



Introduction
Aging is a deteriorative process with decline in physical and functional capabilities. Aging increases the risk of diabetes mellitus (DM), cardiovascular diseases (CVD) and sarcopenia, particularly in the people with conditions such as long-term inflammation and weak responds to oxidative stress. As seen in skeletal muscles the process of aging is associated with weakening of biochemical pathways of oxidative phosphorylation and respiration in mitochondria [1-3]. During the process of aging a progressive deterioration in mitochondrial function results in the accumulation of oxidative stress causing senescence and modulations in the levels of longevity associated proteins such as SIRT1 [4,5].
SIRT1 is a redox-sensitive enzyme that regulates various cellular events including apoptosis, cell survival, endocrine signaling, chromatin remodeling, and gene transcription [6]. Activation of SIRT1 enhances mitochondrial biogenesis via its downstream protein peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) thereby replenishing the metabolic signaling pathways and suppressing inflammatory signaling [7,8]. PGC-1α is a strong transcriptional co-activator of several transcriptional factors and nuclear receptors and it specifically controls metabolic pathways particularly, the oxidative metabolism. One of the major roles of PGC-1α is in the biogenesis of mitochondria and associated oxidative phosphorylation.
Mitochondrial biogenesis and oxidative phosphorylation in muscle, heart and fat tissue can be increased by enhancing the expression of PGC-1α [9-12]. It is also notable that while generally exercise training prevents aging-induced reduction of mitochondrial proteins and associated cellular apoptosis by providing antioxidant benefits, PGC-1α knock-out mice do not display antioxidant effects following exercise [13-15]. Similarly, acute exercise training is known to trigger the activation of the dimeric transcription factor-nuclear factor-kappaB (NFκB). NFκB is activated by various stimulants, including TNF-α. Activation of TNF-α receptor triggers IκB kinase (IKK) to phosphorylate IκB and thereby results in the release of P50/P65 subunits of NFκB complex to enable NFκB nuclear translocation [16,17]. While most studies elucidate these events in skeletal muscles, the long-term implications of exercise training in the longevity mechanisms of aging heart are not clear yet.
Long-term D-galactose IP injection is used to generate animal models for aging studies and they reproduce aging associated oxidative stress [18-21]. In this study, the cardiac effects of long-term exercise on D-galactose induced aging in Sprague-Dawley (SD) rats have been explored.
Results
Exercise training protected cardiac characteristics
Exercise training in control rats and aging rats resulted in a notable reduction in the bodyweight however, the differences weren’t statistically significant. Meanwhile, aging or exercise training did not show any considerable change in cardiac features such as whole heart weight (WHW) and Left ventricle weight (LVW), but the WHW and LVW normalized with body weight showed significant differences (Table 1). The exercise training groups in general showed an increase in WHW/Body weight (BW) and LVW/BW when compared to their respective groups without exercise. Further interpretation of the results shows that exercise training causes a reduction in the body weight of normal as well as induced aging group rats without causing any considerable effect in the heart.
Table 1. Cardiac characteristics of the experimental groups.
| Control (n=9) | Exercise (n=9) | Aging (n=9) | Aging + exercise (n=9) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Body weight (BW, g) | 527.33±66.41 | 485.80±66.41 | 548.67±49.11 | 484.17±37.32# | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tibia (mm) | 46.13±0.34 | 46.62±0.77 | 47.57±0.58 | 46.78±0.17 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Whole Heart weight (WHW, g) | 1.49±0.14 | 1.63±0.32 | 1.59±0.09 | 1.64±0.17 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Left ventricle weight (LVW, g) | 1.03±0.10 | 1.11±0.22 | 1.13±0.06 | 1.13±0.11 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| WHW/ BW (g) | 2.89±0.21 | 3.36±0.18* | 3.03±0.10 | 3.36±0.33# | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LVW/BW (g) | 2±0.16 | 2.29±0.10* | 2.16±0.10 | 2.31±0.18# | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HW/Tibia (mg/mm) | 32.21±2.85 | 34.91±6.22 | 33.50±2.24 | 35±3.10 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HW/Tibia (mg/mm) | 22.27±2.07 | 23.84±4.26 | 23.92±1.56 | 24.11±2.11 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Values are means ± SD among SD rats. BW, body weight; HW, whole heart weight; LVW, left ventricular weight. *P<0.05 indicates significant differences with respect to control group. # P<0.05 indicates significant differences between aging. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Effect of exercise on the histological changes in aging hearts
After eight-weeks of swimming exercise the hearts were excised and the effects of exercise training on the histological changes were analyzed by H & E staining. Heart tissue sections from the aged rats groups (A) showed thinning of the heart muscle but the exercise trained rats (E and AE) showed thicker ventricular walls, showing phenomenon of physiological hypertrophy, when compared to the control (C) and aging (A) groups. Further analysis showed that D-galactose induced aging caused a sever disarrangement in cardiac architecture. The cardiomyocytes in aging group were disordered and showed large interstitial spaces (Figure 1). In contrary, the cardiomyocyte were orderly arranged in groups that underwent swimming exercise (E and AE).
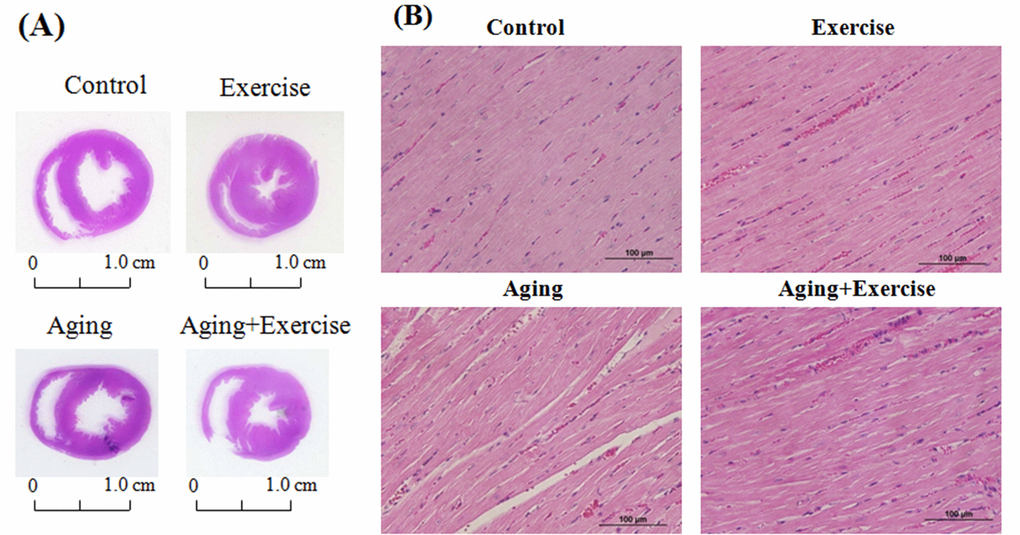
Figure 1. Hematoxylin and eosin stain (H&E stain) showing the cardiac tissue architecture. Representative histopathological analysis of cardiac sections of the left ventricles stained with hematoxylin and eosin. The hematoxylin colors basophilic structures blue-purple and the eosin colors eosinophilic structures in bright pink. The images of myocardial architecture were magnified at 400X.
Exercise training suppressed mediators of inflammation in aging hearts
The western blotting assay was used to assess the effect of swimming exercise on inflammation associated proteins and the results show that cardiac TNF-R, TNF-α, iNOS and COX-2 proteins expressions did not show any significant difference in the control rats (C) and the exercise training (E) rats (Figure 2). As expected, D-galactose induced aging rats (A) showed a significant increase in the levels of TNF-R, TNF-α, pNFκB and COX-2 levels when compared to that of the control (C). Interestingly exercise training significantly reduced the levels of the inflammatory markers in the aging rats. Notably, the COX-2 expression in rat hearts was 2.25 folds in D-galactose induced-aging rats compared with control, and those aging rats subjected to swimming exercise showed only 1.33 folds to that of the control.
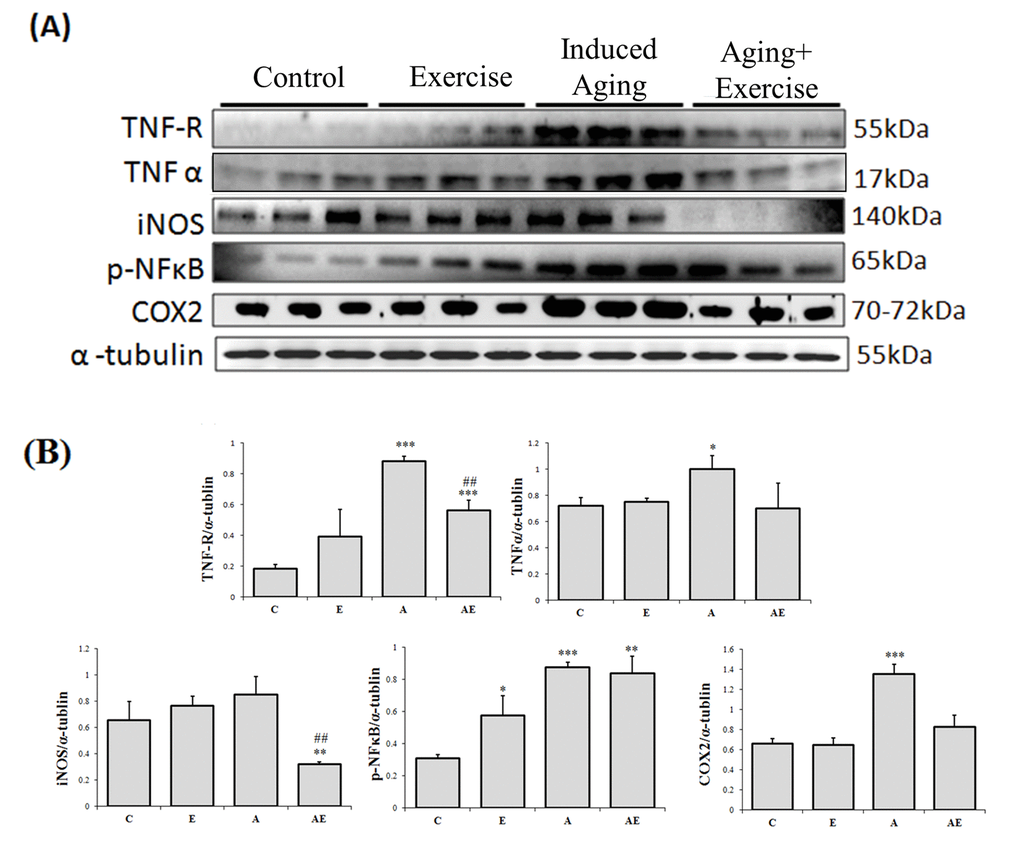
Figure 2. Effect of exercise training in TNF alpha associated Inflammatory pathway. (A) Representative Western blots showing protein products of TNF-R, TNF-α, iNOS, p-NFκB and COX-2 extracted from left ventricles of excised hearts (B) The α-tubulin was used as an internal control. (C) Densitometry bars showing the relative protein levels represented by mean values ± SEM. *P< 0.05, **P< 0.01, ***P< 0.001significant differences with respect to control group. #P<0.05, ##P<0.01, ###P< 0.001 significant differences with respect to aging group.
Exercise training enhances longevity
Analysis on the levels of AMPK and its downstream cellular longevity associated proteins revealed that exercise training in normal rats elevated the levels of pFOXO3a, of pFOXO3a, levels (figure 3). Meanwhile aging rats showed further elevation of pFOXO3a and pAMPKα1 levels and reduced levels of SIRT1, PGC-1α, PPARα and SOD1 however exercise training caused reversion in the levels of pFOXO3a, SIRT1 and PGC-1α and there was no notable difference in the levels of pAMPKα1, PPARα and SOD1 when compared to the aging group.
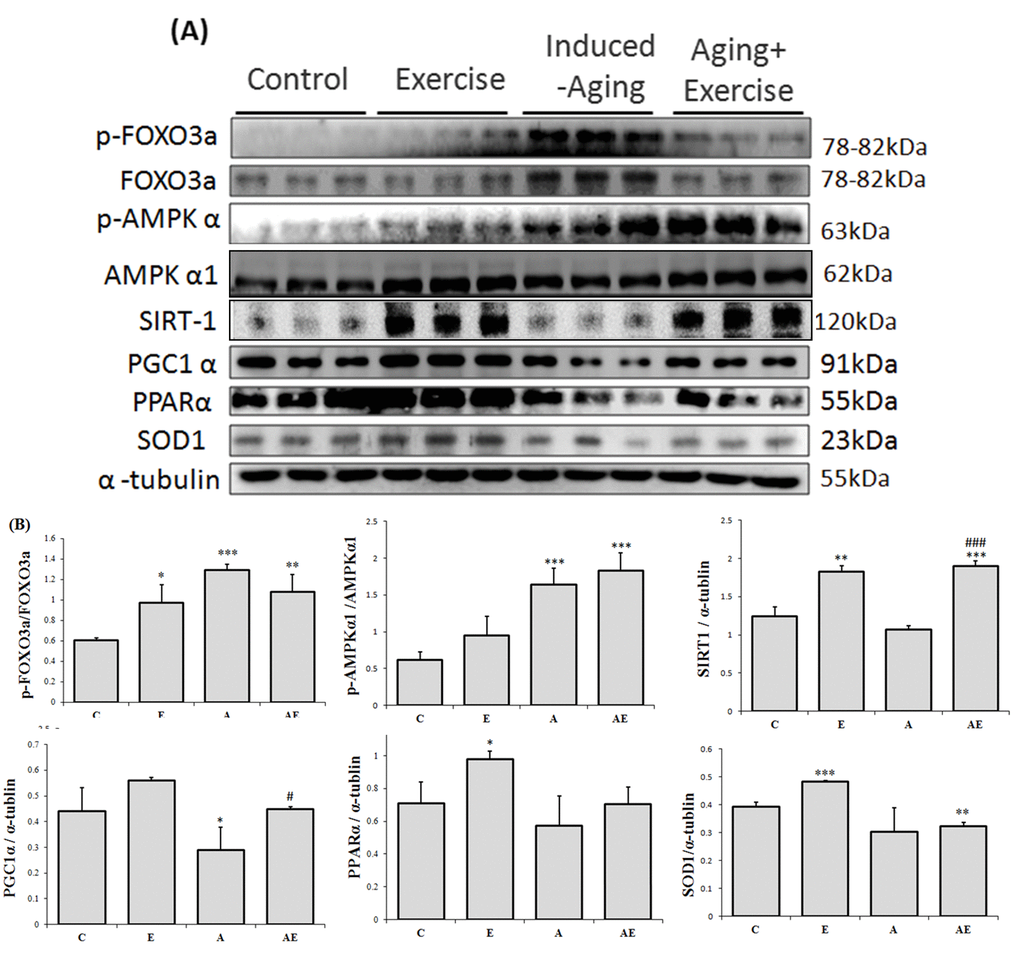
Figure 3. Effect of exercise training on APK associated cellular homeostasis. (A) Representative Western blots showing modulation in the levels of p-FOXO1, FOXO3a, p-FOXO3a, p-AMPKα1, AMPKα1, SIRT1, PGC-1α, PPARα, and SOD-1 extracted from the left ventricles of excised hearts. The α-tubulin was used as an internal control. (B) Densitometric analysis showing relative changes in protein levels represented by mean values ± SEM. *P< 0.05, **P< 0.01, ***P< 0.001significant differences with respect to control group. #P<0.05, ##P<0.01, ###P< 0.001 significant differences with respect to aging group.
Discussion
Previous studies show that various conditions such as hypertension, obesity and diabetes cause cardiac abnormalities in rats. Inflammatory response associated with these conditions trigger cardiac hypertrophy and apoptosis that subsequently result in cardiac dysfunction. Several alternative strategies have been shown to provide cardio-protection against various pathological conditions [22-29]. Exercise training is also known to provide cardio-protection in conditions such as ischemia-reperfusion injury [30,31].
The biochemical and physiological changes in the D-galactose induced aging rat include increase in senescence, oxidative stress, mitochondrial dysfunction and cardiac apoptosis therefore the model very much represents the natural aging animal models [32]. Our results also show that aging group undergoes a 6% increase in whole heart weight and 9% increase in left-ventricle (LV) weight. Increasing whole heart and left-ventricle weight are common phenomenon associated with hypertrophy in conditions such as high blood pressure, obesity and aging [33-35]. Thus, the effects of D-galactose induced aging in heart could be correlated as a potential risk factor of cardiomyopathy. Exercise training is also known to improve heart function from cardiomyopathy conditions as reported by Boyne et al., 2013 [36]. Further, the reduction of the LV weight ratio observed in myocardial infarction animal models was found to be improved with respect to cardiac function and deterioration in cardiac remodeling. The effects of exercise were also correlated with improvements in the inflammatory profile in the chronic heart failure animal models [37]. Exercise training has been shown to provide anti-inflammatory effects as inferred from upregulated NFκB expression and endothelial vascular relaxation [38]. Our results show that expression levels of inflammatory markers such as TNF-α, NFκB, COX-2 and iNOS were steadily increased in aging group but were effectively reverted in exercise training groups therefore it can be interpreted that exercise training could potentially regulate and maintain the cellular homeostasis with respect to inflammation and oxidative stress (Figure 4) [39]. Interestingly, the compensative expression of p-NFκB was observed in D-galactose induced aging when combined with long-term exercise training. The possible reasons might be due to the variation in time and frequency of the exercise training [40].
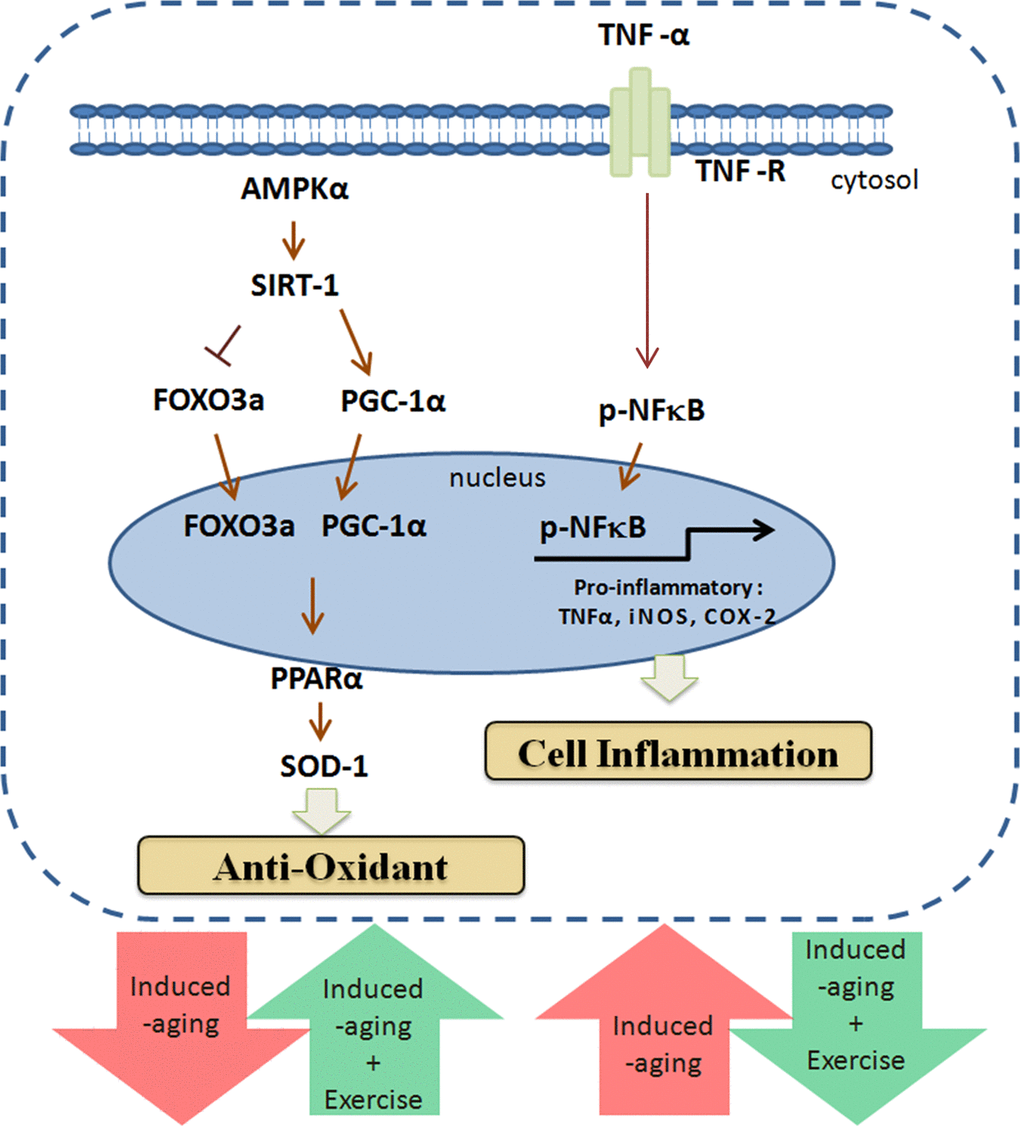
Figure 4. Exercise enhances metabolic adaptation and attenuates inflammation in aging hearts. Long-term exercise training enhances SIRT1, PGC-1α and AMPK in aging hearts to provide protection against aging associated damages.
SIRT1, a deacetylase is involved in the metabolic adaptation of muscle tissue to endurance exercise possibly due to the imbalances in the cytosolic NAD+/NADH ratio caused by muscle contraction [41]. Increase in SIRT1 levels facilitate such metabolic adaptation of muscles [42] and also regulate age-associated changes through various mechanisms that include enhancing the mitochondria biogenesis by deacetylation and activation of PGC-1α, containing oxidative stress and promoting survival signals by activation of FOXO family proteins, reducing apoptosis and proliferation and alleviating pro-inflammatory response of aging [43-45]. Exercise also intensifies the contraction of cardiac muscles which may facilitate SIRT1 upregulation and our results also correlates exercise training with elevated SIRT1 levels.
Aging is associated with increasing stressful conditions like elevated ROS levels and inflammation which effectively trigger cell death which may result in the disarrangement in the cardiomyocyte arrangement causing reduction in the efficiency of the heart muscles [46]. The histological assessment performed in this study shows that exercise training improved the cardiomyocyte arrangement reflecting an improvement in the cardiac function.
In oxidative stress conditions FOXO3a that exist in nucleus induces the expression of inflammatory proteins. The deacetylase SIRT1 removes the acetyl group on the FOXO3a and attenuates its function, thereby protects the cell and also stabilizes the nuclear DNA [47]. The elevated levels of FOXO3a in D-galactose induced aging rat observed in this study were reduced with exercise training which was correlated with elevated SIRT1 levels which potentially contributes to deacetylase activity in cardiac cells.
The energy consumption for exercise is usually subject to glucose transport through AMPK independent pathway. In our results, the AMPK expression was higher in normal rats with exercise training but, interestingly, the p-AMPK level was higher in aging rats and was further increased in the aging rats with exercise training. The results suggest an energy demand that existed in D-galactose induced aging rat hearts, and the AMPK independent pathway was not regulated after exercise training. AMPK is an energy sensor and since it also increases the intracellular NAD+ levels its activity is correlated with SIRT1-enhancment [48]. However since SIRT1 can also activate the predominant AMPK kinase LKB-1, it is also identified as an upstream activator of AMPK. Therefore in certain conditions the effects of exercise are mediated through LKB1/AKT activation and in some they are mediated through SIRT1 induced deacetylation. In this study, although a correlation exists in the levels of AMPK and SIRT1, the active pAMPK levels in the aging groups seem to be independent of SIRT1 levels. Therefore our results also show that, although when AMPK and SIRT1 levels are generally inter-dependable [49] they are independent in some conditions such as aging.
In conclusion, our results demonstrate that long-term swimming exercise can enhance SIRT1, PGC-1α and AMPK in cardiac cells of aging rats and thereby facilitate metabolic adaptation in the heart and attenuates the activation of pro-inflammatory mediators and collectively ameliorates aging induced hypertrophic effects.
Materials and Methods
Animal models
Three months old male Sprague Dawley (SD) rats were purchased from BioLASCO (Taipei, Taiwan). The rats were housed in cages with temperature maintained at 24 ± 2 °C, 55 ± 10% of humidity and 12 h light-dark cycle and were provided a standard laboratory diet (Lab Diet 5001; PMI Nutrition International Inc., Brentwood, MO, USA) and drinking water ad libitum. After one week adaptation the rats were divided into four groups: Control group (C, n=9), Exercise training group (E, n=9), Aging group (A, n=9), Aging group with Exercise training (AE, n=9). The control rats were given 0.9% saline (IP, 1 mL) and the aging induced groups (A and AE) received D-galactose (150 mg/kg body weight) for eight weeks. The swimming training protocol was performed as mentioned in other reports [50]. The Initial exercise training during the first two weeks was given for 20 min/day for five times a week. In the third week the timing of the swimming exercise training time was increased to 30 min per day and from fourth to eighth week the swimming exercise training was given for 60 min/day. The rats swam individually in a water bath (60 × 90 cm, 50 cm depth) with the water temperature maintained at 35 ± 1°C. All protocols were approved by the Institutional Animal Care and Use Committee of China Medical University, Taichung, Taiwan. The study followed the principles of laboratory animal care (NIH publication).
Protein extraction and western blotting
Heart tissue extracts obtained by homogenizing in a lysis buffer (0.05M of pH 7.4 Tris-HCl, 1 mM EDTA, 0.15 M NaCl, 1% NP-40, 0.25% deoxycholic acid) at a ratio of 100 mg tissue/1 mL buffer. The homogenates were kept on ice and the proteins in the supernatants were collected by centrifugation at 13,000 rpm for 40 min and stored at -80 °C. Protein concentrations in heart tissue extracts were determined by the Lowry protein assay. Western blotting was performed following methods mentioned in other reports with slight modification [51]. The protein samples were separated in 8%-12% SDS polyacrylamide gel electrophoresis (SDS-PAGE) under 75 V for 120 min. The separated proteins were subsequently transferred to PVDF membranes (GE healthcare limited, Buckinghamshire, UK) under 50 V for 3 h. The membranes were blocked in 3% bovine serum albumin (BSA) in tris-buffered saline and were then hybridized with primary antibodies including TNF-R (SC-1070, Santa Cruz Biotechnology, California, California, USA), TNF-α (SC-1350, Santa Cruz Biotechnology, California, California, USA), iNOS(610328, BD, New Jersey, USA), p-NFκB (#3033L, Cell Signaling, Maryland, USA) , COX-2(SC-1745, Santa Cruz Biotechnology, California, USA),α-tubulin (SC-5286, Santa Cruz Biotechnology, California, USA), p-FOXO1(#9464, Cell Signaling, Maryland, USA), FOXO3a(#2497, Cell Signaling, Maryland, USA), p-FOXO3a(#9466, Cell Signaling, Maryland, USA), p-AMPKα1(#2535, Cell Signaling, Maryland, USA), AMPKα1(SC-19128, Santa Cruz Biotechnology, California, USA), SIRT-1(SC-74465, Santa Cruz Biotechnology, California, USA), PGC-1α(AF-1817a, ABGENT, San Diego , USA ), PPAR-α(SC-9000, Santa Cruz Biotechnology, California, USA), SOD1(SC-8647, Santa Cruz Biotechnology, California, USA), Finally, the blots were hybridized with horseradish peroxidase-labeled secondary antibodies and pictures were then taken with Fujifilm LAS-3000 (GE healthcare UK limited., Buckinghamshire, UK).
Hemotoxyline and eosin staining
The embedded rat heart sections were cut into 0.2 µm thick slices and were deparaffinized by immersing in xylene and rehydrated by subjecting to graded alcohol (100%-60%). All slices were stained with hematoxylin and eosin (H&E) and photomicrographs were obtained using Zeiss Axiophot microscopes.
Statistical analysis
The results shown are means ± SD of three independent experiments. Statistical differences were evaluated by ANOVA one-way analysis of variants. p< 0.05 was considered statistically significant
Conflicts of Interest
The authors have no conflicts of interest to declare.
Funding
This study is supported in part by China Medical University and Asia University (CMU103-ASIA-15 and CMU103-ASIA-17).
References
- 1. Conley KE, Jubrias SA, Esselman PC. Oxidative capacity and ageing in human muscle. J Physiol. 2000; 526:203–10. https://doi.org/10.1111/j.1469-7793.2000.t01-1-00203.x [PubMed]
- 2. Nair KS. Aging muscle. Am J Clin Nutr. 2005; 81:953–63. https://doi.org/10.1093/ajcn/81.5.953 [PubMed]
- 3. Nunnari J, Suomalainen A. Mitochondria: in sickness and in health. Cell. 2012; 148:1145–59. https://doi.org/10.1016/j.cell.2012.02.035 [PubMed]
- 4. Kwong LK, Sohal RS. Age-related changes in activities of mitochondrial electron transport complexes in various tissues of the mouse. Arch Biochem Biophys. 2000; 373:16–22. https://doi.org/10.1006/abbi.1999.1495 [PubMed]
- 5. Tonkin J, Villarroya F, Puri PL, Vinciguerra M. SIRT1 signaling as potential modulator of skeletal muscle diseases. Curr Opin Pharmacol. 2012; 12:372–76. https://doi.org/10.1016/j.coph.2012.02.010 [PubMed]
- 6. Sakamoto J, Miura T, Shimamoto K, Horio Y. Predominant expression of Sir2alpha, an NAD-dependent histone deacetylase, in the embryonic mouse heart and brain. FEBS Lett. 2004; 556:281–86. https://doi.org/10.1016/S0014-5793(03)01444-3 [PubMed]
- 7. Waldman M, Cohen K, Yadin D, Nudelman V, Gorfil D, Laniado-Schwartzman M, Kornwoski R, Aravot D, Abraham NG, Arad M, Hochhauser E. Regulation of diabetic cardiomyopathy by caloric restriction is mediated by intracellular signaling pathways involving ‘SIRT1 and PGC-1α’. Cardiovasc Diabetol. 2018; 17:111. https://doi.org/10.1186/s12933-018-0754-4 [PubMed]
- 8. Wu Z, Puigserver P, Andersson U, Zhang C, Adelmant G, Mootha V, Troy A, Cinti S, Lowell B, Scarpulla RC, Spiegelman BM. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell. 1999; 98:115–24. https://doi.org/10.1016/S0092-8674(00)80611-X [PubMed]
- 9. Lehman JJ, Barger PM, Kovacs A, Saffitz JE, Medeiros DM, Kelly DP. Peroxisome proliferator-activated receptor gamma coactivator-1 promotes cardiac mitochondrial biogenesis. J Clin Invest. 2000; 106:847–56. https://doi.org/10.1172/JCI10268 [PubMed]
- 10. St-Pierre J, Lin J, Krauss S, Tarr PT, Yang R, Newgard CB, Spiegelman BM. Bioenergetic analysis of peroxisome proliferator-activated receptor gamma coactivators 1alpha and 1beta (PGC-1alpha and PGC-1beta) in muscle cells. J Biol Chem. 2003; 278:26597–603. https://doi.org/10.1074/jbc.M301850200 [PubMed]
- 11. Lin J, Wu H, Tarr PT, Zhang CY, Wu Z, Boss O, Michael LF, Puigserver P, Isotani E, Olson EN, Lowell BB, Bassel-Duby R, Spiegelman BM. Transcriptional co-activator PGC-1 alpha drives the formation of slow-twitch muscle fibres. Nature. 2002; 418:797–801. https://doi.org/10.1038/nature00904 [PubMed]
- 12. Arany Z, He H, Lin J, Hoyer K, Handschin C, Toka O, Ahmad F, Matsui T, Chin S, Wu PH, Rybkin II, Shelton JM, Manieri M, et al. Transcriptional coactivator PGC-1 alpha controls the energy state and contractile function of cardiac muscle. Cell Metab. 2005; 1:259–71. https://doi.org/10.1016/j.cmet.2005.03.002 [PubMed]
- 13. Leick L, Lyngby SS, Wojtaszewski JF, Pilegaard H. PGC-1alpha is required for training-induced prevention of age-associated decline in mitochondrial enzymes in mouse skeletal muscle. Exp Gerontol. 2010; 45:336–42. https://doi.org/10.1016/j.exger.2010.01.011 [PubMed]
- 14. Lanza IR, Short DK, Short KR, Raghavakaimal S, Basu R, Joyner MJ, McConnell JP, Nair KS. Endurance exercise as a countermeasure for aging. Diabetes. 2008; 57:2933–42. https://doi.org/10.2337/db08-0349 [PubMed]
- 15. Adhihetty PJ, Ljubicic V, Hood DA. Effect of chronic contractile activity on SS and IMF mitochondrial apoptotic susceptibility in skeletal muscle. Am J Physiol Endocrinol Metab. 2007; 292:E748–55. https://doi.org/10.1152/ajpendo.00311.2006 [PubMed]
- 16. Zhang Y, Moreno-Villanueva M, Krieger S, Ramesh GT, Neelam S, Wu H. Transcriptomics, NF-κB Pathway, and Their Potential Spaceflight-Related Health Consequences. Int J Mol Sci. 2017; 18:1166. https://doi.org/10.3390/ijms18061166
- 17. Mourkioti F, Rosenthal N. NF-kappaB signaling in skeletal muscle: prospects for intervention in muscle diseases. J Mol Med (Berl). 2008; 86:747–59. https://doi.org/10.1007/s00109-008-0308-4 [PubMed]
- 18. Zhong Y, Hu YJ, Chen B, Peng W, Sun Y, Yang Y, Zhao XY, Fan GR, Huang X, Kong WJ. Mitochondrial transcription factor A overexpression and base excision repair deficiency in the inner ear of rats with D-galactose-induced aging. FEBS J. 2011; 278:2500–10. https://doi.org/10.1111/j.1742-4658.2011.08176.x [PubMed]
- 19. Lu J, Zheng YL, Luo L, Wu DM, Sun DX, Feng YJ. Quercetin reverses D-galactose induced neurotoxicity in mouse brain. Behav Brain Res. 2006; 171:251–60. https://doi.org/10.1016/j.bbr.2006.03.043 [PubMed]
- 20. Li X, Zhang Y, Yuan Y, Sun Y, Qin Y, Deng Z, Li H. Protective Effects of Selenium, Vitamin E, and Purple Carrot Anthocyanins on D-Galactose-Induced Oxidative Damage in Blood, Liver, Heart and Kidney Rats. Biol Trace Elem Res. 2016; 173:433–42. https://doi.org/10.1007/s12011-016-0681-8 [PubMed]
- 21. Bo-Htay C, Palee S, Apaijai N, Chattipakorn SC, Chattipakorn N. Effects of d-galactose-induced ageing on the heart and its potential interventions. J Cell Mol Med. 2018; 22:1392–410. https://doi.org/10.1111/jcmm.13472 [PubMed]
- 22. Chang Lee SN, Ho TJ, Shibu MA, Day CH, Viswanadha VP, Lai CH, Chen YL, Hsieh DJ, Chen YS, Huang CY. Protective effects of electroacupuncture at LR3 on cardiac hypertrophy and apoptosis in hypertensive rats. Acupunct Med. 2016; 34:201–08. https://doi.org/10.1136/acupmed-2015-010782 [PubMed]
- 23. Liao HE, Shibu MA, Kuo WW, Pai PY, Ho TJ, Kuo CH, Lin JY, Wen SY, Viswanadha VP, Huang CY. Deep sea minerals prolong life span of streptozotocin-induced diabetic rats by compensatory augmentation of the IGF-I-survival signaling and inhibition of apoptosis. Environ Toxicol. 2016; 31:769–81. https://doi.org/10.1002/tox.22086 [PubMed]
- 24. Chen YF, Shibu MA, Fan MJ, Chen MC, Viswanadha VP, Lin YL, Lai CH, Lin KH, Ho TJ, Kuo WW, Huang CY. Purple rice anthocyanin extract protects cardiac function in STZ-induced diabetes rat hearts by inhibiting cardiac hypertrophy and fibrosis. J Nutr Biochem. 2016; 31:98–105. https://doi.org/10.1016/j.jnutbio.2015.12.020 [PubMed]
- 25. Hsieh YL, Shibu MA, Lii CK, Viswanadha VP, Lin YL, Lai CH, Chen YF, Lin KH, Kuo WW, Huang CY. Andrographis paniculata extract attenuates pathological cardiac hypertrophy and apoptosis in high-fat diet fed mice. J Ethnopharmacol. 2016; 192:170–77. https://doi.org/10.1016/j.jep.2016.07.018 [PubMed]
- 26. Asokan Shibu M, Kuo WW, Kuo CH, Day CH, Shen CY, Chung LC, Lai CH, Pan LF, Vijaya Padma V, Huang CY. Potential phytoestrogen alternatives exert cardio-protective mechanisms via estrogen receptors. Biomedicine (Taipei). 2017; 7:11. https://doi.org/10.1051/bmdcn/2017070204 [PubMed]
- 27. Shibu MA, Kuo CH, Chen BC, Ju DT, Chen RJ, Lai CH, Huang PJ, Viswanadha VP, Kuo WW, Huang CY. Oolong tea prevents cardiomyocyte loss against hypoxia by attenuating p-JNK mediated hypertrophy and enhancing P-IGF1R, p-akt, and p-Badser136 activity and by fortifying NRF2 antioxidation system. Environ Toxicol. 2018; 33:220–33. https://doi.org/10.1002/tox.22510 [PubMed]
- 28. Shibu MA, Agrawal DC, Huang CY. Mushrooms: A Pandora Box of Cardioprotective Phytochemicals. In: Agrawal DC, Tsay H-S, Shyur L-F, Wu Y-C and Wang S-Y, eds. Medicinal Plants and Fungi: Recent Advances in Research and Development. (Singapore: Springer Singapore), 2017; pp. 337-362.
- 29. Marthandam Asokan S, Mariappan R, Muthusamy S, Velmurugan BK. Pharmacological benefits of neferine - A comprehensive review. Life Sci. 2018; 199:60–70. https://doi.org/10.1016/j.lfs.2018.02.032 [PubMed]
- 30. Lee Y, Min K, Talbert EE, Kavazis AN, Smuder AJ, Willis WT, Powers SK. Exercise protects cardiac mitochondria against ischemia-reperfusion injury. Med Sci Sports Exerc. 2012; 44:397–405. https://doi.org/10.1249/MSS.0b013e318231c037 [PubMed]
- 31. Fiuza-Luces C, Santos-Lozano A, Joyner M, Carrera-Bastos P, Picazo O, Zugaza JL, Izquierdo M, Ruilope LM, Lucia A. Exercise benefits in cardiovascular disease: beyond attenuation of traditional risk factors. Nat Rev Cardiol. 2018; 15:731–43. https://doi.org/10.1038/s41569-018-0065-1 [PubMed]
- 32. Bo-Htay C, Palee S, Apaijai N, Chattipakorn SC, Chattipakorn N. Effects of d-galactose-induced ageing on the heart and its potential interventions. J Cell Mol Med. 2018; 22:1392–410. https://doi.org/10.1111/jcmm.13472 [PubMed]
- 33. Huang CY, Yang AL, Lin YM, Wu FN, Lin JA, Chan YS, Tsai FJ, Tsai CH, Kuo CH, Lee SD. Anti-apoptotic and pro-survival effects of exercise training on hypertensive hearts. J Appl Physiol. 2012; 112:883–91. https://doi.org/10.1152/japplphysiol.00605.2011 [PubMed]
- 34. Lee SD, Shyu WC, Cheng IS, Kuo CH, Chan YS, Lin YM, Tasi CY, Tsai CH, Ho TJ, Huang CY. Effects of exercise training on cardiac apoptosis in obese rats. Nutr Metab Cardiovasc Dis. 2013; 23:566–73. https://doi.org/10.1016/j.numecd.2011.11.002 [PubMed]
- 35. Oxenham H, Sharpe N. Cardiovascular aging and heart failure. Eur J Heart Fail. 2003; 5:427–34. https://doi.org/10.1016/S1388-9842(03)00011-4 [PubMed]
- 36. Boyne P, Dunning K, Carl D, Gerson M, Khoury J, Kissela B. High-intensity interval training in stroke rehabilitation. Top Stroke Rehabil. 2013; 20:317–30. https://doi.org/10.1310/tsr2004-317 [PubMed]
- 37. Nunes RB, Alves JP, Kessler LP, Dal Lago P. Aerobic exercise improves the inflammatory profile correlated with cardiac remodeling and function in chronic heart failure rats. Clinics (São Paulo). 2013; 68:876–82. https://doi.org/10.6061/clinics/2013(06)24 [PubMed]
- 38. El Assar M, Angulo J, Rodríguez-Mañas L. Oxidative stress and vascular inflammation in aging. Free Radic Biol Med. 2013; 65:380–401. https://doi.org/10.1016/j.freeradbiomed.2013.07.003 [PubMed]
- 39. Chung SW, Kim MK, Chung JH, Kim DH, Choi JS, Anton S, Seo AY, Park KY, Yokozawa T, Rhee SH, Yu BP, Chung HY. Peroxisome proliferator-activated receptor activation by a short-term feeding of zingerone in aged rats. J Med Food. 2009; 12:345–50. https://doi.org/10.1089/jmf.2007.0660 [PubMed]
- 40. Chen X, Tang X, Jiang X, Wang D, Peng M, Liu L. [Diagnosis and treatment of unilateral gluteal muscle contracture]. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi. 2011; 25:530–32. [PubMed]
- 41. Robergs RA, Ghiasvand F, Parker D. Biochemistry of exercise-induced metabolic acidosis. Am J Physiol Regul Integr Comp Physiol. 2004; 287:R502–16. https://doi.org/10.1152/ajpregu.00114.2004 [PubMed]
- 42. Suwa M, Nakano H, Radak Z, Kumagai S. Endurance exercise increases the SIRT1 and peroxisome proliferator-activated receptor gamma coactivator-1alpha protein expressions in rat skeletal muscle. Metabolism. 2008; 57:986–98. https://doi.org/10.1016/j.metabol.2008.02.017 [PubMed]
- 43. Yeung F, Hoberg JE, Ramsey CS, Keller MD, Jones DR, Frye RA, Mayo MW. Modulation of NF-kappaB-dependent transcription and cell survival by the SIRT1 deacetylase. EMBO J. 2004; 23:2369–80. https://doi.org/10.1038/sj.emboj.7600244 [PubMed]
- 44. Lavu S, Boss O, Elliott PJ, Lambert PD. Sirtuins--novel therapeutic targets to treat age-associated diseases. Nat Rev Drug Discov. 2008; 7:841–53. https://doi.org/10.1038/nrd2665 [PubMed]
- 45. Huang CC, Wang T, Tung YT, Lin WT. Effect of Exercise Training on Skeletal Muscle SIRT1 and PGC-1α Expression Levels in Rats of Different Age. Int J Med Sci. 2016; 13:260–70. https://doi.org/10.7150/ijms.14586 [PubMed]
- 46. Mariappan D, Niemann R, Gajewski M, Winkler J, Chen S, Choorapoikayil S, Bitzer M, Schulz H, Hescheler J, Sachinidis A. Somitovasculin, a novel endothelial-specific transcript involved in the vasculature development. Arterioscler Thromb Vasc Biol. 2009; 29:1823–29. https://doi.org/10.1161/ATVBAHA.109.190751 [PubMed]
- 47. Oellerich MF, Potente M. FOXOs and sirtuins in vascular growth, maintenance, and aging. Circ Res. 2012; 110:1238–51. https://doi.org/10.1161/CIRCRESAHA.111.246488 [PubMed]
- 48. Cantó C, Gerhart-Hines Z, Feige JN, Lagouge M, Noriega L, Milne JC, Elliott PJ, Puigserver P, Auwerx J. AMPK regulates energy expenditure by modulating NAD+ metabolism and SIRT1 activity. Nature. 2009; 458:1056–60. https://doi.org/10.1038/nature07813 [PubMed]
- 49. Cantó C, Jiang LQ, Deshmukh AS, Mataki C, Coste A, Lagouge M, Zierath JR, Auwerx J. Interdependence of AMPK and SIRT1 for metabolic adaptation to fasting and exercise in skeletal muscle. Cell Metab. 2010; 11:213–19. https://doi.org/10.1016/j.cmet.2010.02.006 [PubMed]
- 50. Hart KJ, Shaw JM, Vajda E, Hegsted M, Miller SC. Swim-trained rats have greater bone mass, density, strength, and dynamics. J Appl Physiol (1985). 2001; 91:1663–68. https://doi.org/10.1152/jappl.2001.91.4.1663 [PubMed]
- 51. Wang ZH. Anti-glycative effects of asiatic acid in human keratinocyte cells. Biomedicine (Taipei). 2014; 4:19. https://doi.org/10.7603/s40681-014-0019-9 [PubMed]