



Introduction
The oxidative stress caused by reactive oxygen species (ROS) is assumed to significantly contribute to aging and numerous age-related pathologies. Mitochondria are known as a place, where the most intensive ROS production can occur. In the recent years, mitochondria-targeted antioxidants has been developed [1-4]. Research was the series of papers published by our group in 1969-1970, where mitochondria-addressed penetrating synthetic cations were described and the idea to use these cations as "electric locomotives" targeting non-charged compounds to mitochondria was put forward [5,6]. In the late nineties, Murphy and coworkers initiated the practical realization of this idea [1,7-9]. They synthesized and tested several mitochondria-targeted antioxidants conjugated to the lipophilic alkyltriphenylphosphonium cations. The ubiquinone moiety linked to triphenylphosphonium cation by C10 aliphatic chain, MitoQ (Figure 1), seemed to be the most promising [1,4,9].
In 2005, an attempt was undertaken in our group to replace the ubiquinone moiety in MitoQ by plasto-quinone. As a result, a series of mitochondria-targeted antioxidants named SkQ has been synthesized [2,10]. There were two main reasons for this modification [1]. Plastoquinone playing in chloroplasts the same role of an electron carrier as ubiquinone does in mitochondria always operates under conditions of oxidative stress (elevated oxygen concentration and an intensive ROS production) [2]. It was reported [11-13] that the reactivity of the "tailless" plastoquinol analogs to the peroxyl radicals was indeed higher than that of natural ubiquinols. The advantage of mitochondria-targeted quinones of SkQ type over MitoQ was recently demonstrated by using several biological models. In particular, it was found that very low doses of SkQ1 (nmol/kg per day) prolong life of podospora, ceriodaphnia, drosophila and mice. In mice, SkQ1 doubled median lifespan arrested development of such traits of the senescence process as involution of thymus and decline of other immunity mechanisms; osteoporosis; disappearance of regular estrous cycles in females, cataract, retinopathies, balding, catinies, hypothermia, chromosome aberrations, peroxidation of lipids and proteins, etc. [10,14-20].
Figure 1.
The structure of the mitochondria-targeted
hydroquinones and other phenolics studied in this work.
Until recently, the reactivity of the mitochondria-targeted antioxidants has, in fact, not been quantitatively determined. This was done in the present paper. The structure of the compounds studied is presented in Figure 1. The chain-breaking antioxidant activity was characterized by the rate constant for reaction of QH2 with the lipid peroxyl radical, LO2•, formed from ML or cardiolipin: LO2• + QH2¾→ LOOH + QH• k1 [1] which competes with the reaction of chain propagation of lipid peroxidation LO2• + LH (+O2) ¾→ LOOH + LO2• k2[2].
Results
Figure 2 shows that SkQ1 is almost completely reduced to SkQ1H2 by NaBH4. For SkQ1, the m/z value was found to be 537.08, which corresponds to the theoretically calculated one. As expected, the m/z value for SkQ1H2 proved to be 539.1, i.e. m/z increased by two units as compared with that for SkQ1. Similar results were also obtained for the reduction of other mitochondria-targeted quinones.
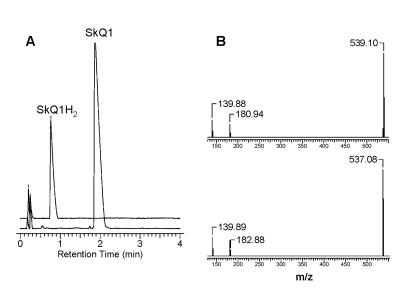
Figure 2. The reduction of
SkQ1 by NaBH4 as studied by UPLC-MS-MS analysis. (A) -
Reverse-phase HPLC chromatograms before and after the addition of NaBH4.
(B) - MS/MS spectra of SkQ1 before reduction (at the bottom) and after
reduction (at the top). Details of the protocol are given in the text.
The non-inhibited oxidation of ML in Triton micelles is a chain process, which rate, R0, was found to be proportional to [ML] and square root of [AAPH] (not shown) as it was reported in our preceding papers [21,22]. Such relationships are also inherent in the lipid peroxidation in other aqueous microheterogeneous systems [23-25]. They correspond to the "classic" kinetic scheme with bimolecular chain termination [26,27].
AAPH + LH + (O2) ¾→ LO2• + products RIN (0) LO2• + LH + ¾→ LOOH + L• k2 [2] L• + O2¾→ LO2• k3 [3] LO2• + LO2•¾→ products 2k4 [4] All the tested QH2 displayed a pronounced chain-breaking antioxidant activity as this is exemplified by Figure 3 for SkQ1H2. When SkQ1H2 was added, the rate of oxidation, R, dramatically decreased. As SkQ1H2 was progressively consumed due to reaction [1], R increased with time and eventually reaches the level of non-inhibited oxidation. As a result, the pronounced induction period was observed (Figure 3A).
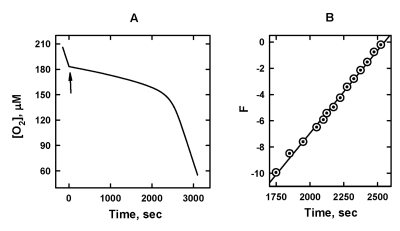
Figure 3. The effect of 5 μM SkQ1H2 on the
kinetics of oxygen consumption caused by oxidation of 20 mM ML in micellar
solution of 50 mM Triton X-100 in 50mM phosphate buffer, pH 7.4, 37 °C. Oxidation was initiated by 3
mM AAPH. (A) [O2] trace; arrow shows addition of SkQ1H2.
(B) plot A in the axes of Eq. 7.
Quantitatively similar [O2] traces were observed with all the other tested QH2 as well as with α-tocopherol and its synthetic analog 6-hydroxy-2,2,5,7,8-pentamethylchromane (HPMC). As for C12TPP, a compound that has no hydroquinone moiety (Figure 1), it did not display any inhibiting activity (not shown). Meanwhile, oxidized form of SkQ1 showed a weak inhibition of ML oxidation, but only during a very short period of time (Figure 4). Most likely, the inhibition is caused in this case by a minor contamination of SkQ1H2 to SkQ1. A similar effect was also observed with other mitochondria-targeted Q. This suggests that mitochondria-targeted quinones by themselves do not act as a chain-breaking antioxidant.
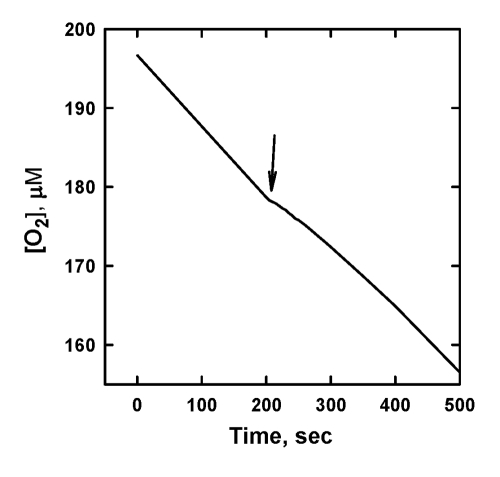
Figure 4. The effect of
addition of 10 μM SkQ1 on the
kinetics of oxygen consumption during the oxidation of 20 mM ML in 50 mM
micellar solution of 50 mM Triton X-100 in 50mM phosphate buffer, pH 7.40,
37 °C, initiated by
3 mM AAPH. Arrow shows the moment when SkQ1 was added.
The reduced forms of mitochondria-targeted quinones studied in this work are p-hydroquinones. Acting as chain-breaking antioxidants during the chain peroxidation of styrene p-hydroquinones, "tailless" analogs of mitochondria-targeted antioxidants show a very high inhibiting activity [11], sometimes comparable with that of α-tocopherol (k1 = 3.3 × 106M-1s-1 [26]). For instance, k1 for Me3BQH2 was found to be as much as 2.2 × 106M-1s-1 (Table 1). The behavior of p-hydroquinones in such a system does not differ from that of monophenolic antioxidants [26,27]. The situation dramatically changes when going to the peroxidation of ML in aqueous micelles [12,28]. The matter is that p-hydroxy-substituted phenoxyl radicals QH• formed in reaction [1] having, as a rule, pK less than 5 [29] undergo fast deprotonation at neutral pH: QH•¾→ Q•- + H+ [5] with the formation of semiquinone anion, Q•-, which reacts readily with molecular oxygen, forming O2•- [30,31]: Q•- + O2¾→ Q + O2•- [6] In turn, O2•- may react with oxidation substrate and QH2, most likely in its protonated form, HO2•. Both reactions result in a decrease in the inhibitory activity of QH2 [28]. SOD removes O2•-and thus arrests the mentioned undesirable reactions with the participation of O2•- (HO2•). This was a reason why SOD was always added to our system.
Table 1.
Kinetic parameters characterizing the antioxidant activity of the reduced forms of mitochondria-targeted quinones and their analogs in micellar solution of 50 mM Triton X-100, 50 mM phosphate buffer, pH 7.4, at 37 °C. Oxidation of ML or cardiolipin was initiated by AAPH. Notes: nd - not determined; a structures of QH2 are given in Figure 1; b figures in brackets are the number of independent experiments; c ML is replaced by cardiolipin; d determined during styrene oxidation in the bulk.
| QH2a | k1/k2b | k1 × 105, M-1s-1 |
| SkQ1H2 | 3670 ± 280 (7) 1980 ± 170 (3)c | 2.2 ± 0.2 nd |
| SkQ3H2 | 2720 ± 210 (4) | 1.6 ± 0.1 |
| SkQ5H2 | 2670 ± 180 (5) | 1.6 ± 0.1 |
| MitoQH2 | 970 ± 55 (6) 520 ± 37 (3)c | 0.58 ± 0.03 nd |
| DMQH2 | 1260 ± 85 (4) | 0.76 ± 0.5 |
| Me3BQH2 | 2170 ± 130 (4) | 1.3 ± 0.1 23d |
| Me4BQH2 | 5020 ± 380 (3) | 3.0 ± 0.2 |
| Ubiquinol-0 | 700 ± 45 (3) | 0.42 ± 0.03 4.4d |
| α-tocopherol | 1170 ± 70 (4) | 0.70 ± 0.04 |
| HPMC | 8680 ± 700 (4) | 5.2 ± 0.4 |
The [O2] traces recorded during the induction period of the inhibited oxidation of ML were used to determine k1. On the base of a reductive kinetic scheme, which includes reactions (0), [1], [2], and [4], the following equation can be deduced [11,12] where [LH] is the concentration of the oxidation substrate (in our case ML). Figure 3B depicts the original [O2] trace (Figure 3A) in the axes of Eq. [7]. It is seen that the plot of F vs. time is a straight line as predicted by Eq. [7]. The kinetic behavior of all the other QH2 studied proved to be was similar. The value of k1/k2 can be calculated from the slope of this straight line by using Eq. 7. It should be noted that this way of calculation of k1/k2 does not require the knowledge in RIN and the starting concentration of QH2. The values of k1/k2 are listed in Table 1. The absolute values of k1were calculated from k1/k2 assuming k2 = 60 M-1s-1 [22].
The k1 values are also listed in Table 1.
With two QH2, SkQ1H2 and MitoQH2, similar experiments were conducted by using the same testing system, but with substituting ML by cardiolipin, the most oxidizable phospholipid component in mitochondria membranes [32,33]. As seen from Figure 5, both [O2] traces during the induction period of the inhibited oxidation and the plots of F vs. time are very similar to those for ML. The value of k1/k2 was calculated from the slope of the plot B (Figure 5) by using Eq. [7] assuming that each molecule of cardiolipin contains four fatty acid residue with 87 % linoleate in the cardiolipin sample used in this work (see http://www.avantilipids.com). These data are also presented in Table 1. Unfortunately, the absolute values of k1 could not be calculated, as k2 for the oxidation of cardiolipin has never been reported.
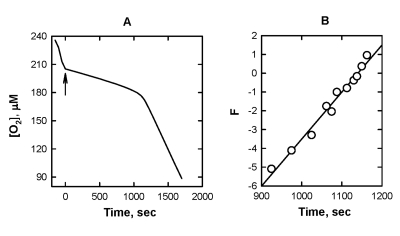
Figure 5. The effect of
addition of 10 μM SkQ1H2
on the kinetics of oxygen consumption during the oxidation of 2.6 mg mL-1 cardiolipin 50
mM micellar solution of 50 mM Triton X-100 in 50mM phosphate buffer, pH
7.40, 37 °C, initiated by
3 mM AAPH. Arrow shows the moment when SkQ1H2 was added.
Discussion
In this paper, the reactivity of the reduced forms of the mitochondria-targeted quinones as chain-breaking anti-oxidants has systematically been studied. As may be seen from Table 1, the k1 value for SkQ1H2, SkQ3H2 and SkQ5H2 are significantly higher than that for MitoQH2. This is in line with the data for simple "tailless" analogs of SkQ1H2 and MitoQH2, namely Me3BQH2, Me4BQH2 and Ubiquinol-0. The same tendency was earlier observed when effects of "tailless" analogues on the chain oxidation of styrene in bulk [11] and ML peroxidation in SDS micelles were studied [12]. Possible reasons why methyl-substituted p-hydroquinones are better antioxidants than methoxy-substituted p-hydroquinones were described elsewhere [11,26]. In brief, the effect under consideration is, the most probably, stereoelectronic by its nature. The matter is that o-methoxy group forms H-bond with oxygen belonging to the adjacent OH group. This causes the decrease in overlap between p-type orbital of oxygen atom of OH-group and the aromatic π-electron cloud (the increase of the dihedral angle between the aromatic ring and O - H bond). The latter results in strengthening O - H bond as compared with that in o-methyl substituted QH2, where such an intramolecular H-bond is absent.
Among mitochondria-targeted QH2 studied in this work, SkQ1H2 showed the highest reactivity towards the lipid peroxyl radicals (Table 1). This observation is in line with data obtained in our group by using several biological models [2,10,14]. However, we recognize that the highest value of k1 for SkQ1H2 is likely not the only reason for the outstanding biological activity of SkQ1. It should be taken into account that k1 given in Table 1 are effective values and cannot be directly attributed to the elementary reaction [1]. The genuine values of k1 can be determined during the chain oxidation in non-polar media, for instance in styrene [11,34,35]. When going to the oxidation of fatty acid (ester) in bulk [12,36]and further to the oxidation in aqueous micelles and liposomes [12,26,37], the experimentally determined k1 values significantly decrease, nearly by one order of magnitude (see data for ubiquinol-0, Table 1). A reason for such a reduction of k1 was repeatedly discussed. The mentioned decrease in k1 is not specific of QH2. A similar effect has earlier been also reported for the oxidation inhibited by monophenolics [25,26,37,38]. The formation of H-bonds between the OH-group of phenolics and the carboxy-group of ML has been suggested as the main reason for the k1 decrease when going from the oxidation of non-polar hydrocarbon to that of fatty acid (ester) [36]. Recently, hydrogen bonding between phenols and fatty acid esters was directly observed by using the NMR technique [39]. Most likely, this is also true for QH2 studied in this work. The further decrease in k1 when going from ML oxidation in bulk to that in aqueous micelles may be explained by the additional formation of H-bonds between QH2 and water molecules as this was earlier suggested for monophenolics [23,37,38].
A general specific feature of reduced forms of the studied mitochondria-targeted quinoles is that their reactivity is actually very close to that of their "tailless" analogs (Table 1). This is in contrast to the couple "α-tocopherol having the long aliphatic chain its "tailless" analog HPMC. The k1 value for α-tocopherol is nearly one order of magnitude lower than that for HPMC (Table 1). This effect was reported to be even more pronounced in the SDS micelles [23,37,38]. The essential feature of our testing system and related microheterogeneous systems is that the concentration of the antioxidants tested is much lower than that of the oxidation substrate (in our case ML). While every micelle (microreactor) contains several molecules of ML, only a few micelles contain an antioxidant. Under these conditions, a fast LO2• reduction by an antioxidant is possible only if an antioxidant is capable of fast transferring from one microreactor to another, the characteristic time of this transfer being shorter than the time of the occurrence of a single kinetic chain. The antioxidants with a rather long aliphatic residue like α-tocopherol commonly do not meet such a requirement [37]. The fact that the values of k1 for the mitochondria-targeted quinols actually do not differ from that of their "tailless" analogs (Table 1) means that all of them are capable of the fast transfer from one microreactor to another. This is in line with a high reported ability of SkQ and MitoQ to easily penetrate through biological membranes [14].
Materials and Methods
Methyl linoleate and Triton X-100 were purchased from Sigma, heart bovine cardiolipin disodium salt was received from Avanti PolarLipids. The water-soluble initiator 2,2'-azobis(2-amidinopropan) dihydrochloride (AAPH) was obtained from Polysciences. NaH2PO4 and Na2HPO4 of the highest quality used to prepare buffer solutions were purchased from Merck. The mitochondria-targeted quinones, SkQ1, SkQ3, SkQ5, MitoQ, DMQ as well as C12TPP (see Figure 1) were synthesized in the Mitoengineering Centre of Moscow State University [2]. Trimethylhydroquinone (Me3BQH2) was purchased from Aldrich; 2,3-dimethoxy-5-methyl-benzoqyuinone (ubiquinone-0) was from Sigma; tetramethylbenzoquinone (Me4BQ) was from EGA Chemie. All the other chemicals were of highest available quality.
The reduced forms of the mitochondria-targeted quinones (QH2) were produced by the reduction of corresponding quinones by NaBH4 in the mixture of 50 mM NaH2PO4 (pH 5.0) with ethanol. This process was under control of UPLC-MS-MS (see below). Reduced forms of ubiquinone-0 and tetramethylhydroquinone (Me4BQH2) were produced by reduction of the quinones by Zn powder [21]. The buffer solution (pH 7.40 ± 0.02) was prepared by mixing 50 mM solutions of NaH2PO4 and Na2HPO4. In turn, the solutions of the individual sodium phosphates were prepared with doubly distilled water and were purged from traces of transition metals by Chelex-100 resin (Bio-Rad).
HPLC-diode array detection-electrospray ionization tandem mass spectrometry analysis (UPLC-MS-MS) was performed using an ACQUITY system (Waters, Milford, MA, USA). Chromatography was carried out using an ACQUITY BEH C18 column (2.1 x 50 mm, 1.7 μm) eluted with a gradient of 40-60% acetonitrile (4 min) and 20 mM acetic acid (pH 3.0) delivered at a flow rate of 0.5 mL per min. UV-monitoring was performed at 280 mm. An injection volume of 11.2 μL (full loop) was used in all cases. A Quattro triple-quadrupole mass spectrometer (Micromass-Waters) fitted with a Z-Spray ion interface was used for analyses. Ionization was achieved using electrospray in a positive ionization mode. The following conditions were found to be optimal for the analysis of SkQ1: capillary voltage, 3.0 kV; source block temperature, 120°C; and desolvatation gas (nitrogen) heated to 450°C and delivered at a flow rate of 800 L h-1; cone voltage, 55 V; cone Gas Flow rate, 50 L h-1. MassLynx 4.0 software (Waters) was used for processing.
The standard testing system was composed of 50 mM buffer, pH 7.4, 50 mM Triton X-100, 2-4 mM AAPH, 8-20 mM ML and 20 unit mL-1 SOD. In some experiments, ML was replaced by cardiolipin. The kinetics of oxygen consumption accompanied ML (cardiolipin) oxidation were studied with a computerized 5300 Biological Oxygen Monitor (Yellow Springs Instruments Co., USA) with a Clark electrode as a sensor. The rate of oxidation was measured as a slope of [O2] traces. Experiments were conducted at 37.0 ± 0.1 °C. ML was added to preliminarily thermostated micellar solution of Triton X-100 and AAPH in buffer. Monitoring was started 3-5 min after ML addition and the rate of non-inhibited oxidation (R0) was measured. The tested compounds were then added to a reaction chamber under steady monitoring as a stock solution by using a Hamilton micro-syringe. In more detail, the protocol was described elsewhere [12,21,22].
Acknowledgments
Supported by Mitotechnology LLC, Russia Ministry of Education and Science (grant "Leading Scientific Schools" N 5762.2008.4).
Conflicts of Interest
The authors in this manuscript have no conflict of interest to declare.
References
- 1. Murphy MP and Smith RAJ. Targeting antioxidants to mitochondria by conjugation to lipophilic cations. Annu Rev Pharmacol Toxicol. 2007; 47: 629 -656. [PubMed] .
- 2. Skulachev VP A Biochemical Approach to the Problem of Aging: "Megaproject" on Membrane-Penetrating Ions. The First Results and Prospects. Biochemistry (Moscow). 2007; 72: 1385 -1396. [PubMed] .
- 3. Hoye AT , Davoren JE , Wipe P , Fink MP and Kagan VE. Targeting Mitochondria. Acc Chem Res. 2008; 41: 87 -97. [PubMed] .
- 4. Rocha M and Victor VM. Targeting antioxidants to mitochondria and cardiovascular diseases: The effects of mitoquinone. Med Sci Monit. 2007; 13: RA132 -145. [PubMed] .
- 5. Liberman EA , Topaly VP , Tsofina LM , Jasaitis AA and Skulachev VP. Mechanism of coupling of oxidative phosphorylation and the membrane potential of mitochondria. Nature. 1969; 222: 1076 -1078. [PubMed] .
- 6. Severin SE , Skulachev VP and Yaguzhinsky LS. A possible role of carnitine in Transport of fatty acids through the mitochondrial membrane. Biokhimiya. 1970; 35: 1250 -1257 (Russ). [PubMed] .
- 7. Murphy M P Targeting bioactive compounds to mitochondria. Trends Biotechnol. 1997; 15: 326 -330. [PubMed] .
- 8. Murphy MP and Smith RAJ. Drug delivery to mitochondria: the key to mitochondrial medicine. Adv Drug Deliv Rev. 2000; 41: 235 -250. [PubMed] .
- 9. Smith RAJ , Kelso GF , James AM and Murphy M P. Targeting coenzyme Q derivatives to mitochondria. Meth Enzymol. 2004; 382: 45 -67. [PubMed] .
- 10. Skulachev VP. Method of acting upon organism by targeted delivery of biologically active substances into mitochondria, pharmaceutical composition for carrying out said method, and compound used for the purpose. World patent. .
- 11. Loshadkin D , Roginsky V and Pliss E. Substituted p-hydroquinones as a chain-breaking antioxidant during the oxidation of styrene. Int J Chem Kinetics. 2002; 34: 162 -171. .
- 12. Roginsky V , Barsukova T , Loshadkin D and Pliss E. Substituted para-hydroquinones as an inhibitor of lipid peroxidation. Chem Phys Lipids. 2003; 125: 49 -58. [PubMed] .
- 13. Kruk J , Jemiola-Rzeminska M and Strzalka K. Plastoquinol and α-tocopherol quinol are more active than ubiquinol and α-tocopherol in inhibition of lipid peroxidation. Chem Phys Lipids. 1997; 87: 73 -80. .
- 14. Antonenko YN , Avetisyan AV , Bakeeva LE , Chernyak BV , Chertkov VA , Domnina LV , Ivanova OY , Izyumov DS , Khailova LS , Klishin SS , Korshunova GA and Lyamzaev KG. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 1. Cationic plastoquinone derivatives: synthesis and in vitro studies. Biochemistry (Moscow). 2008; 73: 1273 -1287. [PubMed] .
- 15. Bakeeva LE , Barskov IV , Egorov MV , Isaev NK , Kapelko VI , Kazachenko AV , Kirpatovsky VI , Kozlovsky SV , Lakomkin VL , Levina SB , Pisarenko OI and Plotnikov EY. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 2. Treatment of some ROS- and age-related diseases (heart arrhythmia, heart infarctions, kidney ischemia, and stroke). Biochemistry (Moscow). 2008; 73: 1288 -1299. [PubMed] .
- 16. Agapova LS , Chernyak BV , Domnina LV , Dugina VB , Efimenko AY , Fetisova EK , Ivanova OY , Kalinina NI , Khromova NV , Kopnin BP , Kopnin PB and Korotetskaya MV. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 3. Inhibitory effect of SkQ1 on tumor development from p53-deficient cells. Biochemistry (Moscow). 2008; 73: 1300 -1316. [PubMed] .
- 17. Neroev VV , Archipova MM , Bakeeva LE , Fursova AZh , Grigorian EN , Grishanova AY , Iomdina EN , Ivashchenko ZhN , Katargina LA , Khoroshilova-Maslova IP , Kilina OV and Kolosova NG. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 4. Age-related eye disease. SkQ1 returns vision to blind animals. Biochemistry (Moscow). 2008; 73: 1317 -1328. [PubMed] .
- 18. Anisimov VN , Bakeeva LE , Egormin PA , Filenko OF , Isakova EF , Manskikh VN , Mikhelson VM , Panteleeva AA , Pasyukova EG , Pilipenko DI , Piskunova TS and Popovich IG. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 5. SkQ1 prolongs lifespan and prevents development of traits of senescence. Biochemistry (Moscow). 2008; 73: 1329 -1342. [PubMed] .
- 19. Skulachev VP , Anisimov VN , Antonenko YN , Bakeeva LE , Chernyak BV , Erichev VP , Filenko OF , Kalinina NI , Kapelko VI , Kolosova NG , Kopnin BP and Korshunova GA. An attempt to prevent senescence: A mitochondrial approach. Biochim Biophys Acta. 2009; 1797: 437 -461. [PubMed] .
- 20. Plotnikov EY , Vasileva AK , Arkhangelskaya AA , Pevzner IB , Skulachev VP and Zorov DB. Interrelations of mitochondrial fragmentation and cell death under ischemia/reoxygenation and UV-irradiation: protective effects of SkQ1, lithium ions and insulin. FEBS Lett. 2008; 582: 3117 -3124. [PubMed] .
- 21. Roginsky VA and Barsukova TK. Kinetics of oxidation of hydroquinones by molecular oxygen. Effect of superoxide dismutase. J Chem Soc Perkin 2 Trans. 2000; N7: 1575 -1582. .
- 22. Roginsky VA Chain-breaking antioxidant activity of natural polyphenols as determined during the chain oxidation of methyl linoleate in Triton X-100 micelles. Arch Biochem Biophys. 2003; 414: 261 -270. [PubMed] .
- 23. Castle L and Perkins MJ. Inhibition kinetics of chain-breaking phenolic antioxidants in SDS micelles. Evidence that intermicellar diffusion rates may be rate-limiting for hydrophobic inhibitors such as alpha-tocopherol. J Amer Chem Soc. 1986; 108: 6381 -6382. .
- 24. Barclay LRC , Locke SJ , MacNeil JM and VanKessel J. Quantitative studies of linoleate monomers sequestered in phosphatidylcholine bilayers. Absolute rate constant in bilayers. Canad J Chem. 1985; 63: 2633 -2638. .
- 25. Barclay LRC , Locke SJ and MacNeil JM. Autoxidation in micelles. Synergism of vitamin C with lipid-soluble vitamin E and water-soluble Trolox. Canad J Chem. 1985; 63: 366 -374. .
- 26. Barclay LRC and Vinqvist MR. Rappoport Z. Phenols as antioxidants The Chemistry of Phenols. Willey 2003; 840 -907. .
- 27. Roginsky VA Phenolic Antioxidants: Efficiency and Reactivity. 1988; Moscow Nauka Russian. .
- 28. Roginsky VA Superoxide dismutase enhances chain-breaking antioxidant capability of hydroquinones. Free Radic Res. 2001; 35: 55 -62. [PubMed] .
- 29. Landolt-Börnstein: Numerical Data and Functional Relationships in Science and Technology - New Series. Group II, V. 13e. 1984; Berlin, Heidelberg Springer-Verlag .
- 30. Wardman P Bioreductive activation of quinones: Redox properties and thiol reactivity. Free Radic Res Commun. 1990; 8: 219 -229. [PubMed] .
- 31. O'Brien PJ Molecular mechanisms of quinone cytotoxicity. Chem-Biol Interact. 1991; 80: 1 -41. [PubMed] .
- 32. Lesnefsky EJ and Hoppel CL. Cardiolipin as an oxidative target in cardiac mitochondria in the aged rat. Biochim Biophys Acta. 2008; 1777: 1020 -1027. [PubMed] .
- 33. Ott M , Gogvadze V , Orrenius S and Zhivotovsky B. Mitochondria, oxidative stress and cell death. Apoptosis. 2007; 12: 913 -922. [PubMed] .
- 34. Litwinienko G and Ingold KU. Solvent effects on the rates and mechanisms of reaction of phenols with free radicals. Acc Chem Res. 2007; 40: 222 -230. [PubMed] .
- 35. Tikhonov I , Roginsky V and Pliss E. The chain-breaking antioxidant activity of phenolic compounds with different number of O-H groups as determined during the oxidation of styrene. Int J Chem Kinet. 2008; In press .
- 36. Roginsky VA Kinetics of oxidation of polyunsaturated fatty acid esters inhibited by substituted phenols. Kinetics and Catalysis (Moscow). 1990; 31: 475 -481. .
- 37. Roginsky VA The inhibiting ability of lipid-soluble and water-soluble phenols at lipid peroxidation in micro-heterogeneous systems. Biol Membr (Moscow). 1990; 4: 437 -451. .
- 38. Prayor WA , Strickland T and Church DF. Comparison of the efficiencies of several natural and synthetic antioxidants in aqueous sodium dodecyl sulfate micelle solutions. J Amer Chem Soc. 1989; 110: 2224 -2229. .
- 39. Litwinienko G , Megiel E and Wojnicz M. Hydrogen bonding between phenols and fatty acid esters: 1H NMR study and ab initio calculations. Org Lett. 2002; 4: 2425 -2428. [PubMed] .