



Introduction
Carnitine, a water-soluble quaternary amine (3-hydroxy-4-N, N, N-trimethyl amino butyric acid), is synthesized from the essential amino acids lysine and methionine [1, 2]. Cells utilize L-carnitine for transporting long chain fatty acids into mitochondria for β-oxidation [3]. L-carnitine is essential for lipid metabolism in all cell types and highly enriched in skeletal and cardiac muscle [4]. Various forms of carnitine, including L-carnitine, acetyl-L-carnitine, and propionyl-L-carnitine, have long been used as dietary supplements for weight loss and enhancing performance [5]. Impaired carnitine uptake due to genetic mutations in the carnitine-transporters can cause cardiomyopathy, skeletal-muscle weakness, and hypoglycemia [6–8]. Certain conditions such as chronic renal failure and antibiotics abuse can also reduce carnitine retention in the body, manifesting symptoms of carnitine deficiency [8, 9]. Carnitine has been approved by the Food and Drug Administration (FDA) to treat primary and certain secondary carnitine-deficiency syndromes [10].
Carnitine levels decrease during aging, which may contribute to the decline in mitochondrial function and aging [11]. In rats, dietary supplementation with high doses of acetyl-L-carnitine and alpha-lipoic acid improves mitochondrial function and decreases oxidative stress, resulting in improved performance on memory-requiring tasks [12–14]. In humans, several studies suggest that carnitine could improve mental health in older adults with mild cognitive impairment or Alzheimer’s disease [15]. Carnitine may also improve other age-associated diseases such as cardiovascular diseases [16–19], atherosclerosis [20], neurometabolic disorders [21] and diabetic symptoms [22–25]. In addition, carnitine has also been used in adjuvant therapy to treat conditions such ashemodialysis [26]. Acetyl-L-carnitine can extend chronological lifespan of budding yeast [27, 28]. Most of the therapeutic effect of carnitine have been linked to cellular redox balance [29, 30]. However, the detailed mechanisms of action at the cellular and molecular levels have not been fully investigated.
The round worms C. elegans has been widely used to study oxidative stress response and aging [31, 32]. In response to chemicals that generate reactive oxygen species (ROS) such as paraquat or juglone, C. elegans activates SKN-1 and DAF-16 dependent transcription programs to alleviate oxidative damage, which leads to lifespan extension [33, 34]. Activation of the SKN-1 pathway contributes to the extended lifespan of glp-1 mutant lacking germline and daf-2 mutant with impaired insulin receptor [35, 36]. The roles of SKN-1 and DAF-16 are conserved in mammals (Nrf2 and FOXO3, respectively), providing useful targets for disease intervention [37]. C. elegans has also been used for study human Aβ toxicity, in which ectopic expression of human Aβ causes paralysis [38]. Very few studies have been carried out in C. elegans regarding carnitine metabolism and function. In one study, L-carnitine supplement in the medium can prevent glucose toxicity on C. elegans survival [39]. In another study, feeding worms with acetyl-carnitine reduces age-related neuronal damage and improves learning behavior [40]. Whether carnitine has a role in OSR and aging in C. elegans has not been reported.
In this study, we show that carnitine supplement in the medium facilitates recovery from oxidative stress during aging, promoting resistance to oxidative toxicity induced by H2O2 and human amyloid protein Aβ(1–42) aggregates. Carnitine’s beneficial effect is dependent on SKN-1, DAF-16 and a potential carnitine transporter T08B1.1. T08B1.1 expression is increased in the longevity mutant glp-1 and required for the long lifespan. Our study is the first of its kind to report the effect of carnitine on aging in C. elegans and reveal novel roles of carnitine in extending lifespan.
Results
L-carnitine shortens the length of oxidative stress response (OSR)
It has been shown that L-carnitine has antioxidant activity in vitro [41]. By using paraquat, a well-known ROS generator affecting mitochondrial respiration and dynamics [42, 43], we tested if L-carnitine would have antioxidant activity in vivo in C. elegans. We raised the animals on agar plate containing nematode growth (NG) medium supplemented with and without 100 μM of L-carnitine from L1 stage to L4 stage, then oxidatively stressed the animals with 1mM paraquat for 24 hours on NGM plate. For convenience, we examined the OSR marker gst-4::gfp [44]. We found strong activation of gst-4:gfp by paraquat treatment, indicating a robust oxidative stress condition. L-carnitine supplementation did not significant affect gst-4:gfp induction by paraquat (Figure 1A, 1B). However, animals treated with L-carnitine had a better recovery from paraquat, as shown by a faster clearance of the OSR marker gst-4::gfp after transferring to paraquat-free medium (Figure 1C, 1D). To confirm these results, we also used another ROS generating compound juglone [45]. Similarly, gst-4::gfp levels decreased faster in animals treated with L-carnitine compared with non-treated animals (Figure 1E), suggesting that L-carnitine promotes oxidative stress recovery in C. elegans.
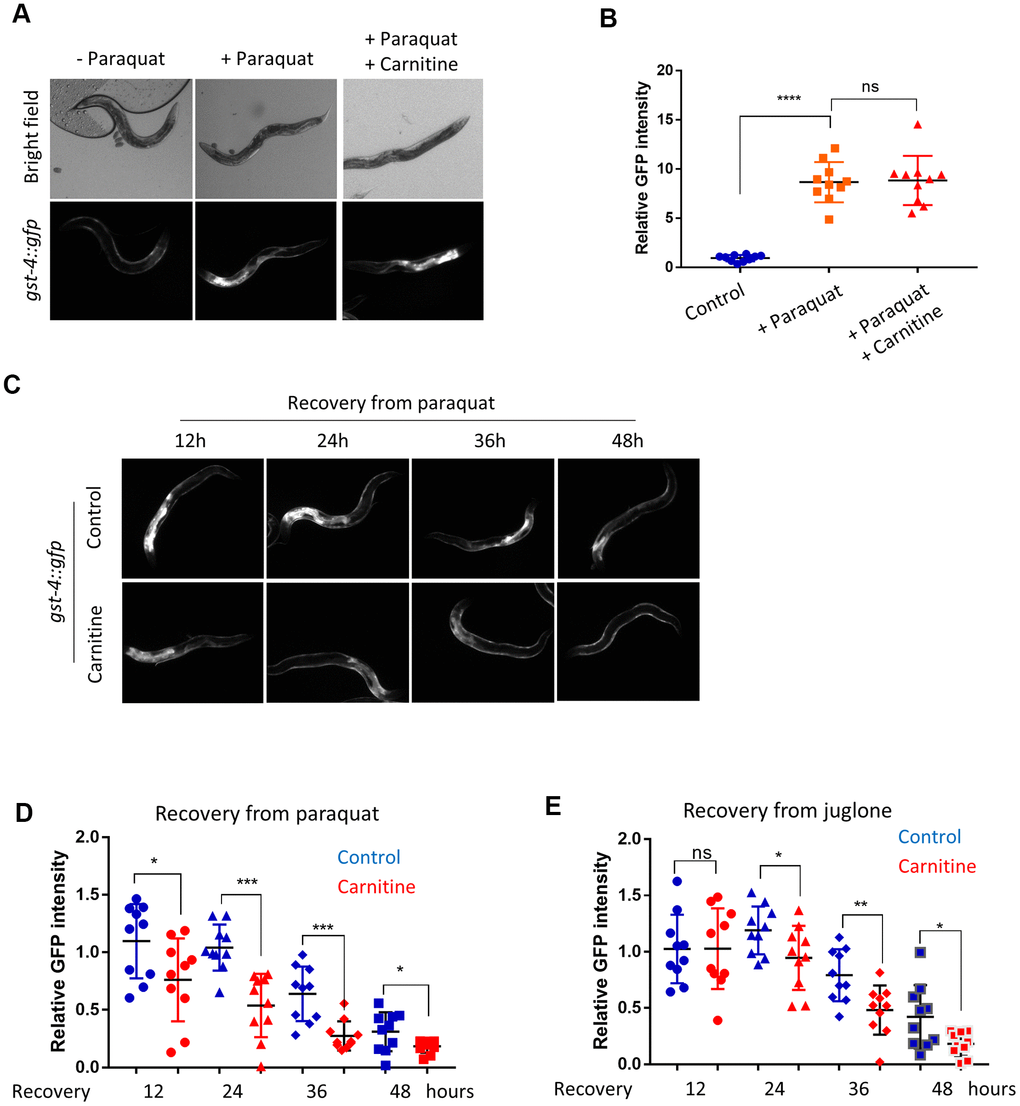
Figure 1. L-carnitine shortens the length of oxidative stress response (OSR) induced by paraquat and juglone. (A) L-carnitine did not affect induction of OSR. C. elegans expressing the OSR marker gst-4::gfp were synchronized at L1 larvae stage and raised on NG medium supplemented with or without 10 μM L-carnitine to L4/young adult stage. The ROS generator paraquat was then added to the medium to the final concentration of 1 mM. After 24 hours, animals were imaged with a fluorescent microscope. Representative images were shown. (B) Quantification of images taken from at least 3 independent experiments in (A) by ImageJ and relative expression levels were plotted. Statistical analysis was performed by two-tailed, unpaired student’s t-test (ns, not significant. ****, P<0.0001). Error bars indicate the standard deviation of the mean. (C) L-carnitine reduced the gst-4::gfp expression during recovery from oxidative stress. C. elegans worms were prepared and treated as in (A). After 24 hours of paraquat treatment, worms were transferred to fresh plate. OSR marker gst-4::gfp were examined at indicated time. (D) Quantification of gst-4::gfp expression in (C). Images from 3 independent experiments were quantified using ImageJ and normalized to the value at time 0. Statistical analysis was performed by two-tailed, unpaired student’s t-test (*, P<0.05. ***, P<0.001). Error bars indicate standard deviation of the mean. (E) L-carnitine reduced gst-4::gfp expression during recovery from juglone treatment. C. elegans worms were prepared and treated as in (C) except that 300 μM juglone (final concentration) was added to the NG medium. Images from 2 independent experiments were quantified using ImageJ and normalized to the average at 12 hours. Statistical analysis was performed by two-tailed, unpaired student’s t-test (ns, not significant. *, P<0.05. **, P<0.001). Error bars indicate standard deviation of the mean.
L-carnitine promotes the recovery from oxidative stress
To further confirm the new finding that L-carnitine improves recovery from oxidative stress, we directly measured the ROS levels. Animals raised with and without 100 μM L-carnitine supplement from L1 stage to L4 stage were challenged with 1mM paraquat for 24 hours, then transferred to paraquat-free medium plate. After 12, 24, 36 and 48 hours, animals were stained with Dihydroethidium (DHE), a widely used ROS indicator [35, 46]. The results showed that L-carnitine treatment robustly reduced the ROS levels in the intestine after 36 and 48 hours of recovery from paraquat (Figure 2A, 2B).
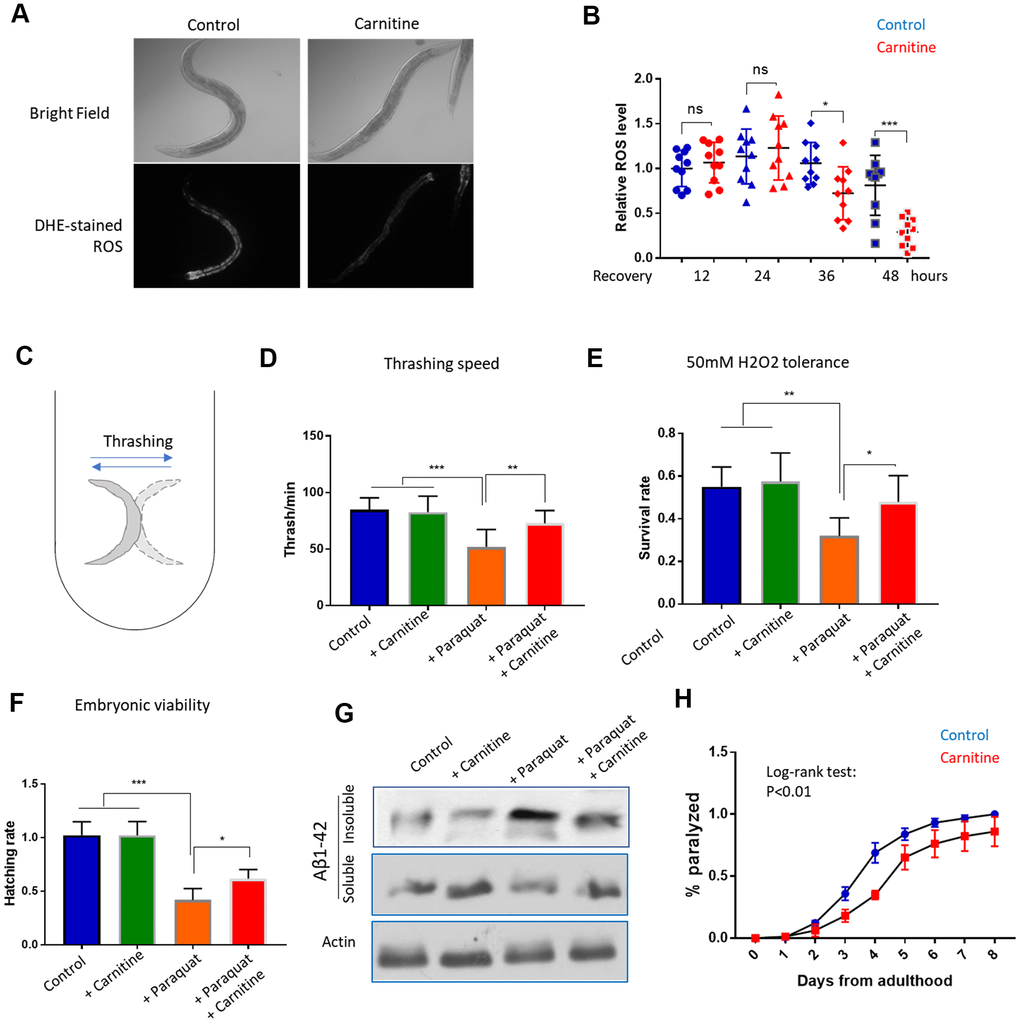
Figure 2. L-carnitine promotes recovery from oxidative stress induced by paraquat and juglone. (A) L-carnitine facilitated the clearance of ROS. N2 wild-type C. elegans were synchronized at L1 larvae stage and raised on NG medium supplemented with or without 10 μM L-carnitine to L4/young adult stage. The ROS generator paraquat was added to the medium to the final concentration of 1mM. After 24 hours, animals were transferred to new paraquat-free plate with and without L-carnitine for recovery. After recovery for 12, 24, 48 hours, worms were stained with ROS dye dihydroethidium (DHE) and imaged with fluorescent microscope. Representative images at 48-hour recovery were shown. (B) Quantification of DHE signal from at least 3 independent experiments in (A) by ImageJ and relative expression levels were plotted. Statistical analysis was performed by two-tailed, unpaired student’s t-test (ns, not significant. *, P<0.05. ***, P<0.001). Error bars indicate standard deviation of the mean. (C) Thrashing assay for C. elegans. Worms were pick from agar plate to 1 mL M9 buffer in 24-well plate and examined under dissecting microscope. Worms were moving left and right rapidly and the movement from one side to the other side then back to the original position was counted as 1 thrash. (D) L-carnitine mitigated the toxicity of paraquat on mobility. Worms were treated with paraquat and L-carnitine as in (A) and recovered for 48 hours. Trashing speed (thrash/min) under different treatments were measured for at least 3-independent experiments with 10 animals/experiment. Statistical analysis was performed by two-tailed, paired student’s t-test (**, P<0.01. ***, P<0.001). Error bars indicate standard deviation of the mean. (E) L-carnitine rescued the H2O2 hypersensitivity of paraquat-treated worms. Worms were treated with paraquat and L-carnitine as in (A) and recovered for 48 hours. Worms were then incubated in 50mM H2O2 for 1 hour. Data were pooled from 2 independent experiments and survival rates under different treatment were compared. Statistical analysis was performed by two-tailed, paired student’s t-test (*, P<0.05. **, P<0.01). Error bars indicate standard deviation of the mean. (F) L-carnitine mitigated the toxicity of paraquat on embryonic survival. Worms were treated with paraquat and L-carnitine as in (A) and recovered for 48 hours.10 worms were transferred to a new agar plate to allow egg laying for 2 hours and total number of eggs were counted. Hatching were examined after 24 hours. Experiments were conducted 2 times with 5 replicates each time. Data were normalized to control group for comparison. Statistical analysis was performed by two-tailed, unpaired student’s t-test (**, P<0.01. ***, P<0.001). Error bars indicate standard deviation of the mean. (G) L-carnitine mitigated paraquat-induced amyloid protein aggregation. Worms expressing human amyloid protein fragment Aβ(1-42) in body wall muscle (CL2006) were treated with paraquat and L-carnitine as in (A) and recovered for 48 hours. Worms were then homogenized in high-salt RAB buffer. Soluble and insoluble fraction were analyzed by western blot using anti-Aβ antibody. Total lysate was analyzed by western using anti-actin antibody. Shown was representative results of 2 independent experiments. (H) L-carnitine mitigated Aβ(1-42)-induced paralysis. Animals were raised at 25° C to young adult stage (day-0) and examined every day thereafter for paralysis. Data pooled from 2 independent experiments (n>120) were plotted. Log-rank test was performed (P<0.01). Error bar indicated the standard deviation of the mean.
Next, we tested multiple heath parameters related to oxidative stress. First, we examined if muscle strength could be improved by L-carnitine, by measuring the bending movement. When in liquid, the worms keep bending or thrashing (Figure 2C), which has been used to indicate the muscle strength and mobility [47]. By treating the animals with paraquat and L-carnitine as mentioned above, we measured the thrashing speed after 48 hours of recovery. The results showed that the impaired mobility by paraquat treatment could be largely rescued by L-carnitine (Figure 2D). L-carnitine did not obviously improve mobility under normal conditions. Second, we tested if the faster recovery from oxidative stress could result in a better tolerance to oxidative damage by H2O2. Similarly, animals treated with paraquat and L-carnitine were collected 48 hours after recovery from paraquat and incubated with 50mM of H2O2 for 1 hour. We found that paraquat-treated worms were sensitive to H2O2 toxicity, which was largely rescued by L-carnitine (Figure 2E). Third, we examined the survival rate of progenies. Consistently, egg hatching was reduced by paraquat, but such reduction was significantly mitigated by L-carnitine (Figure 2F). Fourth, we examined if human amyloid protein aggregation could be mitigated by supplementing L-carnitine in the medium. Amyloid-beta (Aβ1-42) has been known to aggregate upon oxidative stress or during aging [48, 49]. By separating the soluble and insoluble fractions of the whole worm lysate and western blotting, we found that L-carnitine reduced the paraquat-induced Aβ1-42 aggregation (Figure 2G). Interestingly, despite the lack of effect of L-carnitine on Aβ1-42 aggregates under normal culturing conditions (Figure 2G), the animals treated with L-carnitine slightly but significantly decreased amyloid-induced paralysis (Figure 2H). Together, these functional assays confirmed that L-carnitine reduced oxidative stress damage in C. elegans.
L-carnitine decreases oxidative stress during aging and increased lifespan in C. elegans
Since ROS accumulates during aging, L-carnitine could also improve age-related oxidative damage. To test this, C. elegans were cultured from L1 stage on NG medium supplemented with or without 100 μM L-carnitine and gst-4::gfp expression was examined at L4, day-2, day-4 and day-6 of adulthoods. Interestingly, gst-4::gfp was induced after reaching adulthood but gradually declined thereafter, consistent with a previous report [35]. Similar to that of paraquat treatment, aging-induced gst-4::gfp expression was decreased by L-carnitine during the recovery stage but not the induction stage (Figure 3A, 3B). Despite the decline in gst-4::gfp expression after day-4, DHE-stained ROS continued to accumulate from day-2 to day-10. Importantly, worms raised on L-carnitine appeared to have less ROS on day-6 and day-10 of adulthood, suggesting a better recovery from oxidative stress during aging (Figure 3C, 3D).
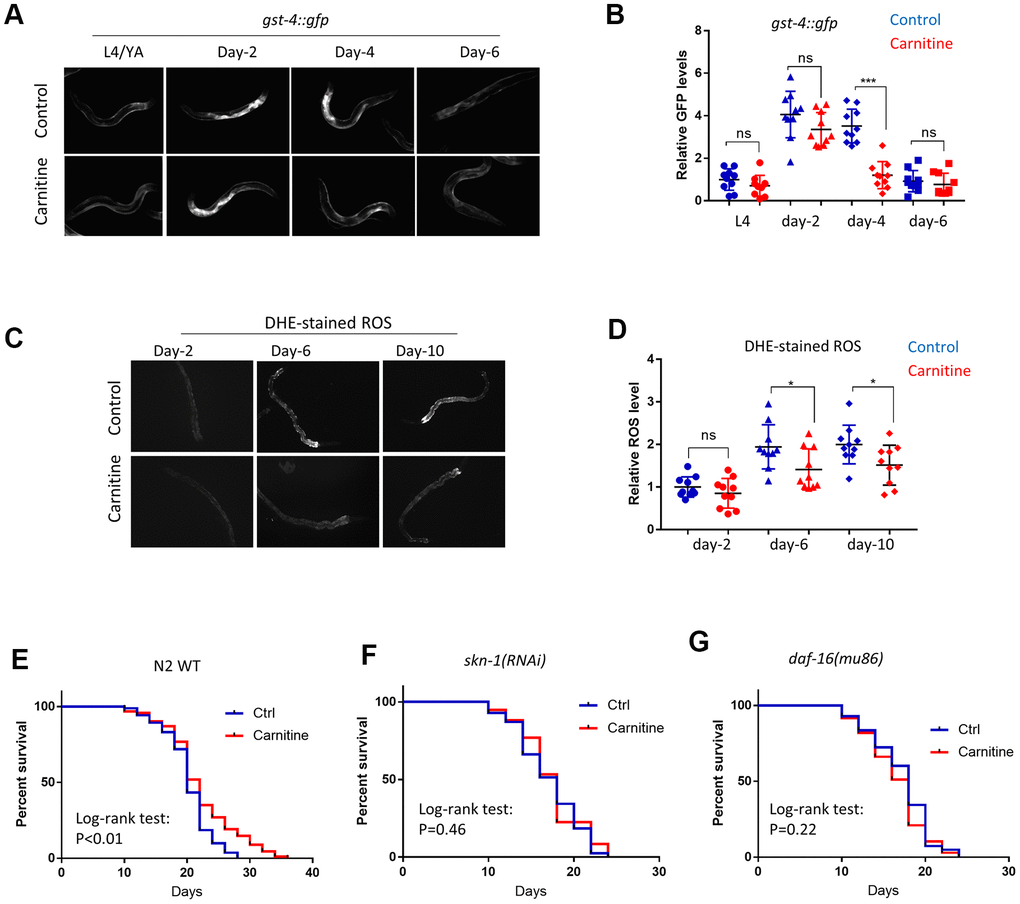
Figure 3. L-carnitine promotes oxidative stress recovery during aging and increased lifespan in C.elegans. (A) L-carnitine promoted recovery from oxidative stress during aging. C. elegans expressing the OSR marker gst-4::gfp were synchronized at L1 larvae stage and raised on NG medium supplemented with or without 10 μM L-carnitine. Worms were imaged with fluorescent microscope at L4/young adult (YA) stage, day-2, day-4 day-6 of adulthood. Representative images of 3 independent experiments were shown. (B) Quantification of images from experiment described in (A). Shown were gst-4::gfp levels normalized to the average of L4/young stage. Statistical analysis was performed by two tailed, unpaired student’s t-test (ns, not significant. ***, P<0.001). Error bars indicate standard deviation of the mean. (C) L-carnitine delayed the ROS accumulation in C. elegans during normal aging. Worms were treated as in (A) and stained with DHE dye at indicated time points, followed by imaging with fluorescent microscope. Representative images from 3 experiments were shown. (D) Quantification of DHE-stained ROS levels described in (C). Images from 3 experiments were quantified by ImageJ and statistically analyzed by two-tailed, unpaired student’s t-test (ns, not significant. *, P<0.05). Error bars indicate standard deviation of the mean. (E) L-carnitine prolonged the lifespan of wild-type N2 C. elegans. Wild-type C. elegans were synchronized at L1 larvae stage and raised on NG medium supplemented with or without 10 μM L-carnitine. 50 μM FUDR was added to prevent reproduction. Dead and viable worms were counted every 2 or 3 days starting from day-10 of adulthood. Experiments were performed for 2 times (n>120) and survival were analyzed by log-rank test (Supplementary Table 1). (F) L-carnitine did not extend lifespan of C. elegans with skn-1 knockdown. Experiments were performed similar to (E) except RNAi bacteria was used. Data were pooled from 2 experiments (n>120) and analyzed by log-rank test (Supplementary Table 1). (G) L-carnitine did not extend lifespan of C. elegans with daf-16 knockdown. Experiments were performed similar to (E) except RNAi bacteria was used. Data were pooled from 2 experiments (n>120) and analyzed with log-rank test (Supplementary Table 1).
We also tested if aging was delayed in C. elegans by supplementing L-carnitine in the medium. Animals were cultured under 20° C on NG medium containing various concentrations of L-carnitine throughout life. 50 μM 5’-Fluoro-2-deoxyuridine (FUDR) was added to inhibit progeny growth. The results showed that L-carnitine extended lifespan of C. elegans at 100, 200 and 500 but not 50 μM (Figure 3E and Supplementary Figure 1A). At 200 and 500 μM, L-carnitine causes slight developmental delay in some worms (Supplementary Figure 1B). We therefore chose 100 μM for all experiments in this study. SKN-1 and DAF-16 are transcription factors known to activate oxidative stress defensing programs and extend lifespan in C. elegans [50]. Both RNAi knockdown prevented L-carnitine from extending lifespan, suggesting that SKN-1 and DAF-16 are both needed for L-carnitine to extend lifespan in C. elegans (Figure 3F, 3G).
The long-lived mutants daf-2 and glp-1 recover better from oxidative stress than wild-type controls
The requirement for DAF-16 and SKN-1 for lifespan extension by L-carnitine prompted us to test the role of L-carnitine in long-lived mutants glp-1 and daf-2. These animals, similar to carnitine-treated animals, require both DAF-16 and SKN-1 for their long lifespan [36, 51]. First, we asked if glp-1(e2144) and daf-2(e2370) mutants would recover from oxidative stress better than wild-type controls. As reported earlier [35], gst-4::gfp expression was much higher in glp-1 mutant reaching adulthood (Figure 4A). To evaluate the recovery, we normalized gst-4::gfp expression levels at day-2 adulthood and examined the speed of decline in GFP intensity. Indeed, glp-1(e2144) showed a faster decline in gst-4::gfp expression than controls (Figure 4A, 4B). Similar results were obtained for daf-2(e2370) mutants (Figure 4B). We then asked if such improved recovery from OSR could lead to enhanced clearance of endogenous ROS levels. Indeed, the experiments showed that both glp-1(e2144) and daf-2(e2370) mutant worms accumulated ROS slower than wild-type control (Figure 4C), suggesting that the speed to recover from OSR plays an important role in aging.
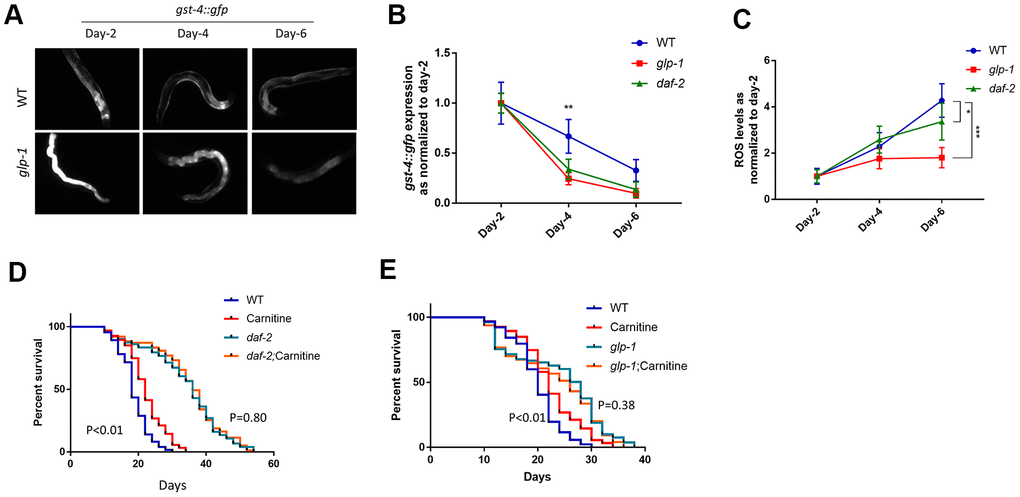
Figure 4. The long-lived mutants daf-2 and glp-1 recover from oxidative stress better than wild-type controls. (A) glp-1 worms showed faster decrease in the expression of the OSR marker gst-4::gfp. Age-matched, gst-4::gfp-expressing WT and glp-1 mutant worms were raised from L1 to L4 at 25° C to deplete germ cells in glp-1 then maintained at 20° C throughout the experiment. gst-4::gfp expression were examined with fluorescent microscope at indicated time points. Representative images were shown. (B) Data from 2 independent experiments described in (A) were normalized to the average of day-2 adulthood. Data were analyzed by two-tailed, paired student’s t-test (**, P<0.01). Error bars indicate standard deviation of the mean. (C) daf-2 worms recovered from oxidative stress faster than wild-type controls. Age-matched worms expressing gst-4::gfp were raised at 20° C. gst-4::gfp expression was examined at indicated time points. Two independent experiments were performed. Data were normalized to the average of day-2 adulthood and analyzed by two-tailed, paired student’s t-test (*, P<0.05. ***, P<0.001). Error bars indicate standard deviation of the mean. (D) L-carnitine did not further increase lifespan of daf-2C. elegans. Age-matched wild-type or daf-2 worms were raised at 20° C throughout life with and without 10 μM L-carnitine supplement. Dead and live worms were counted every 2 or 3 days starting from day-10 of adulthood. Data from 2 experiments were pooled and analyzed by log-rank test (Supplementary Table 2). (E) L-carnitine did not further increase lifespan of glp-1 worms. Age-matched wild-type or glp-1 worms were raised at 25° C from L1 to L4 stage and then maintained at 20° C throughout life. 10 μM L-carnitine supplement was added starting from L1 stage. Dead and live worms were counted every 2 or 3 days starting from day-10 of adulthood. Data from 2 experiments were pooled and analyzed by log-rank test (Supplementary Table 2).
Second, we tested if L-carnitine could further increase the lifespan of glp-1 and daf-2 mutants. In several biological repeats, we found that L-carnitine could no longer increase lifespan of glp-1(e2144) and daf-2(e2370) mutant (Figure 4D, 4E). These results suggest that the activation of DAF-16 and SKN-1 in glp-1 and daf-2 mutants dominate the effect of L-carnitine, making these mutants no longer sensitive to L-carnitine.
A potential carnitine transporter T08B1.1 promotes oxidative stress recovery and increases lifespan in C. elegans
Next, we investigated the mechanisms by taking advantage of the long-lived glp-1 and daf-2 mutants. We focused on genes that are upregulated by both mutants.
One such genes, T08B1.1, encoding a potential cation transporter, showed similarity to the carnitine transporter gene OCTN1 in humans (Figure 5A). Multiple sequence alignment of T08B1.1 with OCTN1 proteins in human, chicken and Drosophila showed 3 conserved protein domains (Supplementary Figure 2), despite moderate similarity at the primary sequence. We confirmed that T08B1.1 mRNA was upregulated in glp-1 and daf-2 mutants by using qPCR (Figure 5B). We tested if T08B1.1 protein could mediate carnitine transport by RNAi knockdown. Animals were cultured in the presence of 10 μM L-carnitine with or without T08B1.1 RNAi bacteria. By using the L-carnitine Assay Kit from Abcam, we showed that glp-1 and daf-2 mutants have higher carnitine content than WT controls after feeding L-carnitine for 48 hours from L1 stage (Figure 5C). L-carnitine levels were all reduced after T08B1.1 knockdown, suggesting T08B1.1 is a potential carnitine transporter.
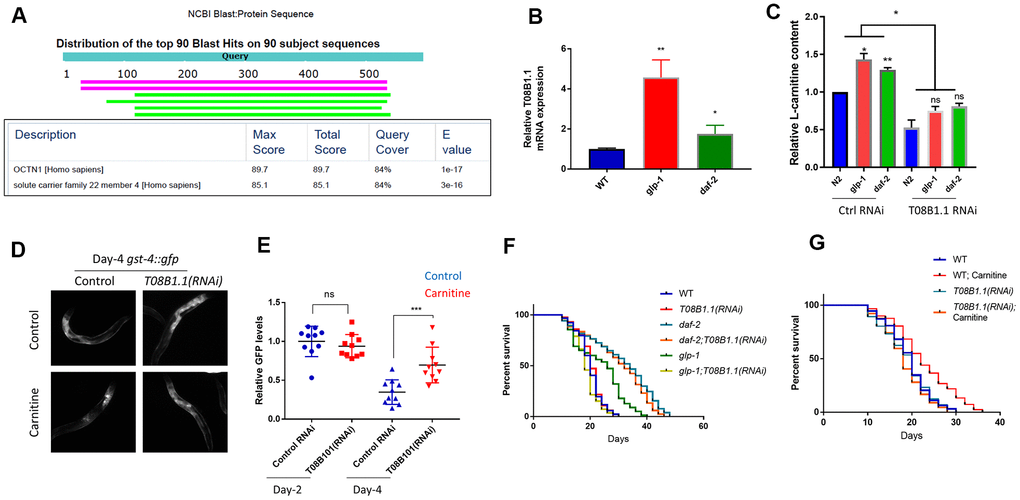
Figure 5. A potential carnitine transporter T08B1.1 promotes oxidative stress recovery and increases lifespan in C. elegans. (A) T08B1.1 is homologous to human carnitine transporter OCTN1. Protein sequence of T08B1.1 was used for blasting search with NCBI website with default setting for homologous sequences in human non-redundant protein sequences database. Table shows blasting results of the top 2 matched sequences. (B) T08B1.1 expression was elevated in glp-1 and daf-2 mutant worms. Age-matched glp-1 worms were raised at 25° C from L1 to L4 stage then changed to 20° C. Age-matched daf-2 worms were kept at 20° C. mRNAs were extracted at day-1 of adulthood and RT-qPCR was performed. Data from 3 experiments (each with 3 replicates) were analyzed with two-tailed, paired student’s t-test (*, P<0.05. **, P<0.01). Error bars indicate standard deviation of the mean. (C) RNAi knocking down of T08B1.1 decreased L-carnitine content in C. elegans. Age-synchronized L1 worms were raised on NG medium supplemented with10 μM L-carnitine until day-1 adulthood. Worms were homogenized and relative L-carnitine content measured with L-carnitine Assay kit. Data from 2 experiments were analyzed with two-tailed, paired student’s t-test (ns, not significant, *P<0.05, **P<0.01). Error bars indicate standard deviation of the mean. (D) RNAi knocking down of T08B1.1 prevented L-carnitine from decreasing oxidative marker gst-4::gfp. Age-synchronized worms expressing gst-4::gfp were raised on NG medium supplemented with10 μM L-carnitine until day-4 adulthood. Worms were imaged for GFP levels. Shown are representative images from day-4 adulthood. (E) Quantification of day-2 and day-4 data from experiments shown in (D). Ten images from 2 experiments were quantified by ImageJ and analyzed with two-tailed, unpaired student’s t-test (***, P<0.001). Error bars indicate standard deviation of the mean. (F) RNAi knocking down of T08B1.1 decreased glp-1 lifespan but not that of wild-type or daf-2 mutant. glp-1 worms were raised at 25° C from L1 to L4 stage and then changed to 20° C. WT and daf-2 worms were kept at 20° C. Dead and live worms were counted every 2 or 3 days starting from day-10 of adulthood. Data from 2 experiments (n>120 for each curve) were analyzed by log-rank test (Supplementary Table 3). (G) RNAi knocking down of T08B1.1 prevented L-carnitine from extending lifespan. Age-matched N2 wild-type worms were kept at 20° C throughout life. Dead and live worms were counted every 2 or 3 days starting from day-10 of adulthood. Data from 2 experiments (n>120 for each curve) were analyzed by log-rank test (Supplementary Table 4).
We continued to ask if T08B1.1 would be required for recovery from oxidative stress during normal aging. To this aim, we fed age-matched L1 C. elegans expressing gst-4:gfp with T08B1.1 RNAi bacteria on NGM plate in the absence or presence of 10 μM L-carnitine and checked GFP levels on day-4 of adulthood. Consistent with previous results, L-carnitine significantly reduced the gst-4::gfp expression, however, knocking down T08B1.1 reversed gst-4::gfp reduction (Figure 5D, 5E), confirming the role of T08B1.1 in oxidative stress recovery. Interestingly, T08B1.1 knockdown decreased the lifespan of glp-1 mutant but did not affect the lifespan of daf-2 mutant (Figure 5F). Consistently, T08B1.1 knockdown also blocked L-carnitine to extend lifespan, supporting the role of T08B1.1 in carnitine transport (Figure 5G).
Discussion
In this study, we found that L-carnitine could facilitate the recovery from oxidative stress, but did not affect the induction of OSR. L-carnitine delayed ROS accumulation during aging, reduced amyloid protein aggregation and prolonged lifespan. We also found that T08B1.1 gene, encoding a potential carnitine transporter, is required for efficient carnitine uptake and lifespan extension in worms treated with L-carnitine and germline mutant (glp-1) mutant. Together, by using the genetically tractable model, we have gained significant insights into the functions of L-carnitine on aging.
The effect of L-carnitine on oxidative stress recovery is unprecedented. L-carnitine is derived from lysine and methionine and such special structure could directly scavenge free radicals [1, 2]. Previous studies have shown that L-carnitine functions to reduce oxidative stress in mice, rat and humans [21, 52]. Among many other proposed mechanisms such as PI3K, AKT, ERK pathways [53, 54], the antioxidant effect of L-carnitine has been attributed to its role in activating Nrf2-dependent transcription [55–57]. However, we show that carnitine does not activate Nrf2 homolog SKN-1 in C. elegans. Rather, L-carnitine serves to facilitate the recovery from oxidative stress by mitigating SKN-1-mediated stress response. Therefore, L-carnitine likely functions through a feedback mechanism to control SKN-1 and oxidative stress. These results contrast with previous thoughts, opening new windows for further investigations into the underlying mechanisms. In addition, there are many antioxidative proteins other than SKN-1 and DAF-16 that could play more important roles in mediating L-carnitine’s effect on lifespan, which could be readily revealed by a genome-wide RNAi screen for suppressors of either markers (for example gst-4::gfp) or lifespan.
Our study has shown a direct role of L-carnitine in lifespan extension. L-carnitine has been shown to improve multiple aging phenotypes, including high blood lipid, high blood glucose, neurodegeneration and weak cardiac function [21, 58–60]. But its role in lifespan remains poorly studied. Only one study has been reported showing that acetyl-L-carnitine can extend the chronological lifespan in the budding yeast [27, 28]. By taking advantage of the genetically tractable model organism C. elegans, we show that L-carnitine was able to extend lifespan in worms, which was mediated through the potential carnitine transporter T08B1.1. Confirming the role of L-carnitine in lifespan regulation, we show that T08B1.1 is elevated in long-lived daf-2 and glp-1 mutants and required for glp-1 mutant to live long. T08B1.1 knockdown does not decrease lifespan of daf-2 mutant, likely due to the fact that the daf-2(1370) mutant relies less on SKN-1-mediated oxidative stress response as shown before [36]. Alternatively, the constitutive activation of SKN-1 in daf-2 mutant could dominate the L-carnitine signaling, making the mutant insensitive to T08B1.1 RNAi knockdown. Our study has established C. elegans as a model for mechanistic study of L-carnitine’s effect on delaying aging, which could boost the discovery of new strategies against aging and related disease.
Materials and Methods
Medium and bacteria and C. elegans strains
Medium used in this study were liquid LB medium for culturing bacterial food OP-50. For culturing RNAi bacteria HT115 bearing a plasmid, 50 ug/mL of Ampicillin was added to liquid LB medium. Nematode grow medium (NGM) were prepared as shown in [61]. C. elegans were raised at 20° C on NGM agar plates seeded with OP-50 bacteria. Worms were maintained by transferring 10 L1 larva to new plate every 3 day. Strains used in this study were: WT strain is N2 (Bristol), CL2006: dvIs2 [pCL12(unc-54/human Abeta peptide 1-42 minigene) + pRF4], daf-16(mu86) I, daf-2(e1370) III, dvIs20[pAF15(gst-4::GFP::NLS)], glp-1(e2144) III, daf-2(e1370) III; dvIs20[pAF15(gst-4::GFP::NLS)], glp-1(e2144) III; dvIs20[pAF15(gst-4::GFP::NLS)]. glp-1(e2144) III mutant worms were phenotypically wild-type at 20º C and raised at 25° C from L1 to L4 stage to inactivate GLP-1 gene product to deplete germline.
Drug treatment
L-carnitine was prepared in 1 mM stock solution and added to the NGM plate to the final concentration of 100 μM at least 12 hours before use. 10 day-1 adult worms were transferred to the plate for egg laying for 2 hours. Age-matched eggs were then allowed to hatch. For paraquat treatment, 10 mM paraquat stock solution was added to the NGM plate to the final concentration of 1 mM at least 12 hours before use. Worms at L4/young adult stage were transferred to paraquat plates for 24 hours and transferred back to paraquat-free NGM plate for recovery. Juglone was added similarly as paraquat except that 300 μM final concentration was used.
ROS detection
To detect ROS levels by using microscope, animals were stained with fluorescent dye Dihydroethidium (DHE). Specifically, worms raised on agar plates were washed with M9 buffer and then incubated in M9 buffer containing 3 μM DHE for 30 min. Worms were washed extensively in M9 buffer before the ROS signal was imaged with fluorescence microscope. Fluorescence intensity was quantified by using ImageJ software.
Western blot of protein aggregates
Equal number of Aβ-expressing worms (CL2006) at L1 stage were raised to young adult stage and treated as indicated. Worms were washed from plates with ice-cold M9 buffer to remove bacteria. Worms were then sonicated in 1X RIPA Buffer (20 mM Tris-HCl pH 7.5. 150 mM NaCl, 1 mM Na2EDTA. 1 mM EGTA. 1% NP-40) then centrifuged at 18,400Xg for 20 min at 4° C. The supernatant is soluble fraction and the pellet is insoluble fraction. The insoluble pellet was washed 3 times with RIPA buffer then resuspended in 75 ml denaturing buffer (8 M Urea, 2% SDS, 50 mM DTT, 50 mM Tris pH 8). Protein samples from soluble and insoluble factions were boiled in SDS loading buffer and separated by SDS-PAGE and total protein were transferred to PVDF membrane for western blot analysis with anti-Aβ antibody (Abcam, ab10148) and β-actin (Abcam, ab14128).
Aβ-induced paralysis assay
CL2006 worms expressing human Aβ(1-42) in body-wall muscles were allowed to lay eggs to new plates for 2 hours. Age-matched progenies were allowed to grow on NGM medium plates at 25° C. 50 μM FUDR (5-Fluoro-2′-deoxyuridine) was added at L4/young adult stage to inhibit reproduction. Worms were gradually paralyzed during adulthood. Worms that failed to move forward or backward when touched by a platinum wire were scored as “paralyzed”. Worms were scored for paralysis at the indicated time points starting from day-1 of adulthood.
RNAi knockdown
RNAi knockdown was carried out by feeding worms with bacteria expressing double strain RNA (dsRNA) on NGM agar plate. All RNAi bacteria were from the Ahringer RNAi library [62]. To prepare the RNAi agar plate, RNAi bacteria were cultured at 37° C overnight and sub-cultured to early log phase at OD600=0.4. 100 ul of bacteria were added directly on NGM plate agar plate containing 50ug/ml Ampicillin and 1mM Isopropyl β-D-1-thiogalactopyranoside (IPTG) to induce dsRNA expression. For RNAi knockdown, 10 day-1 adult worms were transferred to the RNAi plate for egg laying for 2 hours. Age-matched eggs were then allowed to hatch and grow on RNAi bacteria plate. 50 μM FUDR (5-Fluoro-2′-deoxyuridine) was added at L4/young adult stage to inhibit reproduction.
Lifespan assay
For lifespan assay in C. elegans, age-matched L1 worms were raised on either OP-50 or RNAi bacteria plate until L4/young adult stage, then 50 μM FUDR (5-Fluoro-2′-deoxyuridine) was added to inhibit reproduction. Worms may be transferred to new plates as required. The number of live and dead worms was recorded every 2 or 3 days starting from day 10. Exploded and bagged worms and worms with protruding vulva were censored. Death was defined as lack of any visible movement for 5 seconds after touching the tail and head with a platinum wire.
Real time quantitative PCR
Age- matched worms were washed from NGM plates with ice-cold M9 buffer. Worms were washed 2 more times by centrifugation to remove residual bacteria. Total mRNA was extracted with TRIzol™ Plus RNA Purification Kit (ThermoFisher Scientific) by following manufacture’s protocol. mRNA was reverse-transcribed using One-step RT-qPCR kits (Takara). Quantitative PCR was performed using SYBR Green 2X Mater Mix (Applied Biosystems). Gene expression levels were normalized to actin (ACT-1). Primers were designed by Roche Universal Probe Library System Assay Design. Primers for T08B1.1 are: 5’-cgaagttattatggctggatcttc-3’ and 5’-tttgacgtccaaaatggtca-3’. Primers for ACT-1 are: 5’-tcggtatgggacagaaggac-3’ and 5’-catcccagttggtgacgata-3’.
Carnitine content measurement
Worms were grown on NGM medium plate containing 100 μM L-carnitine from hatching. On the day of experiment, worms were washed extensively with M9 buffer to remove bacteria and L-carnitine. Worms were then homogenized by sonication. Worm lysate were centrifuged at 18,400Xg for 20 min at 4° C. L-carnitine levels in the lysate were detected with L-carnitine Assay Kit (Abcam) by following manufacturer’s protocol. Protein concentration was measured by using BCA protein Assay Kit (Abcam) and served as input control.
Author Contributions
D.L. and Z.-L.O. designed the study. D.L., X.Z. and L.L performed the experiments. L.L. provided critical reagents. D.L. and Z.-L.O. wrote the manuscript. All authors revised the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Funding
This work was supported by the Key Research and Development Program of Hunan Province (Grant No: 2019DK2011).
References
- 1. Rebouche CJ, Bosch EP, Chenard CA, Schabold KJ, Nelson SE. Utilization of dietary precursors for carnitine synthesis in human adults. J Nutr. 1989; 119:1907–13. https://doi.org/10.1093/jn/119.12.1907 [PubMed]
- 2. Solarska K, Lewińska A, Karowicz-Bilińska A, Bartosz G. The antioxidant properties of carnitine in vitro. Cell Mol Biol Lett. 2010; 15:90–97. https://doi.org/10.2478/s11658-009-0036-y [PubMed]
- 3. Longo N, Frigeni M, Pasquali M. Carnitine transport and fatty acid oxidation. Biochim Biophys Acta. 2016; 1863:2422–35. https://doi.org/10.1016/j.bbamcr.2016.01.023 [PubMed]
- 4. Adeva-Andany MM, Calvo-Castro I, Fernández-Fernández C, Donapetry-García C, Pedre-Piñeiro AM. Significance of l-carnitine for human health. IUBMB Life. 2017; 69:578–94. https://doi.org/10.1002/iub.1646 [PubMed]
- 5. Mielgo-Ayuso J, Calleja-Gonzalez J, Marqués-Jiménez D, Caballero-García A, Córdova A, Fernández-Lázaro D. Effects of creatine supplementation on athletic performance in soccer players: a systematic review and meta-analysis. Nutrients. 2019; 11:757. https://doi.org/10.3390/nu11040757 [PubMed]
- 6. Nezu J, Tamai I, Oku A, Ohashi R, Yabuuchi H, Hashimoto N, Nikaido H, Sai Y, Koizumi A, Shoji Y, Takada G, Matsuishi T, Yoshino M, et al. Primary systemic carnitine deficiency is caused by mutations in a gene encoding sodium ion-dependent carnitine transporter. Nat Genet. 1999; 21:91–94. https://doi.org/10.1038/5030 [PubMed]
- 7. Frigeni M, Balakrishnan B, Yin X, Calderon FR, Mao R, Pasquali M, Longo N. Functional and molecular studies in primary carnitine deficiency. Hum Mutat. 2017; 38:1684–99. https://doi.org/10.1002/humu.23315 [PubMed]
- 8. Stanley CA. Carnitine deficiency disorders in children. Ann N Y Acad Sci. 2004; 1033:42–51. https://doi.org/10.1196/annals.1320.004 [PubMed]
- 9. Bernardini I, Rizzo WB, Dalakas M, Bernar J, Gahl WA. Plasma and muscle free carnitine deficiency due to renal fanconi syndrome. J Clin Invest. 1985; 75:1124–30. https://doi.org/10.1172/JCI111806 [PubMed]
- 10. Buist NR. Historical perspective on clinical trials of carnitine in children and adults. Ann Nutr Metab. 2016 (Suppl 3); 68:1–4. https://doi.org/10.1159/000448320 [PubMed]
- 11. Ames BN, Liu J. Delaying the mitochondrial decay of aging with acetylcarnitine. Ann N Y Acad Sci. 2004; 1033:108–16. https://doi.org/10.1196/annals.1320.010 [PubMed]
- 12. Liu J, Head E, Gharib AM, Yuan W, Ingersoll RT, Hagen TM, Cotman CW, Ames BN. Memory loss in old rats is associated with brain mitochondrial decay and RNA/DNA oxidation: partial reversal by feeding acetyl-L-carnitine and/or R-alpha -lipoic acid. Proc Natl Acad Sci USA. 2002; 99:2356–61. https://doi.org/10.1073/pnas.261709299 [PubMed]
- 13. Liu J, Killilea DW, Ames BN. Age-associated mitochondrial oxidative decay: improvement of carnitine acetyltransferase substrate-binding affinity and activity in brain by feeding old rats acetyl-L- carnitine and/or R-alpha -lipoic acid. Proc Natl Acad Sci USA. 2002; 99:1876–81. https://doi.org/10.1073/pnas.261709098 [PubMed]
- 14. Hagen TM, Liu J, Lykkesfeldt J, Wehr CM, Ingersoll RT, Vinarsky V, Bartholomew JC, Ames BN. Feeding acetyl-L-carnitine and lipoic acid to old rats significantly improves metabolic function while decreasing oxidative stress. Proc Natl Acad Sci USA. 2002; 99:1870–75. https://doi.org/10.1073/pnas.261708898 [PubMed]
- 15. Montgomery SA, Thal LJ, Amrein R. Meta-analysis of double blind randomized controlled clinical trials of acetyl-L-carnitine versus placebo in the treatment of mild cognitive impairment and mild Alzheimer’s disease. Int Clin Psychopharmacol. 2003; 18:61–71. https://doi.org/10.1097/00004850-200303000-00001 [PubMed]
- 16. Ferrari R, Merli E, Cicchitelli G, Mele D, Fucili A, Ceconi C. Therapeutic effects of l-carnitine and propionyl-L-carnitine on cardiovascular diseases: a review. Ann N Y Acad Sci. 2004; 1033:79–91. https://doi.org/10.1196/annals.1320.007 [PubMed]
- 17. Tarantini G, Scrutinio D, Bruzzi P, Boni L, Rizzon P, Iliceto S. Metabolic treatment with l-carnitine in acute anterior ST segment elevation myocardial infarction. A randomized controlled trial. Cardiology. 2006; 106:215–23. https://doi.org/10.1159/000093131 [PubMed]
- 18. Omori Y, Ohtani T, Sakata Y, Mano T, Takeda Y, Tamaki S, Tsukamoto Y, Kamimura D, Aizawa Y, Miwa T, Komuro I, Soga T, Yamamoto K. L-carnitine prevents the development of ventricular fibrosis and heart failure with preserved ejection fraction in hypertensive heart disease. J Hypertens. 2012; 30:1834–44. https://doi.org/10.1097/HJH.0b013e3283569c5a [PubMed]
- 19. Jing ZC, Wu BX, Peng JQ, Li XL, Pan L, Zhao SP, Li DY, Yu ZX, Gong JB, Zhao QY, Cao JN, Sheng GT, Li J, et al. Effect of intravenous L-carnitine in Chinese patients with chronic heart failure. European Heart Journal Supplements. 2016; 18:A27–A36.
- 20. Koeth RA, Wang Z, Levison BS, Buffa JA, Org E, Sheehy BT, Britt EB, Fu X, Wu Y, Li L, Smith JD, DiDonato JA, Chen J, et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med. 2013; 19:576–85. https://doi.org/10.1038/nm.3145 [PubMed]
- 21. Ribas GS, Vargas CR, Wajner M. L-carnitine supplementation as a potential antioxidant therapy for inherited neurometabolic disorders. Gene. 2014; 533:469–76. https://doi.org/10.1016/j.gene.2013.10.017 [PubMed]
- 22. Mingrone G. Carnitine in type 2 diabetes. Ann N Y Acad Sci. 2004; 1033:99–107. https://doi.org/10.1196/annals.1320.009 [PubMed]
- 23. Mingrone G, Greco AV, Capristo E, Benedetti G, Giancaterini A, De Gaetano A, Gasbarrini G. L-carnitine improves glucose disposal in type 2 diabetic patients. J Am Coll Nutr. 1999; 18:77–82. https://doi.org/10.1080/07315724.1999.10718830 [PubMed]
- 24. Bene J, Hadzsiev K, Melegh B. Role of carnitine and its derivatives in the development and management of type 2 diabetes. Nutr Diabetes. 2018; 8:8. https://doi.org/10.1038/s41387-018-0017-1 [PubMed]
- 25. Li S, Chen X, Li Q, Du J, Liu Z, Peng Y, Xu M, Li Q, Lei M, Wang C, Zheng S, Zhang X, Yu H, et al. Effects of acetyl-L-carnitine and methylcobalamin for diabetic peripheral neuropathy: a multicenter, randomized, double-blind, controlled trial. J Diabetes Investig. 2016; 7:777–85. https://doi.org/10.1111/jdi.12493 [PubMed]
- 26. Yang SK, Xiao L, Song PA, Xu X, Liu FY, Sun L. Effect of L-carnitine therapy on patients in maintenance hemodialysis: a systematic review and meta-analysis. J Nephrol. 2014; 27:317–29. https://doi.org/10.1007/s40620-013-0002-7 [PubMed]
- 27. Palermo V, Falcone C, Calvani M, Mazzoni C. acetyl-L-carnitine protects yeast cells from apoptosis and aging and inhibits mitochondrial fission. Aging Cell. 2010; 9:570–79. https://doi.org/10.1111/j.1474-9726.2010.00587.x [PubMed]
- 28. Barardo D, Thornton D, Thoppil H, Walsh M, Sharifi S, Ferreira S, Anžič A, Fernandes M, Monteiro P, Grum T, Cordeiro R, De-Souza EA, Budovsky A, et al. The DrugAge database of aging-related drugs. Aging Cell. 2017; 16:594–97. https://doi.org/10.1111/acel.12585 [PubMed]
- 29. Speijer D. How the mitochondrion was shaped by radical differences in substrates: what carnitine shuttles and uncoupling tell us about mitochondrial evolution in response to ROS. Bioessays. 2014; 36:634–43. https://doi.org/10.1002/bies.201400033 [PubMed]
- 30. Arockia Rani PJ, Panneerselvam C. Carnitine as a free radical scavenger in aging. Exp Gerontol. 2001; 36:1713–26. https://doi.org/10.1016/s0531-5565(01)00116-4 [PubMed]
- 31. Olsen A, Vantipalli MC, Lithgow GJ. Using caenorhabditis elegans as a model for aging and age-related diseases. Ann N Y Acad Sci. 2006; 1067:120–28. https://doi.org/10.1196/annals.1354.015 [PubMed]
- 32. Antebi A. Genetics of aging in caenorhabditis elegans. PLoS Genet. 2007; 3:1565–71. https://doi.org/10.1371/journal.pgen.0030129 [PubMed]
- 33. Lee SJ, Hwang AB, Kenyon C. Inhibition of respiration extends C. Elegans life span via reactive oxygen species that increase HIF-1 activity. Curr Biol. 2010; 20:2131–36. https://doi.org/10.1016/j.cub.2010.10.057 [PubMed]
- 34. Park SK, Tedesco PM, Johnson TE. Oxidative stress and longevity in caenorhabditis elegans as mediated by SKN-1. Aging Cell. 2009; 8:258–69. https://doi.org/10.1111/j.1474-9726.2009.00473.x [PubMed]
- 35. Wei Y, Kenyon C. Roles for ROS and hydrogen sulfide in the longevity response to germline loss in caenorhabditis elegans. Proc Natl Acad Sci USA. 2016; 113:E2832–41. https://doi.org/10.1073/pnas.1524727113 [PubMed]
- 36. Tullet JM, Hertweck M, An JH, Baker J, Hwang JY, Liu S, Oliveira RP, Baumeister R, Blackwell TK. Direct inhibition of the longevity-promoting factor SKN-1 by insulin-like signaling in C. Elegans. Cell. 2008; 132:1025–38. https://doi.org/10.1016/j.cell.2008.01.030 [PubMed]
- 37. Jiang Y, Bao H, Ge Y, Tang W, Cheng D, Luo K, Gong G, Gong R. Therapeutic targeting of GSK3β enhances the Nrf2 antioxidant response and confers hepatic cytoprotection in hepatitis C. Gut. 2015; 64:168–79. https://doi.org/10.1136/gutjnl-2013-306043 [PubMed]
- 38. Link CD. Expression of human beta-amyloid peptide in transgenic caenorhabditis elegans. Proc Natl Acad Sci USA. 1995; 92:9368–72. https://doi.org/10.1073/pnas.92.20.9368 [PubMed]
- 39. Deusing DJ, Beyrer M, Fitzenberger E, Wenzel U. Carnitine protects the nematode caenorhabditis elegans from glucose-induced reduction of survival depending on the nuclear hormone receptor DAF-12. Biochem Biophys Res Commun. 2015; 460:747–52. https://doi.org/10.1016/j.bbrc.2015.03.101 [PubMed]
- 40. Murakami S, Murakami H. The effects of aging and oxidative stress on learning behavior in C. Elegans. Neurobiol Aging. 2005; 26:899–905. https://doi.org/10.1016/j.neurobiolaging.2004.08.007 [PubMed]
- 41. Gülçin I. Antioxidant and antiradical activities of L-carnitine. Life Sci. 2006; 78:803–11. https://doi.org/10.1016/j.lfs.2005.05.103 [PubMed]
- 42. Cochemé HM, Murphy MP. Complex I is the major site of mitochondrial superoxide production by paraquat. J Biol Chem. 2008; 283:1786–98. https://doi.org/10.1074/jbc.M708597200 [PubMed]
- 43. Zhao F, Wang W, Wang C, Siedlak SL, Fujioka H, Tang B, Zhu X. Mfn2 protects dopaminergic neurons exposed to paraquat both in vitro and in vivo: implications for idiopathic Parkinson’s disease. Biochim Biophys Acta Mol Basis Dis. 2017; 1863:1359–70. https://doi.org/10.1016/j.bbadis.2017.02.016 [PubMed]
- 44. Link CD, Johnson CJ. Reporter transgenes for study of oxidant stress in caenorhabditis elegans. Methods Enzymol. 2002; 353:497–505. https://doi.org/10.1016/s0076-6879(02)53072-x [PubMed]
- 45. Senchuk MM, Dues DJ, Van Raamsdonk JM. Measuring oxidative stress in Caenorhabditis elegans: paraquat and juglone sensitivity assays. Bio Protoc. 2017; 7:e2086. https://doi.org/10.21769/BioProtoc.2086 [PubMed]
- 46. Dues DJ, Schaar CE, Johnson BK, Bowman MJ, Winn ME, Senchuk MM, Van Raamsdonk JM. Uncoupling of oxidative stress resistance and lifespan in long-lived isp-1 mitochondrial mutants in caenorhabditis elegans. Free Radic Biol Med. 2017; 108:362–73. https://doi.org/10.1016/j.freeradbiomed.2017.04.004 [PubMed]
- 47. Bansal A, Zhu LJ, Yen K, Tissenbaum HA. Uncoupling lifespan and healthspan in caenorhabditis elegans longevity mutants. Proc Natl Acad Sci USA. 2015; 112:E277–86. https://doi.org/10.1073/pnas.1412192112 [PubMed]
- 48. McColl G, Roberts BR, Pukala TL, Kenche VB, Roberts CM, Link CD, Ryan TM, Masters CL, Barnham KJ, Bush AI, Cherny RA. Utility of an improved model of amyloid-beta (aβ₁₋₄₂) toxicity in caenorhabditis elegans for drug screening for Alzheimer’s disease. Mol Neurodegener. 2012; 7:57. https://doi.org/10.1186/1750-1326-7-57 [PubMed]
- 49. Cai Y, Wei YH. Stress resistance and lifespan are increased in C. elegans but decreased in S. cerevisiae by mafr-1/maf1 deletion. Oncotarget. 2016; 7:10812–26. https://doi.org/10.18632/oncotarget.7769 [PubMed]
- 50. Robida-Stubbs S, Glover-Cutter K, Lamming DW, Mizunuma M, Narasimhan SD, Neumann-Haefelin E, Sabatini DM, Blackwell TK. TOR signaling and rapamycin influence longevity by regulating SKN-1/Nrf and DAF-16/FoxO. Cell Metab. 2012; 15:713–24. https://doi.org/10.1016/j.cmet.2012.04.007 [PubMed]
- 51. Lin K, Hsin H, Libina N, Kenyon C. Regulation of the caenorhabditis elegans longevity protein DAF-16 by insulin/IGF-1 and germline signaling. Nat Genet. 2001; 28:139–45. https://doi.org/10.1038/88850 [PubMed]
- 52. Zanelli SA, Solenski NJ, Rosenthal RE, Fiskum G. Mechanisms of ischemic neuroprotection by acetyl-L-carnitine. Ann N Y Acad Sci. 2005; 1053:153–61. https://doi.org/10.1196/annals.1344.013 [PubMed]
- 53. Wang W, Lu Y, Xue Z, Li C, Wang C, Zhao X, Zhang J, Wei X, Chen X, Cui W, Wang Q, Zhou W. Rapid-acting antidepressant-like effects of acetyl-l-carnitine mediated by PI3K/AKT/BDNF/VGF signaling pathway in mice. Neuroscience. 2015; 285:281–91. https://doi.org/10.1016/j.neuroscience.2014.11.025 [PubMed]
- 54. Barhwal K, Hota SK, Prasad D, Singh SB, Ilavazhagan G. Hypoxia-induced deactivation of NGF-mediated ERK1/2 signaling in hippocampal cells: neuroprotection by acetyl-L-carnitine. J Neurosci Res. 2008; 86:2705–21. https://doi.org/10.1002/jnr.21722 [PubMed]
- 55. Li J, Zhang Y, Luan H, Chen X, Han Y, Wang C. L-carnitine protects human hepatocytes from oxidative stress-induced toxicity through Akt-mediated activation of Nrf2 signaling pathway. Can J Physiol Pharmacol. 2016; 94:517–25. https://doi.org/10.1139/cjpp-2015-0305 [PubMed]
- 56. Fan Z, Han Y, Ye Y, Liu C, Cai H. L-carnitine preserves cardiac function by activating p38 MAPK/Nrf2 signalling in hearts exposed to irradiation. Eur J Pharmacol. 2017; 804:7–12. https://doi.org/10.1016/j.ejphar.2017.04.003 [PubMed]
- 57. Zhang DM, Guo ZX, Zhao YL, Wang QJ, Gao YS, Yu T, Chen YK, Chen XM, Wang GQ. L-carnitine regulated Nrf2/Keap1 activation in vitro and in vivo and protected oxidized fish oil-induced inflammation response by inhibiting the NF-κB signaling pathway in rhynchocypris lagowski dybowski. Fish Shellfish Immunol. 2019; 93:1100–10. https://doi.org/10.1016/j.fsi.2019.08.041 [PubMed]
- 58. Ringseis R, Keller J, Eder K. Role of carnitine in the regulation of glucose homeostasis and insulin sensitivity: evidence from in vivo and in vitro studies with carnitine supplementation and carnitine deficiency. Eur J Nutr. 2012; 51:1–18. https://doi.org/10.1007/s00394-011-0284-2 [PubMed]
- 59. Asadi M, Rahimlou M, Shishehbor F, Mansoori A. The effect of l-carnitine supplementation on lipid profile and glycaemic control in adults with cardiovascular risk factors: a systematic review and meta-analysis of randomized controlled clinical trials. Clin Nutr. 2020; 39:110–22. https://doi.org/10.1016/j.clnu.2019.01.020 [PubMed]
- 60. DiNicolantonio JJ, Lavie CJ, Fares H, Menezes AR, O’Keefe JH. L-carnitine in the secondary prevention of cardiovascular disease: systematic review and meta-analysis. Mayo Clin Proc. 2013; 88:544–51. https://doi.org/10.1016/j.mayocp.2013.02.007 [PubMed]
- 61. Chaudhuri J, Parihar M, Pires-daSilva A. An introduction to worm lab: from culturing worms to mutagenesis. J Vis Exp. 2011; 47:2293. https://doi.org/10.3791/2293 [PubMed]
- 62. Fraser AG, Kamath RS, Zipperlen P, Martinez-Campos M, Sohrmann M, Ahringer J. Functional genomic analysis of C. Elegans chromosome I by systematic RNA interference. Nature. 2000; 408:325–30. https://doi.org/10.1038/35042517 [PubMed]