



Introduction
Since the first isolation of human Embryonic Stem Cells (ESCs) [1] huge interest has developed in the scientific and clinical communities and in the public in general because of their therapeutic potential. In particular, attention has focused on their potential use in cell-based therapy for diseases that are refractory to conventional treatments, such as neurodegenerative diseases and immunodeficiency, because of their ability to be programmed into new mature differentiated cells of all lineages [2]. The list of pathologies that in theory can be treated using stem cells includes: Alzheimer's disease [3], Parkinson's disease [4], [5], Huntington's disease [6],stroke [7], diabetes [8], cancer [9], age-related disorders [10-12] haematological disorders [13], cardiovascular disease [14, 15] and bone and muscle regeneration [16]. This interest has further increased in the last 6 years since Takhashi and Yamanaka broke the dogma in developmental biology that mammalian somatic cell differentiation is an irreversible process [17, 18]. By transfection in human somatic cells of four transcription factors (Nanog, Sox2, c-Myc and Klf4) they were able to revert the differentiated cells to an embryonic-like state. Because these newly generated cells show the morphology, pluripotency and capacity to form teratomas like ESC, they named these cells, induced pluripotent stem cells (iPSCs). Thus, this revolutionary step in the field has provided the clinical and scientific communities a second tool for cell-based therapy. Although our knowledge of the molecular mechanisms that control the self-renewal and differentiation of stem cells has grown considerably during the past decade, we still need more basic research in order to understand the molecular mechanisms that regulate proliferation, survival and differentiation of stem cells particularly after transplantation and in the pathological environment.
In this review, we will describe the biology of ESCs and iPSCs, emphasising the common features that they share. We will also review the state of the art of stem cells in clinic.
ESCs
The following definition of a stem cell is now widely accepted: stem cells are characterized by their ability to renew themselves through mitotic cell division (self-renewal) and the ability to differentiate into diverse range of specialized cell types. In general, stem cells can be classified into two broad types of mammalian cell: 1) embryonic stem cells (ESCs), that are derived from the inner cell mass of the blastocyst, and 2) adult stem cells. Adult stem cells can be found in several tissues and can be further classified into different subtypes. The two groups can be distinguished on the basis of their ability for self-renewal and differentiation. In particular, the self-renewal potential of ESCs is unlimited, and they can generate all the cell types in the body. In contrast, adult stem cells have limited self-renewal and pluripotency, and are important physiologically in tissue repair and homeostasis in the adult.
The genes and the signalling pathways that control the self-renewal and the cell fate decisions is a molecular signature called “stemness” [19]. The cooperation of intrinsic elements (i.e. transcription factors) and extrinsic signals (i.e. leukemia inhibitor factor, bone morphogenetic protein and fibroblast growth factor) from their microenviroment [20-24] regulates the behaviour of stem cells (self-renewal + pluripotency) (Figure 1A). This shows that self-renewal and pluripotency of stem cells is a complex process that requires the coordination of multiple pathways involved in proliferation and the maintenance of an undifferentiated state [21, 22].
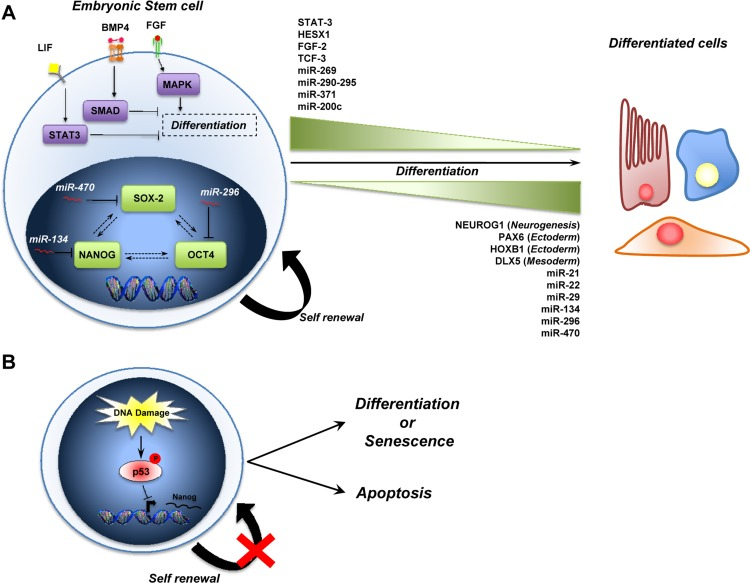
Figure 1. A schematic representation of ESCs biology (A) The stemness of ESCs is maintained by intrinsic (i.e. SOX2, NANOG and OCT4) and by extrinsic pathways (i.e. LIF, BMP4 and FGF). MicroRNAs also play a role in the maintenance of stem cells, and some are expressed during self-renewal (miR-269, miR-290-295 cluster, miR-371, miR-200c) while others are up-regulated during differentiation (miR-21, miR-22, miR-29, miR-134, miR-296, miR-470) (see text for details). (B) Role of p53 in the maintenance of genomic stability in ESCs. During DNA damage, p53 is activated (via Ser315 phosphorylation) and binds the Nanog promoter to repress its expression. The outcome of p53 activation is to induce the differentiation of ESCs into other cell types that they can go into a senescent state or induces apoptosis to preserve genome stability.
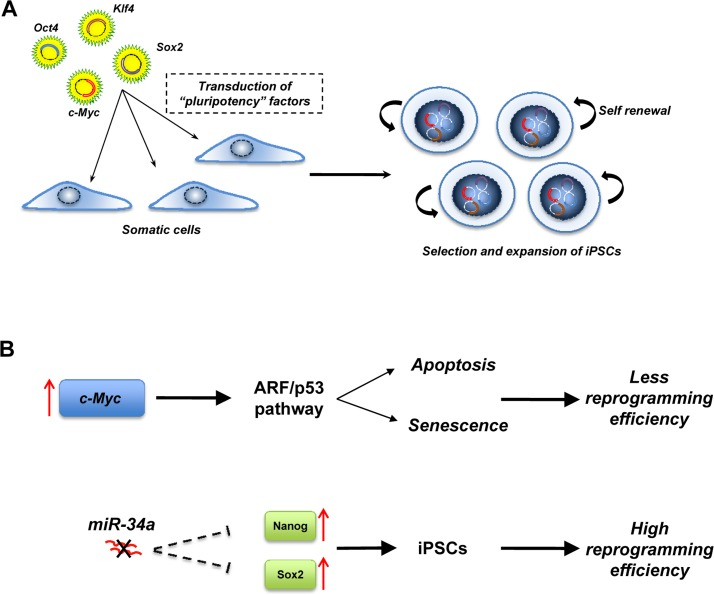
Figure 2. Induced pluripotent stem cells (iPSCs) (A) iPSCs are generated by the transduction in somatic cells of “pluripotency” factors. The resulting iPSCs have similar properties to ESCs. (B) p53 pathways regulate the efficiency of reprogramming. Overexpression of the oncogenic c-myc during reprogramming induces the ARF/p53 pathway that drives the cells to apoptosis or senescence and the miR-34 family negatively regulates the expression of Nanog and Sox2. In a miR-34 null-context, the expression of Nanog and Sox2 is increased, resulting in a higher reprogramming efficiency.
Three transcription factors, Nanog, Oct4 and Sox2 form the core of a regulatory circuit, which maintains stem cells capacity for self-renewal and pluripotency. Indeed, ESCs deficient in Nanog lose pluripotency and differentiate inappropriately [23, 25]. The POU domain transcription factor, Oct4, is also critical for the pluripotency of ESCs. Oct4 is down regulated during differentiation of ESCs in vitro andin vivo [26, 27]. Moreover, Oct4 can interact with Sox2 and bind to the Nanog promoter to regulate the expression of Nanog. The SRY-related HMG-box transcription factor Sox2 is also required for the maintenance of pluripotency in ESC in vivo and in vitro [28-30].
Thus, the Oct4, Sox2 and Nanog transcription factors control the expression of genes, including further transcription factors (such as STAT3, HESX1, FGF-2 and TCF) [31] and other signaling components necessary to maintain the stem cell state. Moreover, they also repress the expression of genes that, if expressed, would promote differentiation (such as NUEROG1, PAX6, HOXB1, DLX5) [32, 33]. This triad also forms an autoregulatory circuit, in which, by binding to their own promoters, as well as to the promoters of Oct4, Sox2 and Nanog they collaborate to sustain their expression.
Among the extrinsic factors, LIF (Leukemia Inhibitor factor) blocks the differentiation of mouse ESCs through the binding to its receptor and subsequent activation of Jak/STAT3 signaling [34, 35]. Activation of this pathway is essential for self-renewal of ESC and is necessary to maintain the undifferentiated state of ESCs[36]. Another extrinsic factor that is critical for the maintenance of the pluripotency of ESCs is bone morphogenetic protein 4 (BMP4) [37, 38]. BMP4 binds the BMP receptor and activates SMAD proteins, which in turn promote the expression of inhibitor of differentiation (Id) proteins. The Id proteins block lineage commitment and permit self-renewal of ESCs [39, 40], for example by blocking the ESC differentiation induced by Fibroblast Growth Factor (FGF) via MAPK signaling [41].
Recently, several observations indicate that the p53 family is involved in the regulation of stem cell biology [42]. The first indication of a direct p53 involvement in this process comes from the observation that p53 regulates Nanog expression [43, 44]. In particular, it has been shown that after induction of DNA damage in mouse embryonic stem cells (mESCs), p53 is phosphorylated at Ser315 and binds to the promoter of Nanog, suppressing its expression. Consequently, the result of p53 activation in this system is to bring mESCs into a more differentiated state where the cells can efficiently undergo p53-dependent cell cycle arrest or apoptosis, promoting the preservation of genomic stability. This role of p53 is supported by the fact that the loss of p53 in human ESCs leads to an increase of genomic instability [45]. Moreover, p53 participates in the control of neural stem cells (NSC). Loss of p53 leads to an elevated proliferation rate as well as an increase in the self-renewal of in vitro propagated p53−/− neural stem cells [46], although the precise contribution of p53 to NSC differentiation is somewhat controversial [47].
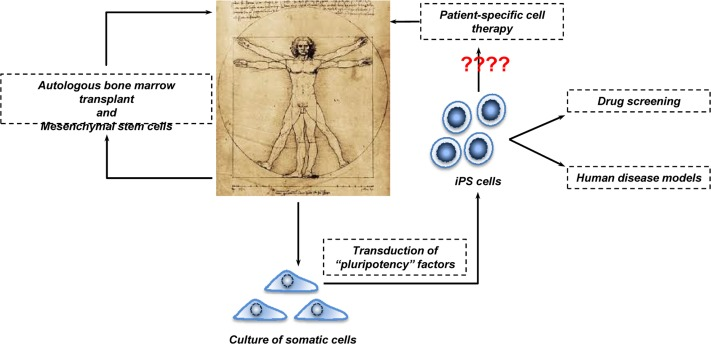
Figure 3. Human cell-based therapy Bone marrow transplantation is widely employed in the clinic for several diseases including cancer and haematological disorders. More recently, MSCs are stepping in. They could be used to treat inflammatory conditions such as Multiple Sclerosis and Pulmonary Fibrosis. The employment of iPSCs in cell-based therapy is a relative new tool. However, because their potential tumorigenicity in the clinical setting remains to be clarified, perhaps iPSCs should stay on the bench meanwhile and be used in drug screening and human disease models.
Another p53 family member, p73 [48-52] is also required for the maintenance of NSCs. Indeed, several experimental findings demonstrate that p73, in particular the TAp73 isoform, is a positive regulator of embryonic and adult NSCs. p73 null mice show a reduction in neurogenesis in the subgranular zone of the dentate gyrus and in the subventricular zone, and neurospheres derived from p73 null mice grow more slowly and are smaller. The potential downstream candidates responsible for this phenotype are genes involved in the regulation of proliferation and/or self-renewal pathways [24, 53], and loss of p73 leads to a transcriptional deregulation of Sox-2, Sox-3, Nanog, Notch-I, Notch-2, Hes-5, Jag2, Hey-2 and Deltex.
With the discoveries of microRNAs (miRs), a new layer of complexity has been added to stem cell biology [54, 55]. miRs (for instance miR-269, the miR-290-295 cluster, miR-371 and miR-200c), have been found to be preferentially expressed in undifferentiated stem cells and their expression levels decrease as the stem cells differentiate [56]. In contrast, the expression of other miRs (miR-21, miR-22 and miR-29) increases during the differentiation of ESCs indicating a possible role for them in stem cell differentiation [57, 58]. Moreover, loss of the components of the miR processing machinery, such as Dicer and Drosha-DGCR8-Ddx5 affect both self-renewal and differentiation of ESCs [59, 60]. The Nanog-Oct4-Sox2 triad is also a target of miRs during the differentiation of ESCs. In particular, miR-134, miR-296 and miR-470 are up regulated during the differentiation of mouse embryonic stem cells induced by retinoic acid, and target these three transcription factors. This leads to transcriptional and morphological changes characteristic of differentiating mouse ESCs. More recently, miR-124 and miR-34a have been shown to contribute to neuronal differentiation of ESCs [61-63].
iPSCs
Nuclear transplantation, cellular fusion and cell explantation are the strategies that scientists have employed in order to induce the conversion of differentiated cells into an embryonic state [17]. However, starting from the “simple” assumption that the factors that play a key role in the maintenance of pluripotency of ESC could work as reprograming factors, Yamanaka and colleagues were able to reprogram somatic cells into pluripotent ESCs [17]. They first identified genes that were highly and specifically expressed in the ESC, but not in somatic cells, and divided these into three groups. The first group comprised genes responsible for the maintenance of a pluripotent state (Nanog, Sox2 and Oct3/4). The second group contained c-Myc, Stat-3 and TCl1, tumor-associated genes. Finally, the third group contained genes that have a specific role in ESCs such as Kfl-4, ECAT1 and Esg1. Different combinations of genes in the three groups were transfected into mouse embryonic fibroblasts with the result that a combination of only four factors (Oct4, Sox2, Klf4 and c-Myc), were sufficient to reprogram somatic cells into iPSCs. While Oct-4 and Sox-2 are required for reprogramming, a combination of other factors, such as Oct4, Sox2, Nanog and Lin28, is also able to reprogram somatic cells into iPSCs [64]. However, Nanog is dispensable, although two oncogenic factors (c-Myc and Klf4) are essential for reprogramming. Phenotypically, the iPSC are similar to ESC in many aspects, including morphology, surface markers, gene expression, in-vitro differentiation and teratoma formation when they are injected in immunocompromised mice. However, the precise molecular mechanism of reprogramming remains unclear. While the role of Oct3/4 and Sox2 could be predicted by the fact that both have a role in the control of pluripotency in ESCs, the exact role of c-Myc and Klf4 remains to be clarified. We can speculate that c-Myc and Klf4 act as modifiers of chromatin structure allowing Oct3/4 and Sox2 to bind their target genes that are normally silenced by epigenetic mechanisms in differentiated cells.
While the ability to develop iPSCs from differentiated somatic cells is exciting, the system has two major drawbacks. Firstly, the reprogramming efficiency is very low, suggesting that inside the cell there may be mechanisms that prevent the reprogramming process: secondly, there is the oncogenic potential of iPSCs, as reflected in their ability to form teratomas in mice. Several findings suggest that p53 is responsible for the low efficiency in the reprogramming of somatic cells [65]. Indeed, overexpression of the oncogene c-Myc induces the ARF/p53 pathway driving the cells towards apoptosis or senescence [66]. This is also supported by the fact that the efficiency of reprogramming is higher in a p53 null context [67-71]. Recently, some observations indicate that the miR-34 family [72] may also regulate reprogramming of somatic cells, and deficiency of miR-34 significantly increases reprogramming efficiency and kinetics [73]. In contrast to p53 deficiency, which enhances reprogramming at the expense of iPSC pluripotency, genetic ablation of miR-34 promoted iPSC generation without compromising self-renewal or pluripotency. Suppression of reprogramming by miR-34a was due, at least in part, to repression of pluripotency genes, including Nanog, Sox2 and N-Myc. However, miR-34a ablation only partially phenocopies that of p53 [68] (and combined deletion of miR-34a and p21 similarly fails to reproduce the p53−/− phenotype), suggesting that p53 provides a barrier to iPSC generation through mechanisms and targets not fully characterized. Because inactivation of p53 and/or downstream components of its pathway seem to be critical for reprogramming efficiency, this raises concern about the tumorigenicity of iPSCs, since chromosomal aberrations have been found in iPSCs[74]. Moreover, injection of iPSCs into blastocysts led to an incidence (about 20%) of tumors in the resultant chimeric mice, attributable to reactivation of the c-Myc transgene [77].
Stem Cells in Therapy: myth or reality
As mentioned above, cell-based therapy has generated a lot of excitement in the scientific and clinical communities. In order to understand how far or close we are from the reality, we scanned the public clinical trials database of the National Institute of Health. Typing “stem cells” into http://clinicaltrials.gov database revealed that there are more than 4000 clinical trials in progress at different stages (Recruiting, Phase I/II and Terminated) for a wide range of diseases (reviewed in [76]). Overall, the following picture emerges. First, extensive experience, extending over more than 30 years, on the transplantation of bone marrow hematopoietic stem cells into patients affected by blood pathologies and cancer provides reasonable confidence on the safety and efficacy of cell-based therapy.
A second message is the use of mesenchymal stem cells (MSCs) as alternatives to the stem cell sources described above. MSCs are adult stem cells that are traditionally found in the bone marrow. However, they can be also isolated from other tissues such as cord and peripheral blood. MSCs can be differentiated to form adipocytes, cartilage, bone, muscle and skin, and therefore are under consideration for bone and cartilage repair and autoimmune disease [77]. One advantage of using MSCs in cell-based therapy is the fact that MSCs are of low immunogenicity (due to lack of surface MHC class II expression) and thus well tolerated during transplantation. However, since it has been shown that MSCs support tumor growth in allogenic animals, more basic research is essential in order to better understand the potential and especially the limits of MSCs [78-82].
The alternative to the use of MSCs, and one probably of broader clinical applicability, is to use iPSCs [83]. Like MSCs, these are autologous, and the problem of immunological rejection and use of potent immunosuppressants implicit in ESC therapy does not arise. However, although iPSCs largely resemble ESCs with respect to gene expression profile, epigenetic signature and differentiation capability, their use as therapy is clouded by their potential tumorigenicity. In particular, there is recent evidence reprogramming into iPSCs induces genetic and epigenetic abnormalities [84-89], including copy number variation, point mutations, DNA methylation and alteration of chromosome numbers. The clinical use of iPSCs must therefore proceed with caution. Nevertheless, several studies have already demonstrated that it is possible to generate iPSCs from somatic cells of patients affected by diabetes[90], neurodegenerative diseases [91-93] and psychiatric disorders such as schizophrenia [94]. The generated iPSCs recapitulate in vitro much of the cellular pathology of the disease, and, quite apart from any therapeutic potential, could be useful for more detailed analysis of disease pathology [95, 96], drug screening and associated toxicology.
Acknowledgments
This work was supported by the Medical Research Council.
Conflicts of Interest
The authors of this manuscript have no conflict of interests to declare.
References
- 1. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. Science. 1998; 282: 1145 -1147. [PubMed] .
- 2. Daley GQ. Stem cells: roadmap to the clinic. J Clin Invest. 2010; 120: 8 -10. [PubMed] .
- 3. Crews L, Patrick C, Adame A, Rockenstein E, Masliah E. Modulation of aberrant CDK5 signaling rescues impaired neurogenesis in models of Alzheimer's disease. Cell Death Dis. 2011; 2: e120 [PubMed] .
- 4. Yang D, Zhang ZJ, Oldenburg M, Ayala M, Zhang SC. Human embryonic stem cell-derived dopaminergic neurons reverse functional deficit in parkinsonian rats. Stem Cells. 2008; 26: 55 -63. [PubMed] .
- 5. Chinta SJ and Andersen JK. Prospects and challenges for the use of stem cell technologies to develop novel therapies for Parkinson disease. Cell Cycle. 2011; 10: 4179 -4180. [PubMed] .
- 6. Bano D, Zanetti F, Mende Y, Nicotera P. Neurodegenerative processes in Huntington's disease. Cell Death Dis. 2011; 2: e228 [PubMed] .
- 7. Copeland N, Harris D, Gaballa MA. Human umbilical cord blood stem cells, myocardial infarction and stroke. Clin Med. 2009; 9: 342 -345. [PubMed] .
- 8. Matsumoto S. Islet cell transplantation for Type 1 diabetes. J Diabetes. 2010; 2: 16 -22. [PubMed] .
- 9. Frank NY, Schatton T, Frank MH. The therapeutic promise of the cancer stem cell concept. J Clin Invest. 2010; 120: 41 -50. [PubMed] .
- 10. Zhao J and Pei G. Why cell reprogramming is functionally linked to aging? Aging (Albany NY). 2011; 3: 700 [PubMed] .
- 11. Ratajczak MZ, Kucia M, Liu R, Shin DM, Bryndza E, Masternak MM, Tarnowski M, Ratajczak J, Bartke A. RasGrf1: genomic imprinting, VSELs, and aging. Aging (Albany NY). 2011; 3: 692 -697. [PubMed] .
- 12. Ratajczak MZ, Shin DM, Ratajczak J, Kucia M, Bartke A. A novel insight into aging: are there pluripotent very small embryonic-like stem cells (VSELs) in adult tissues overtime depleted in an Igf-1-dependent manner? Aging (Albany NY). 2010; 2: 875 -883. [PubMed] .
- 13. Persons DA. The challenge of obtaining therapeutic levels of genetically modified hematopoietic stem cells in beta-thalassemia patients. Ann N Y Acad Sci. 2010; 1202: 69 -74. [PubMed] .
- 14. Sekiguchi H, Ii M, Losordo DW. The relative potency and safety of endothelial progenitor cells and unselected mononuclear cells for recovery from myocardial infarction and ischemia. J Cell Physiol. 2009; 219235 -242. .
- 15. Song YH, Pinkernell K, Alt E. Stem cell induced cardiac regeneration: fusion/mitochondrial exchange and/or transdifferentiation? Cell Cycle. 2011; 10: 2281 -2286. [PubMed] .
- 16. Quattrocelli M, Cassano M, Crippa S, Perini I, Sampaolesi M. Cell therapy strategies and improvements for muscular dystrophy. Cell Death Differ. 2010; 17: 1222 -1229. [PubMed] .
- 17. Takahashi K and Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006; 126: 663 -676. [PubMed] .
- 18. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007; 131: 861 -872. [PubMed] .
- 19. Ivanova NB, Dimos JT, Schaniel C, Hackney JA, Moore KA, Lemischka IR. A stem cell molecular signature. Science. 2002; 298: 601 -604. [PubMed] .
- 20. Casati A, Frascoli M, Traggiai E, Proietti M, Schenk U, Grassi F. Cell-autonomous regulation of hematopoietic stem cell cycling activity by ATP. Cell Death Differ. 2011; 18: 396 -404. [PubMed] .
- 21. Abdelalim EM and Tooyama I. NPR-A regulates self-renewal and pluripotency of embryonic stem cells. Cell Death Dis. 2011; 2: e127 [PubMed] .
- 22. Richly H, Aloia L, Di Croce L. Roles of the Polycomb group proteins in stem cells and cancer. Cell Death Dis. 2011; 2: e204 [PubMed] .
- 23. Chambers I, Colby D, Robertson M, Nichols J, Lee S, Tweedie S, Smith A. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell. 2003; 113: 643 -655. [PubMed] .
- 24. Molofsky AV, Pardal R, Morrison SJ. Diverse mechanisms regulate stem cell self-renewal. Curr Opin Cell Biol. 2004; 16: 700 -707. [PubMed] .
- 25. Mitsui K, Tokuzawa Y, Itoh H, Segawa K, Murakami M, Takahashi K, Maruyama M, Maeda M, Yamanaka S. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell. 2003; 113: 631 -642. [PubMed] .
- 26. Pesce M, Gross MK, Scholer HR. In line with our ancestors: Oct-4 and the mammalian germ. Bioessays. 1998; 20: 722 -732. [PubMed] .
- 27. Niwa H, Miyazaki J, Smith AG. Quantitative expression of Oct-3/4 defines differentiation, dedifferentiation or self-renewal of ES cells. Nat Genet. 2000; 24: 372 -376. [PubMed] .
- 28. Avilion AA, Nicolis SK, Pevny LH, Perez L, Vivian N, Lovell-Badge R. Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev. 2003; 17: 126 -140. [PubMed] .
- 29. Graham V, Khudyakov J, Ellis P, Pevny L. SOX2 functions to maintain neural progenitor identity. Neuron. 2003; 39: 749 -765. [PubMed] .
- 30. Basu-Roy U, Ambrosetti D, Favaro R, Nicolis SK, Mansukhani A, Basilico C. The transcription factor Sox2 is required for osteoblast self-renewal. Cell Death Differ. 2010; 17: 1345 -1353. [PubMed] .
- 31. Matthews JR, Sansom OJ, Clarke AR. Absolute requirement for STAT3 function in small-intestine crypt stem cell survival. Cell Death Differ. 2011; 18: 1934 -1943. [PubMed] .
- 32. Boyer LA, Lee TI, Cole MF, Johnstone SE, Levine SS, Zucker JP, Guenther MG, Kumar RM, Murray HL, Jenner RG, Gifford DK, Melton DA, Jaenisch R, et al. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell. 2005; 122: 947 -956. [PubMed] .
- 33. Loh YH, Wu Q, Chew JL, Vega VB, Zhang W, Chen X, Bourque G, George J, Leong B, Liu J, Wong KY, Sung KW, Lee CW, et al. The Oct4 and Nanog transcription network regulates pluripotency in mouse embryonic stem cells. Nat Genet. 2006; 38: 431 -440. [PubMed] .
- 34. Niwa H, Burdon T, Chambers I, Smith A. Self-renewal of pluripotent embryonic stem cells is mediated via activation of STAT3. Genes Dev. 1998; 12: 2048 -2060. [PubMed] .
- 35. Daheron L, Opitz SL, Zaehres H, Lensch MW, Andrews PW, Itskovitz-Eldor J, Daley GQ. LIF/STAT3 signaling fails to maintain self-renewal of human embryonic stem cells. Stem Cells. 2004; 22: 770 -778. [PubMed] .
- 36. Matsuda T, Nakamura T, Nakao K, Arai T, Katsuki M, Heike T, Yokota T. STAT3 activation is sufficient to maintain an undifferentiated state of mouse embryonic stem cells. Embo J. 1999; 18: 4261 -4269. [PubMed] .
- 37. Ying QL, Nichols J, Chambers I, Smith A. BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell. 2003; 115: 281 -292. [PubMed] .
- 38. Ono Y, Calhabeu F, Morgan JE, Katagiri T, Amthor H, Zammit PS. BMP signalling permits population expansion by preventing premature myogenic differentiation in muscle satellite cells. Cell Death Differ. 2011; 18: 222 -234. [PubMed] .
- 39. Norton JD. ID helix-loop-helix proteins in cell growth, differentiation and tumorigenesis. J Cell Sci. 2000; 113: 3897 -3905. [PubMed] .
- 40. Ruzinova MB, Schoer RA, Gerald W, Egan JE, Pandolfi PP, Rafii S, Manova K, Mittal V, Benezra R. Effect of angiogenesis inhibition by Id loss and the contribution of bone-marrow-derived endothelial cells in spontaneous murine tumors. Cancer Cell. 2003; 4: 277 -289. [PubMed] .
- 41. Ying QL, Stavridis M, Griffiths D, Li M, Smith A. Conversion of embryonic stem cells into neuroectodermal precursors in adherent monoculture. Nat Biotechnol. 2003; 21: 183 -186. [PubMed] .
- 42. Abbas HA, Pant V, Lozano G. The ups and downs of p53 regulation in hematopoietic stem cells. Cell Cycle. 2011; 10: 3257 -3262. [PubMed] .
- 43. Lin T, Chao C, Saito S, Mazur SJ, Murphy ME, Appella E, Xu Y. p53 induces differentiation of mouse embryonic stem cells by suppressing Nanog expression. Nat Cell Biol. 2005; 7: 165 -171. [PubMed] .
- 44. Mora-Castilla S, Tejedo JR, Hmadcha A, Cahuana GM, Martin F, Soria B, Bedoya FJ. Nitric oxide repression of Nanog promotes mouse embryonic stem cell differentiation. Cell Death Differ. 2010; 17: 1025 -1033. [PubMed] .
- 45. Xu Y. A new role for p53 in maintaining genetic stability in embryonic stem cells. Cell Cycle. 2005; 4: 3 363 -364. [PubMed] .
- 46. Meletis K, Wirta V, Hede SM, Nister M, Lundeberg J, Frisen J. p53 suppresses the self-renewal of adult neural stem cells. Development. 2006; 133: 363 -369. [PubMed] .
- 47. Armesilla-Diaz A, Bragado P, Del Valle I, Cuevas E, Lazaro I, Martin C, Cigudosa JC, Silva A. p53 regulates the self-renewal and differentiation of neural precursors. Neuroscience. 2009; 158: 1378 -1389. [PubMed] .
- 48. Flores ER. p73 is critical for the persistence of memory. Cell Death Differ. 2011; 18: 381 -382. [PubMed] .
- 49. Agostini M, Tucci P, Chen H, Knight RA, Bano D, Nicotera P, McKeon F, Melino G. p73 regulates maintenance of neural stem cell. Biochem Biophys Res Commun. 2010; 403: 13 -17. [PubMed] .
- 50. Gonzalez-Cano L, Herreros-Villanueva M, Fernandez-Alonso R, Ayuso-Sacido A, Meyer G, Garcia-Verdugo JM, Silva A, Marques MM, Marin MC. p73 deficiency results in impaired self renewal and premature neuronal differentiation of mouse neural progenitors independently of p53. Cell Death Dis. 2010; 1: e109 [PubMed] .
- 51. Talos F, Abraham A, Vaseva AV, Holembowski L, Tsirka SE, Scheel A, Bode D, Dobbelstein M, Bruck W, Moll UM. p73 is an essential regulator of neural stem cell maintenance in embryonal and adult CNS neurogenesis. Cell Death Differ. 2010; 17: 1816 -1829. [PubMed] .
- 52. Fujitani M, Cancino GI, Dugani CB, Weaver IC, Gauthier-Fisher A, Paquin A, Mak TW, Wojtowicz MJ, Miller FD, Kaplan DR. TAp73 acts via the bHLH Hey2 to promote long-term maintenance of neural precursors. Curr Biol. 2010; 20: 2058 -2065. [PubMed] .
- 53. Zimmer B, Kuegler PB, Baudis B, Genewsky A, Tanavde V, Koh W, Tan B, Waldmann T, Kadereit S, Leist M. Coordinated waves of gene expression during neuronal differentiation of embryonic stem cells as basis for novel approaches to developmental neurotoxicity testing. Cell Death Differ. 2011; 18: 383 -395. [PubMed] .
- 54. Gangaraju VK and Lin H. MicroRNAs: key regulators of stem cells. Nat Rev Mol Cell Biol. 2009; 10: 116 -125. [PubMed] .
- 55. Saunders LR, Sharma AD, Tawney J, Nakagawa M, Okita K, Yamanaka S, Willenbring H, Verdin E. miRNAs regulate SIRT1 expression during mouse embryonic stem cell differentiation and in adult mouse tissues. Aging (Albany NY). 2010; 2: 415 -431. [PubMed] .
- 56. He L and Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004; 5: 522 -531. [PubMed] .
- 57. Houbaviy HB, Murray MF, Sharp PA. Embryonic stem cell-specific MicroRNAs. Dev Cell. 2003; 5: 351 -358. [PubMed] .
- 58. Singh SK, Kagalwala MN, Parker-Thornburg J, Adams H, Majumder S. REST maintains self-renewal and pluripotency of embryonic stem cells. Nature. 2008; 453: 223 -227. [PubMed] .
- 59. Bernstein E, Kim SY, Carmell MA, Murchison EP, Alcorn H, Li MZ, Mills AA, Elledge SJ, Anderson KV, Hannon GJ. Dicer is essential for mouse development. Nat Genet. 2003; 35: 215 -217. [PubMed] .
- 60. Kanellopoulou C, Muljo SA, Kung AL, Ganesan S, Drapkin R, Jenuwein T, Livingston DM, Rajewsky K. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev. 2005; 19: 489 -501. [PubMed] .
- 61. Makeyev EV, Zhang J, Carrasco MA, Maniatis T. The MicroRNA miR-124 promotes neuronal differentiation by triggering brain-specific alternative pre-mRNA splicing. Mol Cell. 2007; 27: 435 -448. [PubMed] .
- 62. Agostini M, Tucci P, Steinert JR, Shalom-Feuerstein R, Rouleau M, Aberdam D, Forsythe ID, Young KW, Ventura A, Concepcion CP, Han YC, Candi E, Knight RA, et al. microRNA-34a regulates neurite outgrowth, spinal morphology, and function. Proc Natl Acad Sci U S A. 2011; 108: 21099 -21104. [PubMed] .
- 63. Agostini M, Tucci P, Killick R, Candi E, Sayan BS, Rivetti di Val Cervo P, Nicotera P, McKeon F, Knight RA, Mak TW, Melino G. Neuronal differentiation by TAp73 is mediated by microRNA-34a regulation of synaptic protein targets. Proc Natl Acad Sci U S A. 2011; 108: 21093 -21098. [PubMed] .
- 64. Yu J, Vodyanik MA, Smuga-Otto K, Antosiewicz-Bourget J, Frane JL, Tian S, Nie J, Jonsdottir GA, Ruotti V, Stewart R, Slukvin II, Thomson JA. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007; 318: 1917 -1920. [PubMed] .
- 65. Menendez S, Camus S, Izpisua Belmonte JC. p53: guardian of reprogramming. Cell Cycle. 2010; 9: 3887 -3891. [PubMed] .
- 66. Pelengaris S, Khan M, Evan G. c-MYC: more than just a matter of life and death. Nat Rev Cancer. 2002; 2: 764 -776. [PubMed] .
- 67. Zhao Y, Yin X, Qin H, Zhu F, Liu H, Yang W, Zhang Q, Xiang C, Hou P, Song Z, Liu Y, Yong J, Zhang P, et al. Two supporting factors greatly improve the efficiency of human iPSC generation. Cell Stem Cell. 2008; 3: 475 -479. [PubMed] .
- 68. Hong H, Takahashi K, Ichisaka T, Aoi T, Kanagawa O, Nakagawa M, Okita K, Yamanaka S. Suppression of induced pluripotent stem cell generation by the p53-p21 pathway. Nature. 2009; 4601132 -1135. .
- 69. Kawamura T, Suzuki J, Wang YV, Menendez S, Morera LB, Raya A, Wahl GM, Izpisua Belmonte JC. Linking the p53 tumour suppressor pathway to somatic cell reprogramming. Nature. 2009; 460: 1140 -1144. [PubMed] .
- 70. Marion RM, Strati K, Li H, Murga M, Blanco R, Ortega S, Fernandez-Capetillo O, Serrano M, Blasco MA. A p53-mediated DNA damage response limits reprogramming to ensure iPS cell genomic integrity. Nature. 2009; 460: 7259 1149 -1153. [PubMed] .
- 71. Utikal J, Polo JM, Stadtfeld M, Maherali N, Kulalert W, Walsh RM, Khalil A, Rheinwald JG, Hochedlinger K. Immortalization eliminates a roadblock during cellular reprogramming into iPS cells. Nature. 2009; 460: 1145 -1148. [PubMed] .
- 72. Hermeking H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010; 17: 193 -199. [PubMed] .
- 73. Choi YJ, Lin CP, Ho JJ, He X, Okada N, Bu P, Zhong Y, Kim SY, Bennett MJ, Chen C, Ozturk A, Hicks GG, Hannon GJ, et al. miR-34 miRNAs provide a barrier for somatic cell reprogramming. Nat Cell Biol. 2011; 13: 1353 -1360. [PubMed] .
- 74. Chin MH, Mason MJ, Xie W, Volinia S, Singer M, Peterson C, Ambartsumyan G, Aimiuwu O, Richter L, Zhang J, Khvorostov I, Ott V, Grunstein M, et al. Induced pluripotent stem cells and embryonic stem cells are distinguished by gene expression signatures. Cell Stem Cell. 2009; 5: 111 -123. [PubMed] .
- 75. Okita K, Ichisaka T, Yamanaka S. Generation of germline-competent induced pluripotent stem cells. Nature. 2007; 448: 313 -317. [PubMed] .
- 76. Trounson A, Thakar RG, Lomax G, Gibbons D. Clinical trials for stem cell therapies. BMC Med. 2011; 9: 52 [PubMed] .
- 77. Noren Hooten N and Evans MK. The ultimate transformers: mesenchymal stem cells. Cell Cycle. 2011; 10: 4189 -4190. [PubMed] .
- 78. Egea V, von Baumgarten L, Schichor C, Berninger B, Popp T, Neth P, Goldbrunner R, Kienast Y, Winkler F, Jochum M, Ries C. TNF-alpha respecifies human mesenchymal stem cells to a neural fate and promotes migration toward experimental glioma. Cell Death Differ. 2011; 18: 853 -863. [PubMed] .
- 79. Djouad F, Plence P, Bony C, Tropel P, Apparailly F, Sany J, Noel D, Jorgensen C. Immunosuppressive effect of mesenchymal stem cells favors tumor growth in allogeneic animals. Blood. 2003; 102: 3837 -3844. [PubMed] .
- 80. Semont A, Mouiseddine M, Francois A, Demarquay C, Mathieu N, Chapel A, Sache A, Thierry D, Laloi P, Gourmelon P. Mesenchymal stem cells improve small intestinal integrity through regulation of endogenous epithelial cell homeostasis. Cell Death Differ. 2010; 17: 952 -961. [PubMed] .
- 81. Tome M, Lopez-Romero P, Albo C, Sepulveda JC, Fernandez-Gutierrez B, Dopazo A, Bernad A, Gonzalez MA. miR-335 orchestrates cell proliferation, migration and differentiation in human mesenchymal stem cells. Cell Death Differ. 2011; 18: 985 -995. [PubMed] .
- 82. Zhu W, Huang L, Li Y, Qian H, Shan X, Yan Y, Mao F, Wu X, Xu WR. Mesenchymal stem cell-secreted soluble signaling molecules potentiate tumor growth. Cell Cycle. 2011; 10: 3198 -3207. [PubMed] .
- 83. Anastasia L, Pelissero G, Venerando B, Tettamanti G. Cell reprogramming: expectations and challenges for chemistry in stem cell biology and regenerative medicine. Cell Death Differ. 2010; 17: 1230 -1237. [PubMed] .
- 84. Sarig R and Rotter V. Can an iPS cell secure its genomic fidelity? Cell Death Differ. 2011; 18: 743 -744. [PubMed] .
- 85. Hussein SM, Batada NN, Vuoristo S, Ching RW, Autio R, Narva E, Ng S, Sourour M, Hamalainen R, Olsson C, Lundin K, Mikkola M, Trokovic R, et al. Copy number variation and selection during reprogramming to pluripotency. Nature. 2011; 471: 58 -62. [PubMed] .
- 86. Pasi CE, Dereli-Oz A, Negrini S, Friedli M, Fragola G, Lombardo A, Van Houwe G, Naldini L, Casola S, Testa G, Trono D, Pelicci PG, Halazonetis TD. Genomic instability in induced stem cells. Cell Death Differ. 2011; 18: 745 -753. [PubMed] .
- 87. Gore A, Li Z, Fung HL, Young JE, Agarwal S, Antosiewicz-Bourget J, Canto I, Giorgetti A, Israel MA, Kiskinis E, Lee JH, Loh YH, Manos PD, et al. Somatic coding mutations in human induced pluripotent stem cells. Nature. 2011; 471: 63 -67. [PubMed] .
- 88. Lister R, Pelizzola M, Kida YS, Hawkins RD, Nery JR, Hon G, Antosiewicz-Bourget J, O'Malley R, Castanon R, Klugman S, Downes M, Yu R, Stewart R, et al. Hotspots of aberrant epigenomic reprogramming in human induced pluripotent stem cells. Nature. 2011; 471: 68 -73. [PubMed] .
- 89. Bogomazova AN, Lagarkova MA, Tskhovrebova LV, Shutova MV, Kiselev SL. Error-prone nonhomologous end joining repair operates in human pluripotent stem cells during late G2. Aging (Albany NY). 2011; 3: 584 -596. [PubMed] .
- 90. Ohmine S, Squillace KA, Hartjes KA, Deeds MC, Armstrong AS, Thatava T, Sakuma T, Terzic A, Kudva Y, Ikeda Y. Reprogrammed keratinocytes from elderly type 2 diabetes patients suppress senescence genes to acquire induced pluripotency. Aging (Albany NY). 2012; 4: 60 -73. [PubMed] .
- 91. Park IH, Arora N, Huo H, Maherali N, Ahfeldt T, Shimamura A, Lensch MW, Cowan C, Hochedlinger K, Daley GQ. Disease-specific induced pluripotent stem cells. Cell. 2008; 134: 877 -886. [PubMed] .
- 92. Dimos JT, Rodolfa KT, Niakan KK, Weisenthal LM, Mitsumoto H, Chung W, Croft GF, Saphier G, Leibel R, Goland R, Wichterle H, Henderson CE, Eggan K. Induced pluripotent stem cells generated from patients with ALS can be differentiated into motor neurons. Science. 2008; 321: 1218 -1221. [PubMed] .
- 93. Pfisterer U, Wood J, Nihlberg K, Hallgren O, Bjermer L, Westergren-Thorsson G, Lindvall O, Parmar M. Efficient induction of functional neurons from adult human fibroblasts. Cell Cycle. 2011; 10: 3311 -3316. [PubMed] .
- 94. Brennand KJ, Simone A, Jou J, Gelboin-Burkhart C, Tran N, Sangar S, Li Y, Mu Y, Chen G, Yu D, McCarthy S, Sebat J, Gage FH. Modelling schizophrenia using human induced pluripotent stem cells. Nature. 2011; 473: 221 -225. [PubMed] .
- 95. Ye Z, Liu CF, Jang YY. Hematopoietic cells as sources for patient-specific iPSCs and disease modeling. Cell Cycle. 2011; 10: 2840 -2844. [PubMed] .
- 96. Ho JC, Zhou T, Lai WH, Huang Y, Chan YC, Li X, Wong NL, Li Y, Au KW, Guo D, Xu J, Siu CW, Pei D, et al. Generation of induced pluripotent stem cell lines from 3 distinct laminopathies bearing heterogeneous mutations in lamin A/C. Aging (Albany NY). 2011; 3: 380 -390. [PubMed] .