




Introduction
The genetic basis of longevity in species ranging from yeast to mammals has been investigated primarily through experimental alteration of expression in individual genes. Fewer studies have investigated the genetic architecture of longevity in natural populations at the level of molecular function [1]. One gene whose role in life span has been extensively studied is the Indy gene, in both Drosophila and C. elegans systems [2-6]. Importantly, Indy's positive role in regulating metabolism and insulin sensitivity has recently been genetically demonstrated in mammals [7]. The INDY protein has been shown to possess dicarboxylate transporter activity [7, 8]. As such, it has been proposed that Indy mutations act to regulate the levels of tricarboxylic acid cycle (TCA) intermediates (e.g. citrate, succinate) via effects on transport, effectively inducing a genetic form of caloric restriction (CR) [4, 6], an intervention known to extend life span. While there is abundant evidence for the function of INDY, the mechanism through which experimentally-induced mutations in Indy increase life span have not been elucidated. Upon investigation, we found that Drosophila isolates from around the world vary with respect to a naturally occurring polymorphism caused by the insertion of a transposable element, Hoppel. This mobile element has inserted into the first intron of the Indy gene, and we show that it is often maintained in a heterozygous state. The presence of the Hoppel element in Indy is associated with an increase in fertility and increased longevity. We provide evidence for a selective sweep in the region where Hoppel has inserted, a molecular signature of recent positive selection acting on DNA sequences at the population level.
Results
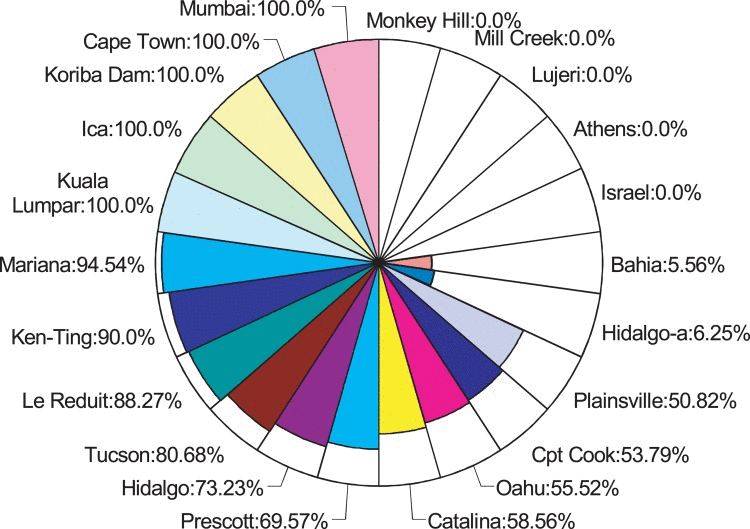
Hoppel polymorphism is found in Drosophila isolates from around the world
In the course of our molecular analyses of Indy gene structure in different Drosophila isolates from around the globe, we found a natural polymorphism present in numerous independent populations. This variant comprises the presence or absence of the transposable element, Hoppel, which has been implicated in regulating gene expression through effects on local chromatin structure [9, 10]. We sought to examine whether heterozygosity for the Hoppel insertion in Indy is maintained in other wild strains. Hoppel is an intronless, defective mobile element related to the P element, is about 1.2kb in average length, and is estimated to have 105 insertions in D. melanogaster euchromatin [11]. Using a PCR based screen, we found that the Hoppel insertion is polymorphic among 22 natural isolate lines (obtained from the Drosophila Species Stock Center, Table S1). Strikingly, many lines were not fixed for the insertion or wild-type alleles (12 of 22 examined lines), even after 5-50 years of inbreeding (60-600 generations) and still maintained the insertional allele at between 6-95 % frequency (Fig. 1). Located in the chromosomal map 75E region, the Indy gene is far from any common natural chromosomal inversions [12]. Thus, heterozygosity for this natural Indy variant is not likely a result of genetic hitchhiking.
Hoppel polymorphisms are associated with increased fertility
Heterozygosity for P{lacW} transgene insertion at the Indy locus has been shown to result in life span extension without a decrement in fecundity under normal laboratory culture conditions, but with a distinct cost under reduced caloric intake [13]. We isolated Hoppel insertion homozygotes (referred to as +/+) and non-insertion homozygotes (referred to as −/−) from three natural lines which demonstrated heterozygosity: Oahu, Hawaii collected in 1955; Captain Cook, Hawaii collected in 2007; and Hidalgo, Mexico collected in 2005. Congenic heterozygotes (−/+) were produced by intercrossing these homozygous genotypes isolated from each natural line (Fig. S1). To examine effects on reproductive fitness, we determined the fecundity of heterozygotes and homozygotes. In all cases, we found that a single copy of the Indy Hoppel insertion conferred highest fecundity (Fig. 2). The strong selection for maximized fecundity in wild populations favors the preservation of the insertion allele, conferring a large benefit (~10% increased egg production) on heterozygotes over the first 15 days of reproduction. Thus, we suggest that the Indy gene has been the subject of selection by the process of heterosis, or heterozygote advantage.
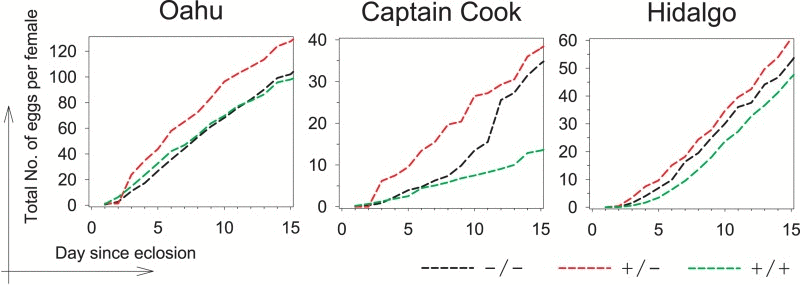
Figure 2. Hoppel insertion in Indy is associated with a reproductive advantage Accumulative number of eggs laid per female for each of the three Hoppel genotypes representing genomes with either no copies of Hoppel in Indy (−/−; in black), one copy of Hoppel in Indy (+/−; in red) or two copies of Hoppel in Indy (+/+; in green), isolated from three different populations collected from distant places or times from the wild (Oahu, Hawaii 1955, Captain Cook, Hawaii 2007 and Hidalgo, Mexico 2005).
Hoppel is associated with selective sweep
A molecular signature of recent positive selection acting on DNA sequences at the population level is decreased polymorphism linked to the advantageous allele, known as selective sweep [14]. Just such a selection on Hoppel is reflected by the DNA sequence polymorphism pattern in the region surrounding the Hoppel insertion site (Fig 3). We compared the sequences of Hoppel homozygotes +/+ and −/− isolates from the Oahu, Captain Cook and Hidalgo strains as well as the fully sequenced Drosophila Genomic Reference Panel (DGRP) lines (n=23) [15] which we also found were polymorphic for Hoppel insertions in Indy. Among the lines carrying Hoppel insertions (n=9), we found a dearth of polymorphisms in sequences neighboring the insertion site as compared to many more polymorphisms seen for sequences in the same Indy region in lines lacking the Hoppel insertion (n=14). Chromosomes from Oahu, Cpt. Cook and Hidalgo with a Hoppel insertion had only 2 polymorphisms over the combined 2 kb region upstream and downstream of the Hoppel insertion, while chromosomes of the same strains from Oahu, Cpt Cook and Hidalgo lacking the Hoppel insertion had a total of 31 polymorphisms over the same interval. For the Drosophila Genomic Reference Panel (DGRP) lines, or RAL lines, chromosomes containing Hoppel had on average 3.1-3.4 polymorphisms over the combined 2 kb region upstream and down stream of the Hoppel insertion site compared to chromosomes without Hoppel which had on average 10-11.5 polymorphisms over the same region. Since the three inbred lines, Oahu, Cpt Cook and Hidalgo are geographically and temporally separated from each other as well as from the DGRP lines, the common of reduced polymorphism around the Hoppel insertion can be best explained as a selective sweep encompassing the Indy locus due to positive selection associated with Hoppel rather than identity-by-descent. (Fig. 3).
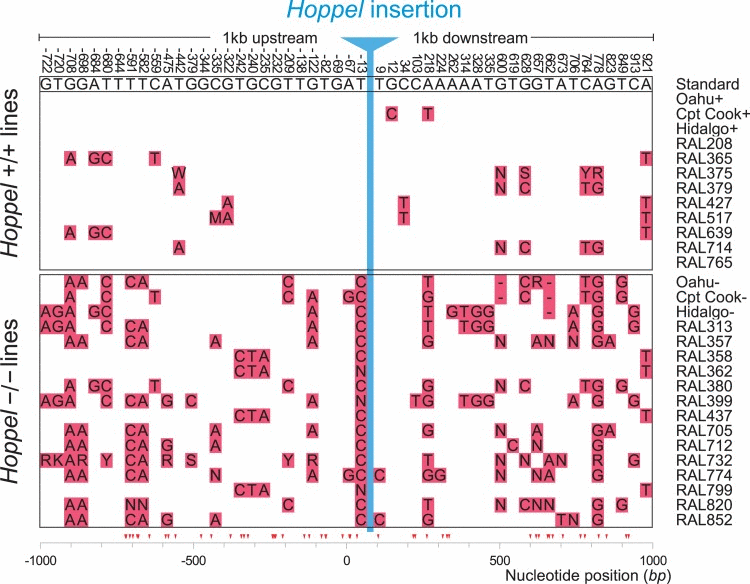
Figure 3. Hoppel insertion in Indy shows evidence for positive selection at the sequence level The DNA sequence polymorphisms 1000bp upstream and downstream from the Hoppel insertion site are shown in red blocks. The standard reference D. melanogaster genome sequence is at the top with nucleotide position for each polymorphic site referenced to the Hoppel insertion site as position 0. The upper half of the figure are strains containing the Hoppel insertion (Hoppel +/+) and the bottom half lines are strains that do not have the Hoppel insertion (Hoppel −/−) (Table. S1).
Hoppel is associated with increased life span
Based upon evolutionary hypotheses regarding the interaction between reproduction and longevity, we anticipated that animals heterozygous for the insertion allele might have shorter life spans due to their increased cost in resources associated with early life reproductive success. Remarkably, when we determined the influence of the presence of the Hoppel insertion on life spans, we found that under two very different dietary conditions; a high calorie diet commonly used in the laboratory, and a low calorie diet likely more realistic of food availability in the wild, heterozygote animals (−/+) for Hoppel significantly outlive congenic homozygotes lacking the Hoppel insertion in Indy (−/−) (Figs. 4A, 4B) despite having a higher early reproductive output (Fig. 2). While the reproductive advantage of being heterozygous for the Hoppel variant of Indy (−/+) is the likely force maintaining heterozygosity, these data demonstrate a naturally occurring genetic variant that, in keeping with similar previous experimental data, is capable of extending life span [4].
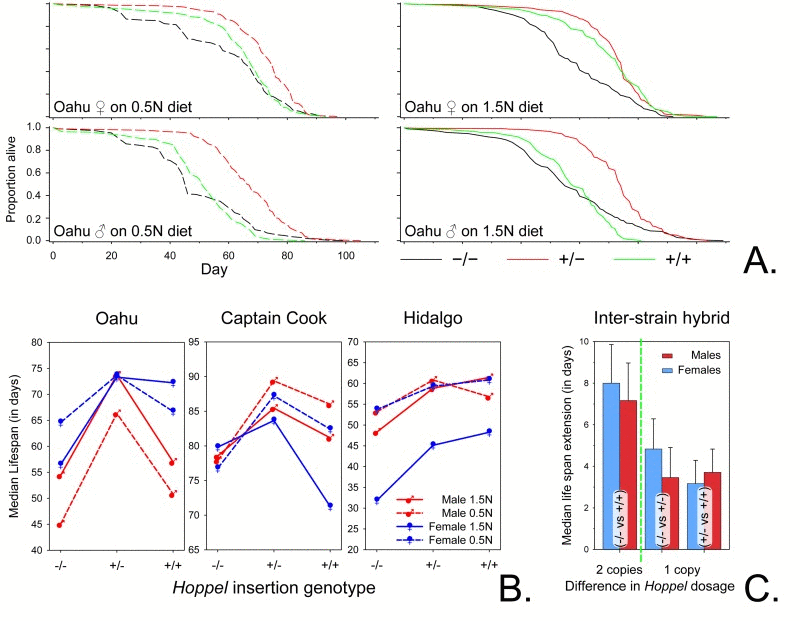
Figure 4. Hoppel insertion extends life span (A) Representative survivorship plots for female and male Hoppel genotypes isolated from Oahu lines with either no chromosome containing Hoppel in Indy (−/−; in black); one chromosome containing Hoppel in Indy (+/−; in red) or both chromosomes containing Hoppel in Indy (+/+; in green) grown on two different foods (1.5N and 0.5N diet) [29]. Survivorship plots for Captain Cook and Hidalgo lines are in Fig. S2 and tests for survivorship and maximum life span are in Table S2. (B) Reaction norms of median life span for Male (red) and Female (blue) Oahu, Captain Cook and Hidalgo lines for each of three different Indy genotypes (no chromosome containing Hoppel in Indy −/−; one chromosome containing Hoppel in Indy +/− or both chromosomes containing Hoppel in Indy +/+) grown on two different types of food (1.5N and 0.5N diet) [29]. (C) The effect of the dosage of each Hoppel containing Indy chromosome on life span extension for inter-strain hybrids between the Oahu, Captain Cook and Hidalgo strains represented as the mean of median life span extension in days for Males (red) and Females (blue) flies. Left of the green dotted line is the mean of the median life span extension for inter-strain hybrids in which both chromosomes have a Hoppel element in Indy (+/+) as compared to inter-strain hybrids with no Hoppel in Indy (−/−). Right of the green dotted line are shown the increase in mean of the median life span upon addition of each chromosome containing Hoppel in Indy; inter-strain hybrids with both chromosomes having a Hoppel in Indy (+/+) as compared to one chromosome having a Hoppel in Indy (+/−) and inter-strain hybrids with one chromosome having a Hoppel in Indy (+/−) as compared to inter-strain hybrids with no chromosome having a Hoppel in Indy (−/−) (Table S3). These data represent 96 separate inter-strain hybrid life spans including two different types of food (1.5N and 0.5N diet) [29]. Error bars are S.E.M.
To demonstrate that the life span extension is likely due to the insertion of Hoppel at the Indy locus (versus other genetic background effects within each strain), we examined the life span of inter-line hybrids where −/− and +/+ from each of the three natural isolate lines were crossed to their counterparts from another line. In these inter-line hybrids, with −/−, +/+ and −/+ genotypes at the Indy locus (and the rest of the genome being heterozygous at all loci), we continued to find a strong association between the presence of Hoppel in Indy and life span extension (Fig. 4C). Examination of the results of 96 separate life spans from these inter-line crosses indicates that on average, a one dose increase of Hoppel insertion extends median life span by ~5 days, while a two dose increase extends median life span by ~8 days.
Hoppel affects Indy mRNA expression
Examination of the molecular underpinnings of how Hoppel insertion at Indy affects longevity suggested that Hoppel, which is inserted within the 1st intron of Indy, and is 2216 base pairs upstream of the translational start, may be exerting its action by modulating the level of Indy expression. Therefore, we performed mRNA expression studies of Indy in various allelic combinations. The expression level of Indy was positively correlated with increasing Hoppel insertion dosage: the heterozygote being intermediate (Fig. 5). Heterozygosity for the Hoppel insertion was shown to confer the longest life span in 10 out of the 12 life spans (Fig. 4B) in males and females, independent of caloric intake. Interestingly, the effect of the Hoppel insertion in these wild strains on Indy transcription is reminiscent of the effect of the experimentally induced mutation, P{lacW}Indy206, which is a transposon insertion into the Hoppel in Indy residing in the normal laboratory strain that also modulates Indy transcription [4, 6].
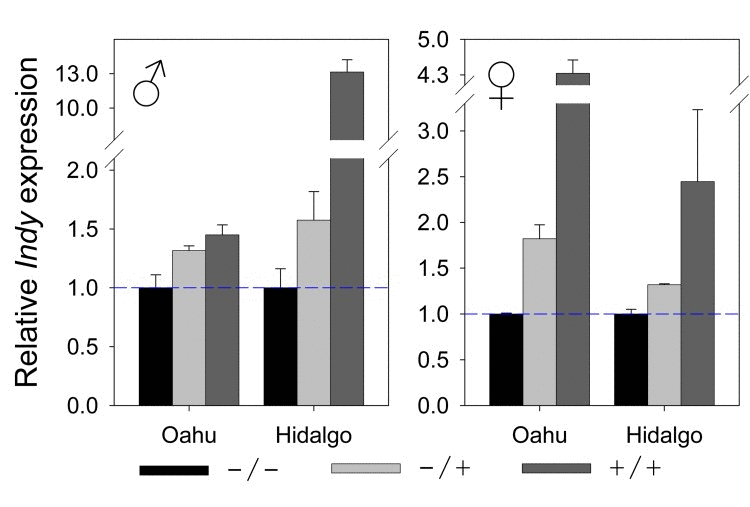
Figure 5. Hoppel insertion is positively correlated with Indy expression levels Indy mRNA expression for female and male Hoppel genotypes isolated from Oahu and Hidalgo lines measured by qPCR: with either no chromosome containing Hoppel in Indy (−/−; in black); one chromosome containing Hoppel in Indy (+/−; in light gray) or both chromosomes containing Hoppel in Indy (+/+; in dark gray). Relative expression level of Indy is compared to the expression level for the strains having no chromosome containing Hoppel in Indy (−/−). Error bars are S.E.M
Discussion
The role of the Indy gene in longevity, based upon molecular genetic interventions in invertebrates, has been controversial [2-6]. Here, we show that natural selection has maintained standing genetic variation at the Indy locus in the wild, with phenotypic consequences, demonstrating that this gene has an evolved role in metabolism, fecundity, and longevity determination. Standard evolutionary paradigms that emphasize metabolic tradeoffs invoke an inverse relationship between early life fecundity and longevity. For instance, when populations are selected for late-life fecundity, they are associated with increased life span [16-19]. However, a recent study suggests that the inverse correlation between lifespan and fecundity may not be a conserved feature of the genetic architecture of longevity [20]. We show, that at least in this case, the molecular lesion caused by Indy Hoppel insertion uncouples this association, allowing for early reproductive success (high fitness) as well as increased longevity. Nevertheless, homozygosity at Indy (in particular the insertion variant), results in decreased fitness, thereby favoring the maintenance of the insertion allele in the heterozygous state, providing one of the few examples of a natural variant transposable element insertion conferring adaptive value in a organism.
The molecular mechanism, by which the heterozygote advantage of Indy on fecundity and longevity is mediated, in both variants in the wild and through experimental manipulations in the laboratory [4, 6], involves modulation of Indy transcription. Additionally, positive effects of altered Indy transcription have been demonstrated in mammals, where genetic manipulation to reduce INDY function imparts significant age-related health benefits on insulin signaling and metabolism that are also found in Indy long-lived mutant flies [7, 21]. The fact that Indy expression varies in the wild and imparts such significant improvements in fitness should not be understated. Both experimental and natural Indy genetic variants appear to act via influences acting on the Hoppel transposon. Numerous studies on metazoans suggest that domestication of transposons serves as a platform for the generation of useful genetic variants and novel gene regulatory networks, upon which natural selection may act [22-27]. In particular, recent data demonstrates that Hoppel double-stranded RNAs are generated in vivo from a trigger site called Hoppel-killer(Hok) on chromosome 4, and that this locus serves as a global regulator of gene silencing directed at Hoppel elements elsewhere in the genome [28]. Thus, the Hoppel insertion in Indy (along with other Hoppel elements throughout the genome) may well be under the influence of complex signals acting through small RNA pathways and heterochromatic gene silencing. The entry of the Indy gene into the Hoppel element global regulatory networks seems to have provided ample positive variation for natural selection to act upon. Further experiments will be necessary to delineate the precise mechanisms through which Indy's inclusion in this network generates the phenotypes we describe. Moreover, if evolution tinkers with Indy expression to regulate important life-history elements in Nature, it suggests that Indy may serve as a natural and important hub for small-molecule intervention linking metabolism, fitness and longevity.
Materials and Methods
Drosophila Stocks
22 inbred Drosophila lines of different geographic origin were obtained from Drosophila Species Stock Center (https://stockcenter.ucsd.edu/). Drosophila Genetic Reference Panel (DGRP) stocks, DGRP lines, were original collected form Raleigh, NC, they were also known as RAL lines. The stock numbers of these lines are summarized in Table S1. All flies were maintained at 25°C in a temperature-controlled incubator at 50% humidity with a 12-hour light/dark cycle.
PCR based Hoppel insertion phenotyping
Multiplex PCR using genomic DNA as template was performed with a combination of three primers: a common forward primer located upstream of Hoppel insertion (Indy-hoppel±1S: 5'-CTACATTGTATACGGAGACATTCG G-3'); two reverse primers, one inside Hoppel (Indyhoppel+3A: 5'-CCATGTAAATTCGTTTCTTCGATC-3') and the other downstream of Hoppel (Indy-hoppel−1A: 5'-CATCTTTCGTCTTGCTATCAGCA-3'). Homozygote for Hoppel insertion (+/+) leads to two amplicons: 392 bp (between Indy-hoppel±1S and Indy-hoppel+3A) and 1423 bp (between Indy-hoppel±1S and Indy-hoppel−1A). Homozygote for without-Hoppel insertion (−/−) can be identified by a single amplicon of 319 bp in size (between Indy-hoppel±1S and Indy-hoppel−1A). Short extension time (15 seconds) was used to eliminate the formation of large amplicon of 1423 bp. Therefore, +/+ homozygote can be visualized as a single amplicon of 392 bp and −/− homozygotes as a 319 bp amplicon. Heterozygote was identified as two amplicons, both the 392 bp and 319 bp ones.
Isolation of +/+ and −/− homozygotes from Captain Cook, Oahu and Hidalgo lines
The crossing scheme for isolating both +/+ and −/− homozygotes from one inbred line is presented in Figure S1. Following this scheme, +/+ and −/− homozygous share the same ‘grandmother’ (generation P). The +/+ and −/− homozygotes, once isolated, were maintained via sib mating as stable stocks. We genotyped the homozygote stocks again at least 7 months after they had been generated and found their Hoppel insertion status unchanged. Therefore, the transposition rate of Hoppel is expected to be low and the homozygote lines can be considered as genetically stable.
Quantification of Hoppel insertion allele frequency
The quantifications of insertion allele frequency were repeated twice with 28 months in between. In the first attempt, before +/+ and −/− homozygotes were isolated from Cpt. Cook, Oahu and Hidalgo lines, 10 adult flies were individually genotyped using the PCR assay described above and the allele frequency was calculated from the number of observed +/+, −/+ and −/− individuals. In the second attempt, performed after +/+ and −/− homozygotes were isolated, pooled genomic DNA from 50 individuals from each inbred lines were genotyped and the PCR product visualized on 1% agrose gels. The amplicon band density was measured with a ChemiDoc XRS camera system (BioRad) and the density ratios between the large and the small amplicon were used to calculate allele frequency through interpolation from a standard curve. The standard curve is established by measuring amplicon density ratio from a series of standard pooled genomic DNA samples of designed allele frequencies. The standard pooled DNA was prepared from 5 individuals and the expected frequencies in the samples ranged from 0.1 (one −/+ and four −/− individuals) to 0.9 (one −/+ and four +/+ individuals). The Hoppel insertion allele frequency in DGRP lines were also assessed using the pooled genomic DNA method, but with 5 individuals in each pooled sample. For the DGRP lines we assayed and reported in Figure 1C, we noticed all of them appear to be homozygous for either (+) or (−).
Life span studies
Flies for demographic experiment were raised and collected from population density controlled broods in order to avoid any confounding effects due to over-crowding. The density controlled broods consisted of 10 replicate vials, each of which were seeded with 25 mating pairs of young adults as parents. The parents were allowed to mate and lay eggs for 2 days before being transferred onto fresh food. After 5 transfers, the parents were discarded. The offsprings from these parents were collected over a period of 24 hours and sorted by sex. 25 males and 25 females were kept in vials containing either 0.5N (5% yeast, 5% sucrose, 2% agar and 0.25% Tegospet in w/v concentration) or 1.5N (same as 0.5N except for 15% yeast and 15% sucrose) diet. For each genotype and dietary treatment, 10 replicate vials were set up so the total sample size is 250 for both sexes. Flies were transferred onto fresh food every two days with the dead flies, if any, removed and the number of death recorded. In the demographic assay for Cpt. Cook, Oahu and Hidalgo lines, heterozygote genotypes (Hoppel −/+) are generated by crossing −/− females to +/+ males. The inter-strain hybrid genotypes were generated from crosses between males and females from different lines, as summarized in Table S3.
Fecundity assay for +/+, −/+ and −/− genotypes for Cpt. Cook, Oahu and Hidalgo lines
Flies for fecundity assay were collected from the density controlled broods set up the same way as for the demographic experiments. Offsprings from the density controlled broods were collect within 8 hours, to ensure only newly eclosed males and females were collected. Groups of 5 males and 5 females were then transferred into vials containing 0.5N diet. The flies were then transferred onto fresh food every day and the number of eggs produced during the previous day were counted and recorded. The average egg production per female was computed from the total eggs of replicate vials and the total number of females. Each genotype was assayed with at least 7 independent biological replicates.
DNA sequencing of the region surrounding Hoppel element insertion in Indy
For +/+ and −/− homozygotes isolated from Cpt. Cook, Oahu and Hidalgo lines, DNA fragments both upstream and downstream of Hoppel element were amplified using the following primers: (Hoppel-BA: 5'-GAATTTCAGGTGTTTCGTGTGGG-3') and (Hoppel-BA-S: 5'-CAGATGTCTGCTATCAC TTTGTGCG-3') for upstream; (Indy-hoppel+1A: 5'-AAATCGTCTTCTAGCACAACACG-3') and (Hoppel-BS: 5'-GCATGAATAATGACCAGTTGAAC CC-3') for downstream. The PCR product was purified and sequenced using BigDye terminator chemistry on an Applied Biosystem platform. All fragments were sequenced on both strands. The sequence for DGRP lines was retrieved from DGRP Freeze 1 Data Release (http://www.hgsc.bcm.tmc.edu/projects/dgrp/freeze1_July_2010/). The coding sequence of Indy is highly conserved and there is only two nonsynonymous polymorphic sites (18822439A/G, resulting in an Ile/Val replacement change; and 18824503A/T, a Phe/Tyr replacement) with low frequency in all DGRP lines, 162 members in total. To confirm the observed effect of Hoppel insertion is not confounded by linked coding sequence polymorphisms in Cpt. Cook, Oahu and Hidalgo lines, we sequenced the coding regions for both +/+ and −/− homozygotes from the aforementioned lines. We found Cpt. Cook −/− homozygotes appeared to be fixed for 18822439G allele and all the other lines 18822439A allele. All of them are fixed for 18824503A. The 18822439A/G and 18824503A/T polymorphism locate at nucleotide position 3L: 18822439 and 18822503 respectively, based on Drosophila genome release version 5.
Indy expression level quantification by qPCR method
Total RNA was extracted from 10-day old flies using TRIzol reagent (Invitrogen) and was further purified using RNA Miniprep columns (Qiagen). Total cDNA was reverse transcribed using iScript cDNA Syntesis kit (BIORAD). RT-qPCR assays were performed on an ABI 7500 Fast Real-Time PCR System thermocycler using SYBR Green PCR Master Mix (ABI) following standard protocols. The relative change in Indy expression level in reference to the level of GAPDH was computed using ddCT method provided by ABI 7500 Fast System Software. The qPCR primers for GAPDH detection were: GAPDH-F: 5'-GACGAAATCAAGGCTAAGGTCG-3'. GAPDH-R: 5'-AATGGGTGTCGCTGAAGAAGTC-3'. The qPCR primers for Indy transcript detection were: (Indy-A-9Q: 5'-TGTTTCAGTCCCTGGC −3') and (Indy-S-9Q: 5'-TGGGCGGAGTACTAACC −3'), located in exon 8 and exon 9 respectively. We sequenced Indy exon 8 and exon 9 and confirmed the primer binding sites are conserved and all our lines carry the identical sequence. The qPCR assays were all performed with 4 replicates.
Supplementary Materials
Acknowledgments
Additional results and statistic tests are presented in supplementary tables and figures. The authors would like to thank Drs. David Rand and Marc Tatar for reading the manuscript and providing helpful comments, and Suzanne Hosier and Adam Kroll for technical assistance. This work was supported by NIA grants AG16667, AG24353 and AG25277 to SLH. SLH and RAR are Ellison Medical Research Foundation Senior Investigators and SLH is a recipient of a Glenn Award for Research in Biological Mechanisms of Aging.
Conflicts of Interest
The authors of this paper declare no conflict of interests.
References
- 1. De Luca M, Roshina NV, Geiger-Thornsberry GL, Lyman RF, Pasyukova EG, Mackay TF. Dopa decarboxylase (Ddc) affects variation in Drosophila longevity. Nat Genet. 2003; 34: 429 -433. [PubMed] .
- 2. Fei YJ, Liu JC, Inoue K, Zhuang L, Miyake K, Miyauchi S, Ganapathy V. Relevance of NAC-2, an Na+-coupled citrate transporter, to life span, body size and fat content in Caenorhabditis elegans. Biochem J. 2004; 379: 191 -198. [PubMed] .
- 3. Fei YJ, Inoue K, Ganapathy V. Structural and functional characteristics of two sodium-coupled dicarboxylate transporters (ceNaDC1 and ceNaDC2) from Caenorhabditis elegans and their relevance to life span. J Biol Chem. 2003; 278: 6136 -644. [PubMed] .
- 4. Rogina B, Reenan RA, Nilsen SP, Helfand SL. Extended life-span conferred by cotransporter gene mutations in Drosophila. Science. 2000; 290: 2137 -2140. [PubMed] .
- 5. Toivonen JM, Walker GA, Martinez-Diaz P, Bjedov I, Driege Y, Jacobs HT, Gems D, Partridge L. No influence of Indy on lifespan in Drosophila after correction for genetic and cytoplasmic background effects. PLoS Genet. 2007; 3: e95 [PubMed] .
- 6. Wang PY, Neretti N, Whitaker R, Hosier S, Chang C, Lu D, Rogina B, Helfand SL. Long-lived Indy and calorie restriction interact to extend life span. Proc Natl Acad Sci U S A. 2009; 106: 9262 -9267. [PubMed] .
- 7. Birkenfeld AL, Lee HY, Guebre-Egziabher F, Alves TC, Jurczak MJ, Jornayvaz FR, Zhang D, Hsiao JJ, Martin-Montalvo A, Fischer-Rosinsky A, Spranger J, Pfeiffer AF, Jordan J, Fromm MF, Konig J, Lieske S, Carmean CM, Frederick DW, Weismann D, Knauf F, Irusta PM, De Cabo R, Helfand SL, Samuel VT, Shulman GI. Deletion of the Mammalian INDY Homolog Mimics Aspects of Dietary Restriction and Protects against Adiposity and Insulin Resistance in Mice. Cell Metab. 2011; 14: 184 -195. [PubMed] .
- 8. Knauf F, Rogina B, Jiang Z, Aronson PS, Helfand SL. Functional characterization and immunolocalization of the transporter encoded by the life-extending gene Indy. Proc Natl Acad Sci U S A. 2002; 99: 14315 -14319. [PubMed] .
- 9. Haynes KA, Caudy AA, Collins L, Elgin SC. Element 1360 and RNAi components contribute to HP1-dependent silencing of a pericentric reporter. Curr Biol. 2006; 16: 2222 -2227. [PubMed] .
- 10. Savva YA, Jepson JEC, Chang Y-J, Whitaker R, Jones BC, St Laurent G, Tackett MR, Kapranov P, Jiang N, Du G, Helfand SL, Reenan RA. RNA editing regulates transposon-mediated heterochromatic gene silencing. Nature Comm. 2013; In Press .
- 11. Kaminker JS, Bergman CM, Kronmiller B, Carlson J, Svirskas R, Patel S, Frise E, Wheeler DA, Lewis SE, Rubin GM, Ashburner M, Celniker SE. The transposable elements of the Drosophila melanogaster euchromatin: a genomics perspective. Genome Biol. 2002; 3: RESEARCH0084 .
- 12. Krimbas CB and Powell JR. Drosophila Inversion Polymorphism 1992; Boca Raton, Ann Arbor, London, Tokyo CRC Press .
- 13. Marden JH, Rogina B, Montooth KL, Helfand SL. Conditional tradeoffs between aging and organismal performance of Indy long-lived mutant flies. Proc. Natl. Acad. Sci. USA. 2003; 100: 3369 -3373. [PubMed] .
- 14. Wootton JC, Feng X, Ferdig MT, Cooper RA, Mu J, Baruch DI, Magill AJ, Su XZ. Genetic diversity and chloroquine selective sweeps in Plasmodium falciparum. Nature. 2002; 418: 320 -323. [PubMed] .
- 15. Mackay TF, Richards S, Stone EA, Barbadilla A, Ayroles JF, Zhu D, Casillas S, Han Y, Magwire MM, Cridland JM, Richardson MF, Anholt RR, Barron M, Bess C, Blankenburg KP, Carbone MA, Castellano D, Chaboub L, Duncan L, Harris Z, Javaid M, Jayaseelan JC, Jhangiani SN, Jordan KW, Lara F, Lawrence F, Lee SL, Librado P, Linheiro RS, Lyman RF, Mackey AJ, Munidasa M, Muzny DM, Nazareth L, Newsham I, Perales L, Pu LL, Qu C, Ramia M, Reid JG, Rollmann SM, Rozas J, Saada N, Turlapati L, Worley KC, Wu YQ, Yamamoto A, Zhu Y, Bergman CM, Thornton KR, Mittelman D, Gibbs RA. The Drosophila melanogaster Genetic Reference Panel. Nature. 2012; 482: 173 -178. [PubMed] .
- 16. Luckinbill L, Arking R, Clare MJ, Cirocco WC, Buck S. Selection for delayed senescence in Drosophila melanogaster. Evolution. 1984; 38: 996 -1003. .
- 17. Luckinbill L and Clare M. Selection for life span in Drosophila melanogaster. Heredity. 1985; 55: 9 -18. [PubMed] .
- 18. Rose M. Laboratory evolution of postponed senescence in Drosophila melanogaster. Evolution. 1984; 38: 1004 -1009. .
- 19. Partridge L, Prowse N, Pignatelli P. Another set of responses and correlated responses to selection on age at reproduction in Drosophila melanogaster. Proc Biol Sci. 1999; 266: 255 -261. [PubMed] .
- 20. Jones OR, Scheuerlein A, Salguero-Gomez R, Camarda CG, Schaible R, Casper BB, Dahlgren JP, Ehrlen J, Garcia MB, Menges ES, Quintana-Ascencio PF, Caswell H, Baudisch A, Vaupel JW. Diversity of ageing across the tree of life. Nature. 2014; 505: 169 -173. [PubMed] .
- 21. Shulman GI and Helfand SL. Indy knockdown in mice mimics elements of dietary restriction. Aging (Albany NY). 2011; 3: 701 [PubMed] .
- 22. Slotkin RK and Martienssen R. Transposable elements and the epigenetic regulation of the genome. Nat Rev Genet. 2007; 8: 272 -285. [PubMed] .
- 23. Levin HL and Moran JV. Dynamic interactions between transposable elements and their hosts. Nat Rev Genet. 2011; 12: 615 -627. [PubMed] .
- 24. Perrat PN, DasGupta S, Wang J, Theurkauf W, Weng Z, Rosbash M, Waddell S. Transposition-driven genomic heterogeneity in the Drosophila brain. Science. 2013; 340: 91 -95. [PubMed] .
- 25. Singer T, McConnell MJ, Marchetto MC, Coufal NG, Gage FH. LINE-1 retrotransposons: mediators of somatic variation in neuronal genomes? Trends Neurosci. 2010; 33: 345 -354. [PubMed] .
- 26. Feschotte C. Transposable elements and the evolution of regulatory networks. Nat Rev Genet. 2008; 9: 397 -405. [PubMed] .
- 27. Wood JG and Helfand SL. Chromatin structure and transposable elements in aging. Front Genet. 2013; 4: 274 [PubMed] .
- 28. Savva YA, Jepson JE, Chang YJ, Whitaker R, Jones BC, St Laurent G, Tackett MR, Kapranov P, Jiang N, Du G, Helfand SL, Reenan RA. RNA editing regulates transposon-mediated heterochromatic gene silencing. Nat Commun. 2013; 4: 2745 [PubMed] .
- 29. Bross TG, Rogina B, Helfand SL. Behavioral, physical, and demographic changes in Drosophila populations through dietary restriction. Aging Cell. 2005; 4: 309 -317. [PubMed] .