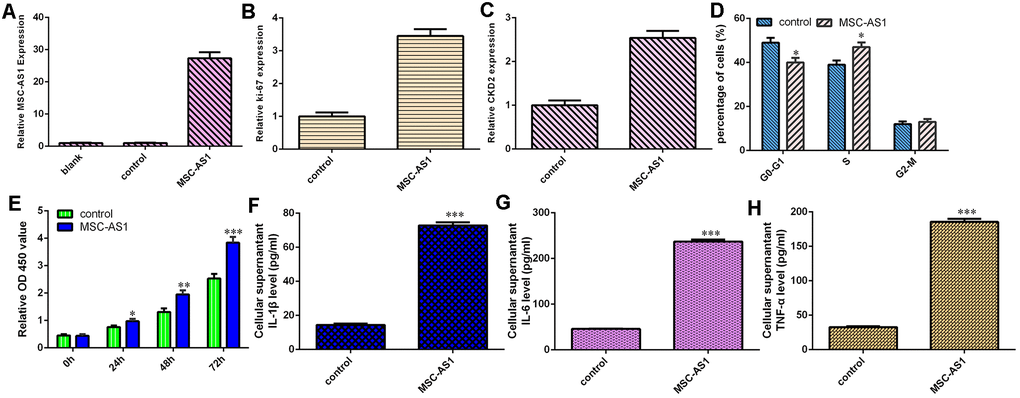
Figure 4. MSC-AS1 increased cell growth, cell cycle progression and inflammatory mediator secretion. (A) The expression of MSC-AS1 was detected by qRT-PCR analysis. (B) The expression of Ki-67 was measured using qRT-PCR analysis. (C) The level of CKD2 was detected by qRT-PCR analysis. (D) MSC-AS1 overexpression induced cell cycle progression in HGC-27 cells. (E) MSC-AS1 overexpression increased HGC-27 cell growth, based on CCK-8 analysis. (F) MSC-AS1 overexpression enhanced IL-1β secretion. (G) MSC-AS1 overexpression promoted IL-6 secretion. (H) Elevated expression of MSC-AS1 increased the secretion of TNF-α. GAPDH was used as the internal control. *p<0.05, **p<0.01 and ***p<0.001.
MSC-AS1 modulated miR-142-5p/DDX5 expression
By utilizing the online tool TargetScan, DDX5 was predicted to be a potential target gene of miR-142-5p (Figure 5A). DDX5 was significantly overexpressed in HGC-27 cells after treatment with the DDX5 mimic (Figure 5B). Overexpression of miR-142-5p suppressed the luciferase activity of wild-type DDX5, but the luciferase activity of mutant DDX5 was not changed (Figure 5C). Moreover, DDX5 and miR-142-5p were enriched in the Ago2-containing beads compared with the input, according to the RIP method (Figure 5D).
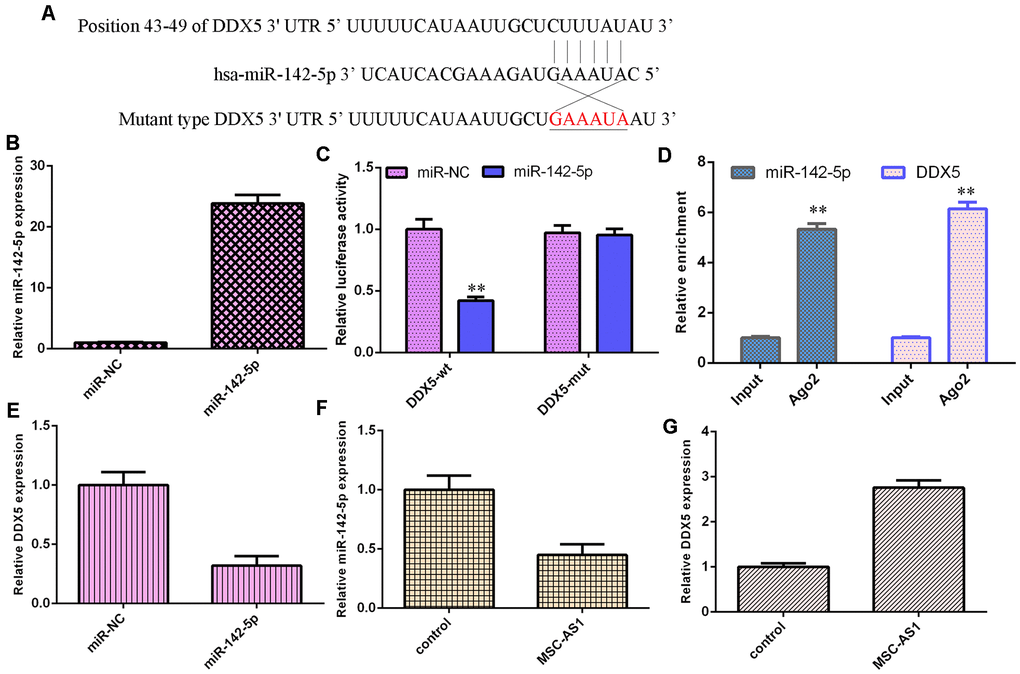
Figure 5. MSC-AS1 modulated miR-142-5p/DDX5 expression. (A) By utilizing an online tool, TargetScan, DDX5 was predicted to be a potential target gene of miR-142-5p. (B) The expression of DDX5 was measured by using qRT-PCR analysis. (C) Overexpression of miR-142-5p suppressed the luciferase activity of wild-type DDX5, but the luciferase activity of mutant DDX5 was not changed. (D) DDX5 and miR-142-5p were enriched in the Ago2-containing beads compared with the input, according to the RIP method. (E) miR-142-5p overexpression decreased DDX5 expression. (F) MSC-AS1 overexpression inhibited miR-142-5p expression. (G) The expression of DDX5 was measured by qRT-PCR assay. **p<0.01.
miR-142-5p overexpression decreased DDX5 expression (Figure 5E). MSC-AS1 overexpression inhibited miR-142-5p expression (Figure 5F) and enhanced DDX5 expression in HGC-27 cells (Figure 5G).
MSC-AS1 induced cell growth, cell cycle progression and inflammatory mediator secretion by modulating DDX5
The level of DDX5 was significantly decreased in HGC-27 cells after treatment with DDX5 siRNA (Figure 6A). Knockdown of DDX5 suppressed the expression of Ki-67 (Figure 6B) and CKD2 (Figure 6C) in MSC-AS1-overexpressing HGC-27 cells. Inhibition of DDX5 expression decreased the cell cycle (Figure 6D) and growth (Figure 6F) of MSC-AS1-overexpressing HGC-27 cells. Knockdown of DDX5 suppressed the secretion of the inflammatory mediators IL-1β, IL-6 and TNF-α (Figure 6F–6H).
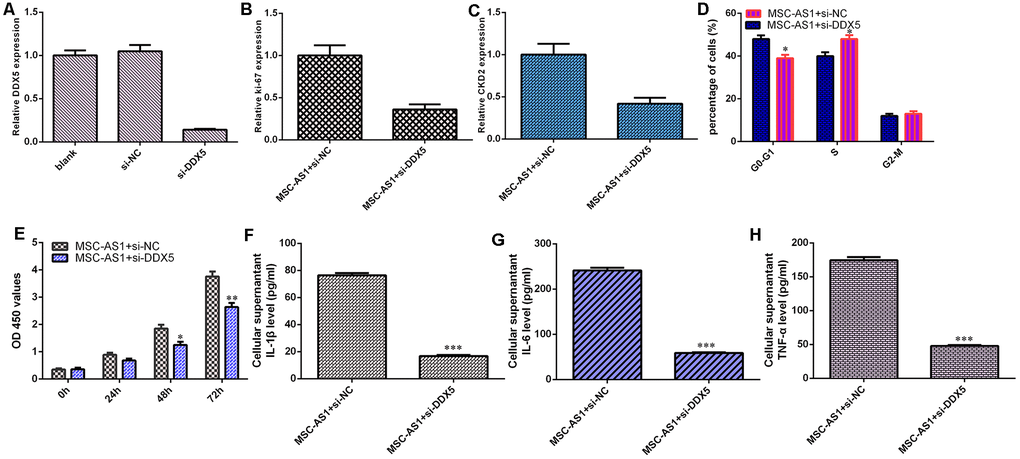
Figure 6. MSC-AS1 induced cell growth, cycle and inflammatory mediators secretion via modulating DDX5. (A) The level of DDX5 was measured by qRT-PCR analysis. (B) The expression of ki-67 was measured by qRT-PCR assay. (C) The expression of CKD2 was detected by qRT-PCR assay. (D) Inhibition expression of DDX5 decreased cell cycle in MSC-AS1-overexpressing HGC-27 cell. (E) Cell proliferation was measured by analyzed with CCK-8 assay. (F) The expression of IL-1β was measured by ELISA. (G) The expression of IL-6 was measured by ELISA. (H) The expression of IL-6 was detected by ELISA. *p<0.05, **p<0.01 and ***p<0.001.
Discussion
Compelling studies have observed that lncRNAs participate in many cell biological processes, including carcinogenesis and development [32–34]. For instance, Sun et al [35]. proved that lncRNA LATS2-AS1-001 suppressed GC development by modulating the YAP1/LATS2 pathway by binding to EZH2. Sun et al. indicated that AK025387 promoted cell invasion and migration in GC [36]. Gao et al. observed that NEAT1 induced GC development by modulating the miR-365a-3p/ABCC4 axis. Zhang et al [37]. showed that NNT-AS1 knockdown suppressed GC development by regulating the miR-142-5p/Wnt/β-catenin/SOX4 signaling pathway. Li et al [38]. showed that IGF2-AS enhanced GC cell migration, invasion and growth by regulating the EZH2/miR-937 axis. Recently, a new lncRNA, MSC-AS1, was identified as a crucial modulator in the development of tumors [26–30]. Cao et al [31]. Found that MSC-AS1 enhanced hepatocellular carcinoma progression by enhancing PGK1 expression. Yao et al [30]. showed that MSC-AS1 increased nasopharyngeal carcinoma development by regulating miR-524-5p/NR4A2. Hu et al [28]. showed that MSC-AS1 regulated renal carcinoma cell migration and growth by modulating the miR-3924/WNT5A/Wnt/β-catenin axis. In the present study, we conducted tests to examine the functional role of MSC-AS1 in GC. We first showed that MSC-AS1 was overexpressed in GC cells (HGC-27, MKN-45, SGC-7901 and MGC-803 cells) compared to GES cells. We observed that MSC-AS1 was upregulated in GC specimens compared with paired normal specimens. MSC-AS1 increased cell growth and cell cycle progression. Moreover, the overexpression of MSC-AS1 enhanced the secretion of the inflammatory mediators IL-1β, IL-6 and TNF-α.
Numerous studies have shown that lncRNAs regulate cell biological processes by sponging miRNA expression [39–41]. For example, Deng et al [42]. showed that DLGAP1-AS1 induced GC aggressiveness by sponging miR-628-5p and regulating the miR-628-5p/AEG-1 axis. Liu et al [43]. showed that SNHG1 enhanced GC cell EMT progression via modulation of the miR-15b/Notch1/DCLK1 axis. Liu et al. proved that TONSL-AS1 modulated CDK1/miR-490-3p to regulate ovarian carcinoma cell growth [44]. Liu et al [45]. indicated that HNF1A-AS1 induced GC metastasis, angiogenesis, invasion and lymphangiogenesis by sponging miR-30b-3p. Shi et al [46]. demonstrated that OIP5-AS1 induced hepatocellular carcinoma angiogenesis, cell migration and cell growth by modulating miR-3163/VEGFA. Furthermore, Zhang et al. showed that knockdown of MSC-AS1 induced sensitivity to cisplatin and suppressed the development of osteosarcoma by sponging miR-142 [29]. We also showed that MSC-AS1 overexpression inhibited miR-142-5p expression in HGC-27 cells. We observed that DDK5 was a target gene of miR-142-5p. The overexpression of miR-142-5p suppressed the luciferase activity of wild-type DDX5, but the luciferase activity of mutant DDX5 was not changed. We showed that miR-142-5p was downregulated in GC specimens compared to paired normal specimens. MSC-AS1 expression was inversely correlated with miR-142-5p expression in GC specimens. MSC-AS1 induced cell growth, cell cycle progression and inflammatory mediator secretion by modulating DDX5.
In summary, we observed that MSC-AS1 was overexpressed in GC cells and specimens and that ectopic expression of MSC-AS1 enhanced cell growth, cell cycle progression and inflammatory mediator secretion by modulating miR-142-5p/DDX5. These results showed that MSC-AS1 acts as a key oncogene in the development of GC.
Materials and Methods
Clinical specimens and cell transfection
A total of forty pairs of GC specimens and control specimens were acquired from GC patients who underwent surgery at The Fourth Hospital of China Medical University (Liaoning, Shengyang). Cell lines (HGC-27, MKN-45, SGC-7901, MGC-803 and GES cells) were acquired from ATCC, USA and cultured in DMEM (Gibco, BRL, UK) supplemented with streptomycin/penicillin and FBS (Gibco, BRL, UK). miR-142-5p mimic, pcDNA-MSC-AS1, siRNA-DDX5 and their controls (20 nM) were obtained from Shanghai GenePharma. Cell transfections were carried out with a Lipofectamine kit (Invitrogen, CA, USA).
RT-qPCR
RNA was extracted from GC cells or specimens using a TRIzol kit following the manufacturer’s protocol (Invitrogen, CA, USA). We utilized SYBR Green reagent (NEWBio) to study miR-142-5p, DDX5 and MSC-AS1 expression with the CFX96 system (VisonBio Scientific). The data were normalized to GAPDH or U6. The 2-ΔΔCt method was carried out to calculate relative fold changes. Primer sequences were noted: MSC-AS1, F, TCAAG AAATG GTGGC TAT and R, GCTCT GAGAC TGGCT GAA; miR-142-5p, F, TCAAG AAATG GTGGC TAT and R, CATAA AGTAG AAAGC ACTACT; U6, F, GCTTC GGCAG CACAT ATACT AAAAT and R, CGCTT CACGA ATTTG CGTGT CAT; GAPDH, F, GTCAA CGGAT TTGGT CTGTA TT and R, AGTCT TCTGG GTGGC AGTGAT.
Luciferase assays
The DDX5 3’-UTR and mutated DDX5 3’-UTR were cloned into the pGL3 plasmid as wild-type or mutant type 3’-UTRs, respectively. GC cells were treated with the wild-type or mutant DDX5 3’-UTR together with miR-142-5p scramble or mimic using a Lipofectamine kit. After transfection for 2 days, the luciferase activity was determined by Dual-Glo luciferase analysis (Promega, WI, USA).
CCK-8 assay, cell cycle analysis and ELISA
Cell growth was detected with a CCK-8 assay kit (Dojindo, Japan) according to the manufacturer’s instructions. These cells were plated in 96-well plates and cultured for 0, 24, 48 and 72 hours. Ten microliters of CCK-8 were added to each well, and the cells were cultured for an additional 3 hours. The absorbance was detected at 450 nm at different time points. To analyze cell cycle progression, GC cells were stained with cell cycle reagent (Thermo) following a standard protocol. The cell cycle was measured with flow cytometry on a Beckman flow cytometer (Dickinson, USA). The protein levels of IL-1β, IL-6 and TNF-α in the cell suspension were detected by ELISA following the manufacturer’s protocol.
RNA Immunoprecipitation (RIP) analysis
The RIP assay was conducted utilizing the Magna RNA-Binding Protein Immunoprecipitation of the RIP Kit (Millipore) following standard instructions. The cells were harvested and then lysed in RIP lysis buffer containing RNase and protease inhibitor, and then, the lysates were treated for 2 hours with buffer containing magnetic beads coated with antibodies against Ago2. IgG served as the negative control. Coprecipitated RNAs were determined by RT-qPCR assay.
Statistical analysis
All the statistical assays were analyzed using SPSS 19.0 (Chicago, IL, USA), and the graphs were generated by Prism 5.0. Student’s t-test was used to compare significant differences between two groups, and the correlation between miR-142-5p and MSC-AS1 in GC was analyzed by Pearson correlation assay. p<0.05 was defined as statistically significant.
Author Contributions
Yan Liu, Lin Li, Xiaoxu Wu, Haiyan Qi, Yan Gao, Yanqi Li, Da Chen collected the related paper, drafted and wrote the manuscript. Yan Liu Yanqi Li, Da Chen and participated in the design of experiments. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Editorial Note
This corresponding author has a verified history of publications using a personal email address for correspondence
References
- 1. Cao C, Xu Y, Du K, Mi C, Yang C, Xiang L, Xie Y, Liu W. LINC01303 functions as a competing endogenous RNA to regulate EZH2 expression by sponging miR-101-3p in gastric cancer. J Cell Mol Med. 2019; 23:7342–48. https://doi.org/10.1111/jcmm.14593 [PubMed]
- 2. Zhou DD, Liu XF, Lu CW, Pant OP, Liu XD. Long non-coding RNA PVT1: emerging biomarker in digestive system cancer. Cell Prolif. 2017; 50:e12398. https://doi.org/10.1111/cpr.12398 [PubMed]
- 3. Liu XF, Hao JL, Xie T, Pant OP, Lu CB, Lu CW, Zhou DD. The BRAF activated non-coding RNA: a pivotal long non-coding RNA in human Malignancies. Cell Prolif. 2018; 51:e12449. https://doi.org/10.1111/cpr.12449 [PubMed]
- 4. Li Z, Shen J, Chan MT, Wu WK. The long non-coding RNA SPRY4-IT1: an emerging player in tumorigenesis and osteosarcoma. Cell Prolif. 2018; 51:e12446. https://doi.org/10.1111/cpr.12446 [PubMed]
- 5. Zhuang K, Han K, Tang H, Yin X, Zhang J, Zhang X, Zhang L. Up-regulation of plasma miR-23b is associated with poor prognosis of gastric cancer. Med Sci Monit. 2016; 22:356–61. https://doi.org/10.12659/msm.895428 [PubMed]
- 6. Zhu W, Zhou K, Zha Y, Chen D, He J, Ma H, Liu X, Le H, Zhang Y. Diagnostic value of serum miR-182, miR-183, miR-210, and miR-126 levels in patients with early-stage non-small cell lung cancer. PLoS One. 2016; 11:e0153046. https://doi.org/10.1371/journal.pone.0153046 [PubMed]
- 7. Zhu M, Zhou X, Du Y, Huang Z, Zhu J, Xu J, Cheng G, Shu Y, Liu P, Zhu W, Wang T. miR-20a induces cisplatin resistance of a human gastric cancer cell line via targeting CYLD. Mol Med Rep. 2016; 14:1742–50. https://doi.org/10.3892/mmr.2016.5413 [PubMed]
- 8. Zhou Q, Zheng X, Chen L, Xu B, Yang X, Jiang J, Wu C. Smad2/3/4 pathway contributes to TGF-β-induced MiRNA-181b expression to promote gastric cancer metastasis by targeting Timp3. Cell Physiol Biochem. 2016; 39:453–66. https://doi.org/10.1159/000445638 [PubMed]
- 9. Zhou N, Qu Y, Xu C, Tang Y. Upregulation of microRNA-375 increases the cisplatin-sensitivity of human gastric cancer cells by regulating ERBB2. Exp Ther Med. 2016; 11:625–30. https://doi.org/10.3892/etm.2015.2920 [PubMed]
- 10. Zheng L, Jiao W, Mei H, Song H, Li D, Xiang X, Chen Y, Yang F, Li H, Huang K, Tong Q. miRNA-337-3p inhibits gastric cancer progression through repressing myeloid zinc finger 1-facilitated expression of matrix metalloproteinase 14. Oncotarget. 2016; 7:40314–28. https://doi.org/10.18632/oncotarget.9739 [PubMed]
- 11. Zheng H, Zhang F, Lin X, Huang C, Zhang Y, Li Y, Lin J, Chen W, Lin X. MicroRNA-1225-5p inhibits proliferation and metastasis of gastric carcinoma through repressing insulin receptor substrate-1 and activation of β-catenin signaling. Oncotarget. 2016; 7:4647–63. https://doi.org/10.18632/oncotarget.6615 [PubMed]
- 12. Zhao J, Nie Y, Wang H, Lin Y. MiR-181a suppresses autophagy and sensitizes gastric cancer cells to cisplatin. Gene. 2016; 576:828–33. https://doi.org/10.1016/j.gene.2015.11.013 [PubMed]
- 13. Zhang Y, Peng Z, Zhao Y, Chen L. microRNA-25 inhibits cell apoptosis of human gastric adenocarcinoma cell line AGS via regulating CCNE1 and MYC. Med Sci Monit. 2016; 22:1415–20. https://doi.org/10.12659/msm.896118 [PubMed]
- 14. Zhang R, Li F, Wang W, Wang X, Li S, Liu J. The effect of antisense inhibitor of miRNA 106b~25 on the proliferation, invasion, migration, and apoptosis of gastric cancer cell. Tumour Biol. 2016; 37:10507–15. https://doi.org/10.1007/s13277-016-4937-x [PubMed]
- 15. Zhang M, Dong BB, Lu M, Zheng MJ, Chen H, Ding JZ, Xu AM, Xu YH. miR-429 functions as a tumor suppressor by targeting FSCN1 in gastric cancer cells. Onco Targets Ther. 2016; 9:1123–33. https://doi.org/10.2147/OTT.S91879 [PubMed]
- 16. Cui Z, Luo Z, Lin Z, Shi L, Hong Y, Yan C. Long non-coding RNA TTN-AS1 facilitates tumorigenesis of papillary thyroid cancer through modulating the miR-153-3p/ZNRF2 axis. J Gene Med. 2019; 21:e3083. https://doi.org/10.1002/jgm.3083 [PubMed]
- 17. Hu R, Zhu Z. ELK1-activated GPC3-AS1/GPC3 axis promotes the proliferation and migration of cervical cancer cells. J Gene Med. 2019; 21:e3099. https://doi.org/10.1002/jgm.3099 [PubMed]
- 18. Refai NS, Louka ML, Halim HY, Montasser I. Long non-coding RNAs (CASC2 and TUG1) in hepatocellular carcinoma: clinical significance. J Gene Med. 2019; 21:e3112. https://doi.org/10.1002/jgm.3112 [PubMed]
- 19. Li Z, Li X, Chen X, Li S, Ho IH, Liu X, Chan MT, Wu WK. Emerging roles of long non-coding RNAs in neuropathic pain. Cell Prolif. 2019; 52:e12528. https://doi.org/10.1111/cpr.12528 [PubMed]
- 20. Zou Y, Zhong Y, Wu J, Xiao H, Zhang X, Liao X, Li J, Mao X, Liu Y, Zhang F. Long non-coding PANDAR as a novel biomarker in human cancer: a systematic review. Cell Prolif. 2018; 51:e12422. https://doi.org/10.1111/cpr.12422 [PubMed]
- 21. Zhu S, Fu W, Zhang L, Fu K, Hu J, Jia W, Liu G. LINC00473 antagonizes the tumour suppressor miR-195 to mediate the pathogenesis of Wilms tumour via IKKα. Cell Prolif. 2018; 51:e12416. https://doi.org/10.1111/cpr.12416 [PubMed]
- 22. Zhao J, Zhang C, Gao Z, Wu H, Gu R, Jiang R. Long non-coding RNA ASBEL promotes osteosarcoma cell proliferation, migration, and invasion by regulating microRNA-21. J Cell Biochem. 2018; 119:6461–69. https://doi.org/10.1002/jcb.26671 [PubMed]
- 23. Yu X, Zheng H, Tse G, Zhang L, Wu WK. CASC2: an emerging tumour-suppressing long noncoding RNA in human cancers and melanoma. Cell Prolif. 2018; 51:e12506. https://doi.org/10.1111/cpr.12506 [PubMed]
- 24. Yu X, Zheng H, Tse G, Chan MT, Wu WK. Long non-coding RNAs in melanoma. Cell Prolif. 2018; 51:e12457. https://doi.org/10.1111/cpr.12457 [PubMed]
- 25. Xu R, Feng F, Yu X, Liu Z, Lao L. LncRNA SNHG4 promotes tumour growth by sponging miR-224-3p and predicts poor survival and recurrence in human osteosarcoma. Cell Prolif. 2018; 51:e12515. https://doi.org/10.1111/cpr.12515 [PubMed]
- 26. Sun Y, Wang P, Yang W, Shan Y, Zhang Q, Wu H. The role of lncRNA MSC-AS1/miR-29b-3p axis-mediated CDK14 modulation in pancreatic cancer proliferation and gemcitabine-induced apoptosis. Cancer Biol Ther. 2019; 20:729–39. https://doi.org/10.1080/15384047.2018.1529121 [PubMed]
- 27. Zhang N, Hu X, He S, Ding W, Wang F, Zhao Y, Huang Z. LncRNA MSC-AS1 promotes osteogenic differentiation and alleviates osteoporosis through sponging microRNA-140-5p to upregulate BMP2. Biochem Biophys Res Commun. 2019; 519:790–96. https://doi.org/10.1016/j.bbrc.2019.09.058 [PubMed]
- 28. Hu Z, Li L, Cheng P, Liu Q, Zheng X, Peng F, Zhang Q. lncRNA MSC-AS1 activates Wnt/β-catenin signaling pathway to modulate cell proliferation and migration in kidney renal clear cell carcinoma via miR-3924/WNT5A. J Cell Biochem. 2020; 121:4085–93. https://doi.org/10.1002/jcb.29594 [PubMed]
- 29. Zhang L, Zhao G, Ji S, Yuan Q, Zhou H. Downregulated long non-coding RNA MSC-AS1 inhibits osteosarcoma progression and increases sensitivity to cisplatin by binding to MicroRNA-142. Med Sci Monit. 2020; 26:e921594. https://doi.org/10.12659/MSM.921594 [PubMed]
- 30. Yao H, Yang L, Tian L, Guo Y, Li Y. LncRNA MSC-AS1 aggravates nasopharyngeal carcinoma progression by targeting miR-524-5p/nuclear receptor subfamily 4 group a member 2 (NR4A2). Cancer Cell Int. 2020; 20:138. https://doi.org/10.1186/s12935-020-01202-1 [PubMed]
- 31. Cao C, Zhong Q, Lu L, Huang B, Li J, Meng L, Wei H. Long noncoding RNA MSC-AS1 promotes hepatocellular carcinoma oncogenesis via inducing the expression of phosphoglycerate kinase 1. Cancer Med. 2020; 9:5174–84. https://doi.org/10.1002/cam4.3080 [PubMed]
- 32. Li Z, Li X, Chen C, Li S, Shen J, Tse G, Chan MTV, Wu WKK. Long non-coding RNAs in nucleus pulposus cell function and intervertebral disc degeneration. Cell Prolif. 2018; 51:e12483. https://doi.org/10.1111/cpr.12483 [PubMed]
- 33. Li Y, Zhou L, Lu C, Shen Q, Su Y, Zhi Z, Wu F, Zhang H, Wen Z, Chen G, Li H, Xia Y, Tang W. Long non-coding RNA FAL1 functions as a ceRNA to antagonize the effect of miR-637 on the down-regulation of AKT1 in Hirschsprung’s disease. Cell Prolif. 2018; 51:e12489. https://doi.org/10.1111/cpr.12489 [PubMed]
- 34. Meng L, Ma P, Cai R, Guan Q, Wang M, Jin B. Long noncoding RNA ZEB1-AS1 promotes the tumorigenesis of glioma cancer cells by modulating the miR-200c/141-ZEB1 axis. Am J Transl Res. 2018; 10:3395–412. [PubMed]
- 35. Sun D, Wang Y, Wang H, Xin Y. The novel long non-coding RNA LATS2-AS1-001 inhibits gastric cancer progression by regulating the LATS2/YAP1 signaling pathway via binding to EZH2. Cancer Cell Int. 2020; 20:204. https://doi.org/10.1186/s12935-020-01285-w [PubMed]
- 36. Sun YY, Zhang H, Ma RR, Zhang GH, Tian YR, Liu L, Liu L, Gao P. Long non-coding RNA AK025387 promotes cell migration and invasion of gastric cancer. Front Oncol. 2020; 10:633. https://doi.org/10.3389/fonc.2020.00633 [PubMed]
- 37. Zhang J, Zhang K, Hou Y. Long non-coding RNA NNT-AS1 knockdown represses the progression of gastric cancer via modulating the miR-142-5p/SOX4/Wnt/β-catenin signaling pathway. Mol Med Rep. 2020; 22:687–96. https://doi.org/10.3892/mmr.2020.11158 [PubMed]
- 38. Li Z, Li Z, Zhong Z, Zhou J, Huang S, Zhou W, Xu J. Retracted: lncRNA IGF2-AS Promotes Cell Proliferation, Migration, and Invasion of Gastric Cancer by Modulating miR-937/EZH2 Axis. Cancer Biother Radiopharm. 2020. [Epub ahead of print]. https://doi.org/10.1089/cbr.2019.3275 [PubMed]
- 39. Tian C, Deng Y, Jin Y, Shi S, Bi H. Long non-coding RNA RNCR3 promotes prostate cancer progression through targeting miR-185-5p. Am J Transl Res. 2018; 10:1562–70. [PubMed]
- 40. Zhang XY, Tang XY, Ma LJ, Guo YL, Li XS, Zhao LM, Tian CJ, Cheng DJ, Chen ZC, Zhang LX. Schisandrin B down-regulated lncRNA BCYRN1 expression of airway smooth muscle cells by improving miR-150 expression to inhibit the proliferation and migration of ASMC in asthmatic rats. Cell Prolif. 2017; 50:e12382. https://doi.org/10.1111/cpr.12382 [PubMed]
- 41. Yu N, Liang Y, Zhu H, Mo H, Pei H. CsA promotes XIST expression to regulate human trophoblast cells proliferation and invasion through miR-144/titin axis. J Cell Biochem. 2017; 118:2208–18. https://doi.org/10.1002/jcb.25867 [PubMed]
- 42. Deng J, Zhang Q, Lu L, Fan C. Long noncoding RNA DLGAP1-AS1 promotes the aggressive behavior of gastric cancer by acting as a ceRNA for microRNA-628-5p and raising astrocyte elevated gene 1 expression. Cancer Manag Res. 2020; 12:2947–60. https://doi.org/10.2147/CMAR.S246166 [PubMed]
- 43. Liu ZQ, He WF, Wu YJ, Zhao SL, Wang L, Ouyang YY, Tang SY. LncRNA SNHG1 promotes EMT process in gastric cancer cells through regulation of the miR-15b/DCLK1/Notch1 axis. BMC Gastroenterol. 2020; 20:156. https://doi.org/10.1186/s12876-020-01272-5 [PubMed]
- 44. Liu Y, Li L, Wang X, Wang P, Wang Z. LncRNA TONSL-AS1 regulates miR-490-3p/CDK1 to affect ovarian epithelial carcinoma cell proliferation. J Ovarian Res. 2020; 13:60. https://doi.org/10.1186/s13048-020-00657-0 [PubMed]
- 45. Liu HT, Ma RR, Lv BB, Zhang H, Shi DB, Guo XY, Zhang GH, Gao P. LncRNA-HNF1A-AS1 functions as a competing endogenous RNA to activate PI3K/AKT signalling pathway by sponging miR-30b-3p in gastric cancer. Br J Cancer. 2020; 122:1825–36. https://doi.org/10.1038/s41416-020-0836-4 [PubMed]
- 46. Shi C, Yang Q, Pan S, Lin X, Xu G, Luo Y, Zheng B, Xie X, Yu M. LncRNA OIP5-AS1 promotes cell proliferation and migration and induces angiogenesis via regulating miR-3163/VEGFA in hepatocellular carcinoma. Cancer Biol Ther. 2020; 21:604–14. https://doi.org/10.1080/15384047.2020.1738908 [PubMed]