



Introduction
Currently, the use of assisted reproductive technology (ART) is widespread [1–3]. It is inevitable for oocytes used in ART to undergo in vitro culture before fertilization. If oocytes are not fertilized at the optimal period, unfertilized oocytes show a time-dependent decline in quality; this phenomenon is called in vitro oocyte aging [4, 5]. Oocyte quality determines the developmental capacity of the embryo after fertilization [4–6]. Compared with high-quality oocytes, poor-quality oocytes have reduced antioxidation activity and weak developmental competence [7, 8].
Many studies point to a series of changes in aged oocytes compared with fresh oocytes, such as zona pellucida hardening [9], mitochondrial dysfunction [6, 10], spindle abnormalities [5, 11] and chromosome aneuploidy [12]. Recent studies have shown that a decline in antioxidant capacity occurs during the oocyte aging process [13], which produces superfluous free radicals, including reactive oxygen species (ROS), inducing many cascades that influence oocyte quality, such as DNA damage, and apoptotic pathways [14–16]. Oocyte mitochondria are very vulnerable to oxidative stress and DNA damage, and dysfunctional mitochondria impair the production of adenosine triphosphate (ATP); moreover, an insufficient ATP supply is an important reason for decreased oocyte fertilization competence and altered embryo development [17]. Oxidative stress is also a factor that induces apoptosis and chromosome instability [18]. It is well known that antioxidants can partially counteract the negative effects of oxidative stress [19, 20]. Therefore, the balance between antioxidant and pro-oxidant substances is a critical factor influencing the quality of aged oocytes.
As a pentacyclic triterpene in Centella asiatica [21], asiatic acid (AA) has been shown to have many bioactivities, including antiapoptotic [22], anticancer [23], anti-inflammatory [24], neuroprotective [25], antifibrotic [26], and hypoglycemic action [27, 28]. In addition, AA effectively protects against oxidation [29], which could guard against the damage to the neuronal system resulting from quinolinic acid [30], alleviate seizures caused by kainic acid [31], and play an important role in preventing LPS/d-GalN-induced fulminant hepatic failure (FHF) by limiting oxidative stress [32]. Furthermore, our previous results suggested that AA exerts beneficial effects on porcine early embryonic development by ameliorating oxidative stress and preserving mitochondrial function [33]. It has been reported that AA might also be beneficial to the prevention or alleviation of brain aging [34].
Despite the well-known biological characteristics and physiological functions of AA and its beneficial protective effects, the protective effect and underlying mechanisms of AA on mammalian oocytes have not been defined. Thus, we hypothesized that AA could protect porcine oocytes from deterioration during the in vitro aging (IVA) process and could improve subsequent embryonic developmental competence. In the present research, we first explored the effects of AA supplementation during the IVA process on morphological changes and subsequent developmental competence of aged porcine oocytes. Second, we further analyzed the effects of AA on oxidative stress, apoptosis and mitochondrial function in porcine oocytes via the IVA model to identify the associated underlying mechanism. The results could shed light on the underlying beneficial effects of AA on aged oocyte quality.
Results
AA reduces morphological defects during the IVA process of porcine oocytes
Abnormal morphology often results in morphological changes. Therefore, we first investigated whether AA supplementation promoted the maintenance of the morphology of aged porcine oocytes. After in vitro maturation (IVM), mature oocytes were collected for IVA, and morphological changes were recorded at 24 and 48 h (Figure 1A–1D). As shown in Figure 1E, the percentage of abnormal oocytes increased in a time-dependent manner (IVA 24 h, 3.70 ± 0.81%, n=245; IVA 48 h, 37.52 ± 2.17%, n=245; p<0.05). Compared with the non-AA-supplemented group, the group of in vitro aged porcine oocytes supplemented with 10 μM AA showed a decrease in the percentage of oocytes with abnormal morphology at 24 h (0.85 ± 0.50%, n=200; 3.70 ± 0.81%, n=245; p<0.05). However, the morphological defects were not dramatically reduced by AA supplementation at 48 h of IVA (36.41 ± 1.97%, n=200; 37.52 ± 2.17%, n=245; p>0.05).
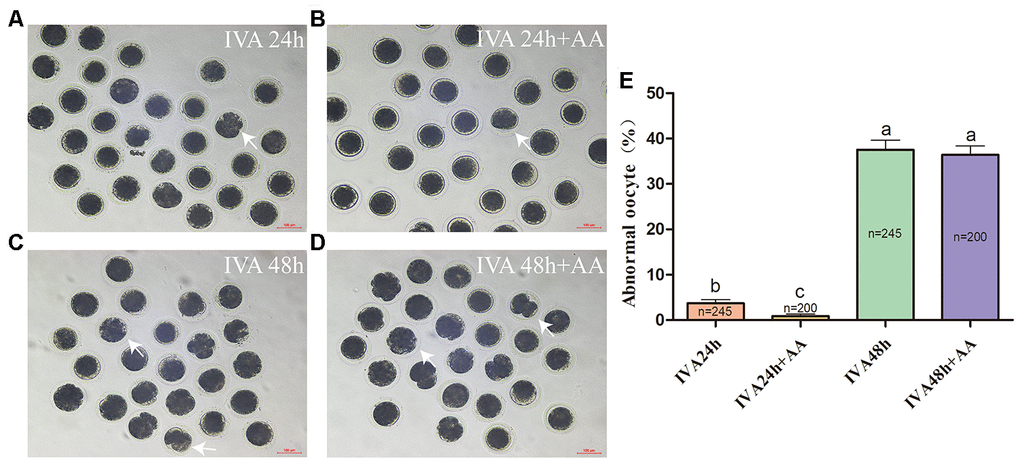
Figure 1. Changes in the morphology of in vitro aged porcine oocytes after AA supplementation. (A) 24-h in vitro aged oocytes; (B) 24-h in vitro aged oocytes supplemented with 10 μM AA; (C) 48-h in vitro aged oocytes; (D) 48-h in vitro aged oocytes supplemented with 10 μM AA; (E) morphological changes in porcine oocytes recovered at different culture time points from the AA supplementation groups. Representative morphologically abnormal in vitro aged oocytes (white arrows) were examined by optical microscopy. Scale bar=100 μm. The numbers of oocytes examined in each group are shown by the bars. Statistically significant differences are indicated by different letters (p<0.05).
AA supplementation improves the embryonic developmental potential of porcine in vitro aged oocytes after parthenogenetic activation
To extend our finding that supplementation with AA ameliorated the time-dependent deterioration of porcine oocyte quality, oocytes were parthenogenetically activated to evaluate subsequent embryonic development (Figure 2A). The results showed that AA supplementation during the IVA process ameliorated the subsequent declines in developmental competence. Although the percentage of activated oocytes that reached the 2-cell stage (52.77 ± 2.25%, n=159; 45.18 ± 4.00%, n=162; 45.76 ± 1.78%, n=160; p>0.05; Figure 2B) and 4-cell stage (72.25 ± 6.42%, n=159; 68.37 ± 4.36%, n=162; 70.50 ± 5.14%, n=160; p>0.05; Figure 2C) showed no obvious change, when in vitro aged porcine oocytes were supplemented with AA, the percentage of activated oocytes that reached the blastocyst stage was obviously improved compared with that not supplemented with AA (50.19 ± 2.85%, n=155; 12.27 ± 2.91%, n=160; 27.19 ± 4.94%, n=183; p<0.05; Figure 2D). Furthermore, the average total number of cells in blastocysts derived from AA-supplemented in vitro aged oocytes tended to be higher than that in blastocysts derived from non-supplemented in vitro aged oocytes (56.24 ± 1.13, n=41; 25.26 ± 1.18, n=39; 41.75 ± 2.96, n=40; p<0.05; Figure 2E). The average diameter of blastocysts derived from AA-supplemented in vitro aged oocytes was also decreased compared with that derived from non-supplemented in vitro aged oocytes (199.57 ± 5.86, n=49; 163.10 ± 4.18, n=41;180.31 ± 3.51, n=42; p<0.05; Figure 2F).
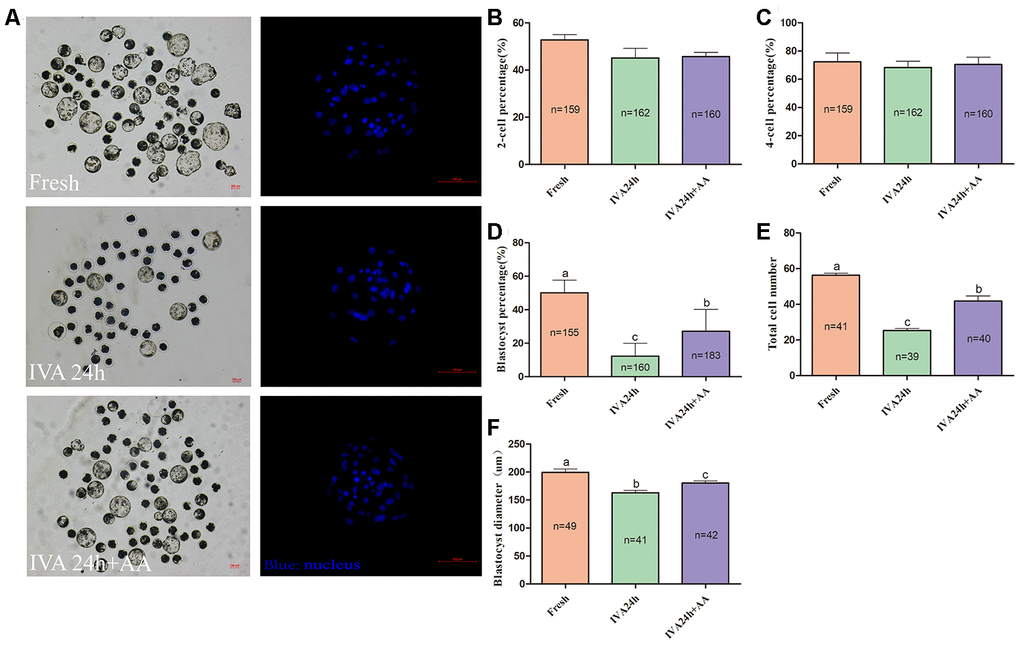
Figure 2. Effect of AA supplementation on the developmental competence of in vitro aged porcine oocytes after parthenogenetic activation. (A) Blastocyst formation at day 7 in each group (Left); and representative fluorescence images of blastocyst staining with Hoechst 33342 in each group (Right). Scale bar=100 μm. (B) Percentage of 2-cell embryos, (C) 4-cell embryos and (D) blastocyst formation in each group. (E) Total number of cells in blastocysts from each group. (F) Diameter of blastocysts from each group. The number of embryos examined in each group is shown by the bars. Statistically significant differences are indicated by different letters (p<0.05).
AA supplementation ameliorates sperm the binding ability of porcine in vitro aged oocytes and subsequent embryonic developmental competence after in vitro fertilization
The results of the present research showed that the decreased oocyte developmental competence induced by IVA was ameliorated by AA supplementation after in vitro fertilization. Sperm-egg binding assay results suggested that the average number of sperm that bound to the zona pellucida of in vitro aged oocytes tended to be higher in the AA-supplemented group than the non-supplemented in vitro aged group (162.26 ± 7.53, n=35; 36.34 ± 2.85, n=38; 101.28 ± 6.83, n=38; p<0.05; Figure 3A and 3B). The above results showed that AA supplementation could improve the quality of porcine oocytes. Therefore, in vitro fertilization was carried out with fresh, in vitro aged and AA-supplemented in vitro aged porcine oocytes (Figure 4A). The results showed that 23.06 ± 0.72% (n=190) of fresh oocytes could develop into the blastocyst stage. For in vitro aged porcine oocytes, the percentage of oocytes that developed into the blastocyst stage decreased to 6.23 ± 0.48% (n=194). However, in the AA-supplemented in vitro aged oocyte group, the percentage of oocytes that developed into the blastocyst stage was significantly increased (12.30 ± 0.67%, n=193). Similar to the blastocyst formation results, the average total number of cells in blastocysts derived from AA-supplemented in vitro aged oocytes was also increased compared with that of blastocysts derived from non-supplemented in vitro aged oocytes (45.68 ± 0.76, n=36; 25.22 ± 0.71, n=33; 34.85 ± 0.67, n=33; p<0.05; Figure 4C).
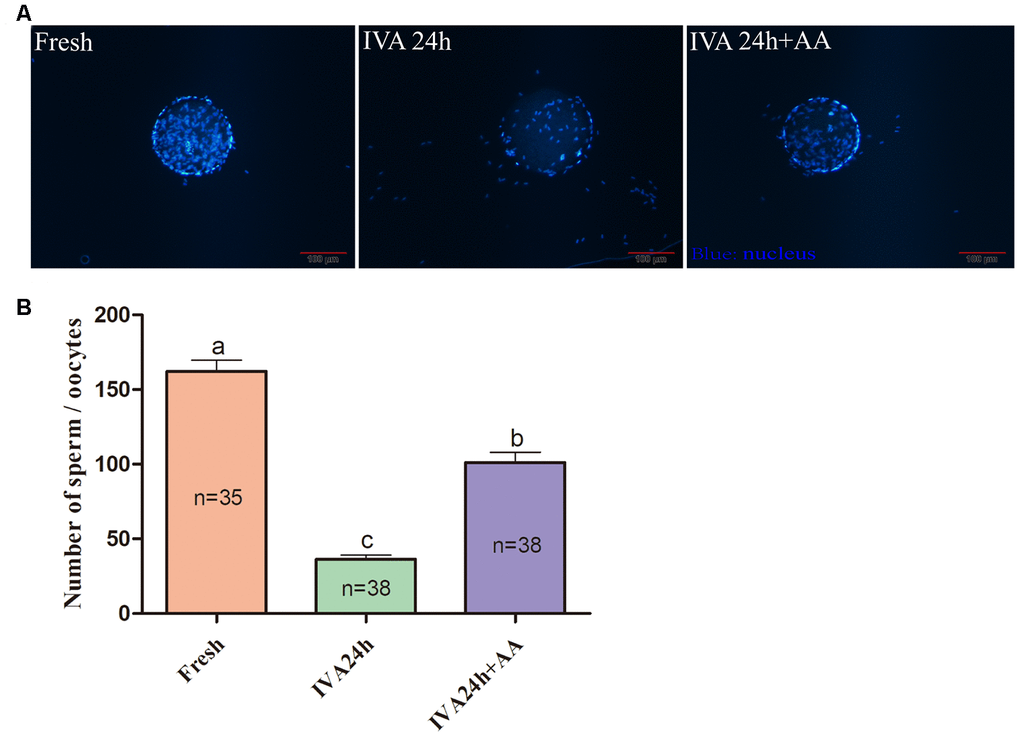
Figure 3. Effect of AA supplementation on sperm binding to the zona pellucida of porcine in vitro aged oocytes after in vitro fertilization. (A) Representative fluorescence images of sperm binding to the surface of the zona pellucida surrounding oocytes staining with Hoechst 33342 in each group. Scale-bar=100 μm. (B) Number of sperm binding to the surface of the zona pellucida surrounding oocytes in each group. The number of oocytes examined in each group is shown by the bars. Statistically significant differences are indicated by different letters (p<0.05).
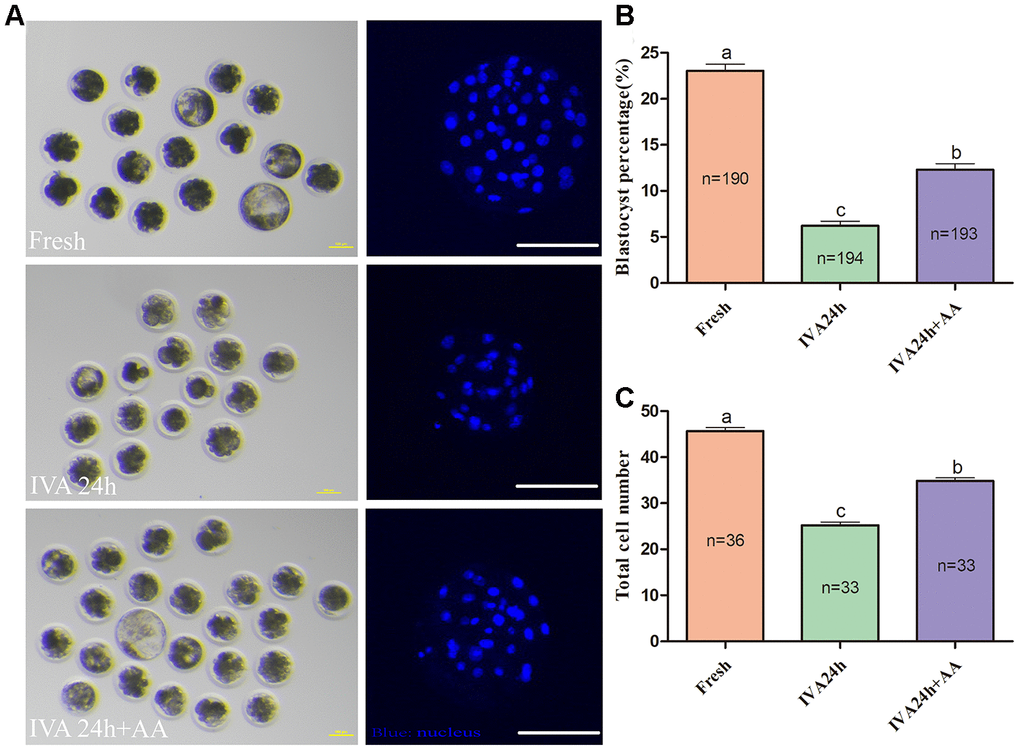
Figure 4. Effect of AA supplementation on the developmental competence of porcine in vitro aged oocytes after in vitro fertilization. (A) Blastocyst formation at day 7 in each group (Left); and representative fluorescence images of blastocyst staining with Hoechst 33342 in each group (Right). Scale bar=100 μm. (B) Percentage of blastocyst formation in each group. (C) Total number of cells in blastocysts from each group. The number of embryos examined in each group is shown by the bars. Statistically significant differences are indicated by different letters (p<0.05).
AA supplementation ameliorates oxidation resistance in porcine IVA oocytes
Because oxidative stress was associated with oocyte quality and subsequent embryonic developmental competence, we further assessed the free radical scavenging capacity of AA in in vitro aged porcine oocytes. The DCFH fluorescent reaction was used to analyze intracellular ROS levels in oocytes. As shown in Figure 5A and 5B, intracellular ROS levels were markedly increased in in vitro aged porcine oocytes compared with fresh oocytes, while AA supplementation effectively alleviated the increase in intracellular ROS levels during the IVA process of porcine oocytes. In accordance with the above results, we further assessed the intracellular GSH levels of in vitro aged porcine oocytes. As shown in Figure 6A and 6B, the intracellular GSH levels of in vitro aged porcine oocytes were markedly decreased compared with those of fresh oocytes, while AA supplementation effectively alleviated the decline in intracellular GSH levels in porcine oocytes during the IVA process.
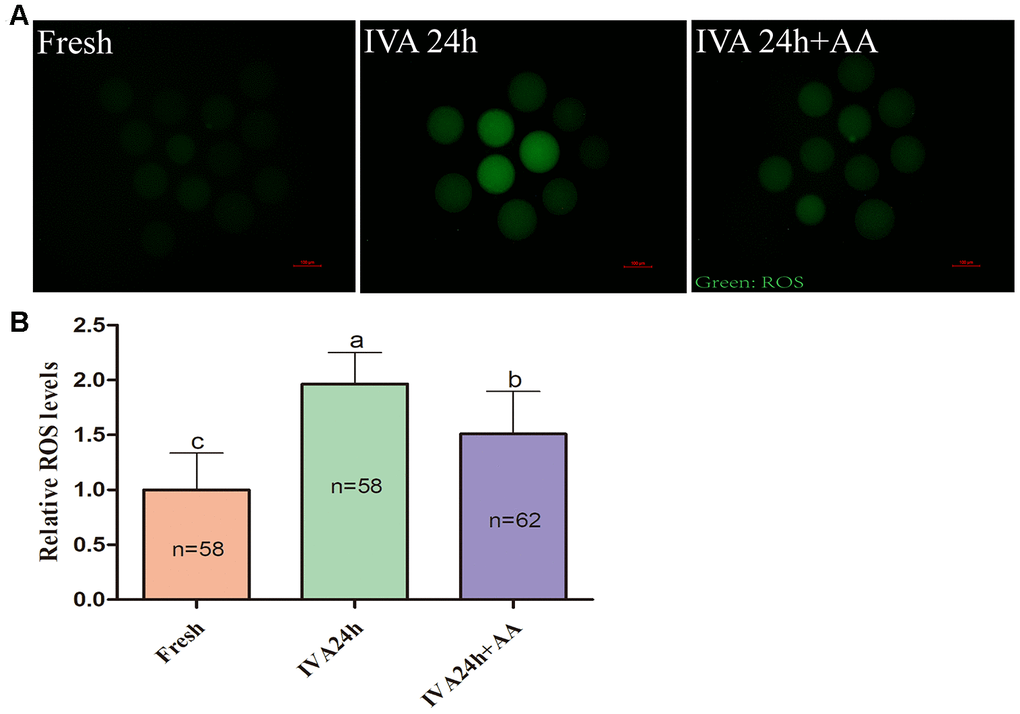
Figure 5. Effects of AA supplementation on the production of intracellular ROS in porcine IVA oocytes. (A) Representative fluorescence images of intracellular ROS in oocytes in each group. Scale bar=100 μm. (B) Relative intracellular ROS levels in oocytes in each group. The number of oocytes examined in each group is shown by the bars. Statistically significant differences are indicated by different letters (p<0.05).
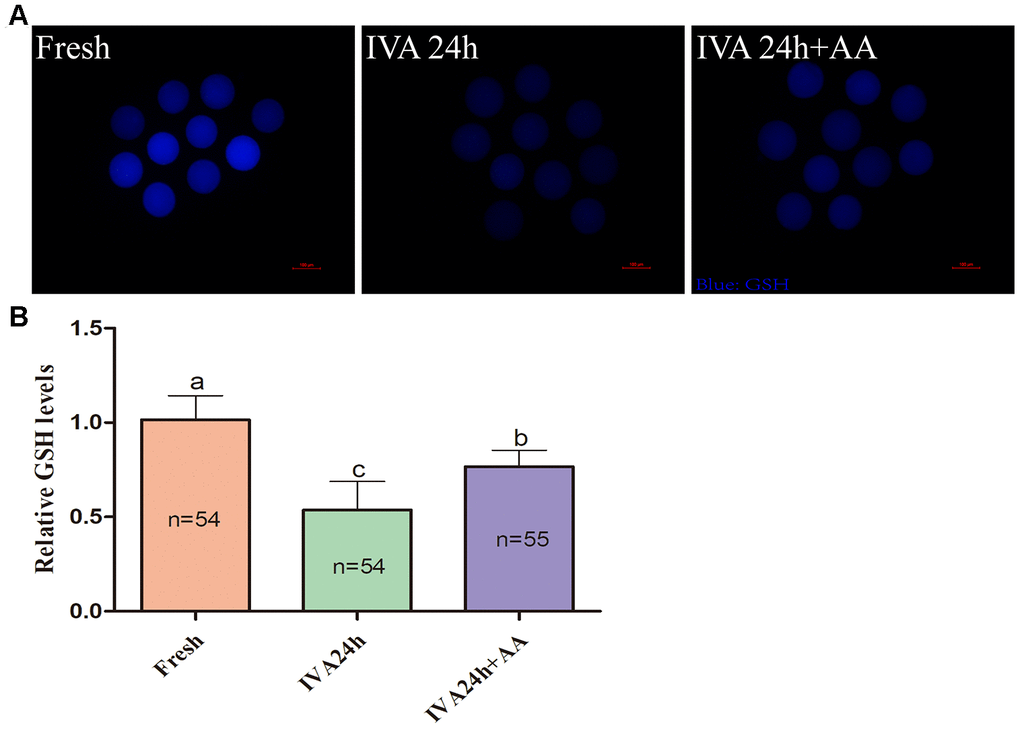
Figure 6. Effects of AA supplementation on the production of intracellular GSH in porcine IVA oocytes. (A) Representative fluorescence images of intracellular GSH in oocytes in each group. Scale bar=100 μm. (B) Relative intracellular GSH levels in oocytes in each group. The number of oocytes examined in each group is shown by the bars. Statistically significant differences are indicated by different letters (p<0.05).
AA supplementation decreased the early apoptosis of in vitro aged porcine oocytes
To further define the role of AA during the IVA process of porcine oocytes, the early apoptosis of in vitro aged porcine oocytes was monitored via the Annexin-V and TUNEL assays. As shown in Figure 7A and 7B, only 34.84 ± 2.26% (n=63) and 1.33 ± 1.33% (n=79) of fresh oocytes were positive for Annexin-V and TUNEL, respectively; however, the proportion of Annexin-V and TUNEL positive oocytes among in vitro aged oocytes were increased to 67.78 ± 3.98% (n=65) and 10.74 ± 1.03%, respectively (n=74); p<0.05. In contrast, in oocytes supplemented with AA during the process of IVA, the proportion of Annexin-V and TUNEL positive oocytes was reduced to 50.14 ± 2.00% (n=67) and 3.89 ± 2.04%, respectively (n=68); p<0.05.
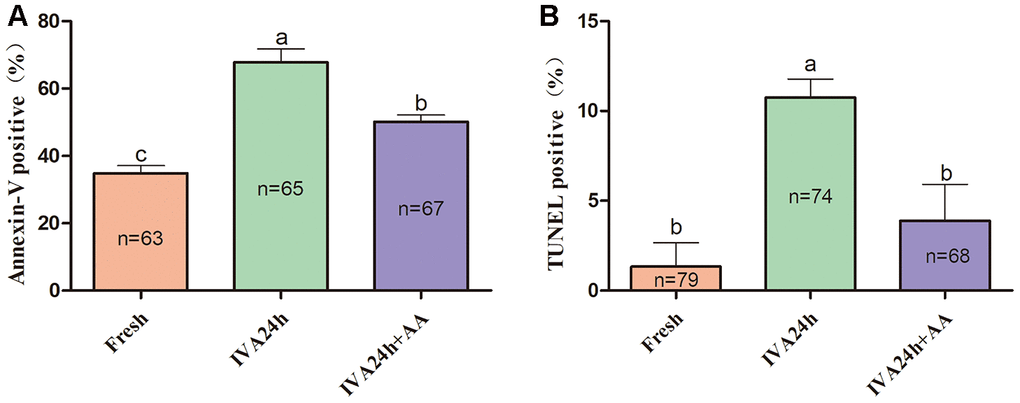
Figure 7. Effects of AA supplementation on the early apoptosis of in vitro aged porcine oocytes. (A) Percentage of Annexin-V and (B) TUNEL positive oocytes in each group. The number of oocytes examined in each group is shown by the bars. Statistically significant differences are indicated by different letters (p<0.05).
AA supplementation alleviated mitochondrial function in porcine IVA oocytes
Mitochondria play important roles in the regulation of cellular energy metabolism, free radical generation and apoptosis. Therefore, we evaluated ATP levels and mitochondrial membrane potential (MMP) in porcine IVA oocytes. As shown in Figure 8A and 8B, the ATP levels of in vitro aged porcine oocytes were markedly decreased compared with those fresh oocytes, while AA supplementation effectively alleviated the decline in ATP levels during the IVA process of porcine oocytes. The MMP of in vitro aged porcine oocytes was evaluated using the JC-1 fluorescent dye (Figure 9A). Quantification showed that the MMP of in vitro aged porcine oocytes was markedly decreased compared with that of fresh oocytes, while AA supplementation effectively alleviated the decrease in MMP during the IVA process of porcine oocytes (Figure 9B).
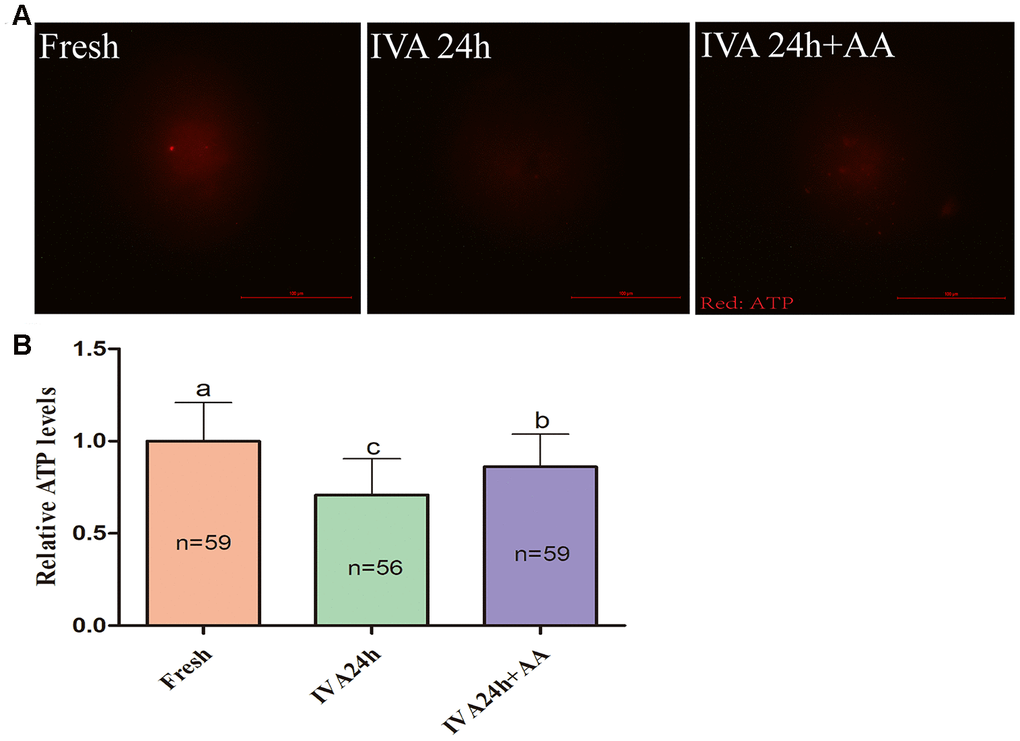
Figure 8. Effects of AA supplementation on the ATP production of in vitro aged porcine oocytes. (A) Representative fluorescence images of ATP staining in oocytes from each group. Scale bar=100 μm. (B) Relative ATP levels in oocytes from each group. The number of oocytes examined in each group is shown by the bars. Statistically significant differences are indicated by different letters (p<0.05).
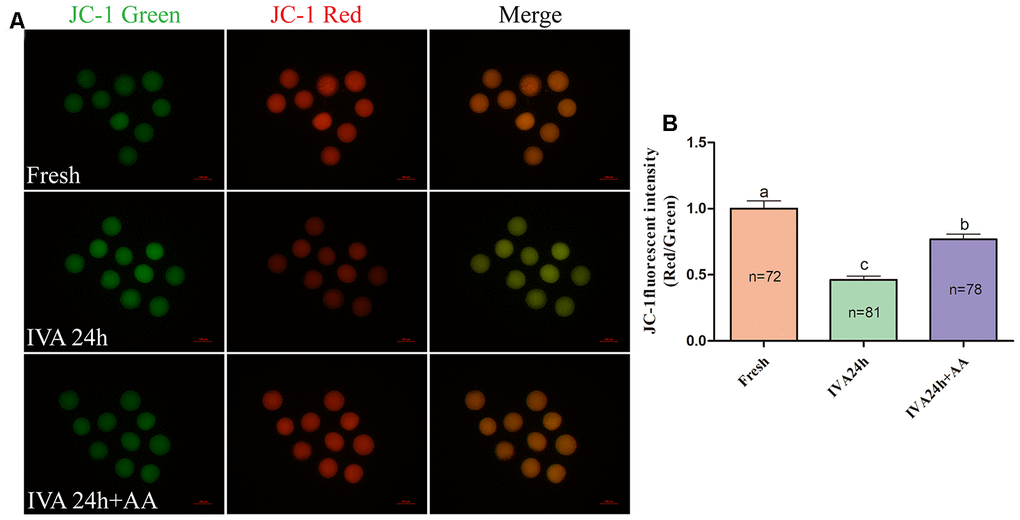
Figure 9. Effects of AA supplementation on the mitochondrial function of in vitro aged porcine oocytes. (A) Representative fluorescent images of JC-1 stained oocytes from each group. Scale bar=100 μm. (B) Relative JC-1 fluorescence intensity (red/green) in oocytes from each group. The number of oocytes examined in each group is shown by the bars. Statistically significant differences are indicated by different letters (p<0.05).
Discussion
The IVM of pig oocytes is used not only for understanding the event of meiosis but also for helping breed, clone and genome edit pigs for animal production. Previous research has shown that compared with that of other species oocytes, the maturation time of pig oocytes is prolonged [35]. During IVM, porcine oocytes are particularly susceptible to aging, leading to disturbances in metabolism, stress response and cell cycle regulation [36]. These changes reduce the quality of oocytes [13] and impact later embryonic development [7, 37]. In the present research, we found that AA could delay the process of pig oocyte IVA and subsequently improve embryonic developmental potential. Fertilization is a complex process of sexual reproduction in which the female and male gametes are mixed to generate new individuals [38]. Therefore, fertilization ability is one of the indicators used to evaluate oocyte quality. Our results show that AA can alleviate the decreased fertilization ability of aged oocytes, which is consistent with our results that AA could improve the parthenogenetic activation of developing embryos. In the reproduction process of animals, a particular sperm needs to arrive at, attach to, and mix with the egg plasma membrane; hence, it has to attach to and pass through the zona pellucida [39–41]. In other words, fertilization begins with the binding of the sperm to the zona pellucida. We found that AA could increase the number of sperm that bind to aged oocytes. Early studies suggested that the process of oocyte aging is usually accompanied by zona pellucida changes [5]. Thus, we suspect that AA may have other potential mechanisms to mitigate fertility declines due to aging by altering certain components of the zona pellucida.
Previous studies have shown that the quality of oocytes decreases gradually during the IVA process and that aged oocytes are sensitive to oxidative stress [37, 42]. Thus, this process of oocyte aging easily results in oxidative stress via the accumulation of free radicals. The increase in intracellular ROS could reduce the developmental potential of oocytes [43]. Mammalian cells have a wide range of antioxidants that have the ability to clear ROS, including non-enzymatic antioxidants [44] and several enzymatic antioxidants [45]. GSH, as an intrinsic antioxidant in oocytes, plays an important role in maintaining the redox state in oocytes [46]. GSH deficiency could lead to increased intracellular oxidative stress [47], germ cell apoptosis [48], and widespread mitochondrial damage [49], and a high intracellular GSH content could improve the quality of the oocytes [50]. In addition, a lack of GSH synthesis is the basis of oxidative stress during aging [51]. Our results demonstrate that AA supplementation clearly ameliorated oxidative stress because it not only decreased intracellular ROS levels but also maintained intracellular GSH levels during the process of IVA. Similarly, AA was reported to reduce intracellular ROS levels in microglial cells [52], HepG2 cells [53], and SH-SYS5Y cells [54] and increase superoxide dismutase activity and intracellular GSH production in spinal cord injury in rats [55]. Therefore, we hypothesized that AA might reduce oxidative stress by removing intracellular ROS and increasing GSH synthesis, thus delaying the decline in porcine oocyte quality during the process of IVA.
Functional mitochondria play an important role in normal oocyte function, since within both oocytes and early embryos, these organelles are the major source of ATP production [56]. Proton pumps, expressed on both sides of the mitochondrial membrane, regulate the potential of the mitochondrial membrane by the production of ATP [57]. An insufficient supply of ATP affects oocyte quality, leading to many problems, such as arrested cell division, abnormal cytokinesis, fragmentation [58–60], maturation and fertilization failure [17]. In mammals, the number of mitochondria increases dramatically with the growth of oocytes [61, 62], and after fertilization, the number of mitochondria remains constant from the early stages of embryo development through embryo implantation [62, 63]. Furthermore, abnormal MMP is closely related to fertilization, oocyte maturation, calcium homeostasis maintenance and apoptosis [17, 37, 64]. Therefore, it is very important for oocytes to have sufficient properly functioning mitochondria to ensure appropriate embryonic development. Increasing data have proven that changes in mitochondrial function are inextricably linked with stress-induced damage to oocyte developmental capacity [65]. Recent research shows that this impairment decreases mitochondrial ROS production, impairs the ability of mitochondria to sufficiently meet the cellular ATP demand, and induces mitochondrial-mediated apoptosis [66]. Moreover, mitochondria are considered to be a key element in the process of oocyte aging, which is greatly impacted by mitochondrial dysfunction [67, 68]. Our study shows that AA reduced the level of intracellular ROS and increased ATP activity and the red/green fluorescence ratio of JC-1, suggesting that mitochondrial dysfunction and oxidative stress were improved by AA supplementation during the process of porcine oocyte IVA. Since the findings in these experiments are in line with previous studies, it can be concluded that mitochondrial dysfunction can be well reduced by AA in various cells, including murine BV2 microglia cells, rat cortical neuronal cells [69, 70] and human A549 cells [71], and these results further confirm our hypothesis.
Since cell death via apoptosis is the last stage of oocyte aging [72, 73], it is hypothesized that AA can inhibit the activation of the apoptotic pathway of aged oocytes. We found that AA could delay the commencement of apoptosis in aged oocytes by Annexin-V and TUNEL assays. This function of AA is consistent with the results of other studies, which highlighted that AA protects against lactate-induced apoptosis through controlling the lactate signaling cascade and mediating mitochondria-dependent caspase activation and oxidative stress [71]; moreover, because of its antioxidant, mito-protective and antiapoptotic properties, pretreatment with AA before rotenone exposure in SH-SY5Y cells prevented marked ROS overproduction, mitochondrial dysfunction and apoptosis [54]. In early studies, the aging of humans and mammals was shown to be closely related to an increased level of ROS in mitochondria with age, which subsequently triggered apoptosis [74]. The problems associated with IVA appear to have a similar mechanism to the body's response to aging, which needs further study.
In summary, we found that AA could effectively improve the quality of in vitro aged porcine oocytes by relieving oxidative stress, reducing apoptosis, improving mitochondrial function, and promoting subsequent embryonic development (Figure 10). In other words, as an outstanding antioxidant that can be extracted from natural products, AA is expected to prevent the in vitro aging-induced decrease in oocyte quality.
Figure 10. Schematic representation of the protective action of AA on porcine IVA oocytes.
Materials and Methods
All chemicals used in this experiment were obtained from Sigma-Aldrich, a company based in St. Louis, America, except for those that are specifically defined.
Oocyte collection and IVM
Porcine ovaries were collected from a local abattoir and transported to the laboratory in 0.9% saline at 30-35° C. Follicular fluid was aspirated from 3-8 mm antral follicles using a syringe needle. Only evenly distributed cumulus oocyte complexes (COCs) with more than three layers of cumulus cells were collected under a stereomicroscope (S22-LGB, Nikon). After that, the COCs were washed three times in Tyrode’s lactate-HEPES buffered medium and were cultured in IVM medium (tissue culture medium 199 supplemented with 0.6 mM L-cysteine, 0.91 mM Na pyruvate, 10 ng/mL EGF, 10% porcine follicular fluid, 10 IU/mL follicle-stimulating hormone, 10 IU/mL luteinizing hormone and 75 mg/mL kanamycin) for 42 h at 38.5° C in an atmosphere of 5% CO2 and 100% humidity covered with mineral oil.
IVA of porcine oocytes
At the end of IVM, the expanded cumulus cells were removed from COCs by treatment with 0.1% hyaluronidase, and only matured oocytes with normal morphology were collected for IVA. The oocytes were then cultured in IVM medium, treated or not treated with 10 μM AA, covered with mineral oil in a damp environment of 38.5° C and 5% CO2 and cultured for 24 or 48 h. The final concentration of AA used in the present research was selected according to our preliminary experiment.
Parthenogenetic activation and in vitro fertilization
The parthenogenetic activation and in vitro fertilization procedures were performed as described in our previous research [33]. After parthenogenetic activation and in vitro fertilization, activated or fertilized oocytes were washed in porcine zygote medium (PZM)-3 and then transferred into four-well plates containing 500 μL PZM-3 covered with mineral oil and cultured for 7 days at 38.5° C in an atmosphere of 5% CO2 and 100% humidity without changing the medium. Blastocyst rates and diameters were analyzed on day 7, and blastocyst diameters were analyzed via NIH ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Assessment of the total number of cells in blastocysts
To quantifying the total number of cells contained in blastocysts, day 7 blastocysts derived from embryos that underwent parthenogenetic activation and in vitro fertilization were fixed in PBS-PVA with 4% PFA for 30 min at room temperature. Then, the blastocysts were washed with PBS-PVA three times and incubated with 10 μg/mL Hoechst 33342 for 5 min to label the nuclei. Next, blastocysts were gently mounted onto glass slides and examined under a fluorescence microscope (IX73, Olympus, Tokyo, Japan). NIH ImageJ software was used to analyze the total number of nuclei.
Sperm binding assay
Frozen boar sperm samples were used in this research, and the percentage of motile spermatozoa after thawing was 70%-90%. Approximately 15-20 oocytes were transferred into 60 μl droplets of mTBM for 30 min and then co-incubated with 20 μl diluted sperm for 1 h at 38.5° C in an atmosphere of 5% CO2 and 100% humidity. Afterwards, the oocytes were fixed in PBS-PVA with 4% PFA for 30 min at room temperature and stained with 10 μg/mL Hoechst 33342. Next, the oocytes were gently mounted onto glass slides and examined under a fluorescence microscope. The number of sperm bound to each oocyte was analyzed via NIH ImageJ software.
Evaluation of intracellular ROS and GSH levels in oocytes
The intracellular ROS and GSH levels in fresh and aged oocytes were measured using the fluorescent probes 2',7’-dichlorodihydrofluorescein diacetate (10 μM, Thermo Fisher Scientific, C400) and 4-chloromethyl-6,8-difluoro-7-hydroxycoumarin (10 μM, Thermo Fisher Scientific, C12881), respectively. Oocytes from each treatment group were incubated in the dark for 25 min at 38.5° C in an atmosphere of 5% CO2 and 100% humidity. After incubation, the oocytes were washed three times with PBS-PVA, and 10 μL droplets were removed. The fluorescence signal was captured using an epifluorescence microscope, and the fluorescence intensity of oocytes was analyzed using NIH ImageJ software.
Evaluation of oocyte apoptosis
The apoptosis of fresh and aged oocytes was measured via Annexin-V and TUNEL assays. Staining procedures were performed according to the product manual. For the Annexin-V assay, the oocytes were incubated in 100 μL Annexin-V binding solution with 5 μL Annexin-V and fluorescein isothiocyanate (FITC) conjugate for 30 min in the dark at room temperature. For the TUNEL assay, the oocytes were incubated with 50 μL TUNEL reaction mixture for 1 h at 37° C in the dark. The fluorescence signal was captured using a confocal microscope.
ATP measurement in oocytes
The ATP content of individual oocytes was measured using a fluorescent antibody (Abcam, 198367). Briefly, oocytes were collected, washed twice in PBS-PVA and then fixed in PBS-PVA with 4% PFA for 10 min at room temperature. Afterwards, the oocytes were washed three times with ice-cold PBS-PVA and then incubated with PBS-PVA containing 0.1% Triton X-100 for membrane permeabilization for 1 h. After being blocked for 30 min in 1.5% bovine serum albumin prepared in PBS-PVA (blocking solution), the oocytes were incubated with the antibody diluted in blocking solution (1:100, v/v) overnight at 4° C. After thoroughly washing with PBS-PVA, the fluorescence signal was captured using an epifluorescence microscope, and the fluorescence intensity of oocytes was analyzed using NIH ImageJ software.
JC-1 staining
To measure MMP, fresh and aged oocytes were washed three times with PBS-PVA. They were measured using the fluorescent probe 5,5’,6,6’-tetrachloro-1,1’,3,3’-tetraethyl-imidacarbocyanine iodide (2 μM, JC-1, Dojindo, China) for 30 min. After incubation, the oocytes were washed three times with the solution in the kit. The red and green fluorescence signals were captured using a fluorescence microscope connected to a digital camera. The fluorescence intensity of each oocyte was analyzed using NIH ImageJ software.
Statistical analysis
SPSS software version 11.0 (IBM, USA) was used to analyze all the data collected. The data from the two groups were compared with Student's t-test, the results are presented as the mean ± standard error of the mean, and considerable discrepancies are indicated by different letters (p<0.05).
Author Contributions
Shuang Liang, Bo-Xing Sun, Xiao Xia Li and Wei-Yi Hu conceived the experiment. Wei-Yi Hu, Xiao Xia Li and Yun Fei Diao performed the experiments. Wei-Yi Hu and Xiao Xia Li analyzed the data and wrote the article. Jia-Jia Qi and Da-Li Wang helped with the analysis of the results and revised the figures. Shuang Liang, Bo-Xing Sun and Jia-Bao Zhang assisted in the analyses of the results and helped revise the article.
Conflicts of Interest
All the authors declare that they have no conflicts of interest.
Funding
This research was supported by the National Natural Science Foundation of China (31802060), Jilin Scientific and Technological Development Program of China (20170307019NY, 20190201159JC) and the Modern Agricultural Industry Technology System (CARS-37).
Editorial Note
This corresponding author has a verified history of publications using a personal email address for correspondence
References
- 1. Davies MJ, Rumbold AR, Moore VM. Assisted reproductive technologies: a hierarchy of risks for conception, pregnancy outcomes, and treatment decisions - ERRATUM. J Dev Orig Health Dis. 2018; 9:241–46. https://doi.org/10.1017/S2040174417000757 [PubMed]
- 2. Kocourkova J, Burcin B, Kucera T. Demographic relevancy of increased use of assisted reproduction in European countries. Reprod Health. 2014; 11:37. https://doi.org/10.1186/1742-4755-11-37 [PubMed]
- 3. Sunderam S, Kissin DM, Zhang Y, Folger SG, Boulet SL, Warner L, Callaghan WM, Barfield WD. Assisted reproductive technology surveillance - United States, 2016. MMWR Surveill Summ. 2019; 68:1–23. https://doi.org/10.15585/mmwr.ss6804a1 [PubMed]
- 4. Lord T, Aitken RJ. Oxidative stress and ageing of the post-ovulatory oocyte. Reproduction. 2013; 146:R217–27. https://doi.org/10.1530/REP-13-0111 [PubMed]
- 5. Miao YL, Kikuchi K, Sun QY, Schatten H. Oocyte aging: cellular and molecular changes, developmental potential and reversal possibility. Hum Reprod Update. 2009; 15:573–85. https://doi.org/10.1093/humupd/dmp014 [PubMed]
- 6. Takahashi T, Igarashi H, Amita M, Hara S, Matsuo K, Kurachi H. Molecular mechanism of poor embryo development in postovulatory aged oocytes: mini review. J Obstet Gynaecol Res. 2013; 39:1431–39. https://doi.org/10.1111/jog.12111 [PubMed]
- 7. Lin T, Lee JE, Kang JW, Oqani RK, Cho ES, Kim SB, Il Jin D. Melatonin supplementation during prolonged in vitro maturation improves the quality and development of poor-quality porcine oocytes via anti-oxidative and anti-apoptotic effects. Mol Reprod Dev. 2018; 85:665–81. https://doi.org/10.1002/mrd.23052 [PubMed]
- 8. Kim E, Jeon Y, Kim DY, Lee E, Hyun SH. Antioxidative effect of carboxyethylgermanium sesquioxide (Ge-132) on IVM of porcine oocytes and subsequent embryonic development after parthenogenetic activation and IVF. Theriogenology. 2015; 84:226–36. https://doi.org/10.1016/j.theriogenology.2015.03.006 [PubMed]
- 9. Xu Z, Abbott A, Kopf GS, Schultz RM, Ducibella T. Spontaneous activation of ovulated mouse eggs: time-dependent effects on m-phase exit, cortical granule exocytosis, maternal messenger ribonucleic acid recruitment, and inositol 1,4,5-trisphosphate sensitivity. Biol Reprod. 1997; 57:743–50. https://doi.org/10.1095/biolreprod57.4.743 [PubMed]
- 10. Wilding M, Dale B, Marino M, di Matteo L, Alviggi C, Pisaturo ML, Lombardi L, De Placido G. Mitochondrial aggregation patterns and activity in human oocytes and preimplantation embryos. Hum Reprod. 2001; 16:909–17. https://doi.org/10.1093/humrep/16.5.909 [PubMed]
- 11. Wakayama S, Thuan NV, Kishigami S, Ohta H, Mizutani E, Hikichi T, Miyake M, Wakayama T. Production of offspring from one-day-old oocytes stored at room temperature. J Reprod Dev. 2004; 50:627–37. https://doi.org/10.1262/jrd.50.627 [PubMed]
- 12. Steuerwald NM, Steuerwald MD, Mailhes JB. Post-ovulatory aging of mouse oocytes leads to decreased MAD2 transcripts and increased frequencies of premature centromere separation and anaphase. Mol Hum Reprod. 2005; 11:623–30. https://doi.org/10.1093/molehr/gah231 [PubMed]
- 13. Takahashi T, Takahashi E, Igarashi H, Tezuka N, Kurachi H. Impact of oxidative stress in aged mouse oocytes on calcium oscillations at fertilization. Mol Reprod Dev. 2003; 66:143–52. https://doi.org/10.1002/mrd.10341 [PubMed]
- 14. Mihalas BP, Redgrove KA, McLaughlin EA, Nixon B. Molecular mechanisms responsible for increased vulnerability of the ageing oocyte to oxidative damage. Oxid Med Cell Longev. 2017; 2017:4015874. https://doi.org/10.1155/2017/4015874 [PubMed]
- 15. Lee SE, Kim EY, Choi HY, Moon JJ, Park MJ, Lee JB, Jeong CJ, Park SP. Rapamycin rescues the poor developmental capacity of aged porcine oocytes. Asian-Australas J Anim Sci. 2014; 27:635–47. https://doi.org/10.5713/ajas.2013.13816 [PubMed]
- 16. Lord T, Nixon B, Jones KT, Aitken RJ. Melatonin prevents postovulatory oocyte aging in the mouse and extends the window for optimal fertilization in vitro. Biol Reprod. 2013; 88:67. https://doi.org/10.1095/biolreprod.112.106450 [PubMed]
- 17. Van Blerkom J. Mitochondrial function in the human oocyte and embryo and their role in developmental competence. Mitochondrion. 2011; 11:797–813. https://doi.org/10.1016/j.mito.2010.09.012 [PubMed]
- 18. Liu L, Trimarchi JR, Navarro P, Blasco MA, Keefe DL. Oxidative stress contributes to arsenic-induced telomere attrition, chromosome instability, and apoptosis. J Biol Chem. 2003; 278:31998–2004. https://doi.org/10.1074/jbc.M303553200 [PubMed]
- 19. Sies H. Strategies of antioxidant defense. Eur J Biochem. 1993; 215:213–19. https://doi.org/10.1111/j.1432-1033.1993.tb18025.x [PubMed]
- 20. Sies H. Oxidative stress: oxidants and antioxidants. Exp Physiol. 1997; 82:291–95. https://doi.org/10.1113/expphysiol.1997.sp004024 [PubMed]
- 21. Lv J, Sharma A, Zhang T, Wu Y, Ding X. Pharmacological review on Asiatic acid and its derivatives: a potential compound. SLAS Technol. 2018; 23:111–27. https://doi.org/10.1177/2472630317751840 [PubMed]
- 22. Ren L, Cao QX, Zhai FR, Yang SQ, Zhang HX. Asiatic acid exerts anticancer potential in human ovarian cancer cells via suppression of PI3K/Akt/mTOR signalling. Pharm Biol. 2016; 54:2377–82. https://doi.org/10.3109/13880209.2016.1156709 [PubMed]
- 23. Ding H, Xiong Y, Sun J, Chen C, Gao J, Xu H. Asiatic acid prevents oxidative stress and apoptosis by inhibiting the translocation of α-synuclein into mitochondria. Front Neurosci. 2018; 12:431. https://doi.org/10.3389/fnins.2018.00431 [PubMed]
- 24. Qian Y, Xin Z, Lv Y, Wang Z, Zuo L, Huang X, Li Y, Xin HB. Asiatic acid suppresses neuroinflammation in BV2 microglia via modulation of the Sirt1/NF-κB signaling pathway. Food Funct. 2018; 9:1048–57. https://doi.org/10.1039/c7fo01442b [PubMed]
- 25. Dong SH, Liu YW, Wei F, Tan HZ, Han ZD. Asiatic acid ameliorates pulmonary fibrosis induced by bleomycin (BLM) via suppressing pro-fibrotic and inflammatory signaling pathways. Biomed Pharmacother. 2017; 89:1297–309. https://doi.org/10.1016/j.biopha.2017.03.005 [PubMed]
- 26. Ahmad Rather M, Justin Thenmozhi A, Manivasagam T, Dhivya Bharathi M, Essa MM, Guillemin GJ. Neuroprotective role of Asiatic acid in aluminium chloride induced rat model of Alzheimer’s disease. Front Biosci (Schol Ed). 2018; 10:262–75. https://doi.org/10.2741/s514 [PubMed]
- 27. Sun W, Xu G, Guo X, Luo G, Wu L, Hou Y, Guo X, Zhou J, Xu T, Qin L, Fan Y, Han L, Matsabisa M, et al. Protective effects of Asiatic acid in a spontaneous type 2 diabetic mouse model. Mol Med Rep. 2017; 16:1333–39. https://doi.org/10.3892/mmr.2017.6684 [PubMed]
- 28. Wang X, Lu Q, Yu DS, Chen YP, Shang J, Zhang LY, Sun HB, Liu J. Asiatic acid mitigates hyperglycemia and reduces islet fibrosis in Goto-Kakizaki rat, a spontaneous type 2 diabetic animal model. Chin J Nat Med. 2015; 13:529–34. https://doi.org/10.1016/S1875-5364(15)30047-9 [PubMed]
- 29. Maneesai P, Bunbupha S, Kukongviriyapan U, Senggunprai L, Kukongviriyapan V, Prachaney P, Pakdeechote P. Effect of Asiatic acid on the Ang II-AT1R-NADPH oxidase-NF-κB pathway in renovascular hypertensive rats. Naunyn Schmiedebergs Arch Pharmacol. 2017; 390:1073–83. https://doi.org/10.1007/s00210-017-1408-x [PubMed]
- 30. Loganathan C, Thayumanavan P. Asiatic acid prevents the quinolinic acid-induced oxidative stress and cognitive impairment. Metab Brain Dis. 2018; 33:151–59. https://doi.org/10.1007/s11011-017-0143-9 [PubMed]
- 31. Wang ZH, Mong MC, Yang YC, Yin MC. Asiatic acid and maslinic acid attenuated kainic acid-induced seizure through decreasing hippocampal inflammatory and oxidative stress. Epilepsy Res. 2018; 139:28–34. https://doi.org/10.1016/j.eplepsyres.2017.11.003 [PubMed]
- 32. Lv H, Qi Z, Wang S, Feng H, Deng X, Ci X. Asiatic acid exhibits anti-inflammatory and antioxidant activities against lipopolysaccharide and d-galactosamine-induced fulminant hepatic failure. Front Immunol. 2017; 8:785. https://doi.org/10.3389/fimmu.2017.00785 [PubMed]
- 33. Qi JJ, Li XX, Diao YF, Liu PL, Wang DL, Bai CY, Yuan B, Liang S, Sun BX. Asiatic acid supplementation during the in vitro culture period improves early embryonic development of porcine embryos produced by parthenogenetic activation, somatic cell nuclear transfer and in vitro fertilization. Theriogenology. 2020; 142:26–33. https://doi.org/10.1016/j.theriogenology.2019.09.027 [PubMed]
- 34. Chao PC, Yin MC, Mong MC. Anti-apoptotic and anti-glycative effects of Asiatic acid in the brain of D-galactose treated mice. Food Funct. 2015; 6:542–48. https://doi.org/10.1039/c4fo00862f [PubMed]
- 35. Kalous J, Tetkova A, Kubelka M, Susor A. Importance of ERK1/2 in regulation of protein translation during oocyte meiosis. Int J Mol Sci. 2018; 19:698. https://doi.org/10.3390/ijms19030698 [PubMed]
- 36. Jiang GJ, Wang K, Miao DQ, Guo L, Hou Y, Schatten H, Sun QY. Protein profile changes during porcine oocyte aging and effects of caffeine on protein expression patterns. PLoS One. 2011; 6:e28996. https://doi.org/10.1371/journal.pone.0028996 [PubMed]
- 37. Wang T, Gao YY, Chen L, Nie ZW, Cheng W, Liu X, Schatten H, Zhang X, Miao YL. Melatonin prevents postovulatory oocyte aging and promotes subsequent embryonic development in the pig. Aging (Albany NY). 2017; 9:1552–64. https://doi.org/10.18632/aging.101252 [PubMed]
- 38. Ohto U, Ishida H, Krayukhina E, Uchiyama S, Inoue N, Shimizu T. Structure of IZUMO1-JUNO reveals sperm-oocyte recognition during mammalian fertilization. Nature. 2016; 534:566–69. https://doi.org/10.1038/nature18596 [PubMed]
- 39. Clark GF. The molecular basis of mouse sperm-zona pellucida binding: a still unresolved issue in developmental biology. Reproduction. 2011; 142:377–81. https://doi.org/10.1530/REP-11-0118 [PubMed]
- 40. Bleil JD, Beall CF, Wassarman PM. Mammalian sperm-egg interaction: fertilization of mouse eggs triggers modification of the major zona pellucida glycoprotein, ZP2. Dev Biol. 1981; 86:189–97. https://doi.org/10.1016/0012-1606(81)90329-8 [PubMed]
- 41. Bleil JD, Wassarman PM. Sperm-egg interactions in the mouse: sequence of events and induction of the acrosome reaction by a zona pellucida glycoprotein. Dev Biol. 1983; 95:317–24. https://doi.org/10.1016/0012-1606(83)90032-5 [PubMed]
- 42. Nie J, Sui L, Zhang H, Zhang H, Yan K, Yang X, Lu S, Lu K, Liang X. Mogroside V protects porcine oocytes from in vitro ageing by reducing oxidative stress through SIRT1 upregulation. Aging (Albany NY). 2019; 11:8362–73. https://doi.org/10.18632/aging.102324 [PubMed]
- 43. Kudryavtseva AV, Krasnov GS, Dmitriev AA, Alekseev BY, Kardymon OL, Sadritdinova AF, Fedorova MS, Pokrovsky AV, Melnikova NV, Kaprin AD, Moskalev AA, Snezhkina AV. Mitochondrial dysfunction and oxidative stress in aging and cancer. Oncotarget. 2016; 7:44879–905. https://doi.org/10.18632/oncotarget.9821 [PubMed]
- 44. Aten RF, Duarte KM, Behrman HR. Regulation of ovarian antioxidant vitamins, reduced glutathione, and lipid peroxidation by luteinizing hormone and prostaglandin F2 alpha. Biol Reprod. 1992; 46:401–07. https://doi.org/10.1095/biolreprod46.3.401 [PubMed]
- 45. Sato EF, Kobuchi H, Edashige K, Takahashi M, Yoshioka T, Utsumi K, Inoue M. Dynamic aspects of ovarian superoxide dismutase isozymes during the ovulatory process in the rat. FEBS Lett. 1992; 303:121–25. https://doi.org/10.1016/0014-5793(92)80502-8 [PubMed]
- 46. Gasparrini B, Boccia L, Marchandise J, Di Palo R, George F, Donnay I, Zicarelli L. Enrichment of in vitro maturation medium for buffalo (bubalus bubalis) oocytes with thiol compounds: effects of cystine on glutathione synthesis and embryo development. Theriogenology. 2006; 65:275–87. https://doi.org/10.1016/j.theriogenology.2005.05.036 [PubMed]
- 47. Dickerhof N, Pearson JF, Hoskin TS, Berry LJ, Turner R, Sly PD, Kettle AJ, and AREST CF. Oxidative stress in early cystic fibrosis lung disease is exacerbated by airway glutathione deficiency. Free Radic Biol Med. 2017; 113:236–43. https://doi.org/10.1016/j.freeradbiomed.2017.09.028 [PubMed]
- 48. Lim J, Luderer U. Glutathione deficiency sensitizes cultured embryonic mouse ovaries to benzo[a]pyrene-induced germ cell apoptosis. Toxicol Appl Pharmacol. 2018; 352:38–45. https://doi.org/10.1016/j.taap.2018.05.024 [PubMed]
- 49. Meister A. Mitochondrial changes associated with glutathione deficiency. Biochim Biophys Acta. 1995; 1271:35–42. https://doi.org/10.1016/0925-4439(95)00007-q [PubMed]
- 50. Funahashi H, Cantley TC, Stumpf TT, Terlouw SL, Day BN. Use of low-salt culture medium for in vitro maturation of porcine oocytes is associated with elevated oocyte glutathione levels and enhanced male pronuclear formation after in vitro fertilization. Biol Reprod. 1994; 51:633–39. https://doi.org/10.1095/biolreprod51.4.633 [PubMed]
- 51. Sekhar RV, Patel SG, Guthikonda AP, Reid M, Balasubramanyam A, Taffet GE, Jahoor F. Deficient synthesis of glutathione underlies oxidative stress in aging and can be corrected by dietary cysteine and glycine supplementation. Am J Clin Nutr. 2011; 94:847–53. https://doi.org/10.3945/ajcn.110.003483 [PubMed]
- 52. Chen D, Zhang XY, Sun J, Cong QJ, Chen WX, Ahsan HM, Gao J, Qian JJ. Asiatic acid protects dopaminergic neurons from neuroinflammation by suppressing mitochondrial ros production. Biomol Ther (Seoul). 2019; 27:442–49. https://doi.org/10.4062/biomolther.2018.188 [PubMed]
- 53. Qi Z, Ci X, Huang J, Liu Q, Yu Q, Zhou J, Deng X. Asiatic acid enhances Nrf2 signaling to protect HepG2 cells from oxidative damage through Akt and ERK activation. Biomed Pharmacother. 2017; 88:252–59. https://doi.org/10.1016/j.biopha.2017.01.067 [PubMed]
- 54. Nataraj J, Manivasagam T, Justin Thenmozhi A, Essa MM. Neuroprotective effect of Asiatic acid on rotenone-induced mitochondrial dysfunction and oxidative stress-mediated apoptosis in differentiated SH-SYS5Y cells. Nutr Neurosci. 2017; 20:351–59. https://doi.org/10.1080/1028415X.2015.1135559 [PubMed]
- 55. Jiang W, Li M, He F, Bian Z, He Q, Wang X, Yao W, Zhu L. Neuroprotective effect of Asiatic acid against spinal cord injury in rats. Life Sci. 2016; 157:45–51. https://doi.org/10.1016/j.lfs.2016.05.004 [PubMed]
- 56. Dumollard R, Ward Z, Carroll J, Duchen MR. Regulation of redox metabolism in the mouse oocyte and embryo. Development. 2007; 134:455–65. https://doi.org/10.1242/dev.02744 [PubMed]
- 57. Yao X, Jiang H, Liang S, Shen X, Gao Q, Xu YN, Kim NH. Laminarin enhances the quality of aged pig oocytes by reducing oxidative stress. J Reprod Dev. 2018; 64:489–94. https://doi.org/10.1262/jrd.2018-031 [PubMed]
- 58. Van Blerkom J. Mitochondria in human oogenesis and preimplantation embryogenesis: engines of metabolism, ionic regulation and developmental competence. Reproduction. 2004; 128:269–80. https://doi.org/10.1530/rep.1.00240 [PubMed]
- 59. May-Panloup P, Chretien MF, Malthiery Y, Reynier P. Mitochondrial DNA in the oocyte and the developing embryo. Curr Top Dev Biol. 2007; 77:51–83. https://doi.org/10.1016/S0070-2153(06)77003-X [PubMed]
- 60. Ramalho-Santos J, Varum S, Amaral S, Mota PC, Sousa AP, Amaral A. Mitochondrial functionality in reproduction: from gonads and gametes to embryos and embryonic stem cells. Hum Reprod Update. 2009; 15:553–72. https://doi.org/10.1093/humupd/dmp016 [PubMed]
- 61. Michaels GS, Hauswirth WW, Laipis PJ. Mitochondrial DNA copy number in bovine oocytes and somatic cells. Dev Biol. 1982; 94:246–51. https://doi.org/10.1016/0012-1606(82)90088-4 [PubMed]
- 62. Smith LC, Alcivar AA. Cytoplasmic inheritance and its effects on development and performance. J Reprod Fertil Suppl. 1993; 48:31–43. [PubMed]
- 63. Dumollard R, Duchen M, Carroll J. The role of mitochondrial function in the oocyte and embryo. Curr Top Dev Biol. 2007; 77:21–49. https://doi.org/10.1016/S0070-2153(06)77002-8 [PubMed]
- 64. Bisht S, Dada R. Oxidative stress: major executioner in disease pathology, role in sperm DNA damage and preventive strategies. Front Biosci (Schol Ed). 2017; 9:420–47. https://doi.org/10.2741/s495 [PubMed]
- 65. Roth Z. Symposium review: reduction in oocyte developmental competence by stress is associated with alterations in mitochondrial function. J Dairy Sci. 2018; 101:3642–54. https://doi.org/10.3168/jds.2017-13389 [PubMed]
- 66. Faitg J, Reynaud O, Leduc-Gaudet JP, Gouspillou G. [Skeletal muscle aging and mitochondrial dysfunction: an update]. Med Sci (Paris). 2017; 33:955–62. https://doi.org/10.1051/medsci/20173311012 [PubMed]
- 67. Zhang D, Keilty D, Zhang ZF, Chian RC. Mitochondria in oocyte aging: current understanding. Facts Views Vis Obgyn. 2017; 9:29–38. [PubMed]
- 68. Reynier P, May-Panloup P, Chrétien MF, Morgan CJ, Jean M, Savagner F, Barrière P, Malthièry Y. Mitochondrial DNA content affects the fertilizability of human oocytes. Mol Hum Reprod. 2001; 7:425–29. https://doi.org/10.1093/molehr/7.5.425 [PubMed]
- 69. Park JH, Seo YH, Jang JH, Jeong CH, Lee S, Park B. Asiatic acid attenuates methamphetamine-induced neuroinflammation and neurotoxicity through blocking of NF-kB/STAT3/ERK and mitochondria-mediated apoptosis pathway. J Neuroinflammation. 2017; 14:240. https://doi.org/10.1186/s12974-017-1009-0 [PubMed]
- 70. Zhang X, Wu J, Dou Y, Xia B, Rong W, Rimbach G, Lou Y. Asiatic acid protects primary neurons against C2-ceramide-induced apoptosis. Eur J Pharmacol. 2012; 679:51–59. https://doi.org/10.1016/j.ejphar.2012.01.006 [PubMed]
- 71. Gao C, Wang F, Wang Z, Zhang J, Yang X. Asiatic acid inhibits lactate-induced cardiomyocyte apoptosis through the regulation of the lactate signaling cascade. Int J Mol Med. 2016; 38:1823–30. https://doi.org/10.3892/ijmm.2016.2783 [PubMed]
- 72. Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007; 35:495–516. https://doi.org/10.1080/01926230701320337 [PubMed]
- 73. Wang H, Jo YJ, Oh JS, Kim NH. Quercetin delays postovulatory aging of mouse oocytes by regulating SIRT expression and MPF activity. Oncotarget. 2017; 8:38631–41. https://doi.org/10.18632/oncotarget.16219 [PubMed]
- 74. Isaev NK, Genrikhs EE, Oborina MV, Stelmashook EV. Accelerated aging and aging process in the brain. Rev Neurosci. 2018; 29:233–40. https://doi.org/10.1515/revneuro-2017-0051 [PubMed]