



Introduction
Cells are continuously exposed to a wide variety of physical and chemical stresses such as oxidation, radiation and heavy metals, which cause damage to cellular proteins, lipids and DNA. Organisms have evolved multiple protective mechanisms to counteract these endogenous and exogenous damages. Nevertheless, the effectiveness of these protective pathways seems to decline with age. As such, aging can be defined as the decrease in the probability of successful repair of cellular damage.
One of the major sources of cellular insults is damage to DNA. To counteract detrimental DNA damage, cells are endowed with a complex network of DNA damage response (DDR) proteins which are capable of detecting DNA damage, and then triggering and amplifying a signaling cascade, which ultimately leads to either cell-cycle arrest and DNA repair, or to apoptotic cell death to eliminate permanently damaged cells [1]. The importance of the DDR in maintaining genomic integrity and limiting the effects of aging is highlighted by premature aging phenotype of mice that lack key DNA repair factors [2]. More tellingly, almost all genetic conditions that lead to premature aging in humans have been mapped to genes belonging to the DDR [3]. Mutations in the Werner DNA helicase, which is required for DNA replication and at telomeres, lead to Werner syndrome and the components of the nucleotide excision repair (NER) XPC, ERCC6, ERCC8 and the ERCC1/XPF complex involved in inter-strand DNA crosslink repair are mutated in Cockayne Syndrome and in Trichothiodystrophy (TTD), two prominent premature aging disorders [4]. These findings suggest a prominent, and causal, role for DNA damage responses in aging.
The DDR, like all major nuclear processes such as DNA replication and transcription, operates in the context of the chromatin fiber [5,6]. Chromatin is made up of nucleosomes, repetitive units of 146bp of DNA tightly wrapped around an octameric core of histone proteins (H2A, H2B, H3 and H4). Nucleosomes are further packaged into higher order structures by the action of architectural chromatin proteins such as histone H1 and heterochromatin protein HP1. Based on cytological criteria, chromatin is classified into euchromatin, which is loosely packed and generally transcriptionally active, and into heterochromatin, which is more compacted and generally represents a transcriptionally repressive environment. Nucleosomal histones are modified by complex patterns of post-translational modifications (PTM) such as acetylation, methylation and ubiquitination which appear to dictate the dynamic recruitment of non-histone proteins to chromatin and regulate its function [7]. Furthermore, chromatin structure and function is also determined by the methylation status of DNA itself and by a large number of ATP-dependent remodeling factors. Both the level of chromatin compaction, and hence the accessibility of DNA, and the recruitment of chromatin-associated factors determine the outcome of transcription, DNA replication and DNA damage repair. All these modifications to chromatin structure, and thus its informational content, are inherited through several cycles of cell division and as such represent an epigenetic memory [8].
Chromatin defects in aging
Chromatin defects are associated with aging. The first hints pointing to a possible link between chromatin maintenance and aging came from studies in the yeast S. cerevesiae, where the NADH-dependent Sir2 histone deacetylase Sir2 was found to be important for establishing heterochromatin at telomeres, at ribosomal DNA (rDNA), and at HMR and HMR loci, which encode factors needed for yeast mating type switching [9-14]. Upon prolonged growth, equated to aging in yeast, repetitive rDNA tends to hyper-recombine and form extrachromosomal rDNA circles (ERC), indicative of increased chromatin fragility [15]. Formation of heterochromatin at rDNA sites by overexpression of Sir2 reduces this hyper-recombination and prolongs lifespan, suggesting a contribution of chromatin structure to aging [16]. Further experiments in worms and flies demonstrated a similar role in lifespan extension for Sirt1, the closest orthologue of yeast Sir2 in these organisms [17,18]. Nevertheless, the role of Sirt1 in increased longevity in higher eukaryotes might not just involve heterochromatin maintenance, since in this case the molecular mechanism does not seem to involve ERC stabilization [19]. Furthermore, the analysis is complicated by the fact that in mammals SIRT1 deacetylates a wide variety of non-histone, aging-related transcription factors such as p53, HSF1 and members of the FOXO transcription factors family [20-22]. Identification of the mechanisms of action of SIRT1 in higher organisms will be key to clarifying its role in the aging process.
There are several other clear indications for a role of chromatin and its maintenance in aging. A hallmark of cellular aging is the appearance of characteristic changes in the epigenetic make-up of the genome. Epigenetic changes associated with aging in mammalian cells include loss of DNA methylation at repetitive DNA sequences [23-25], which are generally heterochromatinized, and an increase in DNA methylation at CpG islands in the promoters of specific genes [26,27]. Cells from aged individuals and patients with the premature aging disorder Hutchinson-Gilford Progeria Syndrome (HGPS) are also characterized by loss of heterochromatin, by loss of key architectural chromatin proteins such as HP1 and the histone mehtyltransferase EZH2, and, importantly, by alterations in the levels of heterochromatin-associated histone PTM including H3K9me3 and H3K27me3 [28-31]. Interestingly, both prematurely and normally aged cells exhibit dramatically increased levels of unrepaired DNA damage [30,32].
In addition to epigenetic and structural chromatin defects, there are indications that aging in mammals is accompanied by stochastic deregulation of gene expression. Transcriptional noise at the single cell level increases with age in the mouse heart, most likely as a consequence of oxidative DNA damage [33]. Furthermore, in mammalian cells oxidative DNA damage also seems to relocalize SIRT1 from otherwise transcriptionally repressed genes to sites of DNA damage [34]. This has led to the speculation that, through unknown mechanisms, aging disrupts the epigenetic organization of heterochromatin both at a global and at a gene-specific level, thus leading to elevation of stochastic transcriptional noise and to the disruption of transcriptional programs necessary for proper cell homeostasis [35]. In contrast to this model of stochastically occurring defects in gene expression programs, the aging process seems to also induce a specific transcriptional response, which dampens the somatotrophic IGF-1 axis and helps protecting cells from DNA damage and stress [36].
The study of chromatin in aging also points to a key influence of aberrant chromatin structure on aging-related defects in DNA repair. Impairment of SIRT1 leads to defective DNA damage repair in mammalian cells [34] and a knock-out mouse model for SIRT6 shows signs of premature aging and has defects in the base excision repair pathway [37]. The exact molecular basis for these phenotypes is not clear yet. One possibility is that SIRT6 affects genomic stability by regulating the levels of H3K56Ac [38,39], a PTM important for chromatin assembly and DNA damage tolerance in yeast [40,41].
A molecular mechanism for aging-associated chromatin defects
The molecular mechanisms leading to chromatin defects in aging are largely unknown. Recent analysis of chromatin defects in the premature aging disease HGPS have given some of the first insights into how chromatin ages [42]. HGPS is an extremely rare genetic disease caused by a de novo point mutation in the lamin A (LMNA) gene, a major structural component of the nuclear envelope [43].The pathogenic mutation leads to the production of an internally truncated form of lamin A, referred to as progerin. This protein acts in a dominant-negative gain of function fashion causing the diverse and pronounced chromatin defects. Analysis of the molecular mechanisms involved in bringing about chromatin defects in HGPS and old cells uncovered the NURD complex as a key player in aging [42]. NURD is a ubiquitous chromatin remodeling complex which contains the histone deacetylases HDAC1 and HDAC2 and the ATPases CHD3 and CHD4 as catalytic subunits. NURD has been implicated in transcriptional repression at specific promoters and more recently has also been shown to associate with pericentromeric heterochromatin [44,45]. The protein levels and the activity of several NURD components including HDAC1 and the histone chaperones RBBP4/, are reduced in HGPS cells and normally aged cells. A direct role for NURD loss in aging-associated chromatin defects is indicated by the finding that knock-down of NURD members in normal cells recapitulates aging-related chromatin defects including heterochromatin loss and increased DNA damage [42]. NURD is known to be involved in a variety of chromatin functions and its loss may explain the broad spectrum of chromatin defects seen in aged cells [42].
Chromatin structure as a trigger of aging
There is little doubt that chromatin defects and DNA damage play a part in the aging process. The unresolved question is: how? One recently proposed scenario suggests that DNA damage and the cellular response to it leads to chromatin defects via relocation of epigenetic machinery from its normal distribution in the genome and to structural chromatin changes, eventually resulting in gene misregulation [34] (Figure 1A). An alternative possibility is that the aging process is triggered by loss of chromatin structure, leading to altered epigenetic modifications, and increased susceptibility to DNA damage. In this model DNA damage is a downstream event (Figure 1B). The key question to distinguish between these two models is: what comes first, DNA damage or chromatin defects? A partial answer comes from recent observations in the premature aging disorder HGPS. Upon induction of the dominant negative disease-causing protein in normal skin fibroblasts, chromatin defects occurred prior to DNA damage [42][41]. Further support for a trigger role of chromatin structure in DNA damage and aging, is the observation that suppression of the activity of chromatin modifiers generates high levels of endogenous DNA damage, as seen in the case of several subunits of the NURD complex [42], the SET8 H4K20 histone methylase [46,47], and for the Su(var)3-9 H3K9 histone methylase in Drosophila [48]. In these cases chromatin structural defects clearly precede DNA damage, placing epigenetic and chroma-tin structure changes upstream of DNA damage events.
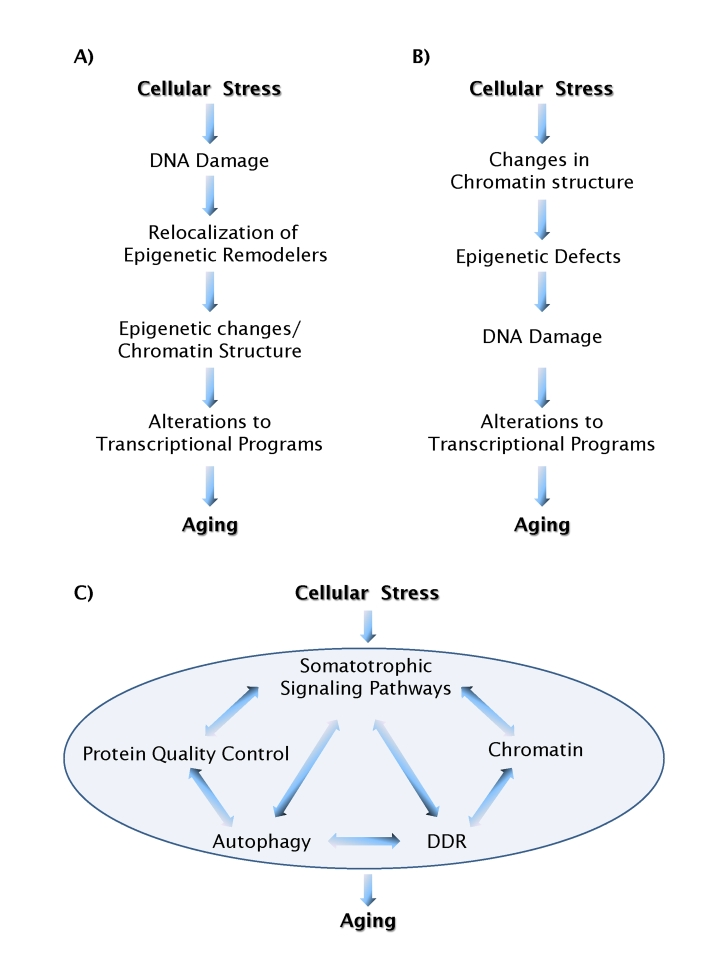
Figure 1. Models of aging pathways. (A) A scenario in which DNA
damage acts as a causal trigger for aging. (B) A scenario in which
chromatin structure acts as a causal trigger for aging. Feedback loops,
which are likely to exist between most individual events, are not shown for
simplicity. (C) Chromatin structure and DNA damage pathways act in
an integrated fashion with a multitude of other cellular process to form a
network of aging processes.
How may aberrant chromatin structure lead to DNA damage and aging? Although only poorly investigated and understood, it is becoming clear that chromatin structure affects the susceptibility of DNA to damage and progression of the DDR [5]. DNA repair occurs with slower kinetics in highly condensed heterochromatin, presumably due to the inability of repair factors to rapidly access the site of damage [49]. Furthermore, heterochromatinized regions of the genome, like nucleoli, centromeres and telomeres tend to be rich in repetitive sequences that are particularly prone to recombination. As such it is possible that the compacted nature of heterochromatin suppresses hyper-recombination of repetitive sequences, the formation of aberrant DNA structures and genomic instability [50]. Another, not-mutually exclusive, possibility is that altered chromatin structure increases the steady-state level of DNA damage due to replication defects such as impaired passage of the replication machinery or to replication fork stalling. It is indeed possible that intact heterochromatin conformation is necessary for the DNA replication machinery to properly proceed through highly repetitive portions of the genome. This last hypothesis is in line with the observation that siRNA silencing of either the histone-chaperones RBBP4/7 [42] or of SET8 [46,47] impairs S-phase progression.
Although these observations point towards an upstream role of chromatin structure in determining DNA stability, it is also true that genome integrity influences chromatin structure. Local DNA damage affects the epigenetic status of chromatin both in the vicinity of a lesion through phosphorylation, acetylation and ubiquitination of nearby histones, but also globally [51]. In response to local DNA damage, the zinc finger protein KAP1 is phosporylated by the ATM kinase and released from heterochromatin, thus facilitating the access of DNA repair factors to these more compacted regions of the genome and also potentially altering chromatin structure at other sites [51]. Furthermore, as previously mentioned, DNA damage results in redistribution of chromatin associated factors and histone modifiers like SIRT1, possibly leading to profound changes in the transcriptional regulation of genes [34]. Clearly, the relationship between chromatin structure and DNA damage is not unidirectional, but rather a mutual one.
A network of aging mechanisms
In our search of molecular mechanisms for biological processes we usually look for linear pathways. What we are learning about the interplay between chromatin structure, epigenetic regulation and DNA repair makes it clear that this is not a one-way street and that these processes are likely connected and linked by feedback mechanisms. The most likely scenario is that chromatin structure, epigenetic status, and DNA repair represent nodes of a network of processes involved in protecting cells from endogenous and exogenous insults, ultimately leading to increased longevity (Figure 1C). Importantly, these processes do not work in isolation, but instead are linked to pathways dedicated to maintain proteostasis such as the heat shock response or autophagy, and hormonal regulation of cellular growth, with the mTOR and IGF-1 pathways, whose role in the regulation of longevity has already been established in mammals [52-54]. In support of a branched network of cellular functions involved in aging, possible connections between the DNA damage, the inhibition of the IGF-I and mTOR pathways have been suggested [55]. This scenario is supported by observations in cells from the ZMPSTE24-/- mouse, a murine model of HGPS in which lamin A processing is impaired. The progeriod ZEMPSTE24-/- mouse shows dramatic alterations in heterochromatin architecture, accompanied by increased DNA damage, by the activation of the authophagic response and by downregulation of the mTOR pathway [32,56].
Aging is a complex process. It is hardly realistic for it to be explained by a single pathway or even a set of closely related pathways. More likely, many diverse cellular functions will contribute to aging and they will do so in a highly inter-dependent manner. The recent investigation of the role of chromatin structure, epigenetic modifications and DNA damage in aging makes this clear. While we are still struggling to understand the precise relationship of these events in the aging process, we are already discovering links to more distantly related events such as signaling pathways and metabolism. Rather than attempting to explain aging as the consequence of degeneration of single pathways, a conceptual framework consisting of a network of affected processes not only reconciles different, at times contentious hypotheses regarding aging mechanisms, but will ultimately lead to an integrated view of these processes and to a more accurate understanding of the molecular basis of aging.
Conflicts of Interest
The authors of this manuscript have no conflict of interest to declare.
References
- 1. Misteli T and Soutoglou E. The emerging role of nuclear architecture in DNA repair and genome maintenance. Nat Rev Mol Cell Biol. 2009; 10: 243 -254. [PubMed] .
- 2. Lombard DB , Chua KF , Mostoslavsky R , Franco S , Gostissa M and Alt FW. DNA repair, genome stability, and aging. Cell. 2005; 120: 497 -512. [PubMed] .
- 3. Ramirez CL , Cadinanos J , Varela I , Freije JM and Lopez-Otin C. Human progeroid syndromes, aging and cancer: new genetic and epigenetic insights into old questions. Cell Mol Life Sci. 2007; 64: 155 -170. [PubMed] .
- 4. Andressoo JO , Hoeijmakers JH and Mitchell JR. Nucleotide excision repair disorders and the balance between cancer and aging. Cell Cycle. 2006; 5: 2886 -2888. [PubMed] .
- 5. Groth A , Rocha W , Verreault A and Almouzni G. Chromatin challenges during DNA replication and repair. Cell. 2007; 128: 721 -733. [PubMed] .
- 6. Li B , Carey M and Workman JL. The role of chromatin during transcription. Cell. 2007; 128: 707 -719. [PubMed] .
- 7. Kouzarides T Chromatin modifications and their function. Cell. 2007; 128: 693 -705. [PubMed] .
- 8. Probst AV , Dunleavy E and Almouzni G. Epigenetic inheritance during the cell cycle. Nat Rev Mol Cell Biol. 2009; 10: 192 -206. [PubMed] .
- 9. Imai S , Armstrong CM , Kaeberlein M and Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000; 403: 795 -800. [PubMed] .
- 10. Rine J and Herskowitz I. Four genes responsible for a position effect on expression from HML and HMR in Saccharomyces cerevisiae. Genetics. 1987; 116: 9 -22. [PubMed] .
- 11. Ivy JM , Klar AJ and Hicks JB. Cloning and characterization of four SIR genes of Saccharomyces cerevisiae. Mol Cell Biol. 1986; 6: 688 -702. [PubMed] .
- 12. Gottschling DE , Aparicio OM , Billington BL and Zakian VA. Position effect at S. cerevisiae telomeres: reversible repression of Pol II transcription. Cell. 1990; 63: 751 -762. [PubMed] .
- 13. Bryk M , Banerjee M , Murphy M , Knudsen KE , Garfinkel DJ and Curcio MJ. Transcriptional silencing of Ty1 elements in the RDN1 locus of yeast. Genes Dev. 1997; 11: 255 -269. [PubMed] .
- 14. Smith JS and Boeke JD. An unusual form of transcriptional silencing in yeast ribosomal DNA. Genes Dev. 1997; 11: 241 -254. [PubMed] .
- 15. Sinclair DA and Guarente L. Extrachromosomal rDNA circles--a cause of aging in yeast. Cell. 1997; 91: 1033 -1042. [PubMed] .
- 16. Kaeberlein M , McVey M and Guarente L. The SIR2/3/4 complex and SIR2 alone promote longevity in Sacharomyces cerevisiae by two different mechanisms. Genes Dev. 1999; 13: 2570 -2580. [PubMed] .
- 17. Rogina B and Helfand SL. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc Natl Acad Sci U S A. 2004; 101: 15998 -16003. [PubMed] .
- 18. Tissenbaum HA and Guarente L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature. 2001; 410: 227 -230. [PubMed] .
- 19. Guarente L and Picard F. Calorie restriction--the SIR2 connection. Cell. 2005; 120: 473 -482. [PubMed] .
- 20. Luo J , Nikolaev AY , Imai S , Chen D , Su F , Shiloh A , Guarente L and Gu W. Negative control of p53 by Sir2alpha promotes cell survival under stress. Cell. 2001; 107: 137 -148. [PubMed] .
- 21. Westerheide SD , Anckar J , Stevens SM Jr , Sistonen L and Morimoto RI. Stress-inducible regulation of heat shock factor 1 by the deacetylase SIRT1. Science. 2009; 323: 1063 -1066. [PubMed] .
- 22. Brunet A , Sweeney LB , Sturgill JF , Chua KF , Greer PL , Lin Y , Tran H , Ross SE , Mostoslavsky R , Cohen HY , Hu LS , Cheng HL and Jedrychowski MP. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science. 2004; 303: 2011 -2015. [PubMed] .
- 23. Wilson VL, Smith RA, Ma S, Cutler RG. Genomic 5-methyldeoxycytidine decreases with age. J Biol Chem. 1987; 262: 9948 -9951. [PubMed] .
- 24. Fuke C , Shimabukuro M , Petronis A , Sugimoto J , Oda T , Miura K , Miyazaki T , Ogura C , Okazaki Y and Jinno Y. Age related changes in 5-methylcytosine content in human peripheral leukocytes and placentas: an HPLC-based study. Ann Hum Genet. 2004; 68: 196 -204. [PubMed] .
- 25. Bjornsson HT , Sigurdsson MI , Fallin MD , Irizarry RA , Aspelund T , Cui H , Yu W , Rongione MA , Ekstrom TJ , Harris TB , Launer LJ , Eiriksdottir G and Leppert MF. Intra-individual change over time in DNA methylation with familial clustering. JAMA. 2008; 299: 2877 -2883. [PubMed] .
- 26. Issa JP, Ottaviano YL, Celano P, Hamilton SR, Davidson NE, Baylin SB. Methylation of the oestrogen receptor CpG island links ageing and neoplasia in human colon. Nat Genet. 1994; 7: 536 -540. [PubMed] .
- 27. Ahuja N , Li Q , Mohan AL , Baylin SB and Issa JP. Aging and DNA methylation in colorectal mucosa and cancer. Cancer Res. 1998; 58: 5489 -5494. [PubMed] .
- 28. Goldman RD, Shumaker DK, Erdos MR, Eriksson M, Goldman AE, Gordon LB, Gruenbaum Y, Khuon S, Mendez M, Varga R, Collins FS. Accumulation of mutant lamin A causes progressive changes in nuclear architecture in Hutchinson-Gilford progeria syndrome. Proc Natl Acad Sci U S A. 2004; 101: 8963 -8968. [PubMed] .
- 29. Scaffidi P and Misteli T. Reversal of the cellular phenotype in the premature aging disease Hutchinson-Gilford progeria syndrome. Nat Med. 2005; 11: 440 -445. [PubMed] .
- 30. Scaffidi P and Misteli T. Lamin A-dependent nuclear defects in human aging. Science. 2006; 312: 1059 -1063. [PubMed] .
- 31. Shumaker DK , Dechat T , Kohlmaier A , Adam SA , Bozovsky MR , Erdos MR , Eriksson M , Goldman AE , Khuon S , Collins FS , Jenuwein T and Goldman RD. Mutant nuclear lamin A leads to progressive alterations of epigenetic control in premature aging. Proc Natl Acad Sci U S A. 2006; 103: 8703 -8708. [PubMed] .
- 32. Liu B , Wang J , Chan KM , Tjia WM , Deng W , Guan X , Huang JD , Li KM , Chau PY , Chen DJ , Pei D , Pendas AM and Cadinanos J. Genomic instability in laminopathy-based premature aging. Nat Med. 2005; 11: 780 -785. [PubMed] .
- 33. Bahar R , Hartmann CH , Rodriguez KA , Denny AD , Busuttil RA , Dolle ME , Calder RB , Chisholm GB , Pollock BH , Klein CA and Vijg J. Increased cell-to-cell variation in gene expression in ageing mouse heart. Nature. 2006; 441: 1011 -1014. [PubMed] .
- 34. Oberdoerffer P , Michan S , McVay M , Mostoslavsky R , Vann J , Park SK , Hartlerode A , Stegmuller J , Hafner A , Loerch P , Wright SM , Mills KD and Bonni A. SIRT1 redistribution on chromatin promotes genomic stability but alters gene expression during aging. Cell. 2008; 135: 907 -918. [PubMed] .
- 35. Sinclair DA and Oberdoerffer P. The ageing epigenome: damaged beyond repair. Ageing Res Rev. 2009; 8: 189 -198. [PubMed] .
- 36. Niedernhofer LJ , Garinis GA , Raams A , Lalai AS , Robinson AR , Appeldoorn E , Odijk H , Oostendorp R , Ahmad A , van Leeuwen W , Theil AF , Vermeulen W and van der Horst GT. A new progeroid syndrome reveals that genotoxic stress suppresses the somatotroph axis. Nature. 2006; 444: 1038 -1043. [PubMed] .
- 37. Mostoslavsky R , Chua KF , Lombard DB , Pang WW , Fischer MR , Gellon L , Liu P , Mostoslavsky G , Franco S , Murphy MM , Mills KD , Patel P and Hsu JT. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell. 2006; 124: 315 -329. [PubMed] .
- 38. Yang B , Zwaans BM , Eckersdorff M and Lombard DB. The sirtuin SIRT6 deacetylates H3 K56Ac in vivo to promote genomic stability. Cell Cycle. 2009; 8: 2662 -2663. [PubMed] .
- 39. Michishita E , McCord RA , Boxer LD , Barber MF , Hong T , Gozani O and Chua KF. Cell cycle-dependent deacetylation of telomeric histone H3 lysine K56 by human SIRT6. Cell Cycle. 2009; 8: 2664 -2666. [PubMed] .
- 40. Masumoto H , Hawke D , Kobayashi R and Verreault A. A role for cell-cycle-regulated histone H3 lysine 56 acetylation in the DNA damage response. Nature. 2005; 436: 294 -298. [PubMed] .
- 41. Li Q , Zhou H , Wurtele H , Davies B , Horazdovsky B , Verreault A and Zhang Z. Acetylation of histone H3 lysine 56 regulates replication-coupled nucleosome assembly. Cell. 2008; 134: 244 -255. [PubMed] .
- 42. Pegoraro G , Kubben N , Wickert U , Gohler H , Hoffmann K and Misteli T. Ageing-related chromatin defects through loss of the NURD complex. Nat Cell Biol. 2009; 11: 1261 -1267. [PubMed] .
- 43. Kudlow BA , Kennedy BK and Monnat RJ Jr. Werner and Hutchinson-Gilford progeria syndromes: mechanistic basis of human progeroid diseases. Nat Rev Mol Cell Biol. 2007; 8: 394 -404. [PubMed] .
- 44. Bowen NJ , Fujita N , Kajita M and Wade PA. Mi-2/NuRD: multiple complexes for many purposes. Biochim Biophys Acta. 2004; 1677: 52 -57. [PubMed] .
- 45. Helbling Chadwick L , Chadwick BP , Jaye DL and Wade PA. The Mi-2/NuRD complex associates with pericentromeric heterochromatin during S phase in rapidly proliferating lymphoid cells. Chromosoma. 2009; 118: 445 -457. [PubMed] .
- 46. Jorgensen S , Elvers I , Trelle MB , Menzel T , Eskildsen M , Jensen ON , Helleday T , Helin K and Sorensen CS. The histone methyltransferase SET8 is required for S-phase progression. J Cell Biol. 2007; 179: 1337 -1345. [PubMed] .
- 47. Tardat M , Murr R , Herceg Z , Sardet C and Julien E. PR-Set7-dependent lysine methylation ensures genome replication and stability through S phase. J Cell Biol. 2007; 179: 1413 -1426. [PubMed] .
- 48. Peng JC and Karpen GH. Heterochromatic genome stability requires regulators of histone H3 K9 methylation. PLoS Genet. 2009; 5: e1000435 [PubMed] .
- 49. Goodarzi AA , Noon AT , Deckbar D , Ziv Y , Shiloh Y , Lobrich M and Jeggo PA. ATM signaling facilitates repair of DNA double-strand breaks associated with heterochromatin. Mol Cell. 2008; 31: 167 -177. [PubMed] .
- 50. Peng JC and Karpen GH. Epigenetic regulation of heterochromatic DNA stability. Curr Opin Genet Dev. 2008; 18: 204 -211. [PubMed] .
- 51. Ziv Y , Bielopolski D , Galanty Y , Lukas C , Taya Y , Schultz DC , Lukas J , Bekker-Jensen S , Bartek J and Shiloh Y. Chromatin relaxation in response to DNA double-strand breaks is modulated by a novel ATM- and KAP-1 dependent pathway. Nat Cell Biol. 2006; 8: 870 -876. [PubMed] .
- 52. Holzenberger M , Dupont J , Ducos B , Leneuve P , Geloen A , Even PC , Cervera P and Le Bouc Y. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature. 2003; 421: 182 -187. [PubMed] .
- 53. Harrison DE , Strong R , Sharp ZD , Nelson JF , Astle CM , Flurkey K , Nadon NL , Wilkinson JE , Frenkel K , Carter CS , Pahor M , Javors MA and Fernandez E. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature. 2009; 460: 392 -395. [PubMed] .
- 54. Selman C , Tullet JM , Wieser D , Irvine E , Lingard SJ , Choudhury AI , Claret M , Al-Qassab H , Carmignac D , Ramadani F , Woods A , Robinson IC and Schuster E. Ribosomal protein S6 kinase 1 signaling regulates mammalian life span. Science. 2009; 326: 140 -144. [PubMed] .
- 55. Niedernhofer LJ and Robbins PD. Signaling mechanisms involved in the response to genotoxic stress and regulating lifespan. Int J Biochem Cell Biol. 2008; 40: 176 -180. [PubMed] .
- 56. Marino G, Ugalde AP, Salvador-Montoliu N, Varela I, Quiros PM, Cadinanos J, van der Pluijm I, Freije JM, Lopez-Otin C. Premature aging in mice activates a systemic metabolic response involving autophagy induction. Hum Mol Genet. 2008; 17: 2196 -2211. [PubMed] .