



Introduction
Gliomas are the most common and deadliest malignant primary brain tumor in adults and accounts for approximately 80% of the primary malignant tumors in the central nervous system [1]. Based on the World Health Organization (WHO) classification criteria, diffuse glioma could be divided into grades II–IV. Lower-grade gliomas (LGGs), designated as astrocytoma, oligodendroglioma, and mixed oligoastrocytoma of grade II and III, account for approximately 43.2% of all gliomas diagnosed in adults [2, 3]. Glioblastoma (GBM) is the most malignant glioma, which refers to WHO grade IV glioma. Despite advances in comprehensive therapy, such as neurosurgical resection, adjuvant radiotherapy, and alkylating agent temozolomide chemotherapy, patients who suffer from gliomas have a short median survival time due to various reasons [4, 5]. The degree of genetic and phenotypic variations of the glioma cells form intratumoral heterogeneity, allows the most adaptive tumor clones to develop treatment resistance [6]. In order to overcome this issue, hundreds of molecular alterations have been identified in grade II, III, and IV gliomas [7]. Isocitrate dehydrogenase 1 (IDH1) mutation, which occurs early in gliomagenesis, especially for WHO grade II and III gliomas, is an acknowledged molecular alteration [8]. Mutation in IDH1 is a stable marker indicating prognosis in both LGGs and GBM with the incidence of 75% and 12%, respectively [9, 10]. However, previous studies have established gene signatures to divide the IDH1 mutation glioma patients with different clinical features into various groups [9, 11]. However, only a few studies have focused on the wild-type IDH1 patients. Therefore, there is an urgent need to identify specific genes for novel diagnosis and therapeutic strategies for human gliomas.
DAZ interacting zinc finger 3 (DZIP3) is first identified as an RNA-binding RING-dependent ubiquitin ligase [12]. It is involved in various physiological processes, such as regulation of chemokine- or estradiol-induced gene expression and self-renewal [13]. Ito et al. indicated that DZIP3 affects the developmental genes in mouse embryonic stem cells by reorganizing the 3D chromatin conformation, and knockdown of DZIP3 could result in an expansion of the mouse embryonic stem cells [12]. However, to the best of our knowledge, no study has explored the correlation between DZIP3 and gliomas.
In the present study, we recruited 590 glioma patients from CGGA RNA-seq (training set), and GSE 16011 array (validation set) sets to compare the mRNA expression level of DZIP3 from different clinical and molecular glioma subtypes. Gene enrichment analysis and immunohistochemistry (IHC) confirmed that DZIP3 could block glioma progression by affecting angiogenesis. Finally, in lower grade wild-type IDH1 patients, based on DZIP3 expression, two groups with different outcomes were established. One group was regarded as IDH1 mutation-like, and the other group was GBM-like.
Results
DZIP3 expression was downregulated in GBM and IDH1 wild-type glioma
IDH1 status was increasingly recognized as the crucial genetic marker for glioma patients and had been included in the 2016 WHO classification of glioma. To get an overview of DZIP3 status, we examined the expression pattern of DZIP3 across WHO grade and IDH1 status. The results demonstrated that DZIP3 was significantly downregulated in GBM and IDH1 wild-type groups (Figure 1A, 1B) in the CGGA RNA-seq set. Additionally, the results could be validated in 285 glioma patients from GSE 16011 array set (Figure 1C, 1D). To the best of our knowledge, GBM and IDH1 wild-type were markers that indicated malignancy and short survival time in glioma. These results indicated that DZIP3 worked as an anti-oncogene in gliomas. To verify the results above in protein level, we randomly selected 15 patients (5 for grade II, 5 for grade III, 5 for grade IV) to perform IHC to detect the relationship between tumor grade and DZIP3 protein expression and the results showed that DZIP3 protein expression was negatively associated with WHO grade (Figure 1E, 1F).
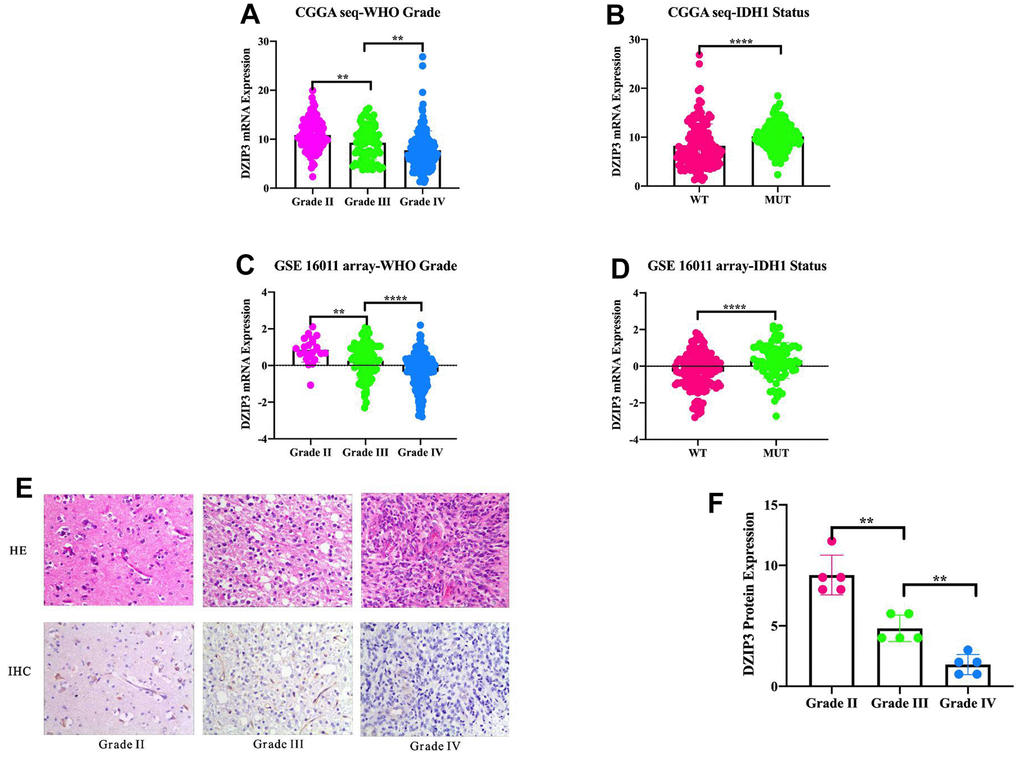
Figure 1. DZIP3 mRNA expression pattern in CGGA RNA-seq and GSE 16011 array cohorts.DZIP3 is enriched in low-grade gliomas in CGGA RNA-seq and GSE 16011 array sets (A, C). DZIP3 is enriched in IDH1-MUT gliomas in CGGA RNA-seq and GSE 16011 array sets (B, D). DZIP3 protein expression is negatively associated with WHO grade (E, F). **p<0.01 and ****p<0.0001.
DZIP3 was closely associated with angiogenesis function in glioma
Based on the aforementioned results, DZIP3 affected the key biological functions to change the malignancy of glioma. A total of 261 genes were found to be significantly negatively related with the expression of DZIP3 as assessed by Pearson’s correlation analysis (Pearson |R| > 0.4) in the CGGA RNA-seq and GSE 16011 array cohorts to explore the biological roles of DZIP3 in gliomas (Figure 2A). Next, we performed GO analysis via DAVID Bioinformatics Resources 6.8 to clarify the biological functions of the related genes mentioned above. The results indicated that genes, negatively associated with DZIP3, were primarily involved in angiogenesis (Figure 2B). IHC was used to evaluate the correlation between DZIP3 expression and CD31 (a novel angiogenesis marker) [14]; DZIP3 mainly expressed on vascular endothelial cells and was negatively associated with CD31 at the protein level indicating that DZIP3 could affect the angiogenesis process of glioma (Figure 3A). Moreover, MVD of CD31 immunoreactivity was performed as described, we found that CD31-MVD was significantly lower in patients with higher DZIP3 expression (Figure 3B).
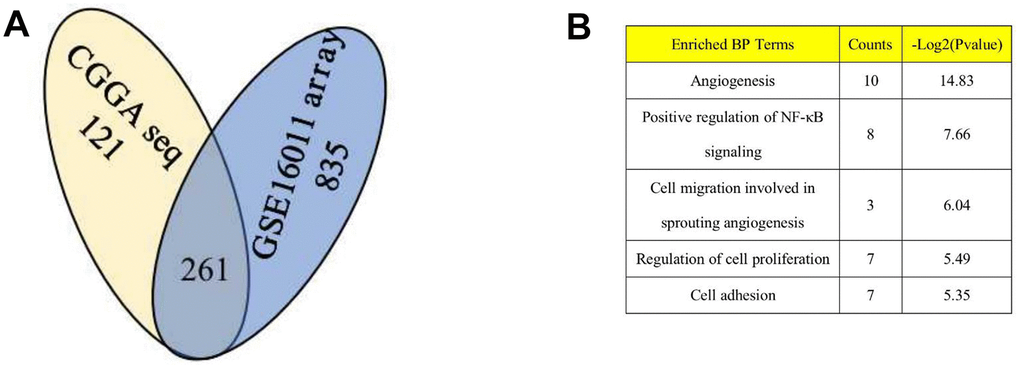
Figure 2. DZIP3 is significantly associated to angiogenesis pathway in gliomas. (A) 261 overlapping genes are negatively associated (Pearson |R| > 0.4) with DZIP3 in CGGA RNA-seq and GSE 16011 array sets; (B) GO analysis of the 261 genes indicates that DZIP3 is involved in angiogenesis, NF-KB and other GO function.
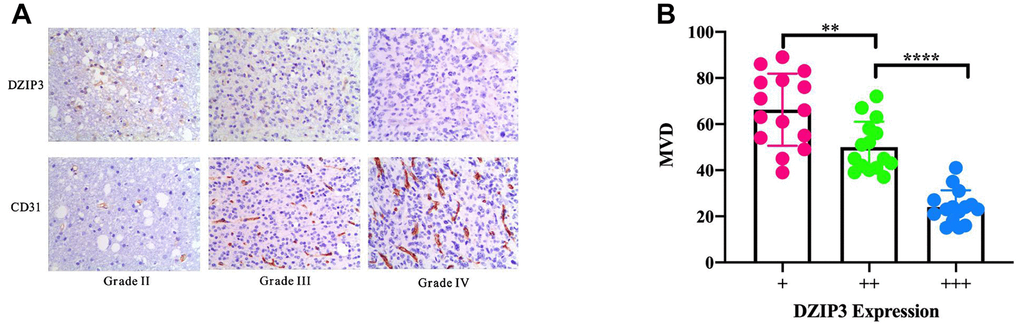
Figure 3. (A) IHC staining of DZIP3 and CD31 in different grades. (B) CD31-MVD is lower in samples with higher DZIP3 expression. **p<0.01 and ****p<0.0001.
High DZIP3 mRNA expression confers improved survival time in all grade and lower grade glioma
In the CGGA RNA-seq set (training set), glioma patients were divided into two groups based on the median expression of DZIP3. Figure 4A, 4B showed that in both all grade and lower grade gliomas, patients in higher DZIP3 expression groups possessed significantly longer survival time than lower expression counterparts (p < 0.0001). However, we cannot attain a similar tendency in the GBM group (Figure 4C, p > 0.05). Furthermore, the results above in the glioma patients from GSE 16011 array set were validated (Figure 4D–4F). Then, Cox regression analysis verified the independence of the clinical prognostic significance of DZIP3 in glioma after adjusting age at diagnosis and WHO grade and IDH1 status in both all grade and lower grade glioma in CGGA RNA-seq cohort (Table 1). These results indicated that DZIP3 plays a crucial biological function in gliomas, especially lower-grade glioma.
Table 1. Univariate and multivariate Cox analysis in CGGA RNA-seq cohort.
| Characteristic | Univariate analysis | Multivariate analysis | ||
| HR (95% CI) | P-value | HR (95% CI) | P-value | |
| All grade | ||||
| Age at diagnosis | 1.039 (1.023-1.056) | < 0.0001 | 0.999 (0.983-1.015) | 0.897 |
| WHO grade | 3.569 (2.751-4.629) | < 0.0001 | 2.791 (2.078-3.749) | < 0.0001 |
| Gender | 1.204 (0.837-1.731) | 0.316 | ||
| IDH1 status | 0.253 (0.172-0.370) | < 0.0001 | 0.574 (0.364-0.905) | 0.017 |
| DZIP3 expression | 0.824 (0.780-0.870) | < 0.0001 | 0.927 (0.877-0.980) | 0.007 |
| Lower grade | ||||
| Age at diagnosis | 1.047 (1.016-1.078) | 0.003 | 1.017 (0.987-1.047) | 0.274 |
| WHO grade | 5.921 (3.137-11.176) | < 0.0001 | 3.511 (1.734-7.110) | < 0.0001 |
| Gender | 1.027 (0.568-1.857) | 0.93 | ||
| I DH1 status | 0.274 (0.153-0.491) | < 0.0001 | 0.445 (0.227-0.873) | 0.018 |
| DZIP3 expression | 0.765 (0.692-0.844) | < 0.0001 | 0.855 (0.776-0.943) | 0.002 |
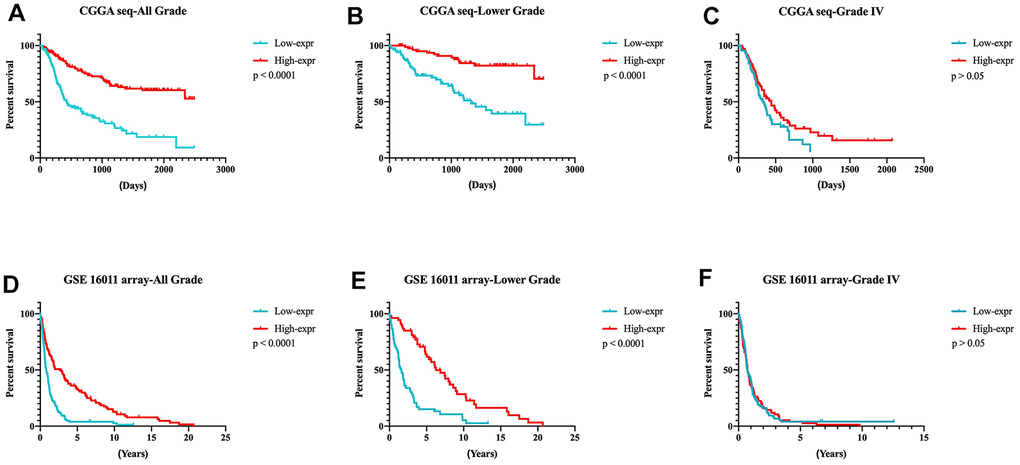
Figure 4. Prognostic significance of DZIP3 in all-grade, lower-grade and GBM groups. (A–C) CGGA RNA-seq cohort; (D–F) GSE 16011 array cohort.
DZIP3 could further stratify IDH1 wild-type lower-grade glioma
According to the results mentioned earlier, higher DZIP3 expression indicated a prolonged survival time in all grade and lower grade glioma, but not in GBM. Next, we focused on 356 lower-grade glioma patients for further research. In lower-grade glioma, DZIP3 expression could predict the survival time in IDH1 mutant and IDH1 wild-type subgroups (Figure 5A–5D). IDH1 mutation was a key genetic event that mainly occurs in lower grade gliomas, while the IDH1 wild-type phenotype constituted the GBM group. IDH1 wild-type lower-grade glioma was a group of patients with high heterogeneity. As shown in Figure 6A, DZIP3 classified IDH1 wild-type lower-grade glioma into GBM-like and IDH1 mutation lower grade-like groups with similar survival tendencies. Also, these results could be validated in GSE 16011 array cohort (Figure 6B), indicating that the introduction of DZIP3 could provide the basis for accurate diagnosis and treatment of glioma, especially lower-grade glioma.
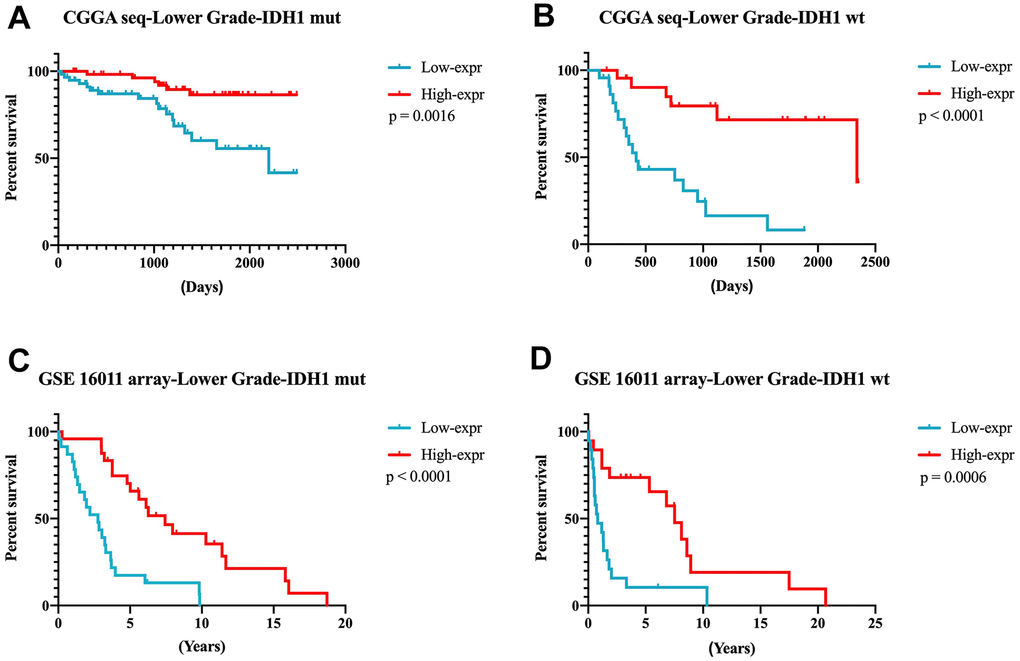
Figure 5. DZIP3 expression could predict the survival time in IDH1-MUT and IDH1-WT subgroups in lower-grade gliomas. (A, B) CGGA RNA-seq cohort; (C, D) GSE 16011 array cohort.
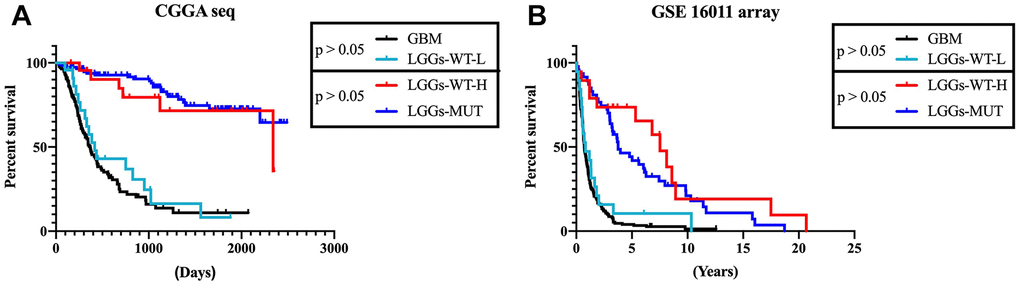
Figure 6. DZIP3 could stratify lower-grade IDH1-WT gliomas patients into IDH1-MUT-like and GBM-like groups. (A) CGGA RNA-seq cohort; (B) GSE 16011 array cohort.
Discussion
The prognosis of patients remains extremely poor, despite aggressive therapy [15]. WHO 2007 glioma classification mainly relies on the morphology and IHC evaluation [16]. Malignant gliomas can be divided into three grades: WHO grade II–IV. WHO grade II and III gliomas are usually described as lower-grade glioma, whereas GBM refers to WHO grade IV glioma [17]. According to 2016 WHO criteria, except for diffuse astrocytic and oligodendroglial tumors, the IDH1 genotypes and status of chromosome arms 1p and 19q have been incorporated into the classification of gliomas [3, 17]. The glioma patients with tumors carrying a mutation in IDH1 and chromosome 1p/19q co-deletion could have a prolonged survival [18, 19]. The classification criteria published in 2016 were more precise than those in 2007. Ueki et al. indicated that IDH-mutated astrocytomas with 19q-loss constitute a subgroup with improved prognosis [20]. Yang et al. suggested that IDH1 and TERT mutation in lower-grade glioma patients is responsible for prolonged survival time compared with other groups [21]. Additionally, the development of sequencing technology in various studies has focused on the stratification of glioma. Based on gene expression profiles, Wu et al. developed a novel molecular classification of IDH-mutant GBM with different clinical characteristics [11]. Moreover, in lower grade glioma, Cheng et al. established a five-gene signature to stratify IDH1-mutant lower-grade glioma with a distinct prognosis [9]. However, only a few studies have focused on IDH1 wild-type lower-grade glioma patients.
In the current study, we utilized two independent cohorts to deduce the role of DZIP3 in glioma. DZIP3, also known as DAZ-interacting zinc finger protein 3, is an RNA-binding RING-type ubiquitin ligase and involved in various biological functions in glioma. Firstly, we tested the expression of DZIP3 between different WHO grades and IDH1 statuses and indicated that DZIP3 expression is lower in GBM and IDH1 wild-type groups. Moreover, based on DZIP3 expression, we could reclassify the IDH1 wild-type lower-grade glioma (Figure 7). Lower-grade glioma patients with lower DZIP3 expression have similar survival time with GBM, while the survival time of the other patients is similar to that of the IDH1 mutation group. In addition, we analyzed the DZIP3 correlated genes using DAVID software. Biomedical analyses revealed that DZIP3 is significantly related to angiogenesis, which is the formation of new blood vessels and a prominent hallmark of glioma [22]. This neovascularization allows for increased oxygen and nutrients to be distributed to rapidly dividing tumor cells [23]. Recent studies designated CD31 (also known as PECAM) as the optimal marker of neoangiogenesis [24]. Herein, we verified the relationship between DZIP3 protein expression and CD31-MVD via IHC method, the results indicated that CD31-MVD was lower in higher DZIP3 protein expression samples.
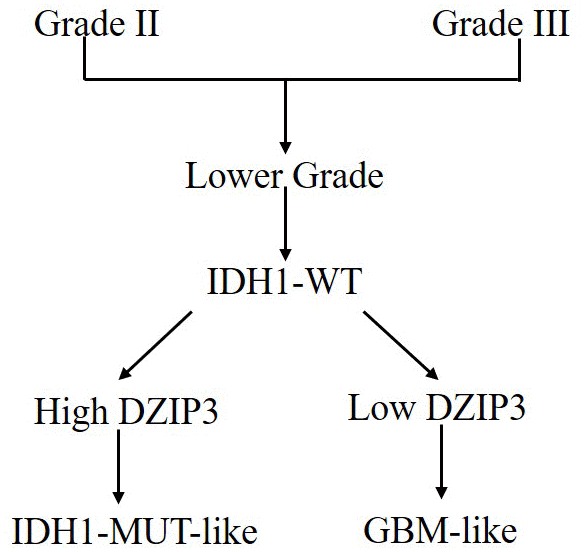
Figure 7. IDH1-MUT-like and GBM-like model for classification of lower-grade IDH1-WT gliomas based on DZIP3 expression.
Taken together, DZIP3 exerts a considerable effect on clinical and molecular conditions via the angiogenesis pathway. Thus, assessing DZIP3 might aid in precise diagnosis of glioma, especially lower grade glioma, and provide a new target for glioma diagnosis and treatment. Therefore, using DZIP3 target medicine for the treatment of glioma warrants for further investigation.
Materials and Methods
Patients and samples
A total of 590 patients with WHO grade II–IV glioma (325 cases from the CGGA RNA-seq cohort and 265 cases from GSE 16011 array cohort) were included in this study. The CGGA RNA-seq cohort (http://www.cgga.org.cn) comprised of 109 grade II, 72 grade III, and 144 grade IV patients. Moreover, 24 grade II, 85 grade III, and 159 grade IV patients constituted the GSE 16011 array cohort (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE16011). The histological diagnoses were determined according to the WHO 2007 criteria [25]. As described previously, the overall survival (OS) time was defined as the period from surgery to death [26]. The present study was approved by the Ethics Committee of Beijing Ditan Hospital and Beijing Tiantan Hospital.
Immunohistochemistry for DZIP3 and CD31 expression
The expression of DZIP3 and CD31 proteins was assessed by IHC. The tumor tissues excised during the operation were immediately placed in 10% formalin for fixation, followed by dehydration, paraffin embedding, and sectioning. Anti-DZIP3 (Abcam) and CD31 (Zhong Shan, Beijing) antibodies were used at a dilution of 1:100. These results were confirmed by two neuropathologists individually. DZIP3 protein expression (semi-quantitative scoring) = expression intensity × expression area. Expression intensity was scored using a 4-point scale from 0 to 3. Expression area was scored using a 5-point scale from 0 to 4 [27]. Moreover, 0-6, 7-9 and 10-12 represented +, ++, +++ respectively.
Counts of microvessel density (MVD)
MVD was based on CD31 immunoreactivity. We attained microscopy of CD31 at low magnification (×20) and three tumor areas with the highest density of distinctly highlighted microvessel (hot spot) within each section were selected for quantitation of MVD. Counts were performed on these fields in the hot spot at ×200 magnification [28].
Bioinformatic analysis
Pearson’s correlation analysis explored the genes related to DZIP3 through the R language. Negatively-related genes with correlation coefficient >0.4 were input into GO analysis in DAVID Bioinformatics Resources 6.8 to understand the biological functions of DZIP3.
Statistical analysis
R language, GraphPad Prism 5, and SPSS 22.0 were used for statistical analyses. Student’s t-test was used to explore the diversity of DZIP3 expression between different clinical and molecular groups. Kaplan–Meier analysis was performed to estimate the survival time of different subgroups, and a log-rank test assessed the prognostic differences. Univariate and multivariate Cox regression analyses were used to determine that DZIP3 is an independent factor to predict the survival time. p < 0.05 was considered as statistically significant [29].
Author Contributions
TYL and XGZ conceived and designed the experiments. PLL performed the experiments. GY and FW analyzed the data. TYL contributed the analysis tools. ESF and PW wrote the paper. All authors gave final approval of the manuscript.
Acknowledgments
We thank the CGGA and GSE 16011 network for sharing large amounts of data. We thank Beijing Ditan Hospital Specimen Bank Project for specimen storage.
Conflicts of Interest
The authors report no conflicts of interest in this work.
Funding
This work is supported by grant from Beijing Medical Management Bureau Cultivation Plan (PX2018079) and the National Key Sci-Tech Special Project of China (No. 2018ZX10302207) and the National Natural Science Foundation of China (NO. 81871013) and Beijing Tiantan Hospital Young Scientist Program (YSP 201705).
References
- 1. Hu H, Mu Q, Bao Z, Chen Y, Liu Y, Chen J, Wang K, Wang Z, Nam Y, Jiang B, Sa JK, Cho HJ, Her NG, et al. Mutational landscape of secondary glioblastoma guides MET-targeted trial in brain tumor. Cell. 2018; 175:1665–78.e18. https://doi.org/10.1016/j.cell.2018.09.038 [PubMed]
- 2. Liu X, Li Y, Li S, Fan X, Sun Z, Yang Z, Wang K, Zhang Z, Jiang T, Liu Y, Wang L, Wang Y. IDH mutation-specific radiomic signature in lower-grade gliomas. Aging (Albany NY). 2019; 11:673–96. https://doi.org/10.18632/aging.101769 [PubMed]
- 3. Louis DN, Perry A, Reifenberger G, von Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD, Kleihues P, Ellison DW. The 2016 world health organization classification of tumors of the central nervous system: a summary. Acta Neuropathol. 2016; 131:803–20. https://doi.org/10.1007/s00401-016-1545-1 [PubMed]
- 4. Behnan J, Finocchiaro G, Hanna G. The landscape of the mesenchymal signature in brain tumours. Brain. 2019; 142:847–66. https://doi.org/10.1093/brain/awz044 [PubMed]
- 5. Guan X, Zhang C, Zhao J, Sun G, Song Q, Jia W. CMTM6 overexpression is associated with molecular and clinical characteristics of malignancy and predicts poor prognosis in gliomas. EBioMedicine. 2018; 35:233–43. https://doi.org/10.1016/j.ebiom.2018.08.012 [PubMed]
- 6. Akgül S, Patch AM, D’Souza RC, Mukhopadhyay P, Nones K, Kempe S, Kazakoff SH, Jeffree RL, Stringer BW, Pearson JV, Waddell N, Day BW. Intratumoural heterogeneity underlies distinct therapy responses and treatment resistance in glioblastoma. Cancers (Basel). 2019; 11:190. https://doi.org/10.3390/cancers11020190 [PubMed]
- 7. Eckel-Passow JE, Lachance DH, Molinaro AM, Walsh KM, Decker PA, Sicotte H, Pekmezci M, Rice T, Kosel ML, Smirnov IV, Sarkar G, Caron AA, Kollmeyer TM, et al. Glioma groups based on 1p/19q, IDH, and TERT promoter mutations in tumors. N Engl J Med. 2015; 372:2499–508. https://doi.org/10.1056/NEJMoa1407279 [PubMed]
- 8. Binder H, Willscher E, Loeffler-Wirth H, Hopp L, Jones DT, Pfister SM, Kreuz M, Gramatzki D, Fortenbacher E, Hentschel B, Tatagiba M, Herrlinger U, Vatter H, et al. DNA methylation, transcriptome and genetic copy number signatures of diffuse cerebral WHO grade II/III gliomas resolve cancer heterogeneity and development. Acta Neuropathol Commun. 2019; 7:59. https://doi.org/10.1186/s40478-019-0704-8 [PubMed]
- 9. Cheng W, Ren X, Zhang C, Cai J, Han S, Wu A. Gene expression profiling stratifies IDH1-mutant glioma with distinct prognoses. Mol Neurobiol. 2017; 54:5996–6005. https://doi.org/10.1007/s12035-016-0150-6 [PubMed]
- 10. Kloosterhof NK, Bralten LB, Dubbink HJ, French PJ, van den Bent MJ. Isocitrate dehydrogenase-1 mutations: a fundamentally new understanding of diffuse glioma? Lancet Oncol. 2011; 12:83–91. https://doi.org/10.1016/S1470-2045(10)70053-X [PubMed]
- 11. Wu F, Chai RC, Wang Z, Liu YQ, Zhao Z, Li GZ, Jiang HY. Molecular classification of IDH-mutant glioblastomas based on gene expression profiles. Carcinogenesis. 2019; 40:853–60. https://doi.org/10.1093/carcin/bgz032 [PubMed]
- 12. Inoue D, Aihara H, Sato T, Mizusaki H, Doiguchi M, Higashi M, Imamura Y, Yoneda M, Miyanishi T, Fujii S, Okuda A, Nakagawa T, Ito T. Dzip3 regulates developmental genes in mouse embryonic stem cells by reorganizing 3D chromatin conformation. Sci Rep. 2015; 5:16567. https://doi.org/10.1038/srep16567 [PubMed]
- 13. Oikawa D, Shiota M, Tokunaga F, Wanibuchi H. Generation of rat monoclonal antibodies specific for DZIP3. Monoclon Antib Immunodiagn Immunother. 2018; 37:153–57. https://doi.org/10.1089/mab.2018.0005 [PubMed]
- 14. Mikkelsen VE, Stensjøen AL, Granli US, Berntsen EM, Salvesen Ø, Solheim O, Torp SH. Angiogenesis and radiological tumor growth in patients with glioblastoma. BMC Cancer. 2018; 18:862. https://doi.org/10.1186/s12885-018-4768-9 [PubMed]
- 15. Hu Y, Zhang M, Tian N, Li D, Wu F, Hu P, Wang Z, Wang L, Hao W, Kang J, Yin B, Zheng Z, Jiang T, et al. The antibiotic clofoctol suppresses glioma stem cell proliferation by activating KLF13. J Clin Invest. 2019; 129:3072–85. https://doi.org/10.1172/JCI124979 [PubMed]
- 16. Aibaidula A, Chan AK, Shi Z, Li Y, Zhang R, Yang R, Li KK, Chung NY, Yao Y, Zhou L, Wu J, Chen H, Ng HK. Adult IDH wild-type lower-grade gliomas should be further stratified. Neuro Oncol. 2017; 19:1327–37. https://doi.org/10.1093/neuonc/nox078 [PubMed]
- 17. Wang N, Xie SY, Liu HM, Chen GQ, Zhang WD. Arterial spin labeling for glioma grade discrimination: correlations with IDH1 genotype and 1p/19q status. Transl Oncol. 2019; 12:749–56. https://doi.org/10.1016/j.tranon.2019.02.013 [PubMed]
- 18. Deluche E, Bessette B, Durand S, Caire F, Rigau V, Robert S, Chaunavel A, Forestier L, Labrousse F, Jauberteau MO, Durand K, Lalloué F. CHI3L1, NTRK2, 1p/19q and IDH status predicts prognosis in glioma. Cancers (Basel). 2019; 11:544. https://doi.org/10.3390/cancers11040544 [PubMed]
- 19. Núñez FJ, Mendez FM, Kadiyala P, Alghamri MS, Savelieff MG, Garcia-Fabiani MB, Haase S, Koschmann C, Calinescu AA, Kamran N, Saxena M, Patel R, Carney S, et al. IDH1-R132H acts as a tumor suppressor in glioma via epigenetic up-regulation of the DNA damage response. Sci Transl Med. 2019; 11:eaaq1427. https://doi.org/10.1126/scitranslmed.aaq1427 [PubMed]
- 20. Otani R, Uzuka T, Higuchi F, Matsuda H, Nomura M, Tanaka S, Mukasa A, Ichimura K, Kim P, Ueki K. IDH-mutated astrocytomas with 19q-loss constitute a subgroup that confers better prognosis. Cancer Sci. 2018; 109:2327–35. https://doi.org/10.1111/cas.13635 [PubMed]
- 21. Yang P, Cai J, Yan W, Zhang W, Wang Y, Chen B, Li G, Li S, Wu C, Yao K, Li W, Peng X, You Y, et al, and CGGA project. Classification based on mutations of TERT promoter and IDH characterizes subtypes in grade II/III gliomas. Neuro Oncol. 2016; 18:1099–108. https://doi.org/10.1093/neuonc/now021 [PubMed]
- 22. Keller S, Schmidt MH. EGFR and EGFRvIII promote angiogenesis and cell invasion in glioblastoma: combination therapies for an effective treatment. Int J Mol Sci. 2017; 18:1295. https://doi.org/10.3390/ijms18061295 [PubMed]
- 23. Beyer S, Fleming J, Meng W, Singh R, Haque SJ, Chakravarti A. The role of miRNAs in angiogenesis, invasion and metabolism and their therapeutic implications in gliomas. Cancers (Basel). 2017; 9:85. https://doi.org/10.3390/cancers9070085 [PubMed]
- 24. Sharma S, Sharma MC, Sarkar C. Morphology of angiogenesis in human cancer: a conceptual overview, histoprognostic perspective and significance of neoangiogenesis. Histopathology. 2005; 46:481–89. https://doi.org/10.1111/j.1365-2559.2005.02142.x [PubMed]
- 25. Zhao Z, Zhang KN, Chai RC, Wang KY, Huang RY, Li GZ, Wang YZ, Chen J, Jiang T. ADAMTSL4, a secreted glycoprotein, is a novel immune-related biomarker for primary glioblastoma multiforme. Dis Markers. 2019; 2019:1802620. https://doi.org/10.1155/2019/1802620 [PubMed]
- 26. Yang P, Liang T, Zhang C, Cai J, Zhang W, Chen B, Qiu X, Yao K, Li G, Wang H, Jiang C, You G, Jiang T. Clinicopathological factors predictive of postoperative seizures in patients with gliomas. Seizure. 2016; 35:93–99. https://doi.org/10.1016/j.seizure.2015.12.013 [PubMed]
- 27. Feng E, Liang T, Wang X, Du J, Tang K, Wang X, Wang F, You G. Correlation of alteration of HLA-F expression and clinical characterization in 593 brain glioma samples. J Neuroinflammation. 2019; 16:33. https://doi.org/10.1186/s12974-019-1418-3 [PubMed]
- 28. Wu ZB, Cai L, Lin SJ, Leng ZG, Guo YH, Yang WL, Chu YW, Yang SH, Zhao WG. Heat shock protein 47 promotes glioma angiogenesis. Brain Pathol. 2016; 26:31–42. https://doi.org/10.1111/bpa.12256 [PubMed]
- 29. Zhang W, Liu Y, Hu H, Huang H, Bao Z, Yang P, Wang Y, You G, Yan W, Jiang T, Wang J, Zhang W. ALDH1A3: a marker of mesenchymal phenotype in gliomas associated with cell invasion. PLoS One. 2015; 10:e0142856. https://doi.org/10.1371/journal.pone.0142856 [PubMed]